Co-Occurrence of Cyanobacteria and Cyanotoxins with Other Environmental Health Hazards: Impacts and Implications

Abstract
:1. Introduction
2. Environmental Intoxications Involving Toxigenic Cyanobacteria and Additional Agents
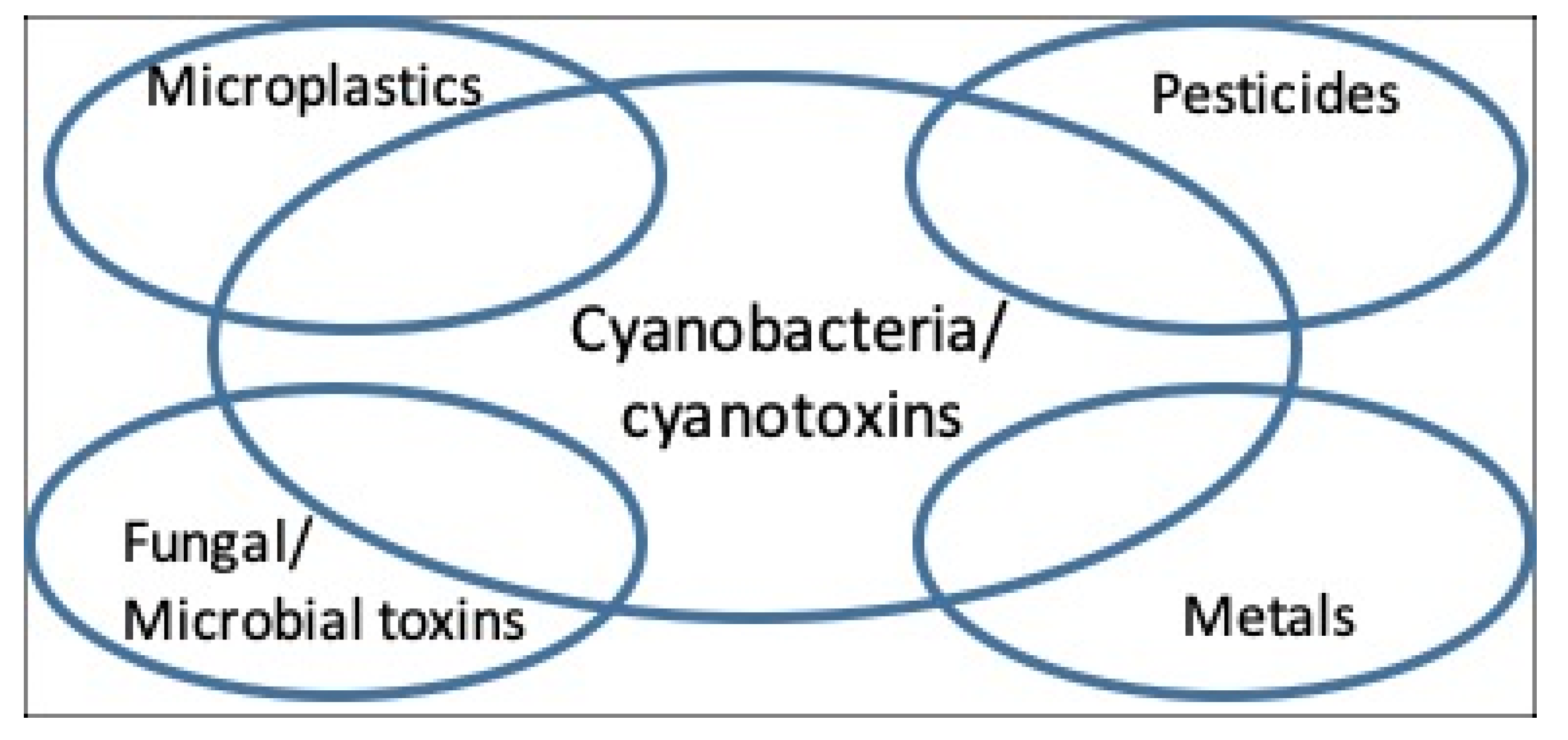
3. Additional Health Hazards: Their Co-Occurrence and Interactions with Cyanobacteria and Cyanotoxins
3.1. Fungal and Algal Toxins
3.2. Microbial Pathogens
3.3. Metals
3.4. Pesticides
3.5. Microplastic and Nanoplastic Particles and Contaminants
4. Implications for Water Safety Guidelines, Legislation, and Water Treatment
5. Concluding Remarks
Funding
Conflicts of Interest
References
- Carmichael, W.W. (Ed.) The Water Environment: Algal Toxins and Health; Plenum Press: New York, NY, USA, 1981; p. 491. [Google Scholar]
- Falconer, I.R. (Ed.) Algal Toxins in Seafood and Drinking Water; Academic Press: London, UK, 1993; p. 224. [Google Scholar]
- Codd, G.A.; Jefferies, T.M.; Keevil, C.W.; Potter, E. (Eds.) Detection Methods for Cyanobacterial Toxins; Royal Society of Chemistry: Cambridge, UK, 1994; p. 191. [Google Scholar]
- Huisman, J.; Matthijs, H.C.P.; Visser, P.M. (Eds.) Harmful Cyanobacteria; Springer: Dordrecht, The Netherlands, 2005; p. 241. [Google Scholar]
- Meriluoto, J.; Spoof, L.; Codd, G.A. (Eds.) Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Wiley: Chichester, UK, 2017; p. 548. [Google Scholar]
- Metcalf, J.S.; Codd, G.A. Cyanotoxins. In Ecology of Cyanobacteria II: Their Diversity in Space and Time; Whitton, B.A., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 651–675. [Google Scholar]
- Testai, E.; Scardala, S.; Vichi, S.; Buratti, F.M.; Funari, E. Risk to human health associated with the environmental occurrence of cyanobacterial neurotoxic alkaloids anatoxins and saxitoxins. Crit. Rev. Toxicol. 2016, 46, 385–418. [Google Scholar] [CrossRef] [PubMed]
- Svircĕv, Z.; Drobac, D.; Tokodi, N.; Mijović, B.; Codd, G.A.; Meriluoto, J. Toxicology of microcystins with reference to cases of human intoxications and epidemiological investigations of exposure to cyanobacteria and cyanotoxins. Arch. Toxicol. 2017, 91, 621–650. [Google Scholar] [CrossRef] [PubMed]
- Buratti, F.M.; Manganelli, M.; Vichi, S.; Stefanelli, M.; Scardala, S.; Testai, E.; Funari, E. Cyanotoxins: Producing organisms, occurrence, toxicity, mechanism of action and human health toxicological risk evaluation. Arch. Toxicol. 2017, 91, 1049–1130. [Google Scholar] [PubMed]
- Pichardo, S.; Caméan, A.M.; Jos, A. In vitro toxicological assessment of cylindrospermopsin: A review. Toxins 2017, 16, 402. [Google Scholar] [CrossRef] [Green Version]
- Metcalf, J.S.; Souza, N.R. Cyanobacteria and their toxins. Sep. Sci. Technol. 2019, 11, 125–148. [Google Scholar]
- Svircĕv, Z.; Lalić, D.; Savić, G.B.; Tokodi, N.; Backovic, D.D.; Chen, L.; Meriluoto, J.; Codd, G.A. Global geographical and historical overview of cyanotoxins distribution and cyanobacterial poisonings. Arch. Toxicol. 2019, 93, 2429–2481. [Google Scholar] [CrossRef]
- Francis, G. Poisonous Australian lake. Nature 1878, 18, 11–12. [Google Scholar] [CrossRef] [Green Version]
- Codd, G.A.; Morton, H.; Baker, P.D. George Francis, a pioneer in the investigation of the quality of South Australia’s drinking water sources (1878–1883). Trans. R. Soc. S. Aust. 2015, 139, 164–170. [Google Scholar] [CrossRef]
- Blaha, L.; Camean, A.M.; Fessard, V.; Gutierrez-Praena, D.; Jos, A.; Marie, B.; Metcalf, J.S.; Pichardo, S.; Puerto, M.; Torokne, A.; et al. Bioassay use in the field of cyanobacteria. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Wiley: Chichester, UK, 2017; pp. 272–279. [Google Scholar]
- Meriluoto, J.; Metcalf, J.S.; Codd, G.A. Selection of analytical methodology for cyanotoxin analysis. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Wiley: Chichester, UK, 2017; pp. 308–312. [Google Scholar]
- Kurmayer, R.; Sivonen, K.; Wilmotte, A.; Salmaso, N. (Eds.) Molecular Tools for the Detection and Quantitation of Toxigenic Cyanobacteria; John Wiley and Sons: Chichester, UK, 2017; p. 402. [Google Scholar]
- Turner, A.D.; Waack, J.; Lewis, A.; Edwards, C.; Lawton, L. Development and single-laboratory validation of a UHPLC-MS/MS method for quantitation of microcystins and nodularin in natural water, cyanobacteria, shellfish and algal supplement tablet powders. J. Chromatogr. B 2018, 1074–1075, 111–123. [Google Scholar] [CrossRef]
- Gimenéz-Campillo, C.; Pastor-Belda, M.; Campillo, N.; Arroyo-Manzanares, N.; Hernández-Cόrdoba, M.; Viñas, P. Determination of cyanotoxins and phycotoxins in seawater and algae-based supplements using ionic liquids and liquid chromatography with time-of-flight mass spectrometry. Toxins 2019, 11, 610. [Google Scholar] [CrossRef] [Green Version]
- Roy-Lachapelle, A.; Duy, S.V.; Munoz, G.; Dinh, Q.T.; Bahl, E.; Simon, D.F.; Sauvé, S. Analysis of multiclass cyanotoxins (microcystins, anabaenopeptins, cylindrospermopsin and anatoxins) in lake water using on-line SPE liquid chromatography high-resolution Orbitrap mass spectrometry. Anal. Methods 2019, 11, 5289. [Google Scholar] [CrossRef] [Green Version]
- Di Pofi, G.; Favero, G.; di Gregorio, F.N.; Ferretti, E.; Viaggiu, E.; Lucentini, L. Multi-residue ultra performance liquid chromataography-high resolution mass spectrometry method for the analysis of 21 cyanotoxins in surface water for human consumption. Talanta 2020, 211, 120738. [Google Scholar] [CrossRef] [PubMed]
- Canter-Lund, H.; Lund, J.W.G. Freshwater Algae: Their Microscopic World Explored; Biopress Limited: Bristol, UK, 1995; p. 360. [Google Scholar]
- Whitton, B.A. (Ed.) Ecology of Cyanobacteria II: Their Diversity in Space and Time; Springer: Dordrecht, The Netherlands, 2012; p. 760. [Google Scholar]
- Islam, M.S.; Miah, M.A.; Hasan, M.K.; Sack, R.B.; Albert, M.J. Detection of non-culturable Vibrio cholera 01 associated with a cyanobacterium from an aquatic environment in Bangladesh. Trans. R. Soc. Trop. Med. Hyg. 1994, 88, 298–299. [Google Scholar] [CrossRef]
- Timbrell, J.A. Introduction to Toxicology; Taylor & Francis: Bristol, UK, 1995; p. 167. [Google Scholar]
- Fitzgeorge, R.B.; Clark, S.A.; Keevil, C.W. Routes of intoxication. In Detection Methods for Cyanobacterial Toxins; Royal Society of Chemistry: Cambridge, UK, 1994; pp. 69–74. [Google Scholar]
- Chia, M.A.; Kramer, B.J.; Jankowiak, J.G.; do Carmo Bittencourt-Oliveira, M.; Gobler, C.J. The individual and combined effects of the cyanotoxins, anatoxin-a and microcystin-LR, on the growth, toxin production and nitrogen fixation of prokaryotic and eukaryotic algae. Toxins 2019, 11, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rymuszka, A.; Sieroslawska, A. Cytotoxic and immunotoxic effects of the mixture containing cyanotoxins on carp cells following in vitro exposure. Cent. Eur. J. Immunol. 2013, 38, 159–163. [Google Scholar] [CrossRef]
- Li, Q.; Gu, P.; Zhang, C.; Luo, X.; Zhang, H.; Zhang, J.; Zheng, Z. Combined toxic effects of anatoxin-a and microcystin-LR on submerged macrophytes and biofilms. J. Hazard. Mater. 2020, 389, 122053. [Google Scholar] [CrossRef]
- Lindsay, J.; Metcalf, J.S.; Codd, G.A. Protection against the toxicity of microcystin-LR and cylindrospermopsin in Artemia salina, and Daphnia spp. by pretreatment with cyanobacterial lipopolysaccharide (LPS). Toxicon 2006, 48, 995–1001. [Google Scholar] [CrossRef]
- Pinheiro, C.; Azevedo, J.; Campos, A.; Vasconcelos, V.; Loureiro, S. The interactive effects of microcystin-LR and cylindrospermopsin on the growth rate of the freshwater alga Chlorella Vulgaris. Ecotoxicology 2016, 25, 745–758. [Google Scholar] [CrossRef] [Green Version]
- Reinikainen, M.; Meriluoto, J.A.O.; Spoof, L.; Harada, K.-I. The toxicities of a polyunsaturated fatty acid and a microcystin to Daphnia Magna. Environ. Toxicol. 2001, 16, 444–448. [Google Scholar] [CrossRef]
- Wang, L.; He, L.; Zeng, H.; Fu, W.; Wang, J.; Tan, Y.; Zheng, C.; Qiu, Z.; Luo, J.; Lv, C.; et al. Low dose microcystin-LR antagonises aflatoxin B1 induced hepatocarcinogenesis through decreased cytochrome P450 1A2 expression and aflatoxin B1-DNA adduct generation. Chemosphere 2020, 248, 126036. [Google Scholar] [CrossRef]
- Meneely, J.P.; Hašlová, J.; Krska, R.; Elliott, C.T. Assessing the combined toxicity of the natural toxins, aflatoxin B1, fumonisin B1 and microcystin-LR by high content analysis. Food Chem. Toxicol. 2018, 121, 527–540. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Wang, S.; Xu, E.G.; Liu, J.; Li, X.; Wang, Z. Synergistic toxicity of microcystin-LR and Cu to zebrafish (Danio rerio). Sci. Total Environ. 2020, 713, 736393. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, J.; Li, E.; Zhang, L.; Wang, X.; Song, L. Combined toxic effects and mechanisms of microcystin-LR and copper on Vallisneria natans (Lous.) Hara seedlings. J. Hazard. Mater. 2017, 328, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Xiao, B.; Song, L.; Wu, X.; Zhang, Y.; Wang, C. Effects of microcystin-LR, linear alkylbenzene sulfonate and their mixture on lettuce (Lactuca sativa L.) seeds and seedlings. Ecotoxicology 2011, 20, 803–814. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.; Steinman, A.D.; Shu, X.; Cao, Q.; Yao, L.; Xie, L. Combined toxic effects of microcystin-LR and phenanthrene on growth and antioxidant system of duckweed (Lemna gibba L.). Ecotoxicol. Environ. Saf. 2019, 185, 109668. [Google Scholar] [CrossRef] [PubMed]
- Hinojosa, M.G.; Prieto, A.I.; Gutiérrez-Praena, D.; Moreno, F.J.; Caméan, A.M.; Jos, A. In vitro assessment of the combination of cylindrospermopsin and the organophosphate chlorpyrifos on the human neuroblastoma SH-SY5Y cell line. Ecotoxicol. Environ. Saf. 2020, 191, 110222. [Google Scholar] [CrossRef]
- Hercog, C.; Štern, A.; Maisanaba, S.; Filipič, M.; Bojana, Ž. Plastics in cyanobacterial blooms—genotoxic effects of binary mixtures of cylindrospermopsin and bisphenols in HepG2 cells. Toxins 2020, 12, 219. [Google Scholar] [CrossRef] [Green Version]
- Fiore, S.M.F.; de Lima, S.T.; Carmichael, W.W.; McKinnie, S.M.K.; Checkan, J.R.; Moore, B.S. Guanitoxin, renaming a cyanobacterial organophosphate toxin. Harmful Algae 2020, 92, 101937. [Google Scholar] [CrossRef]
- Spoof, L.; Catherine, A. Appendix 3: Tables of microcystins and nodularins. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Wiley: Chichester, UK, 2017; pp. 526–537. [Google Scholar]
- Robillot, C.; Vinh, J.E.; Puiseux-Dao, S.; Hennion, M.-C. Hepatotoxin production kinetics of the cyanobacterium Microcystis aeruginosa PCC 7820, as determined by HPLC-mass spectrometry and protein phosphatase bioassay. Environ. Sci. Technol. 2000, 34, 3372–3378. [Google Scholar] [CrossRef]
- Wimmer, K.M.; Strangman, W.; Wright, J.L.C. 7-deoxy-desulfo-cylindrospermopsin and 7-dexoy-desulfo-12-acetylcylindrospermopsin: Two new cylindrospermopsin analogs isolated from a Thai strain of Cylindrospermopsis raciborskii. Harmful Algae 2014, 37, 203–206. [Google Scholar] [CrossRef]
- Ward, C.J.; Codd, G.A. Comparative toxicity of four microcystins of different hydrophobicities to the protozoan, Tetrahymena Pyriformis. J. Appl. Microbiol. 1999, 86, 874–882. [Google Scholar] [CrossRef]
- Harada, K.-I.; Ogawa, K.; Kimura, Y.; Murata, H.; Suzuki, M. Microcystins from Anabaena flos-aquae NRC 525-17. Chem. Res. Toxicol. 1991, 4, 535–540. [Google Scholar] [CrossRef]
- Monteiro, S.; Santos, R.; Bláha, L.; Codd, G.A. Lipopolysaccharide endotoxins. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Wiley: Chichester, UK, 2017; pp. 165–172. [Google Scholar]
- Pouria, S.; de Andrade, A.; Barbosa, J.; Cavalcanti, R.L.; Barreto, V.T.; Ward, C.J.; Preiser, W.; Poon, G.K.; Neild, G.H.; Codd, G.A. Fatal microcystin intoxication in haemodialysis unit in Caruaru, Brazil. Lancet 1998, 352, 21–26. [Google Scholar] [CrossRef]
- Jochimsen, E.M.; Carmichael, W.W.; An, J.S.; Cardo, D.M.; Cookson, S.T.; Holmes, C.E.; Antunes, M.B.; de Melo Filho, D.A.; Lyra, T.M.; Barreto, V.S.; et al. Liver failure and death after exposure to microcystins at a hemodialysis center in Brazil. N. Engl. J. Med. 1998, 338, 873–878. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, W.W.; Azevedo, S.M.; An, J.S.; Molica, R.J.; Jochimsen, E.M.; Lau, S.; Rinehart, K.L.; Shaw, G.R.; Eaglesham, G.K. Human fatalities from cyanobacteria: Chemical and biological evidence for cyanotoxins. Environ. Health Perspect. 2001, 109, 663–668. [Google Scholar] [CrossRef] [PubMed]
- Krienitz, L.; Ballot, A.; Kotut, K.; Wiegand, C.; Pütz, S.; Metcalf, J.S.; Codd, G.A.; Pflugmacher, S. Contribution of hot spring cyanobacteria to the mysterious deaths of Lesser Flamingos at Lake Bogoria, Kenya. FEMS Microbiol. Ecol. 2003, 43, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Krienitz, L.; Ballot, A.; Caspar, P.; Codd, G.A.; Kotut, K.; Metcalf, J.S.; Morrison, L.F.; Pflugmacher, S.; Wiegand, C. Contribution of toxic cyanobacteria to massive deaths of Lesser Flamingos at saline-alkaline lakes. Proc. Int. Assoc. Theor. Appl. Limnol. 2005, 29, 783–786. [Google Scholar] [CrossRef]
- Pennycott, T.; Young, F.M.; Metcalf, J.S.; Codd, G.A. Necrotic enteritis in mute swans associated with cyanobacterial toxins. Vet. Rec. 2004, 154, 575–576. [Google Scholar]
- Landsberg, J.H.; Hendrickson, J.; Tabuchi, M.; Kiryu, Y.; Williams, B.J.; Tomlinson, M.C. A large-scale sustained fish kill in the St John’s River, Florida: A complex consequence of cyanobacterial blooms. Harmful Algae 2020, 92, 101771. [Google Scholar] [CrossRef]
- Hawkins, P.R.; Runnegar, M.T.C.; Jackson, A.R.B.; Falconer, I.R. Severe hepatotoxicity caused by the tropical cyanobacterium (blue-green alga) Cylindrospermopsis raciborskii (Woloszynska) Seenaya and Subba Raju isolated from a domestic water supply. Appl. Environ. Microbiol. 1985, 50, 11292–11295. [Google Scholar] [CrossRef] [Green Version]
- Turner, P.C.; Gammie, A.J.; Hollinrake, K.; Codd, G.A. Pneumonia associated with contact with cyanobacteria. Br. Med. J. 1990, 300, 1440–1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, S.-Z. Drinking water and primary liver cancer. In Primary Liver Cancer; Tang, Z.Y., Wu, M.C., Xia, S.S., Eds.; China Academic Publishers: New York, NY, USA, 1989; pp. 30–37. [Google Scholar]
- Yu, S.-Z. Primary prevention of hepatocellular carcinoma. J. Gastroenterol. Hepatol. 1995, 10, 674–682. [Google Scholar] [CrossRef]
- Ueno, Y.; Nagata, S.; Tsutsumi, T.; Hasegawa, A.; Watanabe, M.; Park, H.D.; Chen, G.-C.; Chen, G.; Yu, S.-Z. Detection of microcystins, a blue-green algal hepatotoxin, in drinking water sampled in Haimen and Fusui, endemic areas of primary liver cancer in China, by highly sensitive immunoassay. Carcinogenesis 1996, 17, 1317–1321. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Zeng, H.; Lin, H.; Wang, J.; Feng, Z.; Chen, J.-A.; Luo, J.; Luo, Y.; Huang, Y.; Wang, L.; et al. Serum microcystin levels positively linked with risk of hepatocellular carcinoma: A case-control study in southwest China. Hepatology 2017, 66, 1519–1528. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, S.A.; Andersen, A.J.C.; Andersen, N.G.; Nielsen, K.F.; Hansen, P.J.; Larsen, T.O. Chemical diversity, origin, and analysis of phycotoxins. J. Nat. Prod. 2016, 79, 662–673. [Google Scholar] [CrossRef] [Green Version]
- Vilariño, N.; Louzao, M.C.; Abal, P.; Cagide, E.; Carrera, C.; Vieytes, M.R.; Botana, L.M. Human poisoning from marine toxins: Unknowns for optimal consumer protection. Toxins 2018, 10, 324. [Google Scholar] [CrossRef] [Green Version]
- Akbar, M.A.; Yusof, N.Y.M.; Tahir, N.I.; Ahmad, A.; Usup, G.; Sahrani, F.K.; Bunawan, H. Biosynthesis of saxitoxin in marine dinoflagellates: An omics perspective. Mar. Drugs 2020, 18, 103. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Kudela, R.M.; Mekebri, A.; Crane, D.; Oates, S.C.; Tinker, M.T.; Staedler, M.; Miller, W.A.; Toy-Choutka, S.; Dominik, C.; et al. Evidence for a novel marine harmful algal bloom: Cyanotoxin (microcystin) transfer from land to sea otters. PLoS ONE 2010, 5, e12576. [Google Scholar] [CrossRef]
- Peacock, M.B.; Gibble, C.M.; Senn, D.B.; Cloern, J.E.; Kudela, R.M. Blurred lines: Multiple freshwater and marine algal toxins at the land-sea interface of San Fransisco Bay, California. Harmful Algae 2018, 73, 138–147. [Google Scholar] [CrossRef]
- Codd, G.A.; Nunn, P.B. Cyanotoxin production beyond the cyanobacteria. Toxicon 2019, 168, 93–94. [Google Scholar] [CrossRef]
- Ballot, A.; Bernard, C.; Fastner, J. Saxitoxin and analogues. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Wiley: Chichester, UK, 2017; pp. 149–154. [Google Scholar]
- Smith, F.M.J.; Wood, S.A.; Wilks, T.; Kelly, D.; Broady, P.A.; Williamson, W.; Gaw, S. Survery of Scytonema (Cyanobacteria) and associated saxitoxins in the littoral zone of recreational lakes in Canterbury, New Zealand. Phycologia 2012, 51, 542–551. [Google Scholar]
- Cullen, A.; D’Agostino, P.M.; Mazmouz, R.; Pickford, R.; Wood, S.A.; Nielan, B.A. Insertions within the saxitoxin biosynthetic gene cluster result in differential toxin profiles. ACS Chem. Biol. 2018, 13, 3107–3114. [Google Scholar] [PubMed]
- Cox, P.A.; Banack, S.A.; Murch, S.J.; Rasmussen, U.; Tien, G.; Bidigare, R.R.; Metcalf, J.S.; Morrison, L.F.; Codd, G.A.; Bergman, B. Diverse taxa of cyanobacteria produce β-N-methylamino-L-alanine, a neurotoxic amino acid. Proc. Natl. Acad. Sci. USA 2005, 102, 5074–5078. [Google Scholar] [PubMed] [Green Version]
- Metcalf, J.S.; Banack, S.A.; Lindsay, J.; Morrison, L.F.; Cox, P.A.; Codd, G.A. Co-occurrence of β-N-methylamino-L-alanine a neurotoxic amino acid with other cyanobacterial toxins in British waterbodies, 1990–2004. Environ. Microbiol. 2008, 10, 702–708. [Google Scholar] [PubMed]
- Faassen, E.J.; Gillissen, F.; Lürling, M. A comparative study of three analytical methods for the determination of the neurotoxin BMAA in cyanobacteria. PLoS ONE 2012, 7, e36667. [Google Scholar] [PubMed]
- Jiang, L.; Eriksson, J.; Lage, S.; Jonasson, S.; Shams, S.; Mehine, M.; Ilag, L.L.; Rasmussen, U. Diatoms: A novel source for the neurotoxin BMAA in aquatic environments. PLoS ONE 2014, 9, e84578. [Google Scholar]
- Violi, J.P.; Facey, J.A.; Mitrovic, S.M.; Colville, A.; Rodgers, K.J. Production of β-methylamino-L-alanine (BMAA) and its isomers in freshwater diatoms. Toxins 2019, 11, 1109–1138. [Google Scholar]
- Jiang, L.; Ilag, L.L. Detection of endogenous BMAA in dinoflagellate (Heterocapsa triquetra) hints at evolutionary conservation and environmental concern. Pubraw Sci. 2014, 2, 1–8. [Google Scholar]
- Metcalf, J.S.; Banack, S.A.; Wessel, R.A.; Lester, M.; Pim, J.G.; Cassani, J.R.; Cox, P.A. Toxin analysis of freshwater cyanobacterial and marine harmful algal blooms on the west coast of Florida and implications for estuarine environments. Neurotox. Res. 2020. [Google Scholar] [CrossRef]
- O’Neal, R.M.; Chen, C.-H.; Reynolds, C.S.; Meghal, S.K.; Koeppe, R.E. The ‘neurotoxicity’ of L-2,4-diaminobutyric acid. Biochem. J. 1968, 106, 699–706. [Google Scholar]
- Schneider, T.; Simpson, C.; Desai, P.; Tucker, M.; Lobner, D. Neurotoxicity of isomers of the environmental toxin L-BMAA. Toxicon 2020, 184, 175–179. [Google Scholar] [PubMed]
- Nunn, P.B.; Codd, G.A. Metabolic solutions to the biosynthesis of some diaminomonocarboxylic acids in nature: Formation in cyanobacteria of the neurotoxins 3-N-methyl-2,3-diaminopropanoic acid (BMAA) and 2,4-diaminobutanoic acid (2,4-DAB). Phytochemistry 2017, 144, 2530270. [Google Scholar] [CrossRef] [PubMed]
- Nunn, P.B.; Codd, G.A. Environmental distribution of the neurotoxin L-BMAA in Paenibacillus species. Toxicol. Res. 2019, 8, 781–783. [Google Scholar] [CrossRef] [PubMed]
- Tu, J.; Chen, L.; Gao, S.; Zhang, J.; Bi, C.; Lu, N.; Lu, Z. Obtaining genome sequences of mutualistic bacteria in single Microcystis colonies. Int. J. Mol. Sci. 2019, 20, 5047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, I.; Webb, P.M.; Schluter, P.J.; Shaw, G.R. Recreational and occupational field exposure to freshwater cyanobacteria—A review of anecdotal and case reports, epidemiological studies and the challenges for epidemiologic assessment. Environ. Health 2006, 5, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, R. An address on cholera and its bacillus. Br. Med. J. 1884, 2, 453. [Google Scholar]
- Islam, M.S.; Drasar, B.S.; Sack, R.B. Probable role of blue-green algae in maintaining endemicity and seasonality of cholera in Bangladesh: A hypothesis. J. Diarrhoeal Dis. Res. 1994, 12, 245–256. [Google Scholar] [PubMed]
- Islam, M.S.; Zaman, M.H.; Islam, M.S.; Ahmed, M.; Clemens, J.D. Environmental reservoirs of Vibrio cholerae. Vaccine 2020, 38, A52–A62. [Google Scholar] [CrossRef]
- Ahmed, M.S.; Raknussaman, M.; Akther, H.; Ahmed, S. The role of cyanobacteria blooms in cholera epidemic in Bangladesh. J. Appl. Sci. 2007, 7, 1785–1789. [Google Scholar]
- Froehlich, B.; Gonzalez, R.; Blackwood, D.; Lauer, K.; Noble, R. Decadal monitoring reveals an increase in Vibrio spp. concentrations in the Neuse River Estuary, North Carolina, USA. PLoS ONE 2019, 14, e0215254. [Google Scholar] [CrossRef]
- Paerl, H.W.; Valdes, L.M.; Peierls, B.J.; Adolf, J.E.; Harding, L.W. Anthropogenic and climate influences on the eutrophication of large estuarine ecosystems. Limnol. Oceanogr. 2006, 51, 448–462. [Google Scholar] [CrossRef] [Green Version]
- Marshall, M.M.; Naumovitz, D.; Ortega, Y.; Sterling, C.R. Waterborne protozoan pathogens. Clin. Microbiol. Rev. 1997, 10, 67–85. [Google Scholar] [CrossRef]
- Lopes, A.M.M.B.; Gomes, L.N.L.; de Cerqueira, M.F.; Filho, C.R.M.; von Sperling, E.; de Pádua, V.L. Dynamic of pathogenic protozoa and cyanobacteria in a reservoir used for water supply in southeastern Brazil. Eng. Sanit. E Ambient. 2017, 22, 25–43. [Google Scholar] [CrossRef]
- Fiore, M.F.; Trevors, J.T. Cell composition and metal tolerance in cyanobacteria. BioMetals 2017, 7, 83–103. [Google Scholar] [CrossRef]
- Baptista, M.S.; Vasconcelos, M.T. Cyanobacteria metal interactions: Requirements, Toxicity and ecological implications. Crit. Rev. Microbiol. 2006, 32, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, B.; Megharaj, M.; Venkateswarlu, K.; Naidu, R.; Sethunathan, N. The impacts of environmental pollutants on microalgae and cyanobacteria. Crit. Rev. Environ. Sci. Technol. 2010, 40, 699–821. [Google Scholar] [CrossRef]
- Gu, P.; Qi, L.; Zhang, W.; Zheng, Z.; Luo, X. Effects of different metal ions (Ca, Cu, Pb, Cd) on formation of cyanobacterial blooms. Ecotoxicol. Environ. Saf. 2020, 189, 109976. [Google Scholar] [CrossRef] [PubMed]
- Heresztyn, T.; Nicholson, B.C. Nodularin concentrations in Lake Alexandrina and Albert, South Australia, during a bloom of the cyanobacterium (blue-green alga) Nodularia spumigena and degradation of the toxin. Environ. Toxicol. Water Qual. 1997, 12, 273–282. [Google Scholar] [CrossRef]
- Falconer, I.R.; Beresford, A.M.; Runnegar, M.T. Evidence of liver damage by toxin from a bloom of the blue-green alga Microcystis aeruginosa. Med. J. Aust. 1983, 1, 511–514. [Google Scholar] [CrossRef]
- Utkilen, H.; Gjolme, N. Iron-stimulated toxin production in Microcystis aeruginosa. Appl. Environ. Microbiol. 1995, 61, 797–800. [Google Scholar] [CrossRef] [Green Version]
- Facey, J.A.; Apte, S.C.; Mitrovic, S.M. A review of the effect of trace metals on freshwater cyanobacterial growth and toxin production. Toxins 2019, 11, 643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harland, F.M.J.; Wood, S.A.; Moltchanova, E.; Williams, W.M.; Gaw, S. Phormidium autumnale growth and anatoxin-a production under iron and copper stress. Toxins 2013, 5, 2504–2521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nunn, P.B.; O’Brien, P.; Pettit, L.D.; Pyburn, S.I. Complexes of zinc, copper and nickel with the non-protein amino acid L-α-amino-β-methylaminopropionic acid: A naturally occurring neurotoxin. J. Inorg. Biochem. 1989, 37, 175–183. [Google Scholar] [CrossRef]
- Humble, A.V.; Gadd, G.M.; Codd, G.A. Binding of copper and zinc to three cyanobacterial microcystins quantified by differential pulse polarography. Water Res. 1997, 31, 1679–1686. [Google Scholar] [CrossRef]
- Rush, T.; Liu, X.; Lobner, D. Synergistic toxicity of the environmental neurotoxins methylmercury and β-N-methylamino-l-alanine. Neuroreport 2012, 23, 216–219. [Google Scholar] [CrossRef]
- Metcalf, J.S.; Bruno, M. Anatoxin-a(S). In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Wiley: Chichester, UK, 2017; pp. 155–159. [Google Scholar]
- Singh, A.K.; Singh, P.P.; Tripathi, V.; Verma, H.; Singh, S.K.; Srivastarva, M.K.; Kumar, A. Distribution of cyanobacteria and their interactions with pesticides in paddy field: A comprehensive review. J. Environ. Manag. 2018, 224, 361–375. [Google Scholar] [CrossRef]
- Staley, Z.R.; Harwood, V.J.; Rohr, J.R. A synthesis of the effects of pesticides on microbial persistence in aquatic ecosystems. Crit. Rev. Toxicol. 2015, 45, 813–836. [Google Scholar] [CrossRef] [Green Version]
- Bettinetti, R.; Quadroni, S.; Crosa, G.; Harper, D.; Dickie, J.; Kyalo, M.; Mavuti, K.; Galassi, S. A preliminary evaluation of the DDT contamination of sediments in Lakes Natron and Bogoria (Eastern Rift Valley, Africa). AMBIO 2011, 40, 341–350. [Google Scholar] [CrossRef] [Green Version]
- Woodward, A.R.; Percival, H.F.; Rauschenberger, R.H.; Gross, T.S.; Rice, K.G.; Conrow, R. Abnormal alligators and organochlorine pesticides in Lake Apopka, Florida. In Wildlife Ecotoxicology Forensics Approached; Elliott, J.E., Bishop, C.A., Morrissey, C.A., Eds.; Springer: New York, NY, USA, 2011; pp. 153–187. [Google Scholar]
- Cerbin, S.; Kraak, M.H.S.; de Voogt, P.; Visser, P.M.; van Donk, E. Combined and single effects of pesticide carbaryl and toxic Microcystis aeruginosa on the life history of Daphnia pulicaria. Hydrobiologia 2010, 643, 129–138. [Google Scholar] [CrossRef] [Green Version]
- Metcalf, J.S.; Beattie, K.A.; Ressler, J.; Gerbersdorf, S.; Pflugmacher, S.; Codd, G.A. Cross-reactivity and performance assessment of four microcystin immunoassays with detoxication products of the cyanotoxin, microcystin-LR. J. Water Supply Res. Technol. AQUA 2002, 51, 145–151. [Google Scholar] [CrossRef]
- Gill, J.P.K.; Sethi, N.; Mohan, A.; Datta, S.; Girdhar, M. Glyphosate toxicity for animals. Environ. Chem. Lett. 2017. [Google Scholar] [CrossRef]
- Zhang, Q.; Qu, Q.; Lu, T.; Ke, M.; Zhu, Y.; Zhang, M.; Zhang, Z.; Du, B.; Pan, X.; Sun, L.; et al. The combined toxicity effect of nanoplastics and glyphosate on Microcystis aeruginosa growth. Environ. Pollut. 2018, 243, 1106–1112. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Qiu, Z.; Zhou, Y.; Du, Y.; Liu, C.; Ye, J.; Hu, X. Physiological effects of the herbicide glyphosate on the cyanobacterium Microcystis aeruginosa. Aquat. Toxicol. 2016, 178, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Malécot, M.; Guével, B.; Pineau, C.; Holbech, B.F.; Bormans, M.; Wiegand, C. Specific proteome response of Unio pictorum mussel to a mixture of glyphosate and microcystin-LR. J. Proteome Res. 2013, 12, 5281–5292. [Google Scholar]
- Moore, C.J. Synthetic polymers in the marine environment: A rapidly increasing long-term threat. Environ. Res. 2008, 108, 131–139. [Google Scholar] [CrossRef]
- Yokota, K.; Waterfield, H.; Hastings, C.; Davidson, E.; Kwietniewski, E.; Wells, B. Finding the missing piece of the aquatic pollution puzzle: Interaction between primary producers and microplastics. Limnol. Oceanogr. Lett. 2017, 2, 91–104. [Google Scholar] [CrossRef] [Green Version]
- Yao, L.; Hui, L.; Yang, Z.; Chen, X.; Xiao, A. Freshwater microplastics pollution: Detecting and visualizing emerging trends based on Citespace II. Chemosphere 2020, 245, 125627. [Google Scholar] [CrossRef]
- Leiser, R.; Wu, G.-M.; Neu, T.R.; Wendt-Potthoff, K. Biofouling, metal sorption and aggregation are related to sinking of microplastics in a stratified reservoir. Water Res. 2020, 176, 115748. [Google Scholar] [CrossRef]
- Feng, L.J.; Sun, X.-D.; Zhu, F.-P.; Feng, Y.; Duan, J.-L.; Xiao, F.; Li, X.-Y.; Shi, Y.; Wang, Q.; Sun, J.-W.; et al. Nanoplastics promote microcystin synthesis and release from cyanobacterial Microcystis aeruginosa. Environ. Sci. Technol. 2020, 54, 3386–3394. [Google Scholar] [CrossRef]
- Hyenstrand, P.; Metcalf, J.S.; Beattie, K.A.; Codd, G.A. Losses of the cyanobacterial toxin microcystin-LR from aqueous solution by adsorption during laboratory manipulations. Toxicon 2001, 39, 589–594. [Google Scholar] [CrossRef]
- Hyenstrand, P.; Metcalf, J.S.; Beattie, K.A.; Codd, G.A. Effects of adsorption to plastics and solvent conditions in the analysis of the cyanobacterial toxin microcystin-LR by high performance liquid chromatography. Water Res. 2001, 35, 3508–3511. [Google Scholar] [CrossRef]
- Hitzfeld, B.C.; Höger, S.; Dietrich, D.R. Cyanobacterial toxins: Removal during water treatment, and human risk assessment. Environ. Health Perspect. 2000, 108, 113–122. [Google Scholar] [PubMed] [Green Version]
- Delgado, L.F.; Charles, P.; Glucina, K.; Morlay, C. The removal of endocrine disrupting compounds, pharmaceutically activated compounds and cyanobacterial toxins during drinking water preparation using activated carbon—A review. Sci. Total Environ. 2012, 435–436, 509–525. [Google Scholar] [CrossRef] [PubMed]
- Hageskal, G.; Lima, N.; Skaar, I. The study of fungi in drinking water. Mycol. Res. 2009, 113, 165–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanches, S.; Crespo, M.T.B.; Pereira, V.J. Drinking water treatment of priority pesticides using low pressure UV photolysis and advanced oxidation processes. Water Res. 2010, 44, 1809–1818. [Google Scholar] [CrossRef]
- Griffini, O.; Bao, M.L.; Burrini, D.; Santianni, D.; Barbieri, C.; Pantani, F. Removal of pesticides during water treatment process at Florence water supply, Italy. J. Water Supply Res. Technol. AQUA 1999, 48, 177–185. [Google Scholar] [CrossRef]
- Ignatowicz, K. Selection of sorbent for removing pesticides during water treatment. J. Hazard. Mater. 2009, 169, 953–957. [Google Scholar] [CrossRef]
- Ma, B.; Xue, W.; Hu, C.; Liu, H.; Qu, J.; Li, L. Characteristics of microplastic removal via coagulation and ultrafiltration during drinking water treatment. Chem. Eng. J. 2019, 359, 159–167. [Google Scholar] [CrossRef]
- Pivokonsky, M.; Cermakova, L.; Novotna, K.; Peer, P.; Cajthaml, T.; Janda, V. Occurrence of microplastics in raw and treated drinking water. Sci. Total Environ. 2018, 643, 1644–1651. [Google Scholar] [CrossRef]
- Elliott, H.A.; Dempsey, B.A.; Maille, P.J. Content and fractionation of heavy metals in water treatment sludges. J. Environ. Qual. 1990, 19, 330–334. [Google Scholar] [CrossRef] [Green Version]
- Chiang, Y.W.; Ghyselbrecht, K.; Santos, R.M.; Martens, J.A.; Swennen, R.; Cappuyns, V.; Meesschaert, B. Adsorption of multi-heavy metals onto water treatment residuals: Sorption capacities and applications. Chem. Eng. J. 2012, 200–202, 405–415. [Google Scholar] [CrossRef] [Green Version]
- Lambert, T.W.; Holmes, C.F.B.; Hrudey, S.E. Adsorption of microcystin-LR by activated carbon and removal in full scale water treatment. Water Res. 1996, 30, 1411–1422. [Google Scholar] [CrossRef]
- Vlad, S.; Anderson, W.B.; Peldszus, S.; Huck, P.M. Removal of the cyanotoxin anatoxin-a by drinking water treatment processes: A review. J. Water Health 2014, 12, 601–617. [Google Scholar] [CrossRef] [PubMed]
- Ho, L.; Tanis-Plant, P.; Kayal, N.; Slyman, N.; Newcombe, G. Optimising water treatment practices for the removal of Anabaena circinalis and its associated metabolites, geosmin and saxitoxins. J. Water Health 2009, 7, 544–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senogles, P.; Shaw, G.; Smith, M.; Norris, R.; Chiswell, R.; Mueller, J.; Sadler, R.; Eaglesham, G. Degradation of the cyanobacterial toxin cylindrospermopsin, from Cylindrospermopsis raciborskii, by chlorination. Toxicon 2000, 38, 1203–1213. [Google Scholar] [CrossRef]
- Falconer, I.R.; Humpage, A.R. Cyanobacterial (blue-green algal) toxins in water supplies: Cylindrospermopsins. Environ. Toxicol. 2006, 21, 299–304. [Google Scholar] [CrossRef]
- Metcalf, J.S.; Codd, G.A. The status and potential of cyanobacteria and their toxins as agents of bioterrorism. In Handbook on Cyanobacteria: Biochemistry, Biotechnology and Applications; Gault, P.M., Marler, H.J., Eds.; Nova Science Publishers: New York, NY, USA, 2009; pp. 259–281. [Google Scholar]
- Chorus, I.; Bartram, J. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; E.&F.N. Spon: London, UK, 1999. [Google Scholar]
- World Health Organization. Microplastics in Drinking Water; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- World Health Organization. Guidelines for Drinking Water Quality, 4th ed.; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Merlo, F.; Maraschi, F.; Piparo, D.; Profumo, A.; Speltini, A. Simultaneous pre-concentration and HPLC-MS/MS quantification of phycotoxins and cyanotoxins in inland and coastal waters. Int. J. Environ. Res. Public Health 2020, 17, 4782. [Google Scholar] [CrossRef]
- Manton, W.I. Total contribution of airborne lead to blood lead. Br. J. Ind. Med. 1985, 42, 168–172. [Google Scholar] [CrossRef]
- Querol, X.; Alastuey, A.; Moreno, T.; Viana, M.M.; Castillo, S.; Pey, J.; Rodríguez, S.; Artiñano, B.; Salvador, P.; Sánchez, M.; et al. Spatial and temportal variation in airborne particulate matter (PM10 and PM2.5) across Spain 1999–2005. Atmos. Environ. 2008, 42, 3964–3979. [Google Scholar] [CrossRef]
- Metcalf, J.S.; Richer, R.; Cox, P.A.; Codd, G.A. Cyanotoxins in desert environments may present a risk to human health. Sci. Total Environ. 2012, 421–422, 118–123. [Google Scholar] [CrossRef]
- Lentz, D.L.; Hamilton, T.L.; Dunning, N.P.; Scarborough, V.L.; Luxton, T.P.; Vonderheide, A.; Tepe, E.J.; Perfetta, C.J.; Brunemann, J.; Grazioso, L.; et al. Molecular genetic and geochemical assays reveal severe contamination of drinking water reservoirs at the ancient Maya city of Tikal. Sci. Rep. 2020, 10, 10316. [Google Scholar] [CrossRef]


{kind=link}
| Toxic Agents | Test Organism/Cell Line | Outcome 1 | Ref. |
|---|---|---|---|
| MC-LR plus Antx-a | Mouse, intranasal bioassay | SYN | [26] |
| MC-LR plus Antx-a | Selenastrum capricornutum | SYN | [27] |
| MC-LR plus Antx-a | Cyprinus carpio, carp cells | SYN (potential) | [28] |
| MC-LR plus Antx-a | Vallisneria natans, and microbial biofilm | ANTAG | [29] |
| MC-LR plus Microcystis LPS | Artemia salina and Daphnia sp. | ANTAG | [30] |
| MC-LR plus CYN | Chlorella vulgaris | SYN | [31] |
| MC-LR plus cyanobacterial λ-linolenic acid | Daphnia magna | ADDIT | [32] |
| MC-LR plus aflatoxin B1 | Human hepatic cells | ANTAG | [33] |
| MC-LR plus aflatoxin B1, and plus fumonisin B1 | HepG2, Caco2, MDBK cell lines | ADDIT, SYN, ANTAG | [34] |
| MC-LR and copper | Danio rerio | SYN | [35] |
| MC-LR plus copper | Vallisneria natans | SYN | [36] |
| MC-LR plus linear alkyl- benzene sulphonate | Lactuca sativa | SYN | [37] |
| MC-LR plus phenanthrene | Lemna gibba | ANTAG, SYN, ADDIT | [38] |
| CYN plus chloropyrifos | Human SH-SY5Y neuroblastoma cell line | ANTAG | [39] |
| CYN plus bisphenols | HepG2 cells | SYN, ADDIT | [40] |
| Toxicant | Treatment Process | References |
|---|---|---|
| Fungal/microbial toxins | SF, C | [123] |
| Pesticides | Oz, AC, | [124,125,126] |
| Microplastics | C, U | [127,128] |
| Metals | C, A | [129,130] |
| Microcystins | AC, Ox, Oz, Cl | [121,131] |
| Anatoxin-a | Oz, Ox, Cl | [132] |
| Saxitoxins | AC, Cl | [133] |
| Cylindrospermopsin | AC, Oz, Cl | [134,135] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Metcalf, J.S.; Codd, G.A. Co-Occurrence of Cyanobacteria and Cyanotoxins with Other Environmental Health Hazards: Impacts and Implications. Toxins 2020, 12, 629. https://doi.org/10.3390/toxins12100629
Metcalf JS, Codd GA. Co-Occurrence of Cyanobacteria and Cyanotoxins with Other Environmental Health Hazards: Impacts and Implications. Toxins. 2020; 12(10):629. https://doi.org/10.3390/toxins12100629
Chicago/Turabian StyleMetcalf, James S., and Geoffrey A. Codd. 2020. "Co-Occurrence of Cyanobacteria and Cyanotoxins with Other Environmental Health Hazards: Impacts and Implications" Toxins 12, no. 10: 629. https://doi.org/10.3390/toxins12100629
APA StyleMetcalf, J. S., & Codd, G. A. (2020). Co-Occurrence of Cyanobacteria and Cyanotoxins with Other Environmental Health Hazards: Impacts and Implications. Toxins, 12(10), 629. https://doi.org/10.3390/toxins12100629