Gas Chromatography–Mass Spectrometry Profiling of Volatile Compounds Reveals Metabolic Changes in a Non-Aflatoxigenic Aspergillus flavus Induced by 5-Azacytidine


Abstract
:1. Introduction
2. Results
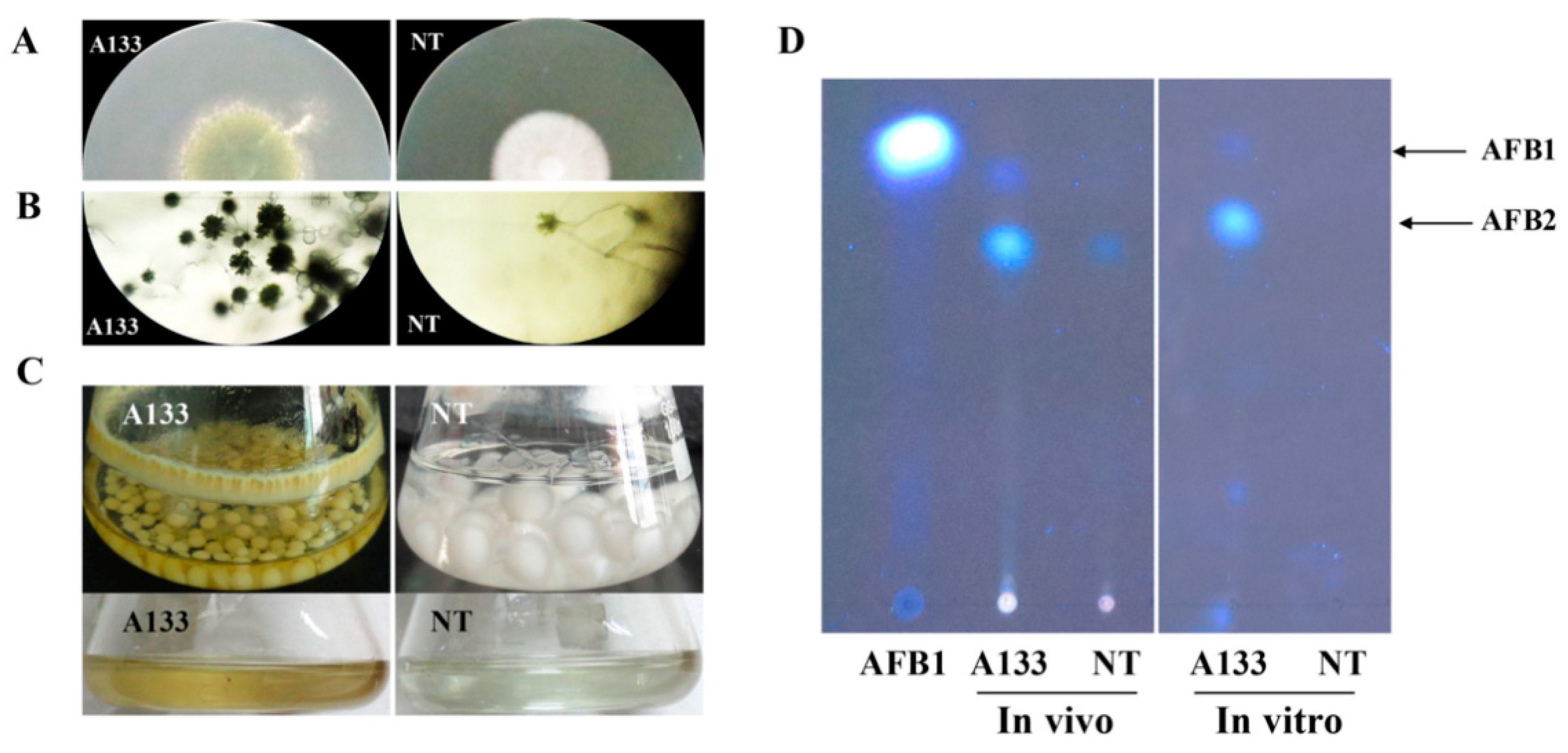
2.1. The NT Mutant Induced by 5-Azacytidine Is Impaired in Conidiation and Mycotoxins Production
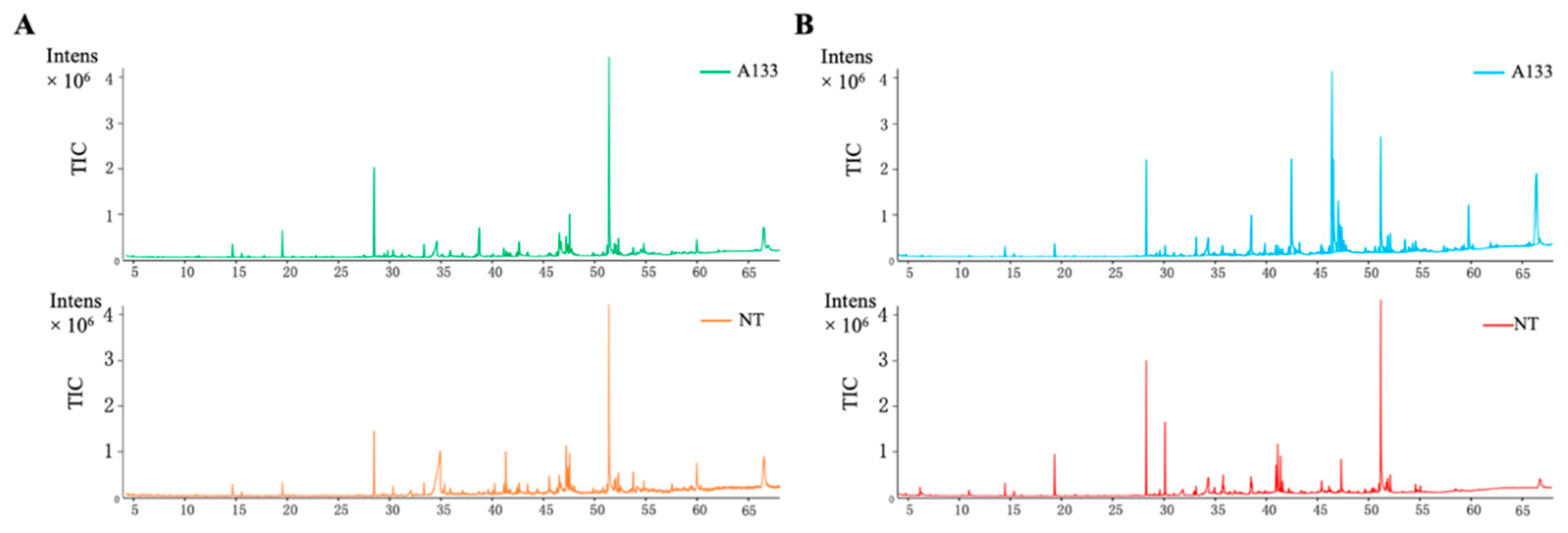
2.2. GC-MS Analysis of Volatiles Extracts
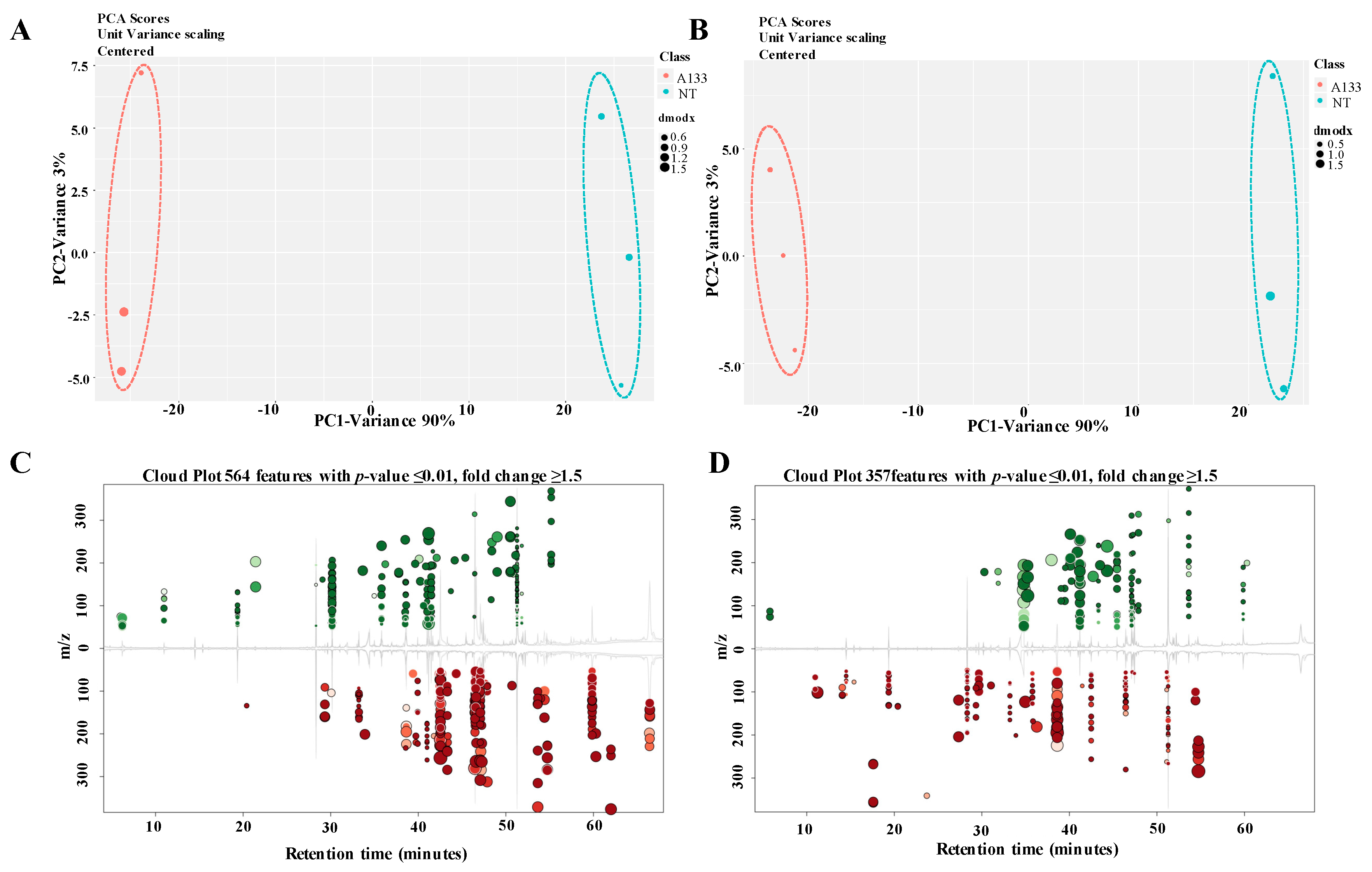
2.3. The A. flavus Non-Aflatoxigenic Mutant Produces Decreasing Volatile Compounds
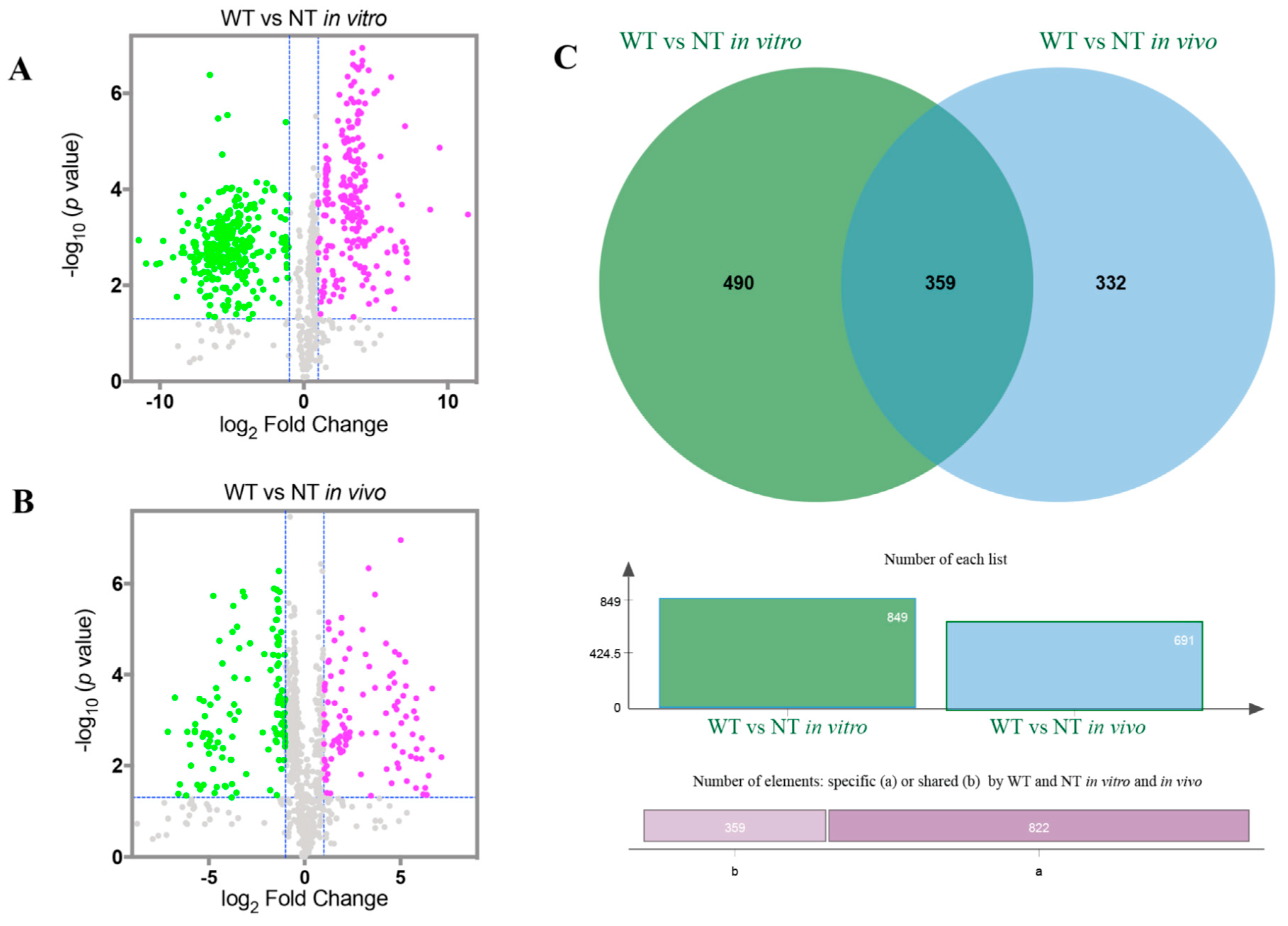
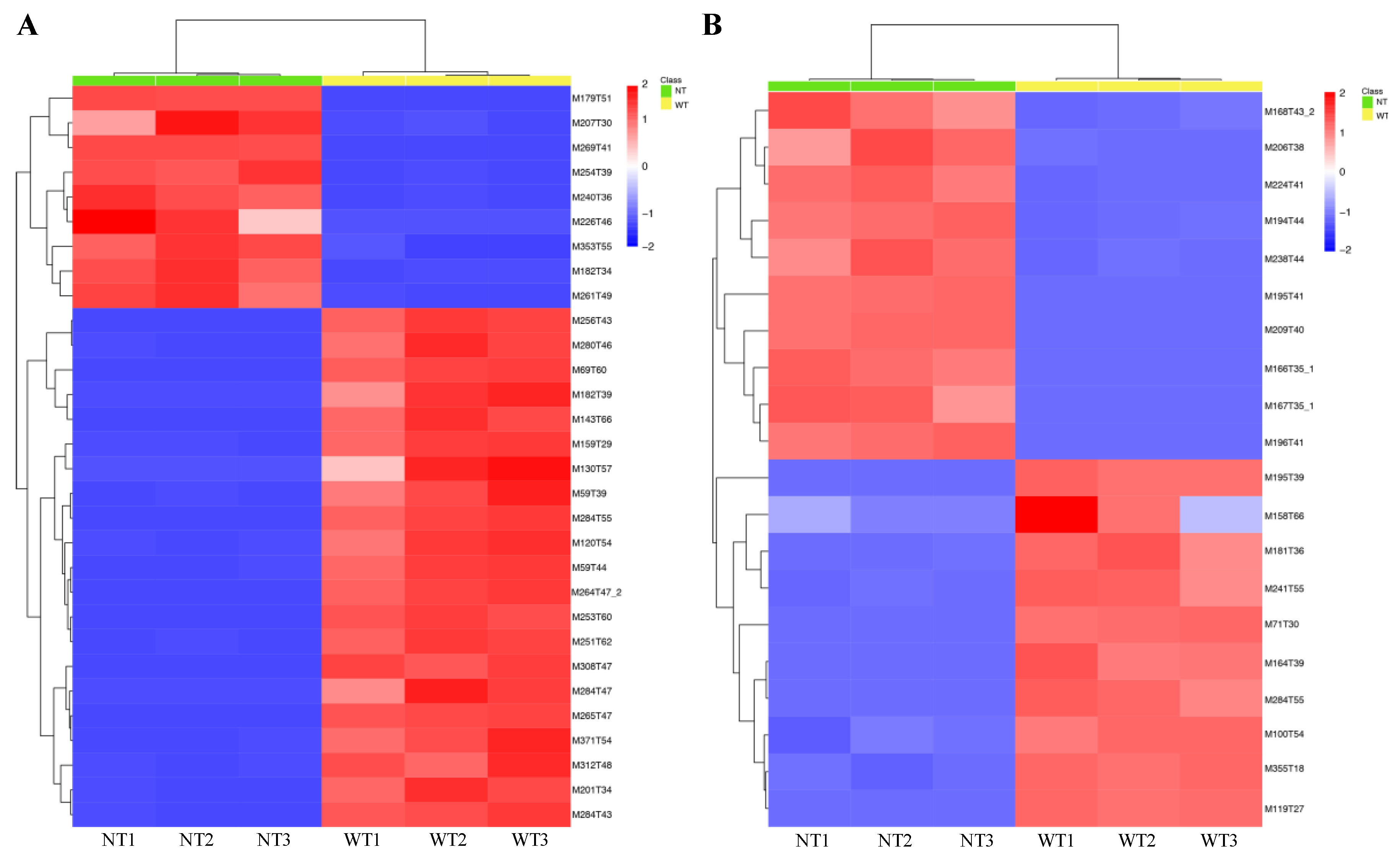
2.4. Screening of the Changed Volatile Compounds in NT Mutant
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Strains and Culture Conditions
5.2. Aflatoxins Analysis
5.3. GC-MS Volatile Analysis
5.4. Data Analysis
5.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yang, K.; Shadkchan, Y.; Tannous, J.; Landero Figueroa, J.A.; Wiemann, P.; Osherov, N.; Wang, S.; Keller, N.P. Contribution of atpase copper transporters in animal but not plant virulence of the crossover pathogen aspergillus flavus. Virulence 2018, 9, 1273–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakrabarti, A.; Singh, R. The emerging epidemiology of mould infections in developing countries. Curr. Opin. Infect. Dis. 2011, 24, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Liu, Y.; Wang, S.; Wu, L.; Xie, R.; Lan, H.; Fasoyin, O.E.; Wang, Y.; Wang, S. Cyclase-associated protein cap with multiple domains contributes to mycotoxin biosynthesis and fungal virulence in aspergillus flavus. J. Agric. Food Chem. 2019, 67, 4200–4213. [Google Scholar] [CrossRef]
- Rushing, B.R.; Selim, M.I. Aflatoxin b1: A review on metabolism, toxicity, occurrence in food, occupational exposure, and detoxification methods. Food Chem. Toxicol. 2019, 124, 81–100. [Google Scholar] [CrossRef] [PubMed]
- Hedayati, M.T.; Pasqualotto, A.C.; Warn, P.A.; Bowyer, P.; Denning, D.W. Aspergillus flavus: Human pathogen, allergen and mycotoxin producer. Microbiology 2007, 153, 1677–1692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruadrew, S.; Craft, J.; Aidoo, K. Occurrence of toxigenic aspergillus spp. And aflatoxins in selected food commodities of asian origin sourced in the west of scotland. Food Chem. Toxicol. 2013, 55, 653–658. [Google Scholar] [CrossRef]
- Tian, J.; Gan, Y.; Pan, C.; Zhang, M.; Wang, X.; Tang, X.; Peng, X. Nerol-induced apoptosis associated with the generation of ros and Ca2+ overload in saprotrophic fungus aspergillus flavus. Appl. Microbiol. Biotechnol. 2018, 102, 6659–6672. [Google Scholar] [CrossRef]
- Tian, J.; Wang, Y.; Lu, Z.; Sun, C.; Zhang, M.; Zhu, A.; Peng, X. Perillaldehyde, a promising antifungal agent used in food preservation, triggers apoptosis through a metacaspase-dependent pathway in aspergillus flavus. J. Agric. Food Chem. 2016, 64, 7404–7413. [Google Scholar] [CrossRef]
- Amaike, S.; Keller, N.P. Aspergillus flavus. Annu. Rev. Phytopathol. 2011, 49, 107–133. [Google Scholar] [CrossRef]
- Chalivendra, S.C.; DeRobertis, C.; Chang, P.K.; Damann, K.E. Cyclopiazonic acid is a pathogenicity factor for aspergillus flavus and a promising target for screening germplasm for ear rot resistance. Mol. Plant-Microbe Interact. MPMI 2017, 30, 361–373. [Google Scholar] [CrossRef] [Green Version]
- Chang, P.K.; Horn, B.W.; Dorner, J.W. Clustered genes involved in cyclopiazonic acid production are next to the aflatoxin biosynthesis gene cluster in aspergillus flavus. Fungal Genet. Biol. 2009, 46, 176–182. [Google Scholar] [CrossRef]
- Gilbert, M.K.; Mack, B.M.; Moore, G.G.; Downey, D.L.; Lebar, M.D.; Joardar, V.; Losada, L.; Yu, J.; Nierman, W.C.; Bhatnagar, D. Whole genome comparison of aspergillus flavus l-morphotype strain nrrl 3357 (type) and s-morphotype strain af70. PLoS ONE 2018, 13, e0199169. [Google Scholar] [CrossRef] [PubMed]
- Payne, G.A.; Nierman, W.C.; Wortman, J.R.; Pritchard, B.L.; Brown, D.; Dean, R.A.; Bhatnagar, D.; Cleveland, T.E.; Machida, M.; Yu, J. Whole genome comparison ofaspergillus flavusanda. Oryzae. Med. Mycol. 2006, 44, 9–11. [Google Scholar] [CrossRef] [PubMed]
- Amare, M.G.; Keller, N.P. Molecular mechanisms of aspergillus flavus secondary metabolism and development. Fungal Genet. Biol. 2014, 66, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Georgianna, D.R.; Fedorova, N.D.; Burroughs, J.L.; Dolezal, A.L.; Bok, J.W.; Horowitz-Brown, S.; Woloshuk, C.P.; Yu, J.; Keller, N.P.; Payne, G.A. Beyond aflatoxin: Four distinct expression patterns and functional roles associated with aspergillus flavus secondary metabolism gene clusters. Mol. Plant Pathol. 2010, 11, 213–226. [Google Scholar] [CrossRef] [PubMed]
- Khaldi, N.; Collemare, J.; Lebrun, M.H.; Wolfe, K.H. Evidence for horizontal transfer of a secondary metabolite gene cluster between fungi. Genome Biol. 2008, 9, R18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roze, L.V.; Chanda, A.; Laivenieks, M.; Beaudry, R.M.; Artymovich, K.A.; Koptina, A.V.; Awad, D.W.; Valeeva, D.; Jones, A.D.; Linz, J.E. Volatile profiling reveals intracellular metabolic changes in aspergillus parasiticus: Vea regulates branched chain amino acid and ethanol metabolism. BMC Biochem. 2010, 11, 33. [Google Scholar] [CrossRef] [Green Version]
- Osbourn, A. Secondary metabolic gene clusters: Evolutionary toolkits for chemical innovation. Trends Genet. 2010, 26, 449–457. [Google Scholar] [CrossRef]
- Pfannenstiel, B.T.; Keller, N.P. On top of biosynthetic gene clusters: How epigenetic machinery influences secondary metabolism in fungi. Biotechnol. Adv. 2019, 37, 107345. [Google Scholar] [CrossRef]
- Sun, J.; Awakawa, T.; Noguchi, H.; Abe, I. Induced production of mycotoxins in an endophytic fungus from the medicinal plant Datura stramonium L. Bioorganic Med. Chem. Lett. 2012, 22, 6397–6400. [Google Scholar] [CrossRef] [PubMed]
- Shwab, E.K.; Bok, J.W.; Tribus, M.; Galehr, J.; Graessle, S.; Keller, N.P. Histone deacetylase activity regulates chemical diversity in aspergillus. Eukaryot. Cell 2007, 6, 1656–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Sun, Y.; Yue, S.; Wang, Y.; Lu, F. Histone deacetylase inhibitors in cancer therapy. Curr. Top. Med. Chem. 2018, 18, 2420–2428. [Google Scholar] [CrossRef] [PubMed]
- Shigemoto, R.; Matsumoto, T.; Masuo, S.; Takaya, N. 5-methylmellein is a novel inhibitor of fungal sirtuin and modulates fungal secondary metabolite production. J. Gen. Appl. Microbiol. 2018, 64, 240–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savickiene, J.; Treigyte, G.; Jazdauskaite, A.; Borutinskaite, V.V.; Navakauskiene, R. DNA methyltransferase inhibitor rg108 and histone deacetylase inhibitors cooperate to enhance nb4 cell differentiation and e-cadherin re-expression by chromatin remodelling. Cell Biol. Int. 2012, 36, 1067–1078. [Google Scholar] [CrossRef]
- Yang, K.; Zhuang, Z.; Zhang, F.; Song, F.; Zhong, H.; Ran, F.; Yu, S.; Xu, G.; Lan, F.; Wang, S. Inhibition of aflatoxin metabolism and growth of aspergillus flavus in liquid culture by a DNA methylation inhibitor. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2015, 32, 554–563. [Google Scholar] [CrossRef]
- Carvajal-Moreno, M.; Vargas-Ortiz, M.; Hernandez-Camarillo, E.; Ruiz-Velasco, S.; Rojo-Callejas, F. Presence of unreported carcinogens, aflatoxins and their hydroxylated metabolites, in industrialized oaxaca cheese from mexico city. Food Chem. Toxicol. 2019, 124, 128–138. [Google Scholar] [CrossRef]
- Qu, S.; Yang, K.; Chen, L.; Liu, M.; Geng, Q.; He, X.; Li, Y.; Liu, Y.; Tian, J. Cinnamaldehyde, a promising natural preservative against aspergillus flavus. Front. Microbiol. 2019, 10, 2895. [Google Scholar] [CrossRef] [Green Version]
- Yang, K.; Liang, L.; Ran, F.; Liu, Y.; Li, Z.; Lan, H.; Gao, P.; Zhuang, Z.; Zhang, F.; Nie, X.; et al. The dmta methyltransferase contributes to aspergillus flavus conidiation, sclerotial production, aflatoxin biosynthesis and virulence. Sci. Rep. 2016, 6, 23259. [Google Scholar] [CrossRef] [Green Version]
- Chang, P.K.; Scharfenstein, L.L.; Li, P.; Ehrlich, K.C. Aspergillus flavus velb acts distinctly from vea in conidiation and may coordinate with flug to modulate sclerotial production. Fungal Genet. Biol. 2013, 58–59, 71–79. [Google Scholar] [CrossRef]
- Han, X.; Qiu, M.; Wang, B.; Yin, W.B.; Nie, X.; Qin, Q.; Ren, S.; Yang, K.; Zhang, F.; Zhuang, Z.; et al. Functional analysis of the nitrogen metabolite repression regulator gene nmra in aspergillus flavus. Front. Microbiol. 2016, 7, 1794. [Google Scholar] [CrossRef] [Green Version]
- Yabe, K.; Nakajima, H. Enzyme reactions and genes in aflatoxin biosynthesis. Appl. Microbiol. Biotechnol. 2004, 64, 745–755. [Google Scholar] [CrossRef] [PubMed]
- Tsitsigiannis, D.I.; Keller, N.P. Oxylipins act as determinants of natural product biosynthesis and seed colonization in aspergillus nidulans. Mol. Microbiol. 2006, 59, 882–892. [Google Scholar] [CrossRef] [PubMed]
- Reverberi, M.; Ricelli, A.; Zjalic, S.; Fabbri, A.A.; Fanelli, C. Natural functions of mycotoxins and control of their biosynthesis in fungi. Appl. Microbiol. Biotechnol. 2010, 87, 899–911. [Google Scholar] [CrossRef] [PubMed]
- Narasaiah, K.V.; Sashidhar, R.B.; Subramanyam, C. Biochemical analysis of oxidative stress in the production of aflatoxin and its precursor intermediates. Mycopathologia 2006, 162, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Jahn, B.; Boukhallouk, F.; Lotz, J.; Langfelder, K.; Wanner, G.; Brakhage, A.A. Interaction of human phagocytes with pigmentless aspergillus conidia. Infect. Immun. 2000, 68, 3736–3739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gowda, H.; Ivanisevic, J.; Johnson, C.H.; Kurczy, M.E.; Benton, H.P.; Rinehart, D.; Nguyen, T.; Ray, J.; Kuehl, J.; Arevalo, B.; et al. Interactive xcms online: Simplifying advanced metabolomic data processing and subsequent statistical analyses. Anal. Chem. 2014, 86, 6931–6939. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Volatile Compound | Retention Time (min) | Quantification Ion (m/z) | Intensity | log2fold | |
|---|---|---|---|---|---|---|
| A133 | NT | |||||
| Fatty acid-derived volatiles | ||||||
| M256T43 | n-Hexadecanoic acid | 42.53 | 256 | 173,803 ± 5899 | 42 ± 22 | −11.48 |
| M284T43 | Hexadecanoicacid,ethylester | 43.31 | 284 | 10,182 ± 273 | 162 ± 57 | −5.97 |
| M59T44 | Normeperidinic acid | 44.33 | 205 | 47,057 ± 1980 | 711 ± 114 | −6.05 |
| M280T46 | 9,12-Octadecadienoicacid(Z,Z)- | 46.47 | 280 | 168,927 ± 10,057 | 48 ± 48 | −10.95 |
| M264T47_2 | 6-Octadecenoic acid | 46.60 | 264 | 49,278 ± 1697 | 0 ± 0 | −9.77 |
| M308T47 | 9,12-Octadecadienoicacid,ethylester | 47.08 | 308 | 14,919 ± 336 | 0 ± 0 | −8.05 |
| M284T47 | n-Propyl9-octadecenoate | 47.14 | 284 | 18,212 ± 1621 | 0 ± 0 | −8.34 |
| M312T48 | Heptadecanoicacid,15-methyl-,ethylester | 47.87 | 312 | 6804 ± 330 | 30 ± 15 | −7.12 |
| M253T60 | 6,9-hexadecadienoic acid | 60.28 | 253 | 42,150 ± 672 | 291 ± 50 | −7.18 |
| M143T66 | Ergosterol | 66.43 | 143 | 407,309 ± 17,088 | 3840 ± 72 | −6.73 |
| Phenylpropanoid derivatives/benzenoids | ||||||
| M159T29 | N-Methyl-N-methoxy-5,6,7,8-tetrahydro-1-naphtamide | 29.33 | 159 | 73,675 ± 2920 | 1314 ± 197 | −5.81 |
| M207T30 | Phenol,2,4-bis(1,1-dimethylethyl)- | 30.16 | 207 | 636 ± 372 | 37,011 ± 4354 | 5.82 |
| M201T34 | 4′-Azidobenzo[1′,2′-b]-1,4-diazabicyclo[2.2.2]octene | 33.91 | 201 | 10,246 ± 446 | 64 ± 64 | −6.66 |
| M254T39 | Phenol,3,5-dimethoxy- | 38.51 | 254 | 85 ± 15 | 3599 ± 101 | 5.4 |
| M182T39 | Flopropione | 38.63 | 182 | 340,108 ± 32,366 | 2423 ± 111 | −7.13 |
| M59T39 | Phthalicacid,isobutylnonylester | 39.40 | 59 | 32,073 ± 2209 | 818 ± 191 | −5.29 |
| M261T49 | Tiaprofenic acid | 49.01 | 260 | 37 ± 37 | 10,811 ± 627 | 7.18 |
| M371T54 | 2,4-Difluorophenol | 53.64 | 371 | 18,882 ± 1098 | 60 ± 60 | -7.6 |
| M353T55 | Phenol,4,4′-methylenebis[2,6-bis(1,1-dimethylethyl)]- | 55.18 | 424 | 1710 ± 250 | 16,636 ± 581 | 3.28 |
| M130T57 | 1H-Indole,4-methyl- | 57.45 | 130 | 87,806 ± 14,251 | 1825 ± 109 | −5.59 |
| M251T62 | Propiophenone,2′-(trimethylsiloxy)- | 62.00 | 222 | 40,280 ± 1350 | 1112 ± 147 | −5.18 |
| Triterpene | ||||||
| M69T60 | 2,6,10,14,18,22-Tetracosahexaene,2,6,10,15,19,23-hexamethyl-,(all-E)- | 59.85 | 410 | 694,833 ± 19,821 | 8465 ± 313 | −6.36 |
| Amino acids derived volatiles | ||||||
| M182T34 | L-Tyrosine | 33.66 | 182 | 68 ± 34 | 7028 ± 315 | 6.34 |
| M240T36 | 6-Methyl-3,5-heptadien-2-one | 35.81 | 124 | 15 ± 15 | 4429 ± 200 | 6.4 |
| M269T41 | 1-Hexadecanamine,N,N-dimethyl- | 41.18 | 269 | 0 ± 0 | 39,265 ± 145 | 9.44 |
| M226T46 | N-Methyl-beta-carboline-3-carboxamide | 46.1 | 225 | 0 ± 0 | 4410 ± 794 | 6.29 |
| M265T47 | 9-Octadecenamide,(Z)- | 47.25 | 265 | 18,661 ± 213 | 0 ± 0 | −8.37 |
| M179T51 | 2-phenyl-N-(5-propan-2-yl-1,3-thiazol-2-yl)acetamide | 50.52 | 260 | 1332 ± 173 | 89,236 ± 370 | 6.07 |
| M120T54 | 4-Morpholineethanol | 54.40 | 131 | 57,950 ± 3565 | 562 ± 108 | −6.69 |
| M284T55 | 1-(2-Thiazolylazo)-2-naphthol | 54.74 | 255 | 33,915 ± 1238 | 170 ± 25 | −7.64 |
| Name | Volatile Compound | Retention Time (min) | Quantification Ion (m/z) | Intensity | log2fold | |
|---|---|---|---|---|---|---|
| A133 | NT | |||||
| Fatty acid-derived volatiles | ||||||
| M119T27 | Trimethylammonioacetate | 27.35 | 117.15 | 32,031 ± 246 | 1471 ± 71 | −4.44 |
| M100T54 | Hexadecanoicacid,2-hydroxy-1-(hydroxymethyl)ethylester | 54.40 | 330.5 | 36,178 ± 753 | 3406 ± 1018 | −3.41 |
| M284T55 | 1,2-Benzenedicarboxylicacid,mono(2-ethylhexyl)ester | 54.74 | 54.74 | 53,296 ± 2260 | 378 ± 61 | −7.14 |
| M158T66 | Ergosterol | 66.44 | 143 | 16,097 ± 6460 | 897 ± 897 | −3.47 |
| Phenylpropanoid derivatives/benzenoids | ||||||
| M355T18 | 2,6-Dichloroindophenol | 17.59 | 268.1 | 33,856 ± 310 | 2579 ± 469 | −3.71 |
| M167T35_1 | Benzo[b]tetrahydrofuran-3-one,5,6-dihydroxy- | 34.79 | 166.027 | 1110 ± 35 | 111,478 ± 7417 | 6.65 |
| M166T35_1 | 2,6-toluenediamine | 35.23 | 122.17 | 4043 ± 615 | 211,094 ± 5606 | 5.71 |
| M209T40 | Isoelemicin | 40.11 | 208.25 | 13,703 ± 254 | 353,432 ± 4258 | 4.69 |
| Saccharide derived volatiles | ||||||
| M195T41 | D-Glucoside | 41.2 | 194.18 | 34,031 ± 185 | 1,301,860 ± 9210 | 5.26 |
| Amino acids derived volatiles | ||||||
| M71T30 | Caulophylline | 29.66 | 204.27 | 112,547 ± 868 | 12,794 ± 455 | −3.14 |
| M181T36 | Theobromine | 36.27 | 180.16 | 11,780 ± 637 | 382 ± 66 | −4.95 |
| M206T38 | N-Acetyl-D-quinovosamine | 37.95 | 205.21 | 170 ± 64 | 7878 ± 634 | 5.54 |
| M195T39 | N,N′-Diacetyl-2-nitro-p-phenylenediamine | 38.6 | 237.21 | 574,864 ± 10,207 | 5272 ± 227 | −6.77 |
| M164T39 | 2,2′-(2-Hydroxy-2-nitrosohydrazinylidene)bis-ethanamine | 38.61 | 163.17 | 54,132 ± 2206 | 1027 ± 49 | −5.72 |
| M224T41 | 4-(Methylnitrosamino)-1-(1-oxido-3-pyridinyl)-1-butanone | 40.90 | 223.23 | 293 ± 38 | 7571 ± 196 | 4.69 |
| M196T41 | 9-Aminoacridine | 41.19 | 194.23 | 4102 ± 160 | 145,064 ± 2468 | 5.14 |
| M168T43_2 | 2,8-Dihydroxyadenine | 42.71 | 167.12 | 455 ± 174 | 11,735 ± 805 | 4.69 |
| M194T44 | Phenylacetylglycine | 43.50 | 194.1 | 1457 ± 103 | 18,559 ± 371 | 3.67 |
| M238T44 | N-Benzylphthalimide | 44.31 | 237.25 | 469 ± 201 | 32,530 ± 1702 | 6.12 |
| M241T55 | Tetramisole hydrochloride | 54.74 | 240.75 | 9247 ± 476 | 243 ± 101 | −5.25 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, F.; Geng, Q.; Wang, X.; Gao, X.; He, X.; Zhao, W.; Lan, H.; Tian, J.; Yang, K.; Wang, S. Gas Chromatography–Mass Spectrometry Profiling of Volatile Compounds Reveals Metabolic Changes in a Non-Aflatoxigenic Aspergillus flavus Induced by 5-Azacytidine. Toxins 2020, 12, 57. https://doi.org/10.3390/toxins12010057
Song F, Geng Q, Wang X, Gao X, He X, Zhao W, Lan H, Tian J, Yang K, Wang S. Gas Chromatography–Mass Spectrometry Profiling of Volatile Compounds Reveals Metabolic Changes in a Non-Aflatoxigenic Aspergillus flavus Induced by 5-Azacytidine. Toxins. 2020; 12(1):57. https://doi.org/10.3390/toxins12010057
Chicago/Turabian StyleSong, Fengqin, Qingru Geng, Xuewei Wang, Xiaoqing Gao, Xiaona He, Wei Zhao, Huahui Lan, Jun Tian, Kunlong Yang, and Shihua Wang. 2020. "Gas Chromatography–Mass Spectrometry Profiling of Volatile Compounds Reveals Metabolic Changes in a Non-Aflatoxigenic Aspergillus flavus Induced by 5-Azacytidine" Toxins 12, no. 1: 57. https://doi.org/10.3390/toxins12010057
APA StyleSong, F., Geng, Q., Wang, X., Gao, X., He, X., Zhao, W., Lan, H., Tian, J., Yang, K., & Wang, S. (2020). Gas Chromatography–Mass Spectrometry Profiling of Volatile Compounds Reveals Metabolic Changes in a Non-Aflatoxigenic Aspergillus flavus Induced by 5-Azacytidine. Toxins, 12(1), 57. https://doi.org/10.3390/toxins12010057