Structural Insight into Integrin Recognition and Anticancer Activity of Echistatin


Abstract
:1. Introduction
2. Results
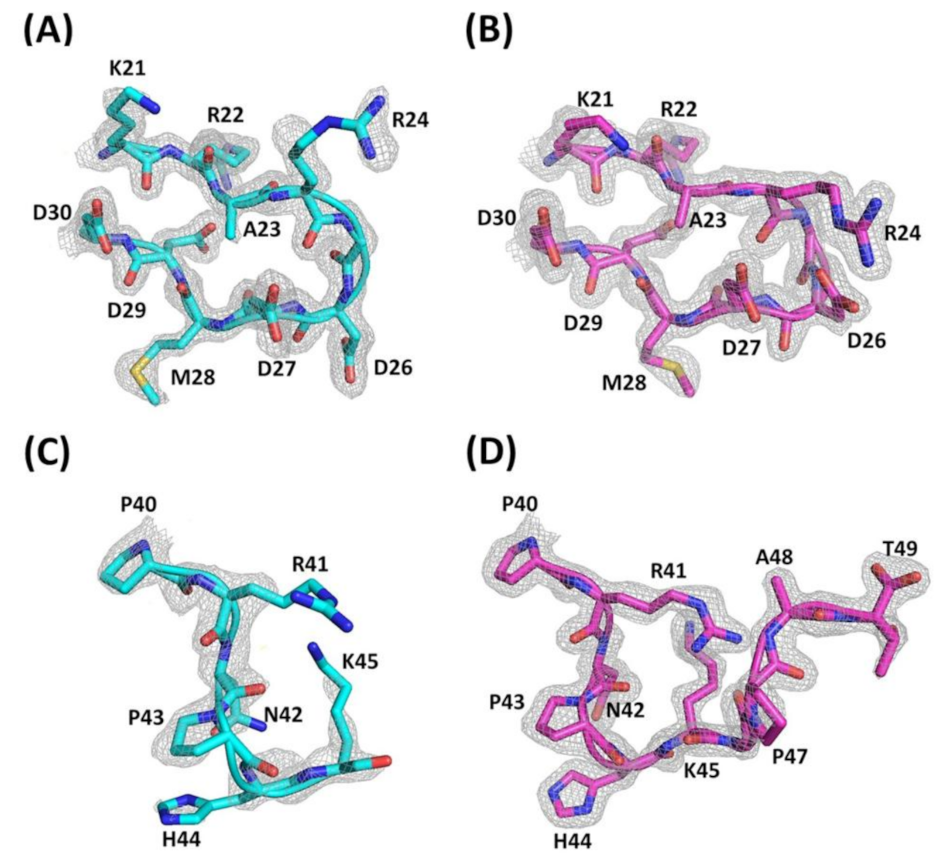
2.1. The Overall Structure of Ech
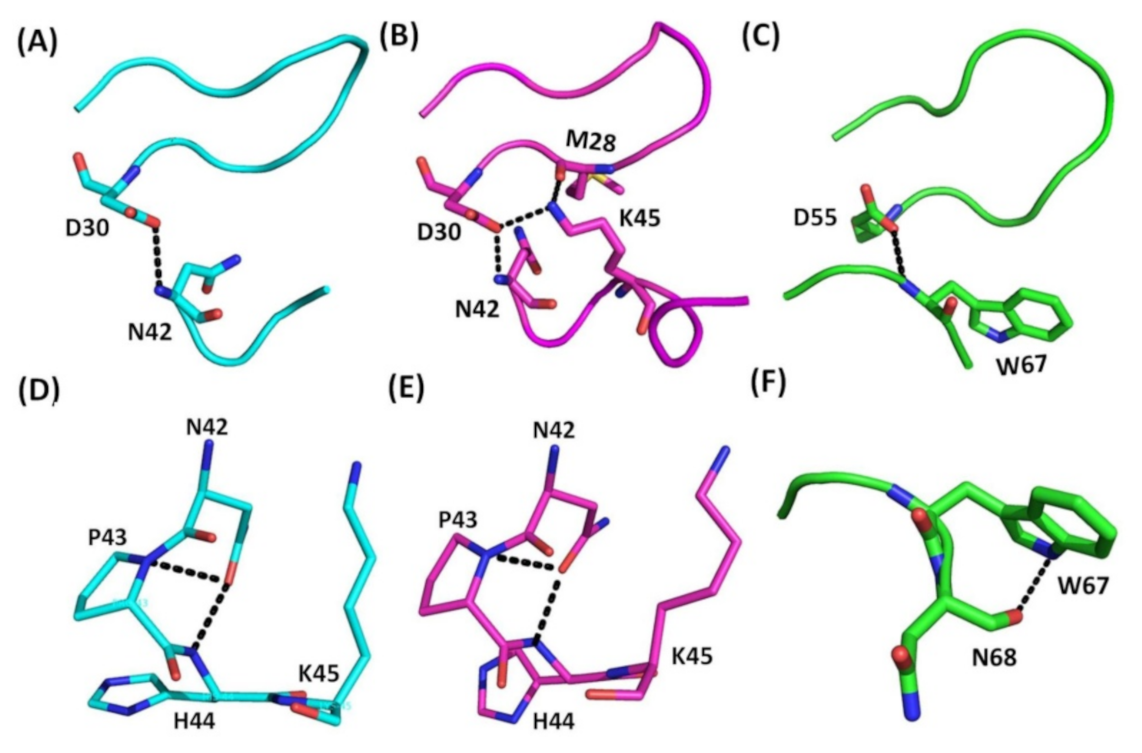
2.2. Structural Comparison of the RGD Loop and the Interactions between the RGD Loop and C-Termini
2.3. Structural Comparison of the C-Terminal Regions
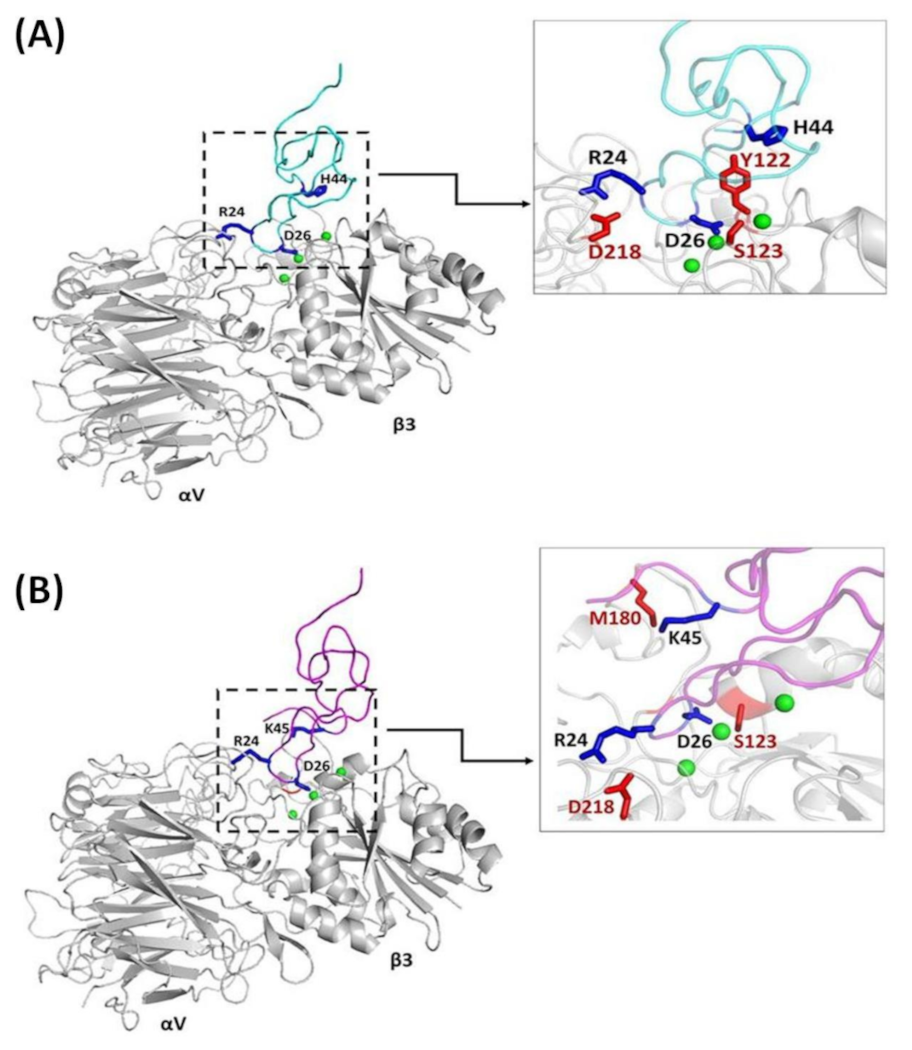
2.4. Structural Differences between the Integrin Complexes of Ech Chain A and Chain B
2.5. The C-Terminal Residues of Ech are Involved in Mediating Integrin Recognition
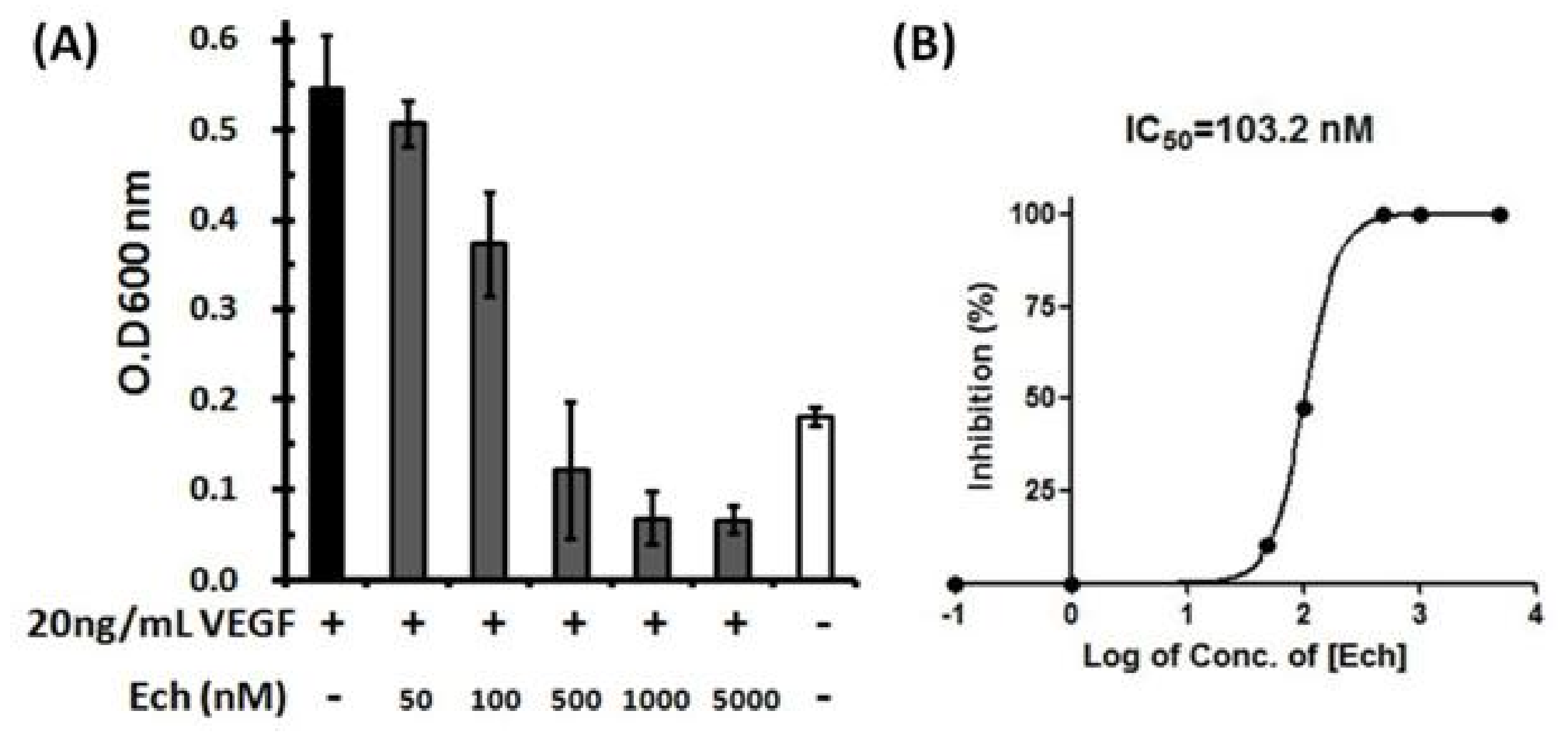
2.6. Inhibition of VEGF-Induced HUVEC Proliferation by Ech
2.7. C-Terminal Residue K45 of Ech Is Responsible for Its Anti-Metastatic Ability
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Expression, Purification, and Characterization of Ech and Its Mutants
5.2. Mass Spectrometric Measurements
5.3. Cell Adhesion Assay
5.4. Crystallization of Ech
5.5. Diffraction Data Collection and Processing
5.6. Structure Determination and Refinement
5.7. NOE Analysis
5.8. Molecular Docking
5.9. VEGF-Induced HUVEC Proliferation Assay
5.10. Cell Migration Assay
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gan, Z.R.; Gould, R.J.; Jacobs, J.W.; Friedman, P.A.; Polokoff, M.A. A potent platelet aggregation inhibitor from the venom of the viper, Echis carinatus. J. Biol. Chem. 1988, 263, 19827–19832. [Google Scholar] [PubMed]
- Savage, B.; Marzec, U.M.; Chao, B.H.; Harker, L.A.; Maraganore, J.M.; Ruggeri, Z.M. Binding of the snake venom-derived proteins applaggin and echistatin to the arginine-glycine-aspartic acid recognition site(s) on platelet glycoprotein IIb.IIIa complex inhibits receptor function. J. Biol. Chem. 1990, 265, 11766–11772. [Google Scholar]
- Atkinson, R.; Saudek, V.; Pelton, J. Echistatin: The refined structure of a disintegrin in solution by 1 H NMR and restrained molecular dynamics. Int. J. Pept. Protein Res. 2009, 43, 563–572. [Google Scholar] [CrossRef]
- Garsky, V.M.; Lumma, P.K.; Freidinger, R.M.; Pitzenberger, S.M.; Randall, W.C.; Veber, D.F.; Gould, R.J.; Friedman, P.A. Chemical synthesis of echistatin, a potent inhibitor of platelet aggregation from Echis carinatus: Synthesis and biological activity of selected analogs. Proc. Natl. Acad. Sci. USA 1989, 86, 4022–4026. [Google Scholar] [CrossRef] [Green Version]
- Cooke, R.M.; Carter, B.G.; Martin, D.M.A.; Murray-Rust, P.; Weir, M.P. Nuclear magnetic resonance studies of the snake toxin echistatin. 1H resonance assignments and secondary structure. JBIC J. Biol. Inorg. Chem. 1991, 202, 323–328. [Google Scholar] [CrossRef]
- Monleon, D.; Esteve, V.; Kovacs, H.; Calvete, J.J.; Celda, B. Conformation and concerted dynamics of the integrin-binding site and the C-terminal region of echistatin revealed by homonuclear NMR. Biochem. J. 2005, 387, 57–66. [Google Scholar] [CrossRef]
- Marcinkiewicz, C.; Vijay-Kumar, S.; McLane, M.A.; Niewiarowski, S. Significance of RGD loop and C-terminal domain of echistatin for recognition of alphaIIb beta3 and alpha(v) beta3 integrins and expression of ligand-induced binding site. Blood 1997, 90, 1565–1575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, T.; Kidera, A. Tailoring echistatin to possess higher affinity for integrin alpha (IIb)beta (3). FEBS Lett. 1996, 387, 11–15. [Google Scholar] [CrossRef] [Green Version]
- McLane, M.A.; Kowalska, M.A.; Silver, L.; Shattil, S.J.; Niewiarowski, S. Interaction of disintegrins with the αIIbβ3 receptor on resting and activated human platelets. Biochem. J. 1994, 301, 429–436. [Google Scholar] [CrossRef]
- Wierzbicka-Patynowski, I.; Niewiarowski, S.; Marcinkiewicz, C.; Calvete, J.J.; Marcinkiewicz, M.M.; McLane, M.A. Structural requirements of echistatin for the recognition of alpha(v)beta(3) and alpha(5)beta(1) integrins. J. Biol. Chem. 1999, 274, 37809–37814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zannetti, A.; Del Vecchio, S.; Iommelli, F.; Del Gatto, A.; De Luca, S.; Zaccaro, L.; Papaccioli, A.; Sommella, J.; Panico, M.; Speranza, A.; et al. Imaging of v 3 Expression by a Bifunctional Chimeric RGD Peptide not Cross-Reacting with v 5. Clin. Cancer Res. 2009, 15, 5224–5233. [Google Scholar] [CrossRef] [Green Version]
- Minea, R.O.; Helchowski, C.M.; Zidovetzki, S.J.; Costa, F.K.; Swenson, S.D.; Markland, F. Vicrostatin – An Anti-Invasive Multi-Integrin Targeting Chimeric Disintegrin with Tumor Anti-Angiogenic and Pro-Apoptotic Activities. PLoS ONE 2010, 5, e10929. [Google Scholar] [CrossRef]
- Chen, Y.-C.; Cheng, C.-H.; Shiu, J.-H.; Chang, Y.-T.; Chang, Y.-S.; Chuang, W.-J.; Lee, J.-C. Expression in Pichia pastoris and characterization of echistatin, an RGD-containing short disintegrin. Toxicon 2012, 60, 1342–1348. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-Y.; Shiu, J.-H.; Hsieh, Y.-H.; Liu, Y.-C.; Chen, Y.-C.; Chen, Y.-C.; Jeng, W.-Y.; Tang, M.-J.; Lo, S.J.; Chuang, W.-J. Effect of D to E mutation of the RGD motif in rhodostomin on its activity, structure, and dynamics: Importance of the interactions between the D residue and integrin. Proteins Struct. Funct. Bioinform. 2009, 76, 808–821. [Google Scholar] [CrossRef]
- Fujii, Y.; Okuda, D.; Fujimoto, Z.; Horii, K.; Morita, T.; Mizuno, H. Crystal Structure of Trimestatin, a Disintegrin Containing a Cell Adhesion Recognition Motif RGD. J. Mol. Biol. 2003, 332, 1115–1122. [Google Scholar] [CrossRef]
- Senn, H.; Klaus, W. The Nuclear Magnetic Resonance Solution Structure of Flavoridin, an Antagonist of the Platelet GP IIb-IIIa Receptor. J. Mol. Biol. 1993, 232, 907–925. [Google Scholar] [CrossRef]
- Shin, J.; Hong, S.Y.; Chung, K.; Kang, I.; Jang, Y.; Kim, D.S.; Lee, W. Solution Structure of a Novel Disintegrin, Salmosin, fromAgkistrondon halysVenom. Biochemistry 2003, 42, 14408–14415. [Google Scholar] [CrossRef]
- Chang, Y.-T.; Shiu, J.-H.; Chuang, W.-J.; Chen, Y.-C.; Chen, C.-Y.; Chang, Y.-S. Effects of the RGD loop and C-terminus of rhodostomin on regulating integrin αIIbβ3 recognition. PLoS ONE 2017, 12, e0175321. [Google Scholar] [CrossRef] [Green Version]
- Saudek, V.; Atkinson, R.A.; Pelton, J.T. Three-dimensional structure of echistatin, the smallest active RGD protein. Biochemistry 1991, 30, 7369–7372. [Google Scholar] [CrossRef]
- Rádis-Baptista, G. Integrins, cancer and snake toxins (mini-review). J. Venom. Anim. Toxins Incl. Trop. Dis. 2005, 11, 217–241. [Google Scholar] [CrossRef]
- Alday-Parejo, B.; Stupp, R.; Rüegg, C. Are Integrins Still Practicable Targets for Anti-Cancer Therapy? Cancers 2019, 11, 978. [Google Scholar] [CrossRef] [Green Version]
- Desgrosellier, J.S.; Cheresh, D.A. Integrins in cancer: Biological implications and therapeutic opportunities. Nat. Rev. Cancer. 2010, 10, 9–22. [Google Scholar] [CrossRef] [Green Version]
- Nieberler, M.; Reuning, U.; Reichart, F.; Notni, J.; Wester, H.-J.; Schwaiger, M.; Weinmüller, M.; Räder, A.; Steiger, K.; Kessler, H. Exploring the Role of RGD-Recognizing Integrins in Cancer. Cancers 2017, 9, 116. [Google Scholar] [CrossRef]
- Selistre-De-Araújo, H.S.; Pontes, C.L.S.; Montenegro, C.F.; Martin, A.C.B.M. Snake Venom Disintegrins and Cell Migration. Toxins 2010, 2, 2606–2621. [Google Scholar] [CrossRef]
- McLane, M.A. Disintegrins in health and disease. Front. Biosci. 2008, 13, 6617–6637. [Google Scholar] [CrossRef]
- McLane, M.A.; Sanchez, E.E.; Wong, A.; Paquette-Straub, C.; Perez, J.C. Disintegrins. Curr. Drug Targets Cardiovasc. Haematol. Disord. 2004, 4, 327–355. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Mercado, E.; Garza-Ocañas, L. Disintegrins obtained from snake venom and their pharmacological potential. Med. Univ. 2017, 19, 32–37. [Google Scholar] [CrossRef]
- Macêdo, J.K.; Fox, J.W.; Castro, M.D.S. Disintegrins from Snake Venoms and their Applications in Cancer Research and Therapy. Curr. Protein Pept. Sci. 2015, 16, 532–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- David, V.; Succar, B.B.; De Moraes, J.A.; Saldanha-Gama, R.F.G.; Barja-Fidalgo, C.; Zingali, R.B. Recombinant and Chimeric Disintegrins in Preclinical Research. Toxins 2018, 10, 321. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.-Y.; Koh, Y.-S.; Chung, K.-H.; Kim, D.-S. Snake venom disintegrin, saxatilin, inhibits platelet aggregation, human umbilical vein endothelial cell proliferation, and smooth muscle cell migration. Thromb. Res. 2002, 105, 79–86. [Google Scholar] [CrossRef]
- Yeh, C.H.; Peng, H.C.; Yang, R.S.; Huang, T.F. Rhodostomin, a snake venom disintegrin, inhibits angiogenesis elicited by basic fibroblast growth factor and suppresses tumor growth by a selective alpha(v)beta (3) blockade of endothelial cells. Mol. Pharmacol. 2001, 59, 1333–1342. [Google Scholar] [CrossRef] [Green Version]
- Sheu, J.-R.; Yen, M.-H.; Kan, Y.-C.; Hung, W.-C.; Chang, P.-T.; Luk, H.-N. Inhibition of angiogenesis in vitro and in vivo: Comparison of the relative activities of triflavin, an Arg-Gly-Asp-containing peptide and anti-αvβ3 integrin monoclonal antibody. Biochim. Biophys. Acta (BBA)—Gen. Subj. 1997, 1336, 445–454. [Google Scholar] [CrossRef]
- Lazarovici, P.; Marcinkiewicz, C.; Lelkes, P.I. From Snake Venom’s Disintegrins and C-Type Lectins to Anti-Platelet Drugs. Toxins 2019, 11, 303. [Google Scholar] [CrossRef] [Green Version]
- Williams, C.J.; Headd, J.J.; Moriarty, N.W.; Prisant, M.G.; Videau, L.L.; Deis, L.N.; Verma, V.; Keedy, D.A.; Hintze, B.J.; Chen, V.B.; et al. MolProbity: More and better reference data for improved all-atom structure validation. Protein Sci. 2018, 27, 293–315. [Google Scholar] [CrossRef]
- Nishida, N.; Yano, H.; Nishida, T.; Kamura, T.; Kojiro, M. Angiogenesis in cancer. Vasc. Health Risk Manag. 2006, 2, 213–219. [Google Scholar] [CrossRef]
- Sumathipala, R.; Xu, C.; Seago, J.; Mould, A.P.; Humphries, M.J.; Craig, S.E.; Patel, Y.; Wijelath, E.S.; Sobel, M.; Rahman, S. The "Linker" Region (Amino Acids 38-47) of the Disintegrin Elegantin Is a Novel Inhibitory Domain of Integrin 5beta1-Dependent Cell Adhesion on Fibronectin: EVIDENCE FOR THE NEGATIVE REGULATION OF FIBRONECTIN SYNERGY SITE BIOLOGICAL ACTIVITY. J. Biol. Chem. 2006, 281, 37686–37696. [Google Scholar] [CrossRef] [Green Version]
- Swenson, S.D.; Minea, R.O.; Tuan, C.D.; Thein, T.-Z.; Chen, T.C.; Markland, F. A Novel Venom-Derived Peptide for Brachytherapy of Glioblastoma: Preclinical Studies in Mice. Molecules 2018, 23, 2918. [Google Scholar] [CrossRef] [Green Version]
- Tome, Y.; Kimura, H.; Kiyuna, T.; Sugimoto, N.; Tsuchiya, H.; Kanaya, F.; Bouvet, M.; Hoffman, R.M. Disintegrin targeting of an αvβ3 integrin-over-expressing high-metastatic human osteosarcoma with echistatin inhibits cell proliferation, migration, invasion and adhesion in vitro. Oncotarget 2016, 7, 46315–46320. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.; Rivera, A.; Tao, L.; Zhang, X. Genetically modified T cells targeting neovasculature efficiently destroy tumor blood vessels, shrink established solid tumors and increase nanoparticle delivery. Int. J. Cancer 2013, 133, 2483–2492. [Google Scholar] [CrossRef]
- Guo, R.-T.; Chou, L.-J.; Chen, Y.-C.; Chen, C.-Y.; Pari, K.; Jen, C.J.; Lo, S.J.; Huang, S.-L.; Lee, C.-Y.; Chang, T.-W.; et al. Expression in Pichia pastoris and characterization by circular dichroism and NMR of rhodostomin. Proteins Struct. Funct. Bioinform. 2001, 43, 499–508. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Cheng, C.-H.; Chen, Y.-C.; Lee, J.-C.; Choua, S.-H.; Huang, W.; Chuang, W.-J. Preparation of amino-acid-type selective isotope labeling of protein expressed in Pichia pastoris. Proteins Struct. Funct. Bioinform. 2005, 62, 279–287. [Google Scholar] [CrossRef]
- Shiu, J.-H.; Chen, C.-Y.; Chen, Y.-C.; Chang, Y.-T.; Chang, Y.-S.; Huang, C.-H.; Chuang, W.-J. Effect of P to A Mutation of the N-Terminal Residue Adjacent to the Rgd Motif on Rhodostomin: Importance of Dynamics in Integrin Recognition. PLoS ONE 2012, 7, e28833. [Google Scholar] [CrossRef] [Green Version]
- Kabsch, W. XDS. Acta Cryst. D 2010, 66, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. Sect. D Biol. Crystallogr. 2004, 60, 2126–2132. [Google Scholar] [CrossRef] [Green Version]
- Murshudov, G.N.; Skubák, P.; Lebedev, A.A.; Pannu, N.S.; Steiner, R.A.; Nicholls, R.A.; Winn, M.D.; Long, F.; Vagin, A.A. REFMAC5 for the refinement of macromolecular crystal structures. Acta Crystallogr. Sect. D Biol. Crystallogr. 2011, 67, 355–367. [Google Scholar] [CrossRef] [Green Version]
- De Vries, S.J.; Van Dijk, M.; Bonvin, A.M.J.J. The HADDOCK web server for data-driven biomolecular docking. Nat. Protoc. 2010, 5, 883–897. [Google Scholar] [CrossRef] [Green Version]
- Van Agthoven, J.F.; Xiong, J.-P.; Alonso, J.L.; Rui, X.; Adair, B.D.; Goodman, S.L.; Arnaout, M.A. Structural basis for pure antagonism of integrin αVβ3 by a high-affinity form of fibronectin. Nat. Struct. Mol. Biol. 2014, 21, 383–388. [Google Scholar] [CrossRef] [Green Version]
- CCP4i Software. Available online: http://structure.usc.edu/ccp4/ (accessed on 9 May 2020).
- PISA Server. Available online: https://www.ebi.ac.uk/pdbe/pisa/pi_link.html (accessed on 22 March 2020).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Crystals PDB Code | Echistatin 6LSQ |
|---|---|
| Data collection | |
| Radiation source | NSRRC TPS05A |
| Wavelength (Å) | 0.99984 |
| Space group | P3221 |
| Unit cell parameters | |
| a (Å) | 33.79 |
| b (Å) | 33.79 |
| c (Å) | 120.90 |
| α | 90.00 |
| β | 90.00 |
| γ | 120.00 |
| Resolution (Å) | 30.00–1.80 (1.91–1.80) |
| No. of reflections | 7695 (1245) |
| Completeness (%) | 92.5 (90.0) |
| I/σ (I) | 7.88 (2.96) |
| CC1/2 | 99.1 (95.1) |
| Refinement | |
| Resolution (Å) | 28.44–1.80 |
| Reflections (work) | 7541 |
| Reflections (free) | 419 |
| Rwork (%) | 21.2 |
| Rfree (%) | 25.5 |
| Geometry deviations | |
| Bond length (Å) | 0.009 |
| Bond angles (°) | 1.461 |
| Mean B values (Å2) | 28.851 |
| Ramachandran plot 1 (%) | |
| Favored | 95.7 |
| Allowed | 4.3 |
| Ligands | RGD Motif | Cα(Ri)-Cα(Di + 2) | Cβ(Ri)-Cβ(Di + 2) | Cζ(Ri)-Cγ(Di + 2) | Cα(Ri)-Cα(Xi + 3) |
|---|---|---|---|---|---|
| Echistatin (NMR) 1 | ARGDDM | 6.2 ± 0.75 | 8.2 ± 1.1 | 12.3 ± 0.7 | 6.3 ± 0.8 |
| Echistatin (A chain) | ARGDDM | 6.5 | 8.6 | 12.4 | 6.2 |
| Echistatin (B chain) | ARGDDM | 5.8 | 6.0 | 4.8 | 5.4 |
| Trimestatin 2 | ARGDFP | 6.7 | 9.2 | 11.1 | 6.8 |
| Ech | αvβ3 | |
|---|---|---|
| Chain A | αv | β3 |
| R22 | N313 | |
| R24 | D150, D218 | |
| G25 | D218 | |
| D26 | S121, Y122, S123, N215, R216, D217 | |
| Y31 | D126 | |
| H44 | Y122 | |
| Ech | αvβ3 | |
|---|---|---|
| Chain B | αv | β3 |
| R24 | D150, D218 | |
| D26 | S121, Y122, S123, N215, R216, D217 | |
| M28 | D126 | |
| Y31 | D126 | |
| K45 | M180 | |
| Protein | Sequence | IC50 (nM) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| (Mutant) | RGD Loop | C-Terminus | αvβ3 | αIIbβ3 | α5β1 | αvβ5 | |||
| Ech | 23ARGDDM | 42NPHKGPAT | 20.7 ± 8.0 | 51.5 ± 3.9 | 132.6 ± 15.7 | 286.4 ± 53.6 | |||
| Ech (P43∆) | 23ARGDDM | 42NP | 132.0 ± 12.0 | 362.3 ± 56.3 | 2469.5 ± 97.7 | 3358.3 ± 1040.4 | |||
| Protein | RGD Loop | C-Terminus | IC50 (nM) | ||
|---|---|---|---|---|---|
| (Mutant) | Sequence | Sequence | αvβ3 | αIIbβ3 | α5β1 |
| Ech | 23ARGDDM | 42NPHKGPAT | 20.7 ± 8.0 | 51.5 ± 3.9 | 132.6 ± 15.7 |
| Ech (P43A) | 23ARGDDM | 42NAHKGPAT | 13.8 ± 1.1 | 67.7 ± 35.0 | 58.5 ± 9.5 |
| Ech (H44A) | 23ARGDDM | 42NPAKGPAT | 16.4 ± 6.9 | 20.7 ± 4.5 | 30.0 ± 5.3 |
| Ech (K45A) | 23ARGDDM | 42NPHAGPAT | 14.7 ± 4.0 | 132.5 ± 53.3 | 84.9 ± 24.9 |
| Protein | RGD Motif | C-Terminal Sequence | IC50 (nM) |
|---|---|---|---|
| Echistatin | ARGDDM | NPHKGPAT | 103.2 |
| Saxatilin | ARGDDM | NPFHA | 100.0 |
| Salmosin | ARGDDL | NPFHA | 130.0–270.0 |
| Triflavin | ARGDFP | WNGL | 100.0–400.0 |
| Rhodostomin | PRGDMP | YH | 108.0 |
| Protein | Sequence | IC50 (nM) | |||
|---|---|---|---|---|---|
| RGD Loop | C-Terminus | A375 | U373MG | Panc-1 | |
| Ech | 23ARGDDM | 42NPHKGPAT | 1.5 | 5.7 | 154.5 |
| Ech (K45E) | 23ARGDDM | 42NPHEGPAT | 13.3 | 27.2 | 1603.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.-C.; Chang, Y.-T.; Chen, C.-Y.; Shiu, J.-H.; Cheng, C.-H.; Huang, C.-H.; Chen, J.-F.; Chuang, W.-J. Structural Insight into Integrin Recognition and Anticancer Activity of Echistatin. Toxins 2020, 12, 709. https://doi.org/10.3390/toxins12110709
Chen Y-C, Chang Y-T, Chen C-Y, Shiu J-H, Cheng C-H, Huang C-H, Chen J-F, Chuang W-J. Structural Insight into Integrin Recognition and Anticancer Activity of Echistatin. Toxins. 2020; 12(11):709. https://doi.org/10.3390/toxins12110709
Chicago/Turabian StyleChen, Yi-Chun, Yao-Tsung Chang, Chiu-Yueh Chen, Jia-Hau Shiu, Chun-Ho Cheng, Chun-Hao Huang, Ju-Fei Chen, and Woei-Jer Chuang. 2020. "Structural Insight into Integrin Recognition and Anticancer Activity of Echistatin" Toxins 12, no. 11: 709. https://doi.org/10.3390/toxins12110709
APA StyleChen, Y. -C., Chang, Y. -T., Chen, C. -Y., Shiu, J. -H., Cheng, C. -H., Huang, C. -H., Chen, J. -F., & Chuang, W. -J. (2020). Structural Insight into Integrin Recognition and Anticancer Activity of Echistatin. Toxins, 12(11), 709. https://doi.org/10.3390/toxins12110709