Garcinol A Novel Inhibitor of Platelet Activation and Apoptosis

 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
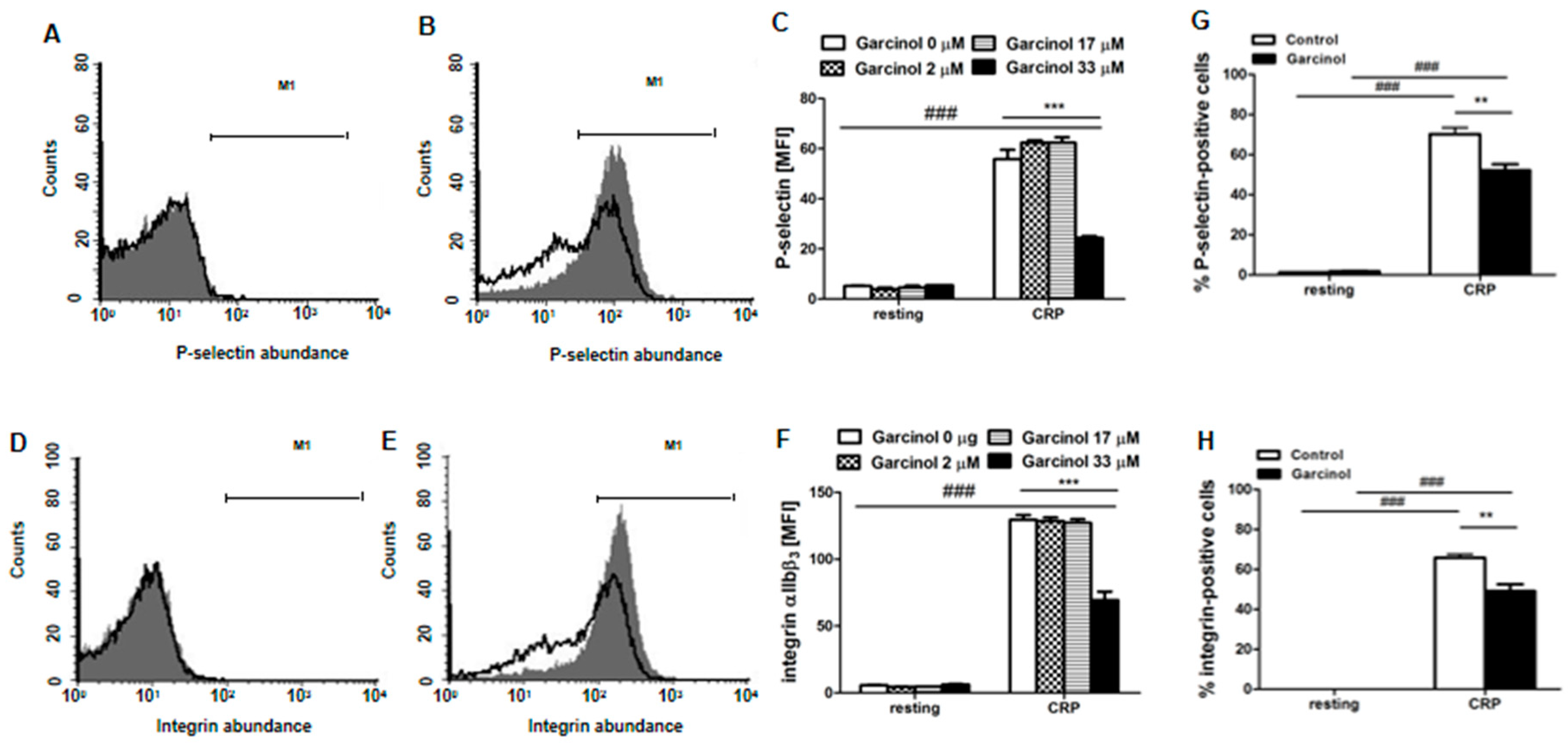
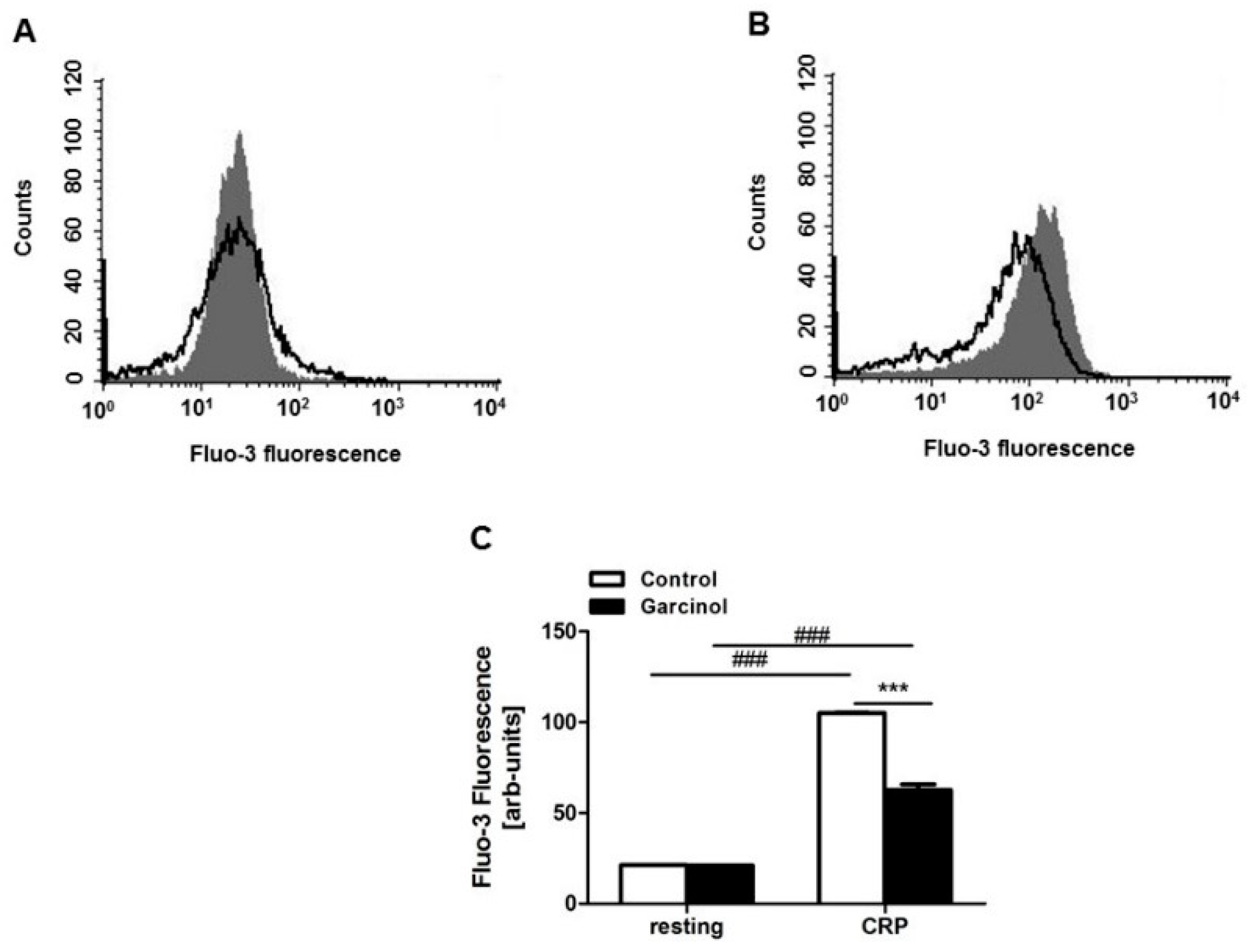
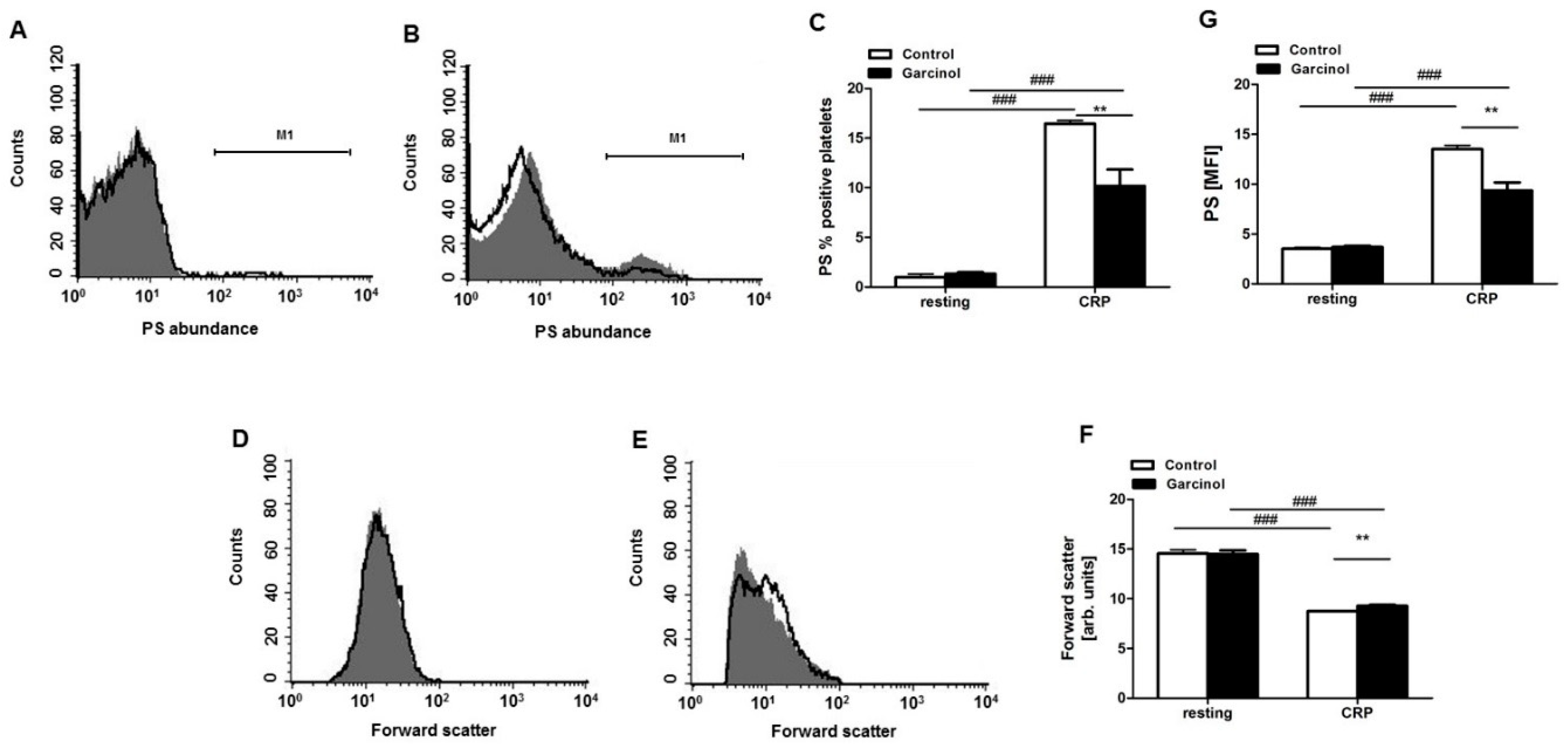
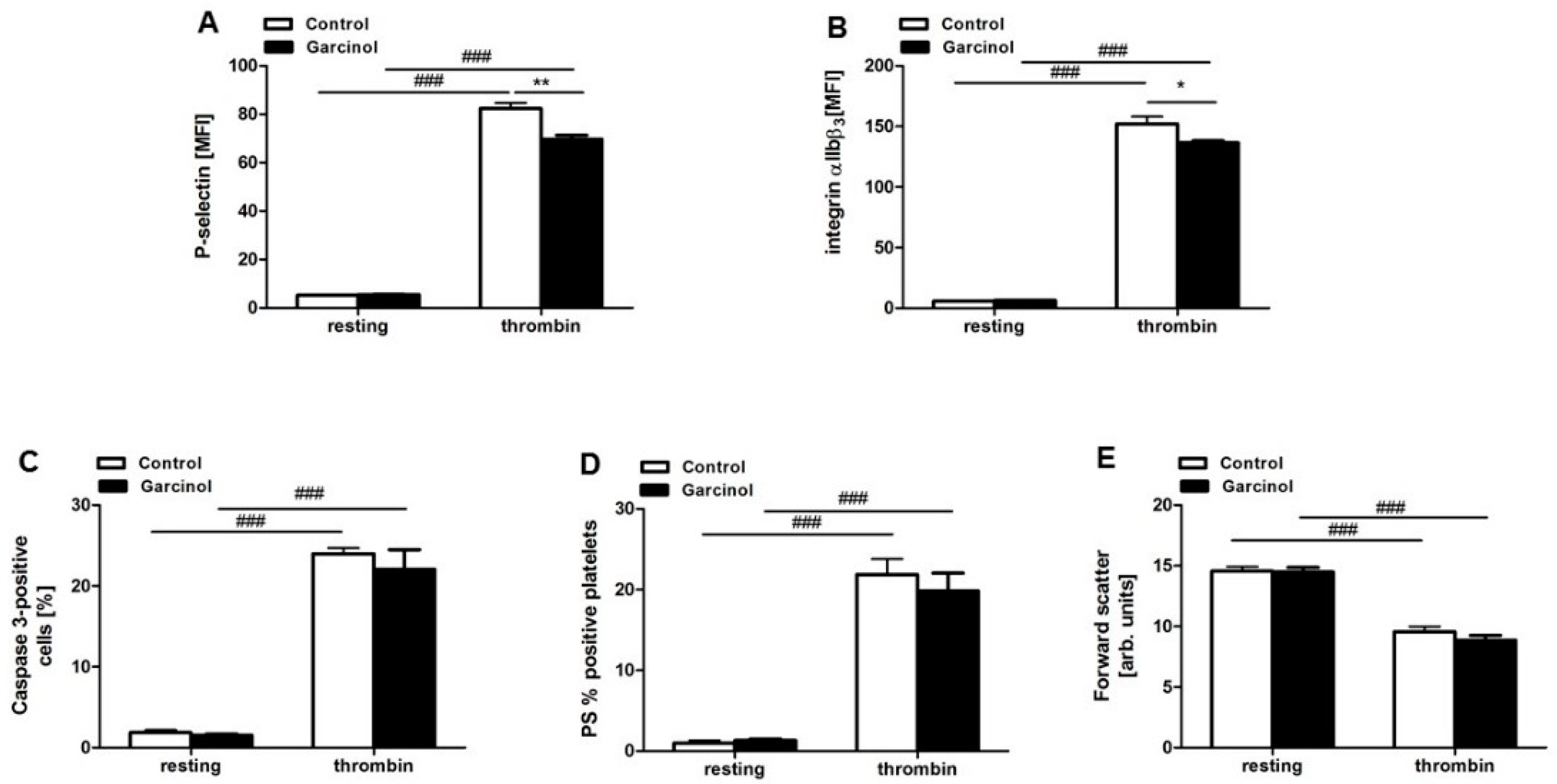
2. Results
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Mice
5.2. Preparation of Mouse Platelets
5.3. Cytosolic Calcium
5.4. P-Selectin and Activated Integrin Abundance
5.5. Phosphatidylserine Exposure and Forward Scatter
5.6. Caspase-3 Activity
5.7. Platelet Aggregation
5.8. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Behera, A.K.; Swamy, M.M.; Natesh, N.; Kundu, T.K. Garcinol and its role in chronic diseases. Adv. Exp. Med. Biol. 2016, 928, 435–452. [Google Scholar] [PubMed]
- Kolodziejczyk, J.; Masullo, M.; Olas, B.; Piacente, S.; Wachowicz, B. Effects of garcinol and guttiferone k isolated from garcinia cambogia on oxidative/nitrative modifications in blood platelets and plasma. Platelets 2009, 20, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Ho, P.C.; Wong, F.C.; Sethi, G.; Wang, L.Z.; Goh, B.C. Garcinol: Current status of its anti-oxidative, anti-inflammatory and anti-cancer effects. Cancer Lett. 2015, 362, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Padhye, S.; Ahmad, A.; Oswal, N.; Sarkar, F.H. Emerging role of garcinol, the antioxidant chalcone from garcinia indica choisy and its synthetic analogs. J. Hematol. Oncol. 2009, 2, 38. [Google Scholar] [CrossRef] [PubMed]
- Hao, F.; Jia, L.H.; Li, X.W.; Zhang, Y.R.; Liu, X.W. Garcinol upregulates gabaa and gad65 expression, modulates bdnf-trkb pathway to reduce seizures in pentylenetetrazole (ptz)-induced epilepsy. Med. Sci. Monit. 2016, 22, 4415–4425. [Google Scholar] [CrossRef] [PubMed]
- Jing, Y.; Ai, Q.; Lin, L.; Dai, J.; Jia, M.; Zhou, D.; Che, Q.; Wan, J.; Jiang, R.; Zhang, L. Protective effects of garcinol in mice with lipopolysaccharide/d-galactosamine-induced apoptotic liver injury. Int. Immunopharmacol. 2014, 19, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, S.; Das, S.N. Garcinol inhibits tumour cell proliferation, angiogenesis, cell cycle progression and induces apoptosis via nf-kappab inhibition in oral cancer. Tumour Biol. 2016, 37, 7175–7184. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Wang, Z.; Wojewoda, C.; Ali, R.; Kong, D.; Maitah, M.Y.; Banerjee, S.; Bao, B.; Padhye, S.; Sarkar, F.H. Garcinol-induced apoptosis in prostate and pancreatic cancer cells is mediated by nf-kappab signaling. Front. Biosci. 2011, 3, 1483–1492. [Google Scholar] [CrossRef]
- Cheng, A.C.; Tsai, M.L.; Liu, C.M.; Lee, M.F.; Nagabhushanam, K.; Ho, C.T.; Pan, M.H. Garcinol inhibits cell growth in hepatocellular carcinoma hep3b cells through induction of ros-dependent apoptosis. Food Funct. 2010, 1, 301–307. [Google Scholar] [CrossRef]
- Parasramka, M.A.; Gupta, S.V. Synergistic effect of garcinol and curcumin on antiproliferative and apoptotic activity in pancreatic cancer cells. J. Oncol. 2012, 2012, 709–739. [Google Scholar] [CrossRef]
- Liao, C.H.; Sang, S.; Ho, C.T.; Lin, J.K. Garcinol modulates tyrosine phosphorylation of fak and subsequently induces apoptosis through down-regulation of src, erk, and akt survival signaling in human colon cancer cells. J. Cell. Biochem. 2005, 96, 155–169. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Wang, Z.; Ali, R.; Maitah, M.Y.; Kong, D.; Banerjee, S.; Padhye, S.; Sarkar, F.H. Apoptosis-inducing effect of garcinol is mediated by nf-kappab signaling in breast cancer cells. J. Cell. Biochem. 2010, 109, 1134–1141. [Google Scholar] [PubMed]
- Sethi, G.; Chatterjee, S.; Rajendran, P.; Li, F.; Shanmugam, M.K.; Wong, K.F.; Kumar, A.P.; Senapati, P.; Behera, A.K.; Hui, K.M.; et al. Inhibition of stat3 dimerization and acetylation by garcinol suppresses the growth of human hepatocellular carcinoma in vitro and in vivo. Mol. Cancer 2014, 13, 66. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Kwon, S.J.; Sang, S.; Ju, J.; Zhou, J.N.; Ho, C.T.; Huang, M.T.; Yang, C.S. Effects of garcinol and its derivatives on intestinal cell growth: Inhibitory effects and autoxidation-dependent growth-stimulatory effects. Free Radic. Biol. Med. 2007, 42, 1211–1221. [Google Scholar] [CrossRef] [PubMed]
- Koeberle, A.; Northoff, H.; Werz, O. Identification of 5-lipoxygenase and microsomal prostaglandin e2 synthase-1 as functional targets of the anti-inflammatory and anti-carcinogenic garcinol. Biochem. Pharmacol. 2009, 77, 1513–1521. [Google Scholar] [CrossRef] [PubMed]
- Fazio, A.; Briglia, M.; Faggio, C.; Alzoubi, K.; Lang, F. Stimulation of suicidal erythrocyte death by garcinol. Cell. Physiol. Biochem. 2015, 37, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Guidetti, G.F.; Canobbio, I.; Torti, M. Pi3k/akt in platelet integrin signaling and implications in thrombosis. Adv. Biol. Regul. 2015, 59, 36–52. [Google Scholar] [CrossRef]
- Lang, F.; Munzer, P.; Gawaz, M.; Borst, O. Regulation of stim1/orai1-dependent ca2+ signalling in platelets. Thromb. Haemost. 2013, 110, 925–930. [Google Scholar]
- Lannan, K.L.; Sahler, J.; Kim, N.; Spinelli, S.L.; Maggirwar, S.B.; Garraud, O.; Cognasse, F.; Blumberg, N.; Phipps, R.P. Breaking the mold: Transcription factors in the anucleate platelet and platelet-derived microparticles. Front. Immunol. 2015, 6, 48. [Google Scholar] [CrossRef]
- Muller-Newen, G.; Stope, M.B.; Kraus, T.; Ziegler, P. Development of platelets during steady state and inflammation. J. Leukoc. Biol. 2017, 101, 1109–1117. [Google Scholar] [CrossRef]
- Kile, B.T. The role of apoptosis in megakaryocytes and platelets. Br. J. Haematol. 2014, 165, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Borst, O.; Schmidt, E.M.; Munzer, P.; Schonberger, T.; Towhid, S.T.; Elvers, M.; Leibrock, C.; Schmid, E.; Eylenstein, A.; Kuhl, D.; et al. The serum-and glucocorticoid-inducible kinase 1 (sgk1) influences platelet calcium signaling and function by regulation of orai1 expression in megakaryocytes. Blood 2012, 119, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Gawaz, M. Role of platelets in coronary thrombosis and reperfusion of ischemic myocardium. Cardiovasc. Res. 2004, 61, 498–511. [Google Scholar] [CrossRef] [PubMed]
- Borst, O.; Munzer, P.; Gatidis, S.; Schmidt, E.M.; Schonberger, T.; Schmid, E.; Towhid, S.T.; Stellos, K.; Seizer, P.; May, A.E.; et al. The inflammatory chemokine cxc motif ligand 16 triggers platelet activation and adhesion via cxc motif receptor 6-dependent phosphatidylinositide 3-kinase/akt signaling. Circ. Res. 2012, 111, 1297–1307. [Google Scholar] [CrossRef] [PubMed]
- Bergmeier, W.; Stefanini, L. Novel molecules in calcium signaling in platelets. J. Thromb. Haemost. 2009, 7, 187–190. [Google Scholar] [CrossRef]
- Varga-Szabo, D.; Braun, A.; Nieswandt, B. Calcium signaling in platelets. J. Thromb. Haemost. 2009, 7, 1057–1066. [Google Scholar] [CrossRef] [PubMed]
- Braun, A.; Varga-Szabo, D.; Kleinschnitz, C.; Pleines, I.; Bender, M.; Austinat, M.; Bosl, M.; Stoll, G.; Nieswandt, B. Orai1 (cracm1) is the platelet soc channel and essential for pathological thrombus formation. Blood 2009, 113, 2056–2063. [Google Scholar] [CrossRef]
- Varga-Szabo, D.; Braun, A.; Nieswandt, B. Stim and orai in platelet function. Cell. Calcium. 2011, 50, 270–278. [Google Scholar] [CrossRef]
- Wang, Y.; Deng, X.; Gill, D.L. Calcium signaling by stim and orai: Intimate coupling details revealed. Sci. Signal. 2010, 3, 42. [Google Scholar] [CrossRef]
- Gyulkhandanyan, A.V.; Mutlu, A.; Freedman, J.; Leytin, V. Markers of platelet apoptosis: Methodology and applications. J. Thromb. Thrombolysis 2012, 33, 397–411. [Google Scholar] [CrossRef]
- Lebois, M.; Josefsson, E.C. Regulation of platelet lifespan by apoptosis. Platelets 2016, 27, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Mammadova-Bach, E.; Nagy, M.; Heemskerk, J.W.M.; Nieswandt, B.; Braun, A. Store-operated calcium entry in thrombosis and thrombo-inflammation. Cell Calcium. 2019, 77, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Harper, M.T.; Poole, A.W. Store-operated calcium entry and non-capacitative calcium entry have distinct roles in thrombin-induced calcium signalling in human platelets. Cell Calcium 2011, 50, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Mahaut-Smith, M.P. A role for platelet trpc channels in the ca2+ response that induces procoagulant activity. Sci. Signal. 2013, 6, 23. [Google Scholar] [CrossRef] [PubMed]
- Mushtaq, M.; Nam, T.S.; Kim, U.H. Critical role for cd38-mediated ca2+ signaling in thrombin-induced procoagulant activity of mouse platelets and hemostasis. J. Biol. Chem. 2011, 286, 12952–12958. [Google Scholar] [CrossRef] [PubMed]
- Lhermusier, T.; Chap, H.; Payrastre, B. Platelet membrane phospholipid asymmetry: From the characterization of a scramblase activity to the identification of an essential protein mutated in scott syndrome. J. Thromb. Haemost. 2011, 9, 1883–1891. [Google Scholar] [CrossRef]
- Badlou, B.A.; Wu, Y.P.; Smid, W.M.; Akkerman, J.W. Platelet binding and phagocytosis by macrophages. Transfusion 2006, 46, 1432–1443. [Google Scholar] [CrossRef]
- Smyth, S.S.; Woulfe, D.S.; Weitz, J.I.; Gachet, C.; Conley, P.B.; Goodman, S.G.; Roe, M.T.; Kuliopulos, A.; Moliterno, D.J.; French, P.A.; et al. G-protein-coupled receptors as signaling targets for antiplatelet therapy. Arterioscler Thromb. Vasc. Biol. 2009, 29, 449–457. [Google Scholar] [CrossRef]
- Bishton, M.J.; Gardiner, E.E.; Harrison, S.J.; Prince, H.M.; Johnstone, R.W. Histone deacetylase inhibitors reduce glycoprotein vi expression and platelet responses to collagen-related peptide. Thromb. Res. 2013, 131, 514–520. [Google Scholar] [CrossRef]
- Saadat, N.; Akhtar, S.; Goja, A.; Razalli, N.H.; Geamanu, A.; David, D.; Shen, Y.; Gupta, S.V. Dietary garcinol arrests pancreatic cancer in p53 and k-ras conditional mutant mouse model. Nutr. Cancer 2018, 70, 1075–1087. [Google Scholar] [CrossRef]
- Cao, H.; Bhuyan, A.A.M.; Umbach, A.T.; Bissinger, R.; Gawaz, M.; Lang, F. Inhibitory effect of afatinib on platelet activation and apoptosis. Cell. Physiol. Biochem. 2017, 43, 2264–2276. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Bissinger, R.; Umbach, A.T.; Al Mamun Bhuyan, A.; Lang, F.; Gawaz, M. Effects of antimalarial tafenoquine on blood platelet activity and survival. Cell. Physiol. Biochem. 2017, 41, 369–380. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Bissinger, R.; Umbach, A.T.; Gawaz, M.; Lang, F. Temsirolimus sensitive stimulation of platelet activity, apoptosis and aggregation by collagen-related peptide. Cell. Physiol. Biochem. 2017, 42, 1252–1263. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Bissinger, R.; Umbach, A.T.; Gawaz, M.; Lang, F. Effect of bexarotene on platelet activation and apoptosis. Cell. Physiol. Biochem. 2017, 42, 838–847. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Qadri, S.M.; Lang, E.; Pelzl, L.; Umbach, A.T.; Leiss, V.; Birnbaumer, L.; Nurnberg, B.; Pieske, B.; Voelkl, J.; et al. Heterotrimeric g-protein subunit galphai2 contributes to agonist-sensitive apoptosis and degranulation in murine platelets. Physiol. Rep. 2018, 6, e13841. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Umbach, A.T.; Bissinger, R.; Gawaz, M.; Lang, F. Inhibition of collagen-related peptide induced platelet activation and apoptosis by ceritinib. Cell. Physiol. Biochem. 2018, 45, 1707–1716. [Google Scholar] [CrossRef] [PubMed]
- Honisch, S.; Gu, S.; Vom Hagen, J.M.; Alkahtani, S.; Al Kahtane, A.A.; Tsapara, A.; Hermann, A.; Storch, A.; Schols, L.; Lang, F.; et al. Chorein sensitive arrangement of cytoskeletal architecture. Cell. Physiol. Biochem. 2015, 37, 399–408. [Google Scholar] [CrossRef]
- Liu, G.; Liu, G.; Alzoubi, K.; Chatterjee, M.; Walker, B.; Munzer, P.; Luo, D.; Umbach, A.T.; Elvira, B.; Chen, H.; et al. Cd44 sensitivity of platelet activation, membrane scrambling and adhesion under high arterial shear rates. Thromb. Haemost. 2015, 115. [Google Scholar]
- Liu, G.; Cao, H.; Liu, G.; Heinzmann, D.; Chen, H.; Umbach, A.T.; Gawaz, M.; Lang, F. Effect of lysosomotropic polyamineoxidase inhibitor mdl-72527 on platelet activation. Cell. Physiol. Biochem. 2016, 38, 1695–1702. [Google Scholar] [CrossRef]
- Liu, G.; Liu, G.; Chatterjee, M.; Umbach, A.T.; Chen, H.; Gawaz, M.; Lang, F. Influence of gamma-secretase inhibitor 24-diamino-5-phenylthiazole dapt on platelet activation. Cell. Physiol. Biochem. 2016, 38, 726–736. [Google Scholar] [CrossRef]
- Liu, G.; Liu, G.; Chen, H.; Alzoubi, K.; Umbach, A.T.; Gawaz, M.; Stournaras, C.; Lang, F. Rapid upregulation of orai1 abundance in the plasma membrane of platelets following activation with thrombin and collagen-related peptide. Cell. Physiol. Biochem. 2015, 37, 1759–1766. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Lin, J.; Wang, B.; He, S.; Wu, C.; Kushwaha, K.K.; Mohabeer, N.; Su, Y.; Fang, H.; Huang, K.; et al. Inflammatory cytokine tslp stimulates platelet secretion and potentiates platelet aggregation via a tslpr-dependent pi3k/akt signaling pathway. Cell. Physiol. Biochem. 2015, 35, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Liu, G.; Chen, H.; Borst, O.; Gawaz, M.; Vortkamp, A.; Schreiber, R.; Kunzelmann, K.; Lang, F. Involvement of ca2+ activated cl- channel ano6 in platelet activation and apoptosis. Cell. Physiol Biochem 2015, 37, 1934–1944. [Google Scholar] [CrossRef] [PubMed]
- De Cuyper, I.M.; Meinders, M.; van de Vijver, E.; de Korte, D.; Porcelijn, L.; de Haas, M.; Eble, J.A.; Seeger, K.; Rutella, S.; Pagliara, D.; et al. A novel flow cytometry-based platelet aggregation assay. Blood 2013, 121, e70–e80. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, H.; Al Mamun Bhuyan, A.; Umbach, A.T.; Ma, K.; Borst, O.; Gawaz, M.; Zhang, S.; Nürnberg, B.; Lang, F. Garcinol A Novel Inhibitor of Platelet Activation and Apoptosis. Toxins 2019, 11, 382. https://doi.org/10.3390/toxins11070382
Cao H, Al Mamun Bhuyan A, Umbach AT, Ma K, Borst O, Gawaz M, Zhang S, Nürnberg B, Lang F. Garcinol A Novel Inhibitor of Platelet Activation and Apoptosis. Toxins. 2019; 11(7):382. https://doi.org/10.3390/toxins11070382
Chicago/Turabian StyleCao, Hang, Abdulla Al Mamun Bhuyan, Anja T. Umbach, Ke Ma, Oliver Borst, Meinrad Gawaz, Shaqiu Zhang, Bernd Nürnberg, and Florian Lang. 2019. "Garcinol A Novel Inhibitor of Platelet Activation and Apoptosis" Toxins 11, no. 7: 382. https://doi.org/10.3390/toxins11070382
APA StyleCao, H., Al Mamun Bhuyan, A., Umbach, A. T., Ma, K., Borst, O., Gawaz, M., Zhang, S., Nürnberg, B., & Lang, F. (2019). Garcinol A Novel Inhibitor of Platelet Activation and Apoptosis. Toxins, 11(7), 382. https://doi.org/10.3390/toxins11070382