Comparative Analysis of Listeria monocytogenes Plasmids and Expression Levels of Plasmid-Encoded Genes during Growth under Salt and Acid Stress Conditions


Abstract
:1. Introduction
2. Results and Discussion
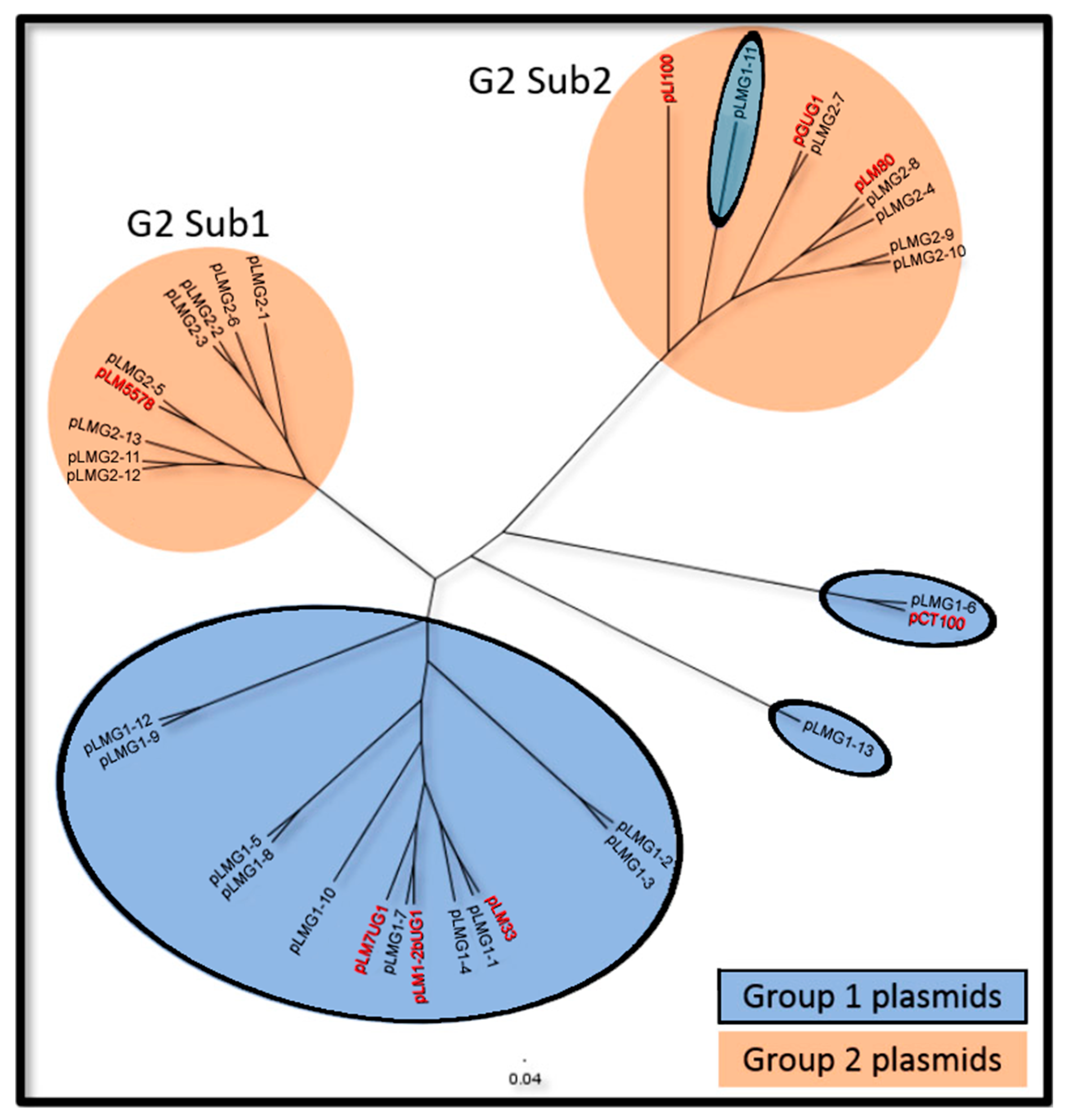
2.1. L. monocytogenes Plasmid Types and Characteristics
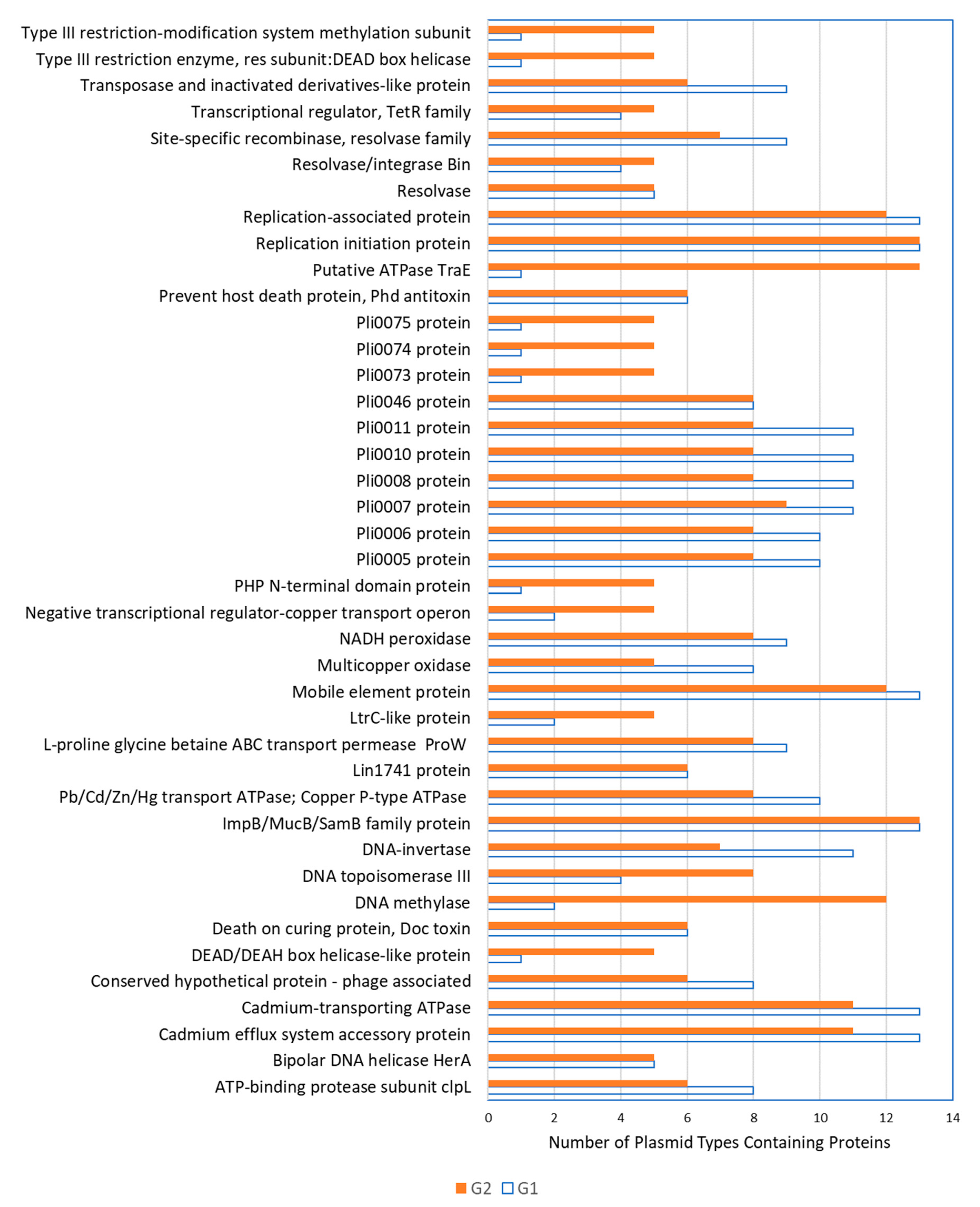
2.2. Genetic Elements Shared by Group 1 and Group 2 Plasmids
2.3. Genetic Elements Associated with Group 1 Plasmids
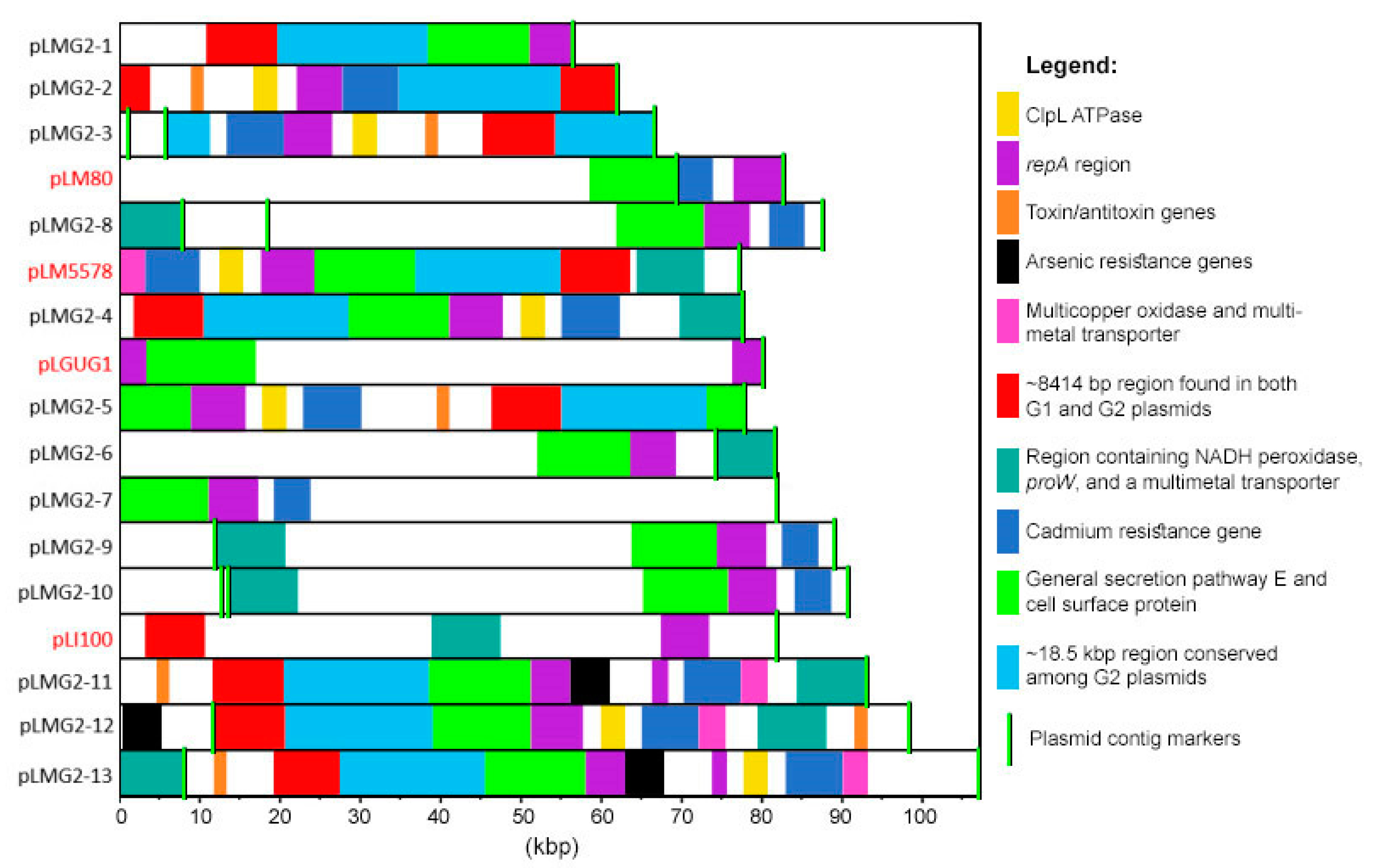
2.4. Genetic Elements Associated with Group 2 Plasmids
2.5. Expression of the L. monocytogenes Plasmid-Encoded Genes During Growth Under Salt and Acid Stress Conditions
2.6. Stress Tolerance Comparisons of Wildtype and Plasmid-Cured Strains
3. Conclusions
4. Materials and Methods
4.1. Strains and Culture Conditions
4.2. Genetic Comparisons of L. Monocytogenes Plasmids
4.3. RNA Isolation and Real-Time qPCR Analysis
4.4. Stress Tolerance Comparisons of Wildtype and Plasmid-Cured Strains
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | Strain | LPD (h) | µmax (A600nm/h) | Nmax (A600nm) |
|---|---|---|---|---|
| BHIB (25 °C) Control | Lm10 | 7.38 ± 0.08 a | 0.09 ± 0.00 a | 0.89 ± 0.01 a |
| Lm10_PC | 8.46 ± 0.05b | 0.09 ± 0.00 a | 0.88 ± 0.01 a | |
| Lm20 | 6.72 ± 0.05 a | 0.09 ± 0.00 a | 0.94 ± 0.02 a | |
| Lm20_PC | 7.59 ± 0.17 b | 0.09 ± 0.00 a | 0.94 ± 0.02 a | |
| BHIB + 6% NaCl | Lm10 | 13.39 ± 1.50 a | 0.05 ± 0.00 a | 0.70 ± 0.06 a |
| Lm10_PC | 17.17 ± 0.10 b | 0.05 ± 0.00 a | 0.60 ± 0.01 a | |
| Lm20 | 12.75 ± 0.64 a | 0.04 ± 0.00 a | 0.62 ± 0.05 a | |
| Lm20_PC | 15.79 ± 0.38 b | 0.04 ± 0.00 a | 0.60 ± 0.02 a | |
| BHIB pH 5 | Lm10 | 10.13 ± 0.28 a | 0.03 ± 0.01 a | 0.56 ± 0.05 a |
| Lm10_PC | 11.17 ± 0.78 a | 0.03 ± 0.01 a | 0.56 ± 0.09 a | |
| Lm20 | 9.70 ± 1.22 a | 0.04 ± 0.02 a | 0.71 ± 0.32 a | |
| Lm20_PC | 12.01 ± 0.76 b | 0.04 ± 0.01 a | 0.55 ± 0.19 a |
| Target Gene | Primer Sequence | Amplicon Length (bp) | Source |
|---|---|---|---|
| 16S rRNA | Fw- TTAGCTAGTTGGTAGGGT Rv- AATCCGGACAACGCTTGC | 550 | [93] |
| ProW | Fw- GGAAGCCGCATTGGATGTTC Rv- CCTCGTAGCCACCAAAGACA | 78 | This study |
| NADH peroxidase | Fw- ACTGGCGAAGGTGTGAAAGA Rv- GGACGAACCCCAACACTGAT | 116 | This study |
| clpL | Fw- TGGGCACTTAACGGATGGAC Rv- TGATCTCGTTGCTCGTCACC | 117 | This study |
| Cell surface protein (A58 G2 plasmid) | Fw- GGTGGAACTAACCGATGCGA Rv- TGACTTGCTTGTTCGTTGCG | 71 | This study |
| Cell surface protein (Lm228) | Fw- CTATCGCGTGACCAAACGTG Rv- GGCGCAGCTTTCATTTCACT | 71 | This study |
| Lipoprotein | Fw- CTGCTTGCGGCGATTTCTTT Rv- TGCCGACATCATTCCACCAA | 90 | This study |
| Secretion system E (A58 G2 plasmid) | Fw- GCCCCTAAGCCGATAGAACG Rv- ACCCGTTCCAGTTTCTCCAG | 84 | This study |
| Secretion system E (Lm228) | Fw- TCTGTTGGGGCAAAAGCATC Rv- TGCAACAATGGCTTTGCGTT | 77 | This study |
| Multicopper oxidase | Fw- ACAGTCCTTGTGAATGGGAAAGT Rv- AAATCCCGAGCATTGGAGCC | 95 | This study |
| Cadmium ATPase (cadA1, Lm106 and Lm228) | Fw- AAACTGGGAAACGTGGGTGT Rv- CATTTCCAATGGCGGTGACG | 107 | This study |
| Cadmium ATPase (cadA2, A58 G2 plasmid) | Fw- AGAACAAGGGAAAACCGCCA Rv- TACTTCATCCGCGACAGCAA | 75 | This study |
| uvrA | Fw- AGGCTGTATGTCACGGTTGG Rv- GCCACTCCGCAATACCTGAT | 100 | This study |
| ebrA | Fw- GGTATGGGGAGTGCTTTTTACCT Rv- TCCTCCACCAGACCATATTGC | 87 | This study |
| Membrane-bound protease | Fw- CGGTCATTCCCCTAATCAACCT Rv- TTGCCATACAAACCCATGTGC | 73 | This study |
References
- Walker, S.J.; Archer, P.; Banks, J.G. Growth of Listeria monocytogenes at refrigeration temperatures. J. Appl. Bacteriol. 1990, 68, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Sorrells, K.M.; Enigl, D.C.; Hatfield, J.R. Effect of pH, acidulant, time, and temperature on the growth and survival of Listeria monocytogenes. J. Food Prot. 1989, 52, 571–573. [Google Scholar] [CrossRef] [PubMed]
- Shabala, L.; Lee, S.H.; Cannesson, P.; Ross, T. Acid and NaCl limits to growth of Listeria monocytogenes and influence of sequence of inimical acid and NaCl levels on inactivation kinetics. J. Food. Prot. 2008, 71, 1169–1177. [Google Scholar] [CrossRef] [PubMed]
- Yildiz, O.; Aygen, B.; Esel, D.; Kayabas, U.; Alp, E.; Sumerkan, B.; Doganay, M. Sepsis and meningitis due to Listeria monocytogenes. Yonsei Med. J. 2007, 48, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Perez-Diaz, J.C.; Vicente, M.F.; Baquero, F. Plasmids in Listeria. Plasmid 1982, 8, 112–118. [Google Scholar] [CrossRef]
- Peterkin, P.I.; Gardiner, M.-A.; Malik, N.; Idziak, E.S. Plasmids in Listeria monocytogenes and other Listeria species. Can. J. Microbiol. 1992, 38, 161–164. [Google Scholar] [CrossRef] [PubMed]
- Lebrun, M.; Loulergue, J.; Chaslus-Dancla, E.; Audurier, A. Plasmids in Listeria monocytogenes in relation to cadmium resistance. Appl. Environ. Microbiol. 1992, 58, 3183–3186. [Google Scholar]
- Kolstad, J.; Caugant, D.A.A.; Rørvik, L.M.; Rørvik, L.M. Differentiation of Listeria monocytogenes isolates by using plasmid profiling and multilocus enzyme electrophoresis. Int. J. Food Microbiol. 1992, 16, 247–260. [Google Scholar] [CrossRef]
- Fistrovici, E.; Collins-Thompson, D.L. Use of plasmid profiles and restriction endonuclease digest in environmental studies of Listeria spp. from raw milk. Int. J. Food Microbiol. 1990, 10, 43–50. [Google Scholar] [CrossRef]
- Dykes, G.A.; Geornaras, I.; Papathanasopoulos, M.A.; von Holy, A. Plasmid profiles of Listeria species associated with poultry processing. Food Microbiol. 1994, 11, 519–523. [Google Scholar] [CrossRef]
- Elhanafi, D.; Dutta, V.; Kathariou, S. Genetic characterization of plasmid-associated benzalkonium chloride resistance determinants in a Listeria monocytogenes strain from the 1998-1999 outbreak. Appl. Environ. Microbiol. 2010, 76, 8231–8238. [Google Scholar] [CrossRef] [PubMed]
- Rakic-Martinez, M.; Drevets, D.A.; Dutta, V.; Katic, V.; Kathariou, S. Listeria monocytogenes strains selected on ciprofloxacin or the disinfectant benzalkonium chloride exhibit reduced susceptibility to ciprofloxacin, gentamicin, benzalkonium chloride, and other toxic compounds. Appl. Environ. Microbiol. 2011, 77, 8714–8721. [Google Scholar] [CrossRef] [PubMed]
- Katharios-Lanwermeyer, S.; Rakic-Martinez, M.; Elhanafi, D.; Ratani, S.; Tiedje, J.M.; Kathariou, S. Coselection of cadmium and benzalkonium chloride resistance in conjugative transfers from nonpathogenic Listeria spp. to other Listeria. Appl. Environ. Microbiol. 2012, 78, 7549–7556. [Google Scholar] [CrossRef] [PubMed]
- Poyart-Salmeron, C.; Carlier, C.; Trieu-Cuot, P.; Courvalin, P.; Courtieu, A.L. Transferable plasmid-mediated antibiotic resistance in Listeria monocytogenes. Lancet 1990, 335, 1422–1426. [Google Scholar] [CrossRef]
- Hadorn, K.; Hächler, H.; Schaffner, A.; Kayser, F.H. Genetic characterization of plasmid-encoded multiple antibiotic resistance in a strain of Listeria monocytogenes causing endocarditis. Eur. J. Hum. Genet. 1993, 12, 928–937. [Google Scholar]
- Kuenne, C.; Voget, S.; Pischimarov, J.; Oehm, S.; Goesmann, A.; Daniel, R.; Hain, T.; Chakraborty, T. Comparative analysis of plasmids in the genus Listeria. PLoS ONE 2010, 5, e12511. [Google Scholar] [CrossRef]
- Liang, L.; Gnaneshan, S.; Garduño, R. Genome sequence of Listeria monocytogenes plasmid pLM-C-273 carrying genes related to stress resistance. Genome 2016, 4, e01125-16. [Google Scholar] [CrossRef] [PubMed]
- Naditz, A.L.; Dzieciol, M.; Wagner, M.; Schmitz-Esser, S. Plasmids contribute to food processing environment–associated stress survival in three Listeria monocytogenes ST121, ST8, and ST5 strains. Int. J. Food Microbiol. 2019, 299, 39–46. [Google Scholar] [CrossRef] [PubMed]
- McLauchlin, J.; Hampton, M.D.D.; Shah, S.; Threlfall, E.J.J.; Wieneke, A.A.A.; Curtis, G.D.W.D.W. Subtyping of Listeria monocytogenes on the basis of plasmid profiles and arsenic and cadmium susceptibility. J. Appl. Microbiol. 1997, 83, 381–388. [Google Scholar] [CrossRef]
- Harvey, J.; Gilmour, A. Characterization of recurrent and sporadic Listeria monocytogenes isolates from raw milk and nondairy foods by pulsed-field gel electrophoresis, monocin typing, plasmid profiling, and cadmium and antibiotic resistance determination. Appl. Environ. Microbiol. 2001, 67, 840–847. [Google Scholar] [CrossRef]
- Xu, D.; Nie, Q.; Wang, W.; Shi, L.; Yan, H. Characterization of a transferable bcrABC and cadAC genes-harboring plasmid in Listeria monocytogenes strain isolated from food products of animal origin. Int. J. Food 2016, 217, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Lebrun, M.; Audurier, A.; Cossart, P. Plasmid-borne cadmium resistance genes in Listeria monocytogenes are similar to cadA and cadC of Staphylococcus aureus and are induced by cadmium. J. Bacteriol. 1994, 176, 3040–3048. [Google Scholar] [CrossRef] [PubMed]
- Mullapudi, S.; Siletzky, R.M.; Kathariou, S. Diverse cadmium resistance determinants in Listeria monocytogenes isolates from the turkey processing plant environment. Appl. Environ. Microbiol. 2010, 76, 627–630. [Google Scholar] [CrossRef] [PubMed]
- Parsons, C.; Lee, S.; Jayeola, V.; Kathariou, S. Novel cadmium resistance determinant in Listeria monocytogenes. Appl. Environ. Microbiol. 2017, 83, e02580-16. [Google Scholar] [CrossRef]
- Parsons, C.; Lee, S.; Kathariou, S. Heavy metal resistance determinants of the foodborne pathogen Listeria monocytogenes. Genes 2018, 10, 11. [Google Scholar] [CrossRef] [PubMed]
- Little, P.; Martin, M.H. A survey of zinc, lead and cadmium in soil and natural vegetation around a smelting complex. Environ. Pollut. 1972, 3, 241–254. [Google Scholar] [CrossRef]
- Kasuya, M.; Teranishi, H.; Aoshima, K.; Katoh, T.; Horiguchi, H.; Morikawa, Y.; Nishijo, M.; Iwata, K. Water pollution by cadmium and the onset of itai-itai disease. Water Sci. Technol. 1992, 25, 149–156. [Google Scholar] [CrossRef]
- Ryan, P.B.; Huet, N.; Macintoshl, D.L. Longitudinal investigation of exposure to arsenic, cadmium, and lead in drinking water. Env. Heal. Perspect. 2000, 108, 731–735. [Google Scholar] [CrossRef]
- Diaz Ricci, J.C.; Hernández, M.E. Plasmid effects on Escherichia coli metabolism. Crit. Rev. Biotechnol. 2000, 20, 79–108. [Google Scholar] [CrossRef]
- Hingston, P.; Chen, J.; Dhillon, B.K.; Laing, C.; Bertelli, C.; Gannon, V.; Tasara, T.; Allen, K.; Brinkman, F.S.L.; Hansen, L.T.; et al. Genotypes associated with Listeria monocytogenes isolates displaying impaired or enhanced tolerances to cold, salt, acid, or desiccation stress. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef]
- Canchaya, C.; Giubellini, V.; Ventura, M.; de los Reyes-Gavilán, C.G.; Margolles, A. Mosaic-like sequences containing transposon, phage, and plasmid elements among Listeria monocytogenes plasmids. Appl. Environ. Microbiol. 2010, 76, 4851–4857. [Google Scholar] [CrossRef]
- Schmitz-Esser, S.; Müller, A.; Stessl, B. Genomes of sequence type 121 Listeria monocytogenes strains harbor highly conserved plasmids and prophages. Front. Microbiol. 2015, 6, 380. [Google Scholar] [CrossRef]
- Kuenne, C.; Billion, A.; Mraheil, M.A.; Strittmatter, A.; Daniel, R.; Goesmann, A.; Barbuddhe, S.; Hain, T.; Chakraborty, T. Reassessment of the Listeria monocytogenes pan-genome reveals dynamic integration hotspots and mobile genetic elements as major components of the accessory genome. BMC Genom 2013, 14. [Google Scholar] [CrossRef] [PubMed]
- Bohlin, J.; Snipen, L.; Hardy, S. Analysis of intra-genomic GC content homogeneity within prokaryotes. BMC Genom 2010, 11. [Google Scholar] [CrossRef]
- Edwards, D.J.; Holt, K.E. Beginner’s guide to comparative bacterial genome analysis using next-generation sequence data. Microb. Inform. Exp. 2013, 3. [Google Scholar] [CrossRef] [PubMed]
- Lebrun, M.; Audurier, A.; Cossart, P. Plasmid-borne cadmium resistance genes in Listeria monocytogenes are present on Tn5422, a novel transposon closely related to Tn917. J. Bacteriol. 1994, 176, 3049–3061. [Google Scholar] [CrossRef]
- Lee, S.; Rakic-Martinez, M.; Graves, L.M.; Ward, T.J.; Siletzky, R.M.; Kathariou, S. Genetic determinants for cadmium and arsenic resistance among Listeria monocytogenes serotype 4b isolates from sporadic human listeriosis patients. Appl. Environ. Microbiol. 2013, 79, 2471–2476. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Ward, T.J.; Jima, D.D.; Parsons, C.; Kathariou, S. The arsenic resistance-associated Listeria genomic island LGI2 exhibits sequence and integration site diversity and a propensity for three Listeria monocytogenes clones with enhanced virulence. Appl. Environ. Microbiol. 2017, 83, e01189-17. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.; Flynn, S.; Li, Y.; Claesson, M.J.; van Pijkeren, J.-P.; Collins, J.K.; van Sinderen, D.; O’Toole, P.W. Characterization of endogenous plasmids from Lactobacillus salivarius UCC118. Appl. Environ. Microbiol. 2008, 74, 3216–3228. [Google Scholar] [CrossRef]
- Tsuchimoto, S.; Ohtsubo, H.; Ohtsubo, E. Two genes, pemK and pemI, responsible for stable maintenance of resistance plasmid R100. J. Bacteriol. 1988, 170, 1461–1466. [Google Scholar] [CrossRef]
- Sobecky, P.A.; Easter, C.L.; Bear, P.D.; Helinski, D.R. Characterization of the stable maintenance properties of the par region of broad-host-range plasmid RK2. J. Bacteriol. 1996, 178, 2086–2093. [Google Scholar] [CrossRef] [Green Version]
- del Solar, G.; Giraldo, R.; Ruiz-Echevarría, M.J.; Espinosa, M.; Díaz-Orejas, R. Replication and control of circular bacterial plasmids. Microbiol. Mol. Biol. Rev. 1998, 62, 434–464. [Google Scholar]
- Van Melderen, L.; Saavedra De Bast, M. Bacterial toxin–antitoxin systems: More than selfish entities? PLoS Genet. 2009, 5, e1000437. [Google Scholar] [CrossRef]
- Bukowski, M.; Rojowska, A.; Wladyka, B. Prokaryotic toxin-antitoxin systems—The role in bacterial physiology and application in molecular biology. Acta Biochim. Pol. 2011, 58. [Google Scholar] [CrossRef]
- Bukowski, M.; Lyzen, R.; Helbin, W.M.; Bonar, E.; Szalewska-Palasz, A.; Wegrzyn, G.; Dubin, G.; Dubin, A.; Wladyka, B. A regulatory role for Staphylococcus aureus toxin–antitoxin system PemIKSa. Nat. Commun. 2013, 4. [Google Scholar] [CrossRef]
- Zielenkiewicz, U.; Ceglowski, P. The toxin-antitoxin system of the streptococcal plasmid pSM19035. J. Bacteriol. 2005, 187, 6094–6105. [Google Scholar] [CrossRef]
- Sletvold, H.; Johnsen, P.J.; Hamre, I.; Simonsen, G.S.; Sundsfjord, A.; Nielsen, K.M. Complete sequence of Enterococcus faecium pVEF3 and the detection of an ω-ε-ζ toxin–antitoxin module and an ABC transporter. Plasmid 2008, 60, 75–85. [Google Scholar] [CrossRef]
- Hayes, F. A family of stability determinants in pathogenic bacteria. J. Bacteriol. 1998, 180, 6415–6418. [Google Scholar]
- Mullins, K.E.; Hang, J.; Clifford, R.J.; Onmus-Leone, F.; Yang, Y.; Jiang, J.; Leguia, M.; Kasper, M.R.; Maguina, C.; Lesho, E.P.; et al. Whole-genome analysis of Bartonella ancashensis, a novel pathogen causing verruga peruana, rural Ancash Region, Peru. Emerg. Infect. Dis. 2017, 23, 430–438. [Google Scholar] [CrossRef]
- Kaplan, E.; Sela, N.; Doron-Faigenboim, A.; Navon-Venezia, S.; Jurkevitch, E.; Cytryn, E. Genomic and functional characterization of qnr-encoding plasmids from municipal wastewater biosolid Klebsiella pneumoniae isolates. Front. Microbiol. 2015, 6, 1354. [Google Scholar] [CrossRef]
- Christen, S.; Srinivas, A.; Bä, P.; Zeller, A.; Pridmore, D.; Bieniossek, C.; Baumann, U.; Erni, B. Regulation of the dha operon of Lactococcus lactis a deviation from the rule followed by the tetr family of transcription regulators. J. Biol. Chem. 2006, 281, 23129–23137. [Google Scholar] [CrossRef]
- Erni, B.; Siebold, C.; Christen, S.; Srinivas, A.; Oberholzer, A.; Baumann, U. Small substrate, big surprise: Fold, function and phylogeny of dihydroxyacetone kinases. Cell. Mol. Life Sci. 2006, 63, 890–900. [Google Scholar] [CrossRef]
- Masaoka, Y.; Ueno, Y.; Morita, Y.; Kuroda, T.; Mizushima, T.; Tsuchiya, T. A two-component multidrug efflux pump, EbrAB, in Bacillus subtilis. J. Bacteriol. 2000, 182, 2307–2310. [Google Scholar] [CrossRef] [PubMed]
- Schulein, R.; Dehio, C. The VirB/VirD4 type IV secretion system of Bartonella is essential for establishing intraerythrocytic infection. Mol. Microbiol. 2002, 46, 1053–1067. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.K.; Huang, K.C. The role of hydrolases in bacterial cell-wall growth. Curr. Opin. Microbiol. 2013, 16, 760–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Gorski, L.; Reynolds, J.; Orozco, E. Role of uvrA in the growth and survival of Listeria monocytogenes under UV radiation and acid and bile stress. J. Food Prot. 2006, 69, 3031–3036. [Google Scholar] [CrossRef] [PubMed]
- Payne, A.; Schmidt, T.B.; Nanduri, B.; Pendarvis, K.; Pittman, J.R.; Thornton, J.A.; Grissett, J.; Donaldson, J.R. Proteomic analysis of the response of Listeria monocytogenes to bile salts under anaerobic conditions. J. Med. Microbiol. 2013, 62, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Prieto, A.I.; Ramos-Morales, F.; Casadesús, J. Repair of DNA damage induced by bile salts in Salmonella enterica. Genetics 2006, 174, 575–584. [Google Scholar] [CrossRef]
- Navarre, W.W.; Schneewind, O. Surface proteins of Gram-positive bacteria and mechanisms of their targeting to the cell wall envelope. Microbiol. Mol. Biol. Rev. 1999, 63, 174–229. [Google Scholar]
- Bierne, H.; Cossart, P. Listeria monocytogenes surface proteins: From genome predictions to function. Microbiol. Mol. Biol. Rev. 2007, 71, 377–397. [Google Scholar] [CrossRef]
- Kroos, L.; Akiyama, Y. Biochemical and structural insights into intramembrane metalloprotease mechanisms. Biochim. Biophys. Acta Biomembr. 2013, 1828, 2873–2885. [Google Scholar] [CrossRef] [Green Version]
- Woodward, J.J.; Iavarone, A.T.; Portnoy, D.A. c-di-AMP secreted by intracellular Listeria monocytogenes activates a host type I interferon response. Science 2010, 328, 1703–1705. [Google Scholar] [CrossRef]
- Gibson, C.M.; Mallett, T.C.; Claiborne, A.; Caparon, M.G. Contribution of NADH oxidase to aerobic metabolism of Streptococcus pyogenes. J. Bacteriol. 2000, 182, 448–455. [Google Scholar] [CrossRef]
- Sakamoto, M.; Komagata, K. Aerobic growth of and activities of NADH oxidase and NADH peroxidase in lactic acid bacteria. J. Ferment. Bioeng. 1996, 82, 210–216. [Google Scholar] [CrossRef]
- Ko, R.; Smith, L.T.; Smith, G.M. Glycine betaine confers enhanced osmotolerance and cryotolerance on Listeria monocytogenes. J. Bacteriol. 1994, 176, 426–431. [Google Scholar] [CrossRef] [PubMed]
- Gerhardt, P.N.; Tombras Smith, L.; Smith, G.M. Osmotic and chill activation of glycine betaine porter II in Listeria monocytogenes membrane vesicles. J. Bacteriol. 2000, 182, 2544–2550. [Google Scholar] [CrossRef] [PubMed]
- Ko, R.; Smith, L.T. Identification of an ATP-driven, osmoregulated glycine betaine transport system in Listeria monocytogenes. Appl. Environ. Microbiol. 1999, 65, 4040–4048. [Google Scholar]
- Khedr, A.; Abbas, M.; Wahid, A. Proline induces the expression of salt-stress-responsive proteins and may improve the adaptation of Pancratium maritimum L. to salt-stress. J. Exp. Bot. 2003, 54, 2553–2562. [Google Scholar] [CrossRef]
- Yoshiba, Y.; Kiyosue, T.; Nakashima, K. Regulation of levels of proline as an osmolyte in plants under water stress. Plant. Cell Physiol. 1997, 38, 1095–1102. [Google Scholar] [CrossRef]
- Sleator, R.D.; Hill, C. Bacterial osmoadaptation: The role of osmolytes in bacterial stress and virulence. FEMS Microbiol. Rev. 2002, 26, 49–71. [Google Scholar] [CrossRef]
- Kempf, B.; Bremer, E. Uptake and synthesis of compatible solutes as microbial stress responses to high-osmolality environments. Arch. Microbiol. 1998, 170, 319–330. [Google Scholar] [CrossRef]
- Frees, D.; Savijoki, K.; Varmanen, P.; Ingmer, H. Clp ATPases and ClpP proteolytic complexes regulate vital biological processes in low GC, Gram-positive bacteria. Mol. Microbiol. 2007, 63, 1285–1295. [Google Scholar] [CrossRef]
- Wall, T.; Båth, K.; Britton, R.; Jonsson, H. The early response to acid shock in Lactobacillus reuteri involves the ClpL chaperone and a putative cell wall-altering esterase. Appl. Environ. Microbiol. 2007, 73, 3924–3935. [Google Scholar] [CrossRef]
- Varcamonti, M.; Arsenijevic, S. Expression of the heat shock gene clpL of Streptococcus thermophilus is induced by both heat and cold shock. Microb. Cell Fact. 2006, 5. [Google Scholar] [CrossRef]
- Gaillot, O.; Pellegrini, E.; Bregenholt, S. The ClpP serine protease is essential for the intracellular parasitism and virulence of Listeria monocytogenes. Mol. Microbiol. 2000, 35, 1286–1294. [Google Scholar] [CrossRef]
- Pöntinen, A.; Aalto-Araneda, M.; Lindström, M.; Korkeala, H. Heat resistance mediated by pLM58 plasmid-borne ClpL in Listeria monocytogenes. mSphere 2017, 2, e00364-17. [Google Scholar] [CrossRef]
- Huston, W.; Jennings, M.; McEwan, A.G. The multicopper oxidase of Pseudomonas aeruginosa is a ferroxidase with a central role in iron acquisition. Mol. Microbiol. 2002, 45, 1741–1750. [Google Scholar] [CrossRef]
- Askwith, C.; Eide, D.; Van Ho, A.; Bernard, P.; Li, L.; Davis-Kaplan, S.; Sipe, D.M.; Kaplan, J. The FET3 gene of S. cerevisiae encodes a multicopper oxidase required for ferrous iron uptake. Cell 1994, 76, 403–410. [Google Scholar] [CrossRef]
- Cornelis, P.; Wei, Q.; Andrews, S.C.; Vinckx, T. Iron homeostasis and management of oxidative stress response in bacteria. Metallomics 2011, 3, 540–549. [Google Scholar] [CrossRef]
- Nucifora, G.; Chu, L.; Misra, T.; Silver, S. Cadmium resistance from Staphylococcus aureus plasmid pI258 cadA gene results from a cadmium-efflux ATPase. Proc. Natl. Acad. Sci. USA 1989, 86, 3544–3548. [Google Scholar] [CrossRef]
- Casey, A.; Fox, E.M.; Schmitz-Esser, S.; Coffey, A.; McAuliffe, O.; Jordan, K. Transcriptome analysis of Listeria monocytogenes exposed to biocide stress reveals a multi-system response involving cell wall synthesis, sugar uptake, and motility. Front. Microbiol. 2014, 5, 68. [Google Scholar] [CrossRef]
- Lin, J.; Sahin, O.; Michel, L.; Zhang, Q. Critical role of multidrug efflux pump CmeABC in bile resistance and in vivo colonization of Campylobacter jejuni. Infect. Immun. 2003, 71, 4250–4259. [Google Scholar] [CrossRef]
- Romanova, N.A.; Wolffs, P.F.G.; Brovko, L.Y.; Griffiths, M.W. Role of efflux pumps in adaptation and resistance of Listeria monocytogenes to benzalkonium chloride. Appl. Environ. Microbiol. 2006, 72, 3498–3503. [Google Scholar] [CrossRef]
- Feng, J.; Gu, Y.; Wang, J.; Song, C.; Yang, C.; Xie, H.; Zhang, W.; Wang, S. Curing the plasmid pMC1 from the poly (γ-glutamic Acid) producing Bacillus amyloliquefaciens LL3 strain using plasmid incompatibility. Appl. Biochem. Biotechnol. 2013, 171, 532–542. [Google Scholar] [CrossRef]
- Goverde, R.L.J.; Kusters, J.G.; Huis in‘t Veld, J.H.J. Growth rate and physiology of Yersinia enterocolitica; influence of temperature and presence of the virulence plasmid. J. Appl. Bacteriol. 1994, 77, 96–104. [Google Scholar] [CrossRef]
- Logue, C.M.; Sherwood, J.S.; Doetkott, C. Growth studies of plasmid bearing and plasmid cured Yersinia enterocolitica GER O:3 in the presence of cefsulodin, irgasan and novobiocin at 25 and 37 °C. J. Appl. Microbiol. 2006, 100, 1299–1306. [Google Scholar] [CrossRef]
- Milillo, M.S. Resistance to food processing sanitizers and heavy metals in Listeria monocytogenes from British Columbia, Canada and antibiogram profiles of clinically relevant Listeria monocytogenes from British Columbia and Alberta, Canada. Masters Thesis, University of British Columbia, Vancouver, BC, Canada, 2015. [Google Scholar]
- Darling, A.A.E.; Mau, B.; Perna, N.N.T. progressiveMauve: multiple genome alignment with gene gain, loss and rearrangement. PLoS ONE 2010, 5, e11147. [Google Scholar] [CrossRef]
- Brettin, T.; Davis, J.; Disz, T.; Edwards, R.; Gerdes, S. RASTtk: A modular and extensible implementation of the RAST algorithm for building custom annotation pipelines and annotating batches of genomes. Sci. Rep. 2015, 5, 8365. [Google Scholar] [CrossRef]
- Overbeek, R.; Olson, R.; Pusch, G.; Olsen, G. The SEED and the Rapid Annotation of microbial genomes using Subsystems Technology (RAST). Nucleic Acids Res. 2013, 42, D206–D214. [Google Scholar] [CrossRef]
- Aziz, R.; Bartels, D.; Best, A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: rapid annotations using subsystems technology. BMC Genom. 2008, 9. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Tasara, T.; Stephan, R. Genes in Listeria monocytogenes as potential internal control references for normalizing mRNA expression levels in stress adaptation models using real-time PCR. FEMS Microbiol. Lett. 2007, 269, 265–272. [Google Scholar] [CrossRef]
- Baranyi, J.; Roberts, T.A. A dynamic approach to predicting bacterial growth in food. Int. J. Food Microbiol. 1994, 23, 277–294. [Google Scholar] [CrossRef]






| Plasmid Type | Sub-Group a | # of Strains b | Strain Origin(s) c | Serotype d,e or Species | Clonal Complex d | Plasmid Size (bp) d,f | # of Plasmid Contigs | Plasmid %GC | # of Predicted Genes | # of MGEs g |
|---|---|---|---|---|---|---|---|---|---|---|
| pLM33 ǂ | MC | - | - | 1/2b | - | 32307 | closed | 36.16 | 36 | 9 |
| pLMG1-1 | MC | 3 | CH | 1/2a, 1/2c (2) | 9 | 25605 | 1 | 36.85 | 29 | 8 |
| pLMG1-2 | MC | 1 | CH | 1/2c | 9 | 38056 | 3 | 36.85 | 42 | 8 |
| pLMG1-3 | MC | 1 | AB | 1/2a | Singleton (ST 839) | 40730 | 3 | 36.85 | 48 | 6 |
| pLMG1-4 | MC | 3 | AB | 1/2c | 9 | 44350 | 3 | 36.85 | 54 | 11 |
| pLMG1-5 | MC | 7 | AB | 1/2c | 9 | 48409 (6), 48460 | 3 | 36.85 | 52 | 12 |
| pLMG1-6 | Outlier | 4 | BC | 4b | 6 | 54735, 54736 (3) | 5 | 34.77 | 64 | 9 |
| pCT100 ǂ | Outlier | - | - | L. innocua | - | 37279 | closed | 34.07 | 34 | 6 |
| pLM1-2bUG1 ǂ | MC | - | - | 1/2b | - | 57780 | closed | 36.04 | 63 | 16 |
| pLMG1-7 | MC | 26 | AB (18), BC (4), CH (4) | 1/2a (17), 1/2b (6), 1/2c (2), 3b | 3 (5), 5 (2), 7 (9), 9 (3), 11 (5), 89, 193 | 57082, 57083 (25) | 3 | 36.04 | 64 | 13 |
| pLM7UG1 ǂ | MC | - | - | 7 | - | 50100 | closed | 35.48 | 55 | 13 |
| pLMG1-8 | MC | 3 | AB, BC (2) | 1/2c | 9 | 58105 | 3 | 36.65 | 69 | 13 |
| pLMG1-9* | MC | 1 | AB | 1/2b | 5 | 62258 | 2 | 35.58 | 75 | 17 |
| pLMG1-10 | MC | 1 | AB | 1/2a/3a | 199 | 70385 | 3 | 36.36 | 83 | 17 |
| pLMG1-11 | G2 Sub2 | 1 | BC | 1/2c | 9 | 75351 | 7 | 36.85 | 88 | 13 |
| pLMG1-12 | MC | 2 | AB, BC | 1/2b | 5 | 78240, 78245 | 4 | 36.14 | 95 | 18 |
| pLMG1-13 | Outlier | 5 | BC | 1/2a | 11 | 87487 (2), 87488, 87574 (2) | 1 | 34.38 | 113 | 15 |
| Plasmid Type | Sub-Group a | # of Strains b | Strain Origin(s) c,d | Serotype d,e or Species | Clonal Complex d | Plasmid Size (bp) d,f | # of Plasmid Contigs | Plasmid %GC | # of Predicted Genes | # of MGEs g |
|---|---|---|---|---|---|---|---|---|---|---|
| pLMG2-1 | Sub1 | 1 | AB | 1/2a | 8 | 55472 | 1 | 37.55 | 63 | 1 |
| pLMG2-2 | Sub1 | 1 | CH | 1/2a | 121 | 61053 | 1 | 36.85 | 64 | 4 |
| pLM80 ǂ | Sub 2 | - | - | 4b | - | 81588 | 2 | 37.54 | 88 | 11 |
| pLMG2-3 | Sub1 | 6 | BC | 1/2a (4), 3a (2) | 321 | 66447 | 3 | 36.85 | 74 | 6 |
| pLMG2-4 | Sub2 | 6 | AB | 1/2a (6) | 8 | 77109, 77221 (2), 77229 (3) | 1 | 36.85 | 83 | 9 |
| pLM5578 ǂ | Sub 1 | - | - | 1/2a | - | 77054 | closed | 36.59 | 76 | 11 |
| pLMG2-5 | Sub1 | 2 | AB | 1/2a | 8, singleton (ST 1018) | 77249 | 1 | 36.85 | 83 | 10 |
| pLGUG1 ǂ | Sub 2 | - | - | L. grayi | - | 79249 | closed | 36.78 | 99 | 8 |
| pLMG2-6 | Sub1 | 2 | AB | 1/2c | 9 | 81510 | 2 | 36.85 | 87 | 5 |
| pLMG2-7 | Sub2 | 3 | AB, BC (2) | 1/2a, 1/2b (2) | 7, 88 (2) | 81644 | 1 | 36.85 | 92 | 7 |
| pLMG2-8* | Sub2 | 1 | AB | 1/2b | 5 | 87369 | 3 | 37.71 | 97 | 10 |
| pLMG2-9 | Sub2 | 1 | BC | 1/2a | 155 | 89025 | 2 | 37.55 | 99 | 10 |
| pLMG2-10 | Sub2 | 4 | AB, BC, CH (2) | 1/2a (2), 1/2b (2) | 5 (2), 204 (2) | 90543 | 3 | 36.85 | 99 | 11 |
| pLI100 ǂ | Sub 2 | - | - | L. innocua | - | 81905 | closed | 35.52 | 84 | 24 |
| pLMG2-11 | Sub1 | 6 | AB (5), BC | 1/2a | 8 | 92204 (5), 99205 | 1 | 36.85 | 98 | 10 |
| pLMG2-12 | Sub1 | 1 | AB | 1/2a | 8 | 98358 | 2 | 36.85 | 108 | 12 |
| pLMG2-13 | Sub1 | 1 | CH | 1/2b | 59 | 107184 | 2 | 36.85 | 120 | 11 |
| Predicted Protein | G1 Plasmid Types | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | |
| Acyltransferase family protein | |||||||||||||
| Alcohol dehydrogenase | |||||||||||||
| Arsenic efflux pump protein | |||||||||||||
| ATPase involved in DNA repair | |||||||||||||
| Cadmium resistance protein | |||||||||||||
| Copper-transporting ATPase | |||||||||||||
| CRISPR-associated protein MTH1087 | |||||||||||||
| Dihydroxyacetone kinase, ATP-dependent | |||||||||||||
| DUF1706 domain-containing protein | |||||||||||||
| Epsilon antitoxin | |||||||||||||
| Glutathione-dependent formaldehyde dehydrogenase | |||||||||||||
| Glycerol dehydrogenase | |||||||||||||
| Glycerol kinase | |||||||||||||
| Hypothetical lambda repressor-like, DNA-binding | |||||||||||||
| Integral membrane protein | |||||||||||||
| Lmo0466 protein | |||||||||||||
| Lmo2276 protein | |||||||||||||
| Membrane proteins related to metalloendopeptidases | |||||||||||||
| Mercuric ion reductase | |||||||||||||
| Mercuric resistance operon regulatory protein | |||||||||||||
| Methyl-accepting chemotaxis protein | |||||||||||||
| Multi antimicrobial extrusion (MATE) family transporter | |||||||||||||
| Myosin heavy chain, nonmuscle type B | |||||||||||||
| Na(+)/H(+) antiporter | |||||||||||||
| Organomercurial lyase | |||||||||||||
| Oxidoreductase (putative) | |||||||||||||
| Permease of the drug/metabolite transporter superfamily | |||||||||||||
| Phage protein | |||||||||||||
| Phosphate regulon transcriptional regulatory protein PhoB | |||||||||||||
| Phosphoenolpyruvate-dihydroxyacetone phosphotransferase ADP-binding subunit DhaL | |||||||||||||
| Phosphoenolpyruvate-dihydroxyacetone phosphotransferase dihydroxyacetone binding subunit DhaK | |||||||||||||
| Phosphoenolpyruvate-dihydroxyacetone phosphotransferase subunit DhaM | |||||||||||||
| Predicted transcriptional regulator of pyridoxine metabolism | |||||||||||||
| Prophage LambdaSa2, site-specific recombinase | |||||||||||||
| Protein involved in cell division | |||||||||||||
| Protoporphyrinogen IX oxidase, novel form, HemJ | |||||||||||||
| pXO2-10 | |||||||||||||
| RelB/StbD replicon stabilization protein (antitoxin to RelE/StbE) | |||||||||||||
| RelE/StbE replicon stabilization toxin | |||||||||||||
| RepB | |||||||||||||
| Replication-associated protein RepB | |||||||||||||
| Rlx-like protein | |||||||||||||
| Site-specific recombinase, DNA invertase | |||||||||||||
| Site-specific recombinase, phage integrase family | |||||||||||||
| Sortase A, LPXTG specific | |||||||||||||
| Tn916, transcriptional regulator, putative | |||||||||||||
| Transcriptional regulator, PadR family | |||||||||||||
| Transcriptional regulator, XRE family | |||||||||||||
| Transcriptional repressor, BlaI/MecI family | |||||||||||||
| Transposase, IS204/IS1001/IS1096/IS1165 | |||||||||||||
| Type I restriction-modification system, restriction subunit R | |||||||||||||
| Zeta toxin | |||||||||||||
| Predicted Protein | G2 Plasmid Types | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | |
| ABC transporter | |||||||||||||
| Cell surface protein | |||||||||||||
| Chromosome (plasmid) partitioning protein ParA | |||||||||||||
| Conjugation protein, TraG/TraD family, (pXO2-16) | |||||||||||||
| Conserved hypothetical protein | |||||||||||||
| General secretion pathway protein E | |||||||||||||
| Hypothetical protein, (pXO1-65) | |||||||||||||
| Hypothetical protein, (pXO2-28) | |||||||||||||
| Invasion associated protein p60 | |||||||||||||
| Lipoprotein, NLP/P60 family | |||||||||||||
| Membrane protein, putative, (pXO2-14) | |||||||||||||
| Membrane-bound protease, CAAX family | |||||||||||||
| Phage protein lin1266 | |||||||||||||
| Pli0009 protein | |||||||||||||
| Pli0068 protein | |||||||||||||
| Secreted antigen GbpB/SagA/PcsB, putative peptidoglycan hydrolase | |||||||||||||
| Thermonuclease | |||||||||||||
| Tn5252, Orf 21 protein, internal deletion | |||||||||||||
| TolA protein | |||||||||||||
| TraG/TraD family protein | |||||||||||||
| Type IV secretory pathway, VirD4 components | |||||||||||||
| Type V secretory pathway, adhesin AidA | |||||||||||||
| Strain | Origin | Serotype | Plasmid Type a | Notable Characteristics |
|---|---|---|---|---|
| A58 | AB | 1/2b | pLMG1-9 pLMG2-4 | - Acid tolerant - Contains two plasmids |
| Lm10 | BC | 1/2a | pLMG2-9 | |
| Lm10_PC | 1/2a | N/A | - Plasmid-cured Lm10 | |
| Lm20 | BC | 1/2c | pLMG1-11 | |
| Lm20_PC | 1/2c | N/A | - Plasmid-cured Lm20 | |
| Lm106 | BC | 1/2b | pLMG1-12 | |
| Lm228 | CH | 1/2b | pLMG2-13 | - Acid/salt tolerant - Harbours the largest plasmid in our strain collection |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hingston, P.; Brenner, T.; Truelstrup Hansen, L.; Wang, S. Comparative Analysis of Listeria monocytogenes Plasmids and Expression Levels of Plasmid-Encoded Genes during Growth under Salt and Acid Stress Conditions. Toxins 2019, 11, 426. https://doi.org/10.3390/toxins11070426
Hingston P, Brenner T, Truelstrup Hansen L, Wang S. Comparative Analysis of Listeria monocytogenes Plasmids and Expression Levels of Plasmid-Encoded Genes during Growth under Salt and Acid Stress Conditions. Toxins. 2019; 11(7):426. https://doi.org/10.3390/toxins11070426
Chicago/Turabian StyleHingston, Patricia, Thomas Brenner, Lisbeth Truelstrup Hansen, and Siyun Wang. 2019. "Comparative Analysis of Listeria monocytogenes Plasmids and Expression Levels of Plasmid-Encoded Genes during Growth under Salt and Acid Stress Conditions" Toxins 11, no. 7: 426. https://doi.org/10.3390/toxins11070426
APA StyleHingston, P., Brenner, T., Truelstrup Hansen, L., & Wang, S. (2019). Comparative Analysis of Listeria monocytogenes Plasmids and Expression Levels of Plasmid-Encoded Genes during Growth under Salt and Acid Stress Conditions. Toxins, 11(7), 426. https://doi.org/10.3390/toxins11070426