Venomics and Cellular Toxicity of Thai Pit Vipers (Trimeresurus macrops and T. hageni)


 ,
,
Abstract
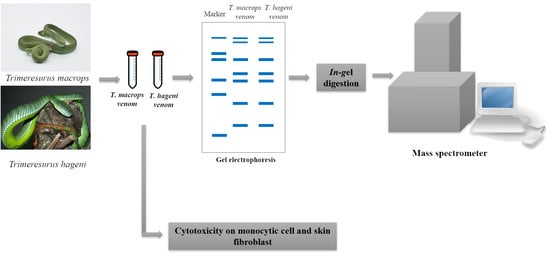
:
1. Introduction
2. Results
2.1. Proteomics Analysis of T. macrops and T. hageni Venom
2.2. Cellular Toxicity of T. macrops and T. hageni Venoms on U937 Monocytic Cells and CRL-1474 Skin Fibroblasts
3. Discussion
4. Materials and Methods
4.1. Venom Collection
4.2. T. macrops and T. hageni Venom Protein Separation
4.3. In-Gel Digestion
4.4. Mass Spectrometric Analysis
4.5. Cell Culture
4.6. Venom Cytotoxicity on Monocytic Cell (U937 Cell) and Skin Fibroblasts (CRL-1474)
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Arruda, L.K.; Santos, A.B. Immunologic responses to common antigens in helminthic infections and allergic disease. Curr. Opin. Allergy Clin. Immunol. 2005, 5, 399–402. [Google Scholar] [CrossRef]
- Malhotra, A.; Thorpe, R.S. A phylogeny of the Trimeresurus group of pit vipers: New evidence from a mitochondrial gene tree. Mol. Phylogenet. Evol. 2000, 16, 199–211. [Google Scholar] [CrossRef]
- Kurohmaru, M.; Matsui, T.; Igarashi, H.; Hattori, S.; Hayashi, Y. Distribution of actin filaments in the seminiferous epithelium of the Habu, Trimeresurus flavoviridis. Anat. Histol. Embryol. 2019, 48, 505–507. [Google Scholar] [CrossRef]
- Blessmann, J.; Nguyen, T.P.N.; Bui, T.P.A.; Krumkamp, R.; Vo, V.T.; Nguyen, H.L. Incidence of snakebites in 3 different geographic regions in Thua Thien Hue province, central Vietnam: Green pit vipers and cobras cause the majority of bites. Toxicon 2018, 156, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Blessmann, J.; Khonesavanh, C.; Outhaithit, P.; Manichanh, S.; Somphanthabansouk, K.; Siboualipha, P. Venomous snake bites in Lao PDR: A retrospective study of 21 snakebite victims in a provincial hospital. Southeast Asian J. Trop. Med. Public Health 2010, 41, 195–202. [Google Scholar] [PubMed]
- Hon, K.L.; Kwok, L.W.; Leung, T.F. Snakebites in children in the densely populated city of Hong Kong: A 10-year survey. Acta Paediatr. 2004, 93, 270–272. [Google Scholar] [CrossRef] [PubMed]
- Hung, D.Z. Taiwan’s venomous snakebite: Epidemiological, evolution and geographic differences. Trans. R. Soc. Trop. Med. Hyg. 2004, 98, 96–101. [Google Scholar] [CrossRef]
- Viravan, C.; Looareesuwan, S.; Kosakarn, W.; Wuthiekanun, V.; McCarthy, C.J.; Stimson, A.F.; Bunnag, D.; Harinasuta, T.; Warrell, D.A. A national hospital-based survey of snakes responsible for bites in Thailand. Trans. R. Soc. Trop. Med. Hyg. 1992, 86, 100–106. [Google Scholar] [CrossRef]
- Zeng, X.; Hu, J.; Liang, X.; Wu, Y.; Yan, M.; Zhu, M.; Fu, Y. Acute cerebral infarction following a Trimeresurus stejnegeri snakebite: A case report. Medicine 2019, 98, e15684. [Google Scholar] [CrossRef]
- Witharana, E.W.R.A.; Gnanathasan, A.; Dissanayake, A.S.; Wijesinghe, S.K.J.; Kadahetti, S.C.L.; Rajapaksha, R.M.J.K. Sri Lankan green pit viper (Trimeresurus trigonocephalus) bites in Deniyaya: A clinico-epidemiological study. Toxicon 2019, S0041, 30414–30423. [Google Scholar] [CrossRef]
- Chotenimitkhun, R.; Rojnuckarin, P. Systemic antivenom and skin necrosis after green pit viper bites. Clin. Toxicol 2008, 46, 122–125. [Google Scholar] [CrossRef] [PubMed]
- Gowda, R.; Rajaiah, R.; Angaswamy, N.; Krishna, S.; Bannikuppe, V.S. Biochemical and pharmacological characterization of Trimersurus malabaricus snake venom. J. Cell. Biochem. 2018, 119, 5904–5912. [Google Scholar] [CrossRef] [PubMed]
- Greene, S.; Galdamez, L.A.; Tomasheski, R. White-Lipped tree viper (Cryptelytrops albolabris) envenomation in an American viper keeper. J. Emerg. Med. 2017, 53, e115–e118. [Google Scholar] [CrossRef] [PubMed]
- Sivaganabalan, R.; Ismail, A.K.; Salleh, M.S.; Mohan, K.; Choo, T.C.; Adnan, A.; Ariff, A.M.; Mohamed, Z.; Thevarajah, N.; Daud, R.; et al. Guideline: Management of Snakebite Ministry of Health Malaysia, 1st ed.; Ministry of Health Malaysia: Putrajaya, Malaysia, 2017.
- Tu, Y.Y.; Chen, C.C.; Chang, H.M. Isolation of immunoglobulin in yolk (IgY) and rabbit serum immunoglobulin (IgG) specific against bovine lactoferrin by immunoaffinity chromatography. Food Res. Int. 2001, 34, 783–789. [Google Scholar] [CrossRef]
- Ralidis, M.P. Medical treatment of reptile envenomation: A review of the current literature. Top. Emerg. Med. 2000, 22, 16–36. [Google Scholar]
- Calvete, J.J.; Sanz, L.; Angulo, Y. Venoms, venomics, antivenomics. FEBS Lett. 2009, 583, 1736–1743. [Google Scholar] [CrossRef] [Green Version]
- Jones, B.K.; Saviola, A.J.; Reilly, S.B.; Stubbs, A.L.; Arida, E.; Iskandar, D.T.; McGuire, J.A.; Yates, J.R.; Mackessy, S.P. Venom Composition in a Phenotypically Variable Pit Viper (Trimeresurus insularis) across the Lesser Sunda Archipelago. J. Proteome Res. 2019, 18, 2206–2220. [Google Scholar] [CrossRef]
- Tan, C.H.; Tan, K.Y.; Ng, T.S.; Quah, E.S.H.; Ismail, A.K.; Khomvilai, S.; Sitprija, V.; Tan, N.H. Venomics of Trimeresurus (Popeia) nebularis, the Cameron Highlands Pit Viper from Malaysia: Insights into Venom Proteome, Toxicity and Neutralization of Antivenom. Toxins 2019, 11, 95. [Google Scholar] [CrossRef] [Green Version]
- Soogarun, S.; Sangvanich, P.; Chowbumroongkait, M.; Jiemsup, S.; Wiwanikit, V.; Pradniwat, P.; Palasuwan, A.; Pawinwongchai, J.; Chanprasert, S.; Moungkote, T. Analysis of green pit viper (Trimeresurus alborabris) venom protein by LC/MS-MS. J. Biochem. Mol. Toxicol. 2008, 22, 225–229. [Google Scholar] [CrossRef]
- Rojnuckarin, P.; Banjongkit, S.; Chantawibun, W.; Akkawat, B.; Juntiang, J.; Noiphrom, J.; Pakmanee, N. Intragumtornchai, Green pit viper (Trimeresurus albolabris and T. macrops) venom antigenaemia and kinetics in humans. Trop. Doct. 2007, 37, 207–210. [Google Scholar] [CrossRef]
- Wilson, D.; Daly, N.L. Venomics: A mini-review. High-Throughput. 2018, 7, 19. [Google Scholar] [CrossRef] [Green Version]
- Whittington, C.M.; Papenfuss, A.T.; Locke, D.P.; Mardis, E.R.; Wilson, R.K.; Abubucker, S.; Mitreva, M.; Wong, E.S.; Hsu, A.L.; Kuchel, P.W.; et al. Novel venom gene discovery in the platypus. Genome Biol. 2010, 11, R95. [Google Scholar] [CrossRef] [Green Version]
- Leslie, C.C. Properties and regulation of cytosolic phospholipase A2. J. Biol. Chem. 1997, 272, 16709–16712. [Google Scholar] [CrossRef] [Green Version]
- Harris, J.B.; Scott-Davey, T. Secreted phospholipases A2 of snake venoms: Effects on the peripheral neuromuscular system with comments on the role of phospholipases A2 in disorders of the CNS and their uses in industry. Toxins 2013, 17, 2533–2571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casewell, N.R.; Wagstaff, S.C.; Wűster, W.; Cook, D.A.N.; Bolton, F.M.S. Medically important differences in snake venom composition are dictated by distinct postgenomic mechanisms. Proc. Natl. Acad. Sci. USA 2014, 111, 9205–9210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Sousa, M.V.; Morhy, L.; Arni, R.K.; Ward, R.J.; Díaz, C.; Gutiérrez, J.M. Amino acid sequence of a myotoxic Lys49-phospholipase A2 homologue from the venom of Cerrophidion (Bothrops) godmani. Biochim. Biophys. Acta 1998, 1384, 204–208. [Google Scholar] [CrossRef]
- Markland, F.S. Snake venoms and the hemostatic system. Toxicon 2000, 36, 1749–1800. [Google Scholar] [CrossRef]
- Matsui, T.; Fujimura, Y.; Titani, K. Snake venom proteases affecting hemostasis and thrombosis. Biochim Biophys. Acta 2000, 1477, 146–156. [Google Scholar] [CrossRef]
- Iwanaga, S.; Takeya, H. Structure and function of snake venom metalloproteinase family. In Methods in Protein Sequence Analysis; Plenum Press: New York, NY, USA, 1993; pp. 107–115. [Google Scholar]
- Bjarnason, J.B.; Fox, J.W. Snake venom metalloendopeptidases: Reprolysins. Methods Enzymol. 1995, 248, 345–368. [Google Scholar]
- McLane, M.A.; Sanchez, E.E.; Wong, A.; Paquette-Straub, C.; Perez, J.C. Disintegrins. Curr. Drug Targets Cardiovasc. Haematol. Disord. 2004, 4, 327–355. [Google Scholar] [CrossRef]
- Normand, P. Snake Venom Disintegrins and Cell Migration. Toxins 2010, 2, 2606–2621. [Google Scholar]
- Krebs, H.A. The Enzymes, 1st ed.; Delmar Publishers: New York, NY, USA, 1933. [Google Scholar]
- Ande, S.R.; Fussi, H.; Knauer, H.; Murkovic, M.; Ghisla, S.; Fröhlich, K.U.; Macheroux, P. Induction of apoptosis in yeast by L-amino acid oxidase from the Malayan pit viper Calloselasma rhodostoma. Yeast 2008, 25, 349–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clemetson, K.J.; Lu, Q.; Clemetson, J.M. Snake C-type lectin-like proteins and platelet receptors. Pathophysiol. Haemost. Thromb. 2005, 34, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Shikamoto, Y.; Fujimoto, Z.; Morita, T.; Mizuno, H. Crystallization and preliminary X-ray analysis of coagulation factor IX-binding protein from habu snake venom at pH 6.5 and 4.6. Acta Crystallogr. Sect. F Struct Biol. Cryst. Commun. 2005, 61, 147–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clemetson, K.J.; Clemetson, J.M. Platelet GPIb complex as a target for anti-thrombotic drug development. Thromb. Haemost. 2008, 99, 473–479. [Google Scholar] [CrossRef]
- Yamazaki, Y.; Hyodo, F.; Morita, T. Wide distribution of cysteine-rich secretory proteins in snake venoms: Isolation and cloning of novel snake venom cysteine-rich secretory proteins. Arch. Biochem. Biophys. 2003, 412, 133–141. [Google Scholar] [CrossRef]
- Peng, M.; Lu, W.; Beviglia, L.; Niewiarowski, S.; Kirby, E.P. Echicetin: A snake venom protein that inhibits binding of von Willebrand factor and alboaggregins to platelet glycoprotein Ib. Blood 1993, 81, 2321–2328. [Google Scholar] [CrossRef]
- Fry, B.G.; Vidal, N.; Norman, J.A.; Vonk, F.J.; Scheib, H.; Ramjan, S.F.; Kuruppu, S.; Fung, K.; Hedges, S.B.; Richardson, M.K.; et al. Early evolution of the venom system in lizards and snakes. Nature 2006, 439, 584–588. [Google Scholar] [CrossRef]
- Reddy, T.; Gibbs, G.M.; Merriner, D.J.; Kerr, J.B.; O’Bryan, M.K. Cysteine-rich secretory proteins are not exclusively expressed in the male reproductive tract. Dev. Dyn. 2008, 237, 3313–3323. [Google Scholar] [CrossRef]
- Yamazaki, Y.; Koike, H.; Sugiyama, Y.; Motoyoshi, K.; Wada, T.; Hishinuma, S.; Mita, M.; Morita, T. Cloning and characterization of novel snake venom proteins that block smooth muscle contraction. Eur. J. Biochem. 2002, 269, 2708–2715. [Google Scholar] [CrossRef]
- Brown, R.L.; Haley, T.L.; West, K.A.; Crabb, J.W. Pseudechetoxin: A peptide blocker of cyclic nucleotide-gated ion channels. Proc. Natl. Acad. Sci. USA 1999, 96, 754–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stransky, S.; Costal-Oliveira, F.; Lopes-de-Souza, L.; Guerra-Duarte, C.; Chávez-Olórtegui, C.; Braga, V.M.M. In vitro assessment of cytotoxic activities of Lachesis muta muta snake venom. PLoS Negl. Trop. Dis. 2018, 12, e0006427. [Google Scholar] [CrossRef] [PubMed]
- Kerkkamp, H.; Bagowski, C.; Kool, J.; van Soolingen, B.; Vonk, F.J.; Vlecken, D. Whole snake venoms: Cytotoxic, anti-metastatic and antiangiogenic properties. Toxicon 2018, 150, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Lewinska, A.; Bocian, A.; Petrilla, V.; Adamczyk-Grochala, J.; Szymura, K.; Hendzel, W.; Kaleniuk, E.; Hus, K.; Petrillova, M.; Wnuk, M. Snake venoms promote stress-induced senescence in human fibroblasts. J. Cell. Physiol. 2019, 234, 6147–6160. [Google Scholar] [CrossRef] [PubMed]
- Debono, J.; Bos, M.H.A.; Frank, N.; Fry, B. Clinical implications of differential antivenom efficacy in neutralising coagulotoxicity produced by venoms from species within the arboreal viperid snake genus Trimeresurus. Toxicol. Lett. 2019, 316, 35–48. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Ontology | % of Protein Components | |
|---|---|---|
| T. macrops | T. hageni | |
| Molecular Function | ||
| binding (GO:0005488) | 12.5 | 33.3 |
| catalytic activity (GO:0003824) | 75.0 | 44.4 |
| structural molecule activity (GO:0005198) | 12.5 | - |
| molecular function regulator (GO:0098772) | - | 22.2 |
| Biological Process | ||
| biological regulation (GO:0065007) | 36.4 | 20.0 |
| cellular process (GO:0009987) | 27.3 | 26.7 |
| metabolic process (GO:0008152) | 9.1 | 6.7 |
| rhythmic process (GO:0048511) | 27.3 | 20.0 |
| immune system process (GO:0002376) | - | 6.7 |
| response to stimulus (GO:0050896) | - | 6.7 |
| localization (GO:0051179) | - | 13.3 |
| Cellular component | ||
| cell (GO:0005623) | 25.0 | 33.3 |
| extracellular region (GO:0005576) | 12.5 | 16.7 |
| membrane (GO:0016020) | 12.5 | - |
| organelle (GO:0043226) | 37.5 | 50.0 |
| protein-containing complex (GO:0032991) | 12.5 | - |
| No. | Accession No. | Protein | Family | Score | M.W. | No. of Peptide | % Sequence Coverage | pI | emPAI |
|---|---|---|---|---|---|---|---|---|---|
| 1 | gi|299493 | Thrombin-like proteinase | Protease | 99 | 2737 | 2 | 54.2 | 4.31 | 4.69 |
| 2 | gi|46395675 | Cysteine-rich venom protein tripurin | Toxin activity | 86 | 3971 | 2 | 33.3 | 9.19 | 2.76 |
| 3 | gi|13959432 | Acidic phospholipase A2 5 | Toxin activity | 442 | 13,870 | 4 | 27.9 | 4.72 | 2.74 |
| 4 | gi|67462321 | Disintegrin trigramin-gamma | Protease | 111 | 7568 | 2 | 28.8 | 4.61 | 2.17 |
| 5 | gi|697351484 | Chain A, crystal structure of an acidic Pla2 | Toxin activity | 443 | 13,710 | 4 | 36.4 | 4.81 | 2.03 |
| 6 | gi|60593434 | Chain A, crystal structure of stecrisp | Toxin activity | 238 | 25,112 | 4 | 18.1 | 5.53 | 1.12 |
| 7 | gi|20177994 | Acidic phospholipase A2 6 | Toxin activity | 146 | 15,712 | 3 | 21 | 4.72 | 0.79 |
| 10 | gi|538259813 | Galactose binding lectin, partial | CHO binding | 79 | 17,654 | 3 | 23 | 5.72 | 0.69 |
| 11 | gi|3552036 | Pallase | Protease | 151 | 26,162 | 3 | 19.4 | 5.89 | 0.62 |
| 12 | gi|344268956 | Phorbol-12-myristate-13-acetate-induced protein 1 | Apoptotic process | 36 | 6008 | 1 | 12.7 | 10.06 | 0.60 |
| 14 | gi|13959429 | Basic phospholipase A2 2 | Toxin activity | 130 | 7813 | 1 | 14.3 | 8.49 | 0.45 |
| 15 | gi|82094948 | Bradykinin-releasing enzyme KR-E-1 | Protease | 102 | 25,335 | 3 | 13.2 | 4.82 | 0.45 |
| 16 | gi|123894851 | Snake venom metalloproteinase | Protease | 364 | 68,673 | 7 | 12.2 | 6.4 | 0.39 |
| 17 | gi|13959619 | Snake venom serine protease 2B | Protease | 71 | 28,915 | 3 | 17.3 | 5.54 | 0.39 |
| 18 | gi|538259847 | 5’-nucleotidase | Nucleotidase activity | 203 | 57,055 | 7 | 15.1 | 8.27 | 0.32 |
| 19 | gi|32469800 | Thrombin-like enzyme contortrixobin | Protease | 102 | 25,396 | 3 | 13.2 | 4.95 | 0.28 |
| 20 | gi|143681919 | Thrombin-like enzyme kangshuanmei | Protease | 99 | 26,415 | 2 | 14.8 | 8.27 | 0.27 |
| No. | Accession No. | Protein | Family | Score | M.W. | No. of Peptides | %Sequence Coverage | pI | emPAI |
|---|---|---|---|---|---|---|---|---|---|
| 1 | gi|82130933 | Snaclec coagulation factor IX-binding protein subunit A | Toxin activity | 145 | 14,631 | 4 | 31.8 | 4.92 | 1.31 |
| 2 | gi|67462321 | Platelet aggregation activation inhibitor | Protease | 162 | 7568 | 2 | 28.8 | 4.61 | 1.16 |
| 3 | gi|60593434 | Chain A, crystal structure of stecrisp | Toxin activity | 174 | 25,112 | 5 | 21.7 | 5.53 | 1.12 |
| 4 | gi|3914268 | Acidic phospholipase A2 2 | Toxin activity | 155 | 13,784 | 4 | 38.5 | 4.95 | 0.94 |
| 5 | gi|538259791 | C-type lectin factor IX/X binding protein A subunit, | CHO binding | 122 | 14,688 | 3 | 30 | 4.92 | 0.87 |
| 6 | gi|32469800 | Thrombin-like enzyme contortrixobin | Protease | 450 | 25,396 | 6 | 18.4 | 4.95 | 0.85 |
| 7 | gi|39655009 | Chain A, crystal structure of a platelet agglutination factor isolated from the venom of Taiwan Habu | Protease | 227 | 15,718 | 5 | 26.7 | 5.09 | 0.79 |
| 8 | gi|126130 | Galactose-specific lectin | CHO binding | 55 | 16,281 | 4 | 31.1 | 5.54 | 0.76 |
| 9 | gi|565235162 | hypothetical protein L345_18387, partial | Unknown | 36 | 5242 | 1 | 37.5 | 3.63 | 0.7 |
| 10 | gi|13959617 | Snake venom serine protease 1 | Protease | 150 | 27,923 | 6 | 22.5 | 5.66 | 0.57 |
| 11 | gi|13959432 | Acidic phospholipase A2 5 | Toxin activity | 65 | 13,870 | 2 | 11.5 | 4.72 | 0.55 |
| 12 | gi|82201344 | Basic phospholipase A2 homolog Ts-R6 | Toxin activity | 56 | 15,477 | 5 | 24.8 | 8.5 | 0.49 |
| 13 | gi|129417 | Acidic phospholipase A2 1 | Toxin activity | 62 | 15,524 | 2 | 10.1 | 6.54 | 0.48 |
| 14 | gi|356581537 | Beta-defensin-like protein 10 | Chemokine receptor binding | 14 | 7492 | 1 | 9 | 9.73 | 0.48 |
| 15 | gi|59727030 | D1E6b phospholipase A2 | Toxin activity | 85 | 15,959 | 2 | 10.1 | 4.79 | 0.47 |
| 16 | gi|360797 | Flavoxobin | Protease | 158 | 25,725 | 5 | 13.6 | 5.2 | 0.44 |
| 17 | gi|538259853 | Phosphodiesterase | Magnesium binding | 128 | 96,526 | 15 | 17 | 8.03 | 0.44 |
| 18 | gi|676251620 | Neurofibromin, | Phosphatid-ylcholine binding | 30 | 7991 | 1 | 8.6 | 8.66 | 0.44 |
| 19 | gi|538259847 | 5ʹ-nucleotidase, | Nucleotidase activity | 177 | 57,055 | 10 | 19.5 | 8.27 | 0.4 |
| 20 | gi|632991866 | Transmembrane protein 184B-like | Unknown | 52 | 9150 | 1 | 13.1 | 5.65 | 0.37 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumkate, S.; Chanhome, L.; Thiangtrongjit, T.; Noiphrom, J.; Laoungboa, P.; Khow, O.; Vasaruchapong, T.; Sitprija, S.; Chaiyabutr, N.; Reamtong, O. Venomics and Cellular Toxicity of Thai Pit Vipers (Trimeresurus macrops and T. hageni). Toxins 2020, 12, 54. https://doi.org/10.3390/toxins12010054
Kumkate S, Chanhome L, Thiangtrongjit T, Noiphrom J, Laoungboa P, Khow O, Vasaruchapong T, Sitprija S, Chaiyabutr N, Reamtong O. Venomics and Cellular Toxicity of Thai Pit Vipers (Trimeresurus macrops and T. hageni). Toxins. 2020; 12(1):54. https://doi.org/10.3390/toxins12010054
Chicago/Turabian StyleKumkate, Supeecha, Lawan Chanhome, Tipparat Thiangtrongjit, Jureeporn Noiphrom, Panithi Laoungboa, Orawan Khow, Taksa Vasaruchapong, Siravit Sitprija, Narongsak Chaiyabutr, and Onrapak Reamtong. 2020. "Venomics and Cellular Toxicity of Thai Pit Vipers (Trimeresurus macrops and T. hageni)" Toxins 12, no. 1: 54. https://doi.org/10.3390/toxins12010054
APA StyleKumkate, S., Chanhome, L., Thiangtrongjit, T., Noiphrom, J., Laoungboa, P., Khow, O., Vasaruchapong, T., Sitprija, S., Chaiyabutr, N., & Reamtong, O. (2020). Venomics and Cellular Toxicity of Thai Pit Vipers (Trimeresurus macrops and T. hageni). Toxins, 12(1), 54. https://doi.org/10.3390/toxins12010054