Fungal Bioactive Anthraquinones and Analogues



Abstract
:1. Introduction
2. Fungal Anthraquinones and Analogues
3. Industrial Application of Anthraquinones
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Turner, W.B.; Aldridge, D.C. Fungal Metabolites II; Academic Press: London, UK, 1983. [Google Scholar]
- Osbourn, A.E.; Lanzotti, V. Plant-Derived Products; Springer: Dordrecht, Germany, 2009. [Google Scholar]
- Dewick, P.M. Medicinanal Natural Products—A Biosynthetic Approach; Wiley and Sons Ltd.: Chicester, UK, 2009. [Google Scholar]
- Locatelli, M. Anthraquinones: Analytical techniques as a novel tool to investigate on the triggering of biological targets. Curr. Drug Targets 2011, 12, 366–380. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liang, C.; Chen, Q.; Yang, Z. Rhein: A potential biological therapeutic drug for intervertebral disc degeneration. Med. Hypotheses 2011, 77, 1105–1107. [Google Scholar] [CrossRef] [PubMed]
- Mizushina, Y.; Nishiumi, S.; Nishida, M.; Yoshida, H.; Azuma, T.; Yoshida, M. Inhibition of repair-related DNA polymerases by vitamin Ks, their related quinone derivatives and associated inflammatory activity. Int. J. Oncol. 2013, 42, 793–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, W.; Bounda, G.A.; Yu, F. Pharmacological potential action of rhein and its diverse signal transduction-a systematic review. World J. Pharm. Res. 2014, 3, 3599–3626. [Google Scholar]
- Sultan, S.; Ghani, N.A.; Shah, S.A.A.; Ismail, N.H.; Noor, M.Z.; Naz, H. Microbial transformation of bioactive anthraquinones—A review. Biosci. Biotechnol. Res. Asia 2013, 10, 577–582. [Google Scholar] [CrossRef]
- Duval, J.; Pecher, V.; Poujol, M.; Lesellier, E. Research advances for the extraction, analysis and uses of anthraquinones: A review. Ind. Crops Prod. 2016, 94, 812–833. [Google Scholar] [CrossRef]
- Gessler, N.N.; Egorova, A.S.; Belozerskaya, T.A. Fungal anthraquinones. Appl. Biochem. Microbiol. 2013, 49, 85–99. [Google Scholar] [CrossRef]
- Fouillaud, M.; Venkatachalam, M.; Girard-Valenciennes, E.; Caro, Y.; Dufossé, L. Anthraquinones and derivatives from marine-derived fungi: Structural diversity and selected biological activities. Mar. Drugs 2016, 14, 64. [Google Scholar] [CrossRef] [Green Version]
- Rai, M.; Gade, A.; Zimowska, B.; Ingle, A.P.; Ingle, P. Marine-derived Phoma-the gold mine of bioactive compounds. Appl. Microbiol. Biotechnol. 2018, 102, 9053–9066. [Google Scholar] [CrossRef]
- Gallagher, R.; Hodges, R. The chemistry of dothistromin, a difuronanthraquinone from Dothistroma pini. Aust. J. Chem. 1972, 25, 2399–2407. [Google Scholar] [CrossRef]
- Danks, A.V.; Hodges, R. Polyhydroxyanthraquinones from Dothistroma pini. Aust. J. Chem. 1974, 27, 1603–1606. [Google Scholar] [CrossRef]
- Li, J.L.; Jiang, X.; Liu, X.; He, C.; Di, Y.; Lu, S.; Huang, H.; Linc, B.; Wangd, D.; Fan, B. Antibacterial anthraquinone dimers from marine derived fungus Aspergillus sp. Fitoterapia 2019, 133, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Nakaishima, S. Studies on the metabolites of phytotoxic fungi. I. Isolation of macrosporin and 6-methylxanthopurpurin 3-methyl ether from Alternaria bataticola Ikata ex Yamamoto. Chem. Pharm. Bull. 1973, 21, 2083–2085. [Google Scholar]
- Evidente, A.; Rodeva, R.; Andolfi, A.; Stoyanova, Z.; Perrone, C.; Motta, A. Phytotoxic polyketides produced by Phomopsis foeniculi, a strain isolated from diseased Bulgarian fennel. Eur. J. Plant Pathol. 2011, 130, 173–182. [Google Scholar] [CrossRef]
- Barash, I.; Karr, A.L.; Strobel, G.A. Isolation and characterization of stemphylin, a chromone glucoside from Stemphylium botryosum. Plant Physiol. 1975, 55, 646–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barash, I.; Netzer, D.; Nachmias, A.; Strobel, G.A. Differential effect of phytotoxins produced by Stemphylium botryosum on susceptible and resistant lettuce cultivars. Phytoparasitica 1978, 6, 95–98. [Google Scholar] [CrossRef]
- Assante, G.; Nasini, G. Identity of the phytotoxin stemphylin from Stemphylium botryosum with altersolanol A. Phytochemistry 1987, 26, 703–705. [Google Scholar] [CrossRef]
- Sakamura, S.; Ichihara, A.; Yoshihara, T. Biologically Active Natural Products; ACS Symposium Series; Cutler, H.G., Ed.; American Chemical Society: Washington, DC, USA, 1988. [Google Scholar]
- Sparapano, L.; Mairota, P.; Lerario, P. Phytotoxins and Plant Pathogenesis; Graniti, A., Durbin, R.D., Ballio, A., Eds.; Springer: Berlin/Heidelberg, Germany, 1989. [Google Scholar]
- Corsaro, M.M.; De Castro, C.; Evidente, A.; Lanzetta, R.; Molinaro, A.; Parrilli, M.; Sparapano, L. Phytotoxic extracellular polysaccharide fractions from Cryphonectria parasitica (Murr.) Barr strains. Carbohydr. Polym. 1998, 37, 167–172. [Google Scholar] [CrossRef]
- Calhoun, L.A.; Findlay, J.A.; Miller, J.D.; Whitney, N.J. Metabolites toxic to spruce budworm from balsam fir needle endophytes. Mycol. Res. 1992, 96, 281–286. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Rank, C.; Nielsen, K.F.; Larsen, T.O. Metabolomics of Aspergillus fumigatus. Med. Mycol. 2009, 47, S53–S71. [Google Scholar] [CrossRef]
- Wijesekara, I.; Zhang, C.; Van Ta, Q.; Vo, T.S.; Li, Y.X.; Kim, S.K. Physcion from marine-derived fungus Microsporum sp. induces apoptosis in human cervical carcinoma HeLa cells. Microbiol. Res. 2014, 169, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Thomson, R.H. Naturally Occurring Quinones; Academic Press: London, UK; New York, NY, USA, 1971. [Google Scholar]
- Engström, K.; Brishammar, S.; Svensson, C.; Bengtsson, M.; Andersson, R. Anthraquinones from some Drechslera species and Bipolaris sorokiniana. Mycol. Res. 1993, 97, 381–384. [Google Scholar] [CrossRef]
- Bouras, N.; Strelkov, S.E. The anthraquinone catenarin is phytotoxic and produced in leaves and kernels of wheat infected by Pyrenophora tritici-repentis. Physiol. Mol. Plant Pathol. 2008, 72, 87–95. [Google Scholar] [CrossRef]
- Brady, S.F.; Wagenaar, M.M.; Singh, M.P.; Janso, J.E.; Clardy, J. The cytosporones, new octaketide antibiotics isolated from an endophytic fungus. Org. Lett. 2000, 2, 4043–4046. [Google Scholar] [CrossRef]
- Brady, S.F.; Singh, M.P.; Janso, J.E.; Clardy, J. Cytoskyrins A and B, new BIA active bisanthraquinones isolated from an endophytic fungus. Org. Lett. 2000, 2, 4047–4049. [Google Scholar] [CrossRef] [PubMed]
- Elespuru, R.K.; Yarmolinsky, M.B. A colorimetric assay of lysogenic induction designed for screening potential carcinogenic and carcinostatic agents. Environ. Mutagen. 1979, 1, 65–78. [Google Scholar] [CrossRef]
- Elespuru, R.K. Environmental Physiology and Medicine, Short-Term Tests for Chemical Carcinogens; Stich, H., San, C., Eds.; Springer: New York, NY, USA, 1981; pp. 1–11. [Google Scholar]
- Elespuru, R.K.; Moore, S.G. Micro-BIA, a colorimetric microtiter assay of λ prophage induction. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 1986, 164, 31–40. [Google Scholar] [CrossRef]
- Tanaka, M.; Ohra, J.; Tsujtno, Y.; Fujimori, T.; Ago, H.; Tsuge, H.Z. Dendryol A, B, C, and D, four new compounds produced by a weed pathogenic fungus Dendryphiella sp. Naturforsch. C 1995, 50, 751–756. [Google Scholar] [CrossRef]
- Imaizumi, S.; Tokumasu, S.; Nitta, N.; Fujimori, T.; Yamada, M. A new disease of paddy weed Eleocharis kuroguwai caused by Dendryphiella sp. Ann. Phytopath. Soc. Jpn. 1992, 58, 158. [Google Scholar]
- Huss, H.; Mayerhofer, H.; Wetschnig, W. Ophiocladium hordei CAV. (Fungi imperfecti), ein für Östereich neuer parasitischer Pilz der Gerste. Der Pflanzenarzt 1987, 40, 11–13. [Google Scholar]
- Miethbauer, S.; Haase, S.; Schmidtke, K.U.; Günther, W.; Heiser, I.; Liebermann, B. Biosynthesis of photodynamically active rubellins and structure elucidation of new anthraquinone derivatives produced by Ramularia collo-cygni. Phytochemistry 2006, 67, 1206–1213. [Google Scholar] [CrossRef] [PubMed]
- Miethbauer, S.; Gaube, F.; Möllmann, U.; Dahse, H.M.; Schmidtke, M.; Gareis, M.; Pickhardt, M.; Liebermann, B. Antimicrobial, antiproliferative, cytotoxic, and tau inhibitory activity of rubellins and caeruleoramularin produced by the phytopathogenic fungus Ramularia collo-cygni. Planta Med. 2009, 75, 1523–1525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miethbauer, S.; Gunther, W.; Schmidtke, K.U.; Heiser, I.; Grafe, S.; Gitter, B.; Liebermann, B. Uredinorubellins and caeruleoramularin, photodynamically active anthraquinone derivatives produced by two species of the genus Ramularia. J. Nat. Prod. 2008, 71, 1371–1375. [Google Scholar] [CrossRef] [PubMed]
- Vinale, F.; Ghisalberti, E.L.; Sivasithamparam, K.; Marra, R.; Ritieni, A.; Ferracane, R.; Woo, S.; Lorito, M. Factors affecting the production of Trichoderma harzianum secondary metabolites during the interaction with different plant pathogens. Lett. Appl. Microbiol. 2009, 48, 705–711. [Google Scholar]
- Lee, Y.M.; Li, H.; Hong, J.; Cho, H.Y.; Bae, K.S.; Kim, M.A.; Kim, D.K.; Jung, J.H. Bioactive metabolites from the sponge-derived fungus Aspergillus versicolor. Arch. Pharm. Res. 2010, 33, 231–235. [Google Scholar] [CrossRef]
- Hawas, U.W.; El-Beih, A.A.; El-Halawany, A.M. Bioactive anthraquinones from endophytic fungus Aspergillus versicolor isolated from red sea algae. Arch. Pharm. Res. 2012, 35, 1749–1756. [Google Scholar] [CrossRef]
- Cimmino, A.; Masi, M.; Evidente, M.; Superchi, S.; Evidente, A. Fungal phytotoxins with potential herbicidal activity: Chemical and biological characterization. Nat. Prod. Rep. 2015, 32, 1629–1653. [Google Scholar] [CrossRef]
- Bick, I.R.; Rhee, C. Anthraquinone pigments from Phoma foveata Foister. Biochem. J. 1966, 98, 112–116. [Google Scholar] [CrossRef] [Green Version]
- Quereshi, S.; Khan, N.A.; Pandey, A.K. Anthraquinone pigment with herbicidal potential from Phoma herbarum FGCC# 54. Chem. Nat. Compd. 2011, 47, 521–523. [Google Scholar]
- Zheng, C.J.; Shao, C.L.; Guo, Z.Y.; Chen, J.F.; Deng, D.S.; Yang, K.L.; Chen, Y.Y.; Fu, X.M.; She, Z.G.; Lin, Y.C.; et al. Bioactive hydroanthraquinones and anthraquinone dimers from a soft coral-derived Alternaria sp. fungus. J. Nat. Prod. 2012, 75, 189–197. [Google Scholar] [CrossRef]
- Niu, C.; Cai, M.; Zhang, Y.; Zhou, X. Biosynthetic origin of the carbon skeleton of a novel anti-tumor compound, haloroquinone, from a marine-derived fungus, Halorosellinia sp. Biotechnol. Lett. 2012, 34, 2119–2124. [Google Scholar] [CrossRef] [PubMed]
- Isaka, M.; Palasarn, S.; Tobwor, P.; Boonruangprapa, T.; Tasanathai, K. Bioactive anthraquinone dimers from the leafhopper pathogenic fungus Torrubiella sp. BCC 28517. J. Antibiot. 2012, 65, 571–574. [Google Scholar] [CrossRef] [PubMed]
- Ayers, S.; Graf, T.N.; Adcock, A.F.; Kroll, D.J.; Shen, Q.; Swanson, S.M.; Matthew, S.; Carcache de Blanco, E.J.; Wani, M.C.; D’Arveaux, B.A.; et al. Cytotoxic xanthone–anthraquinone heterodimers from an unidentified fungus of the order Hypocreales (MSX 17022). J. Antibiot. 2012, 65, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Khamthong, N.; Rukachaisirikul, V.; Phongpaichit, S.; Preedanon, S.; Sakayaroj, J. Bioactive polyketides from the sea fan-derived fungus Penicillium citrinum PSU-F51. Tetrahedron 2012, 68, 8245–8250. [Google Scholar] [CrossRef]
- Zhou, X.M.; Zheng, C.J.; Chen, G.Y.; Song, X.P.; Han, C.R.; Li, G.; Li, G.N.; Fu, Y.H.; Chen, W.H.; Niu, Z.G. Bioactive anthraquinone derivatives from the mangrove-derived fungus Stemphylium sp. 33231. J. Nat. Prod. 2014, 77, 2021–2028. [Google Scholar] [CrossRef]
- Li, H.; Wei, J.; Pan, S.Y.; Gao, J.M.; Tian, J.M. Antifungal, phytotoxic and toxic metabolites produced by Penicillium purpurogenum. Nat. Prod. Res. 2014, 28, 2358–2361. [Google Scholar] [CrossRef]
- Shao, C.; Wang, C.; Wei, M.; Li, S.; She, Z.; Gu, Y.; Lin, Y. Structural and spectral assignments of six anthraquinone derivatives from the mangrove fungus (ZSUH-36). Magn. Reson. Chem. 2008, 46, 886–889. [Google Scholar] [CrossRef]
- Shao, C.; She, Z.; Guo, Z.; Peng, H.; Cai, X.; Zhou, S.; Gu, Y.; Lin, Y. 1H and 13C NMR assignments for two anthraquinones and two xanthones from the mangrove fungus (ZSUH-36). Magn. Reson. Chem. 2007, 45, 434–438. [Google Scholar] [CrossRef]
- Miao, F.P.; Li, X.D.; Liu, X.H.; Cichewicz, R.H.; Ji, N.Y. Secondary metabolites from an algicolous Aspergillus versicolor strain. Mar. Drugs 2012, 10, 131–139. [Google Scholar] [CrossRef]
- Gui, R.-Y.; Xu, L.; Kuang, Y.; Chuang, M., III; Qin, J.-C.; Liu, L.; Yang, S.-X.; Zhao, L.-C. Chaetominine, (+)-alantrypinone, questin, isorhodoptilometrin, and 4-hydroxybenzaldehyde produced by the endophytic fungus Aspergillus sp. YL-6 inhibit wheat (Triticum aestivum) and radish (Raphanus sativus) germination. J. Plant Interact. 2015, 10, 87–92. [Google Scholar]
- Kimura, Y.; Kozawa, M.; Baba, K.; Hata, K. New constitutents of roots of Polygonum cuspidatum. Planta Med. 1983, 48, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Tian, L.; Gu, Q.; Zhu, W. Secalonic acid D: A cytotoxic constituent from marine lichen-derived fungus Gliocladium sp. T31. Arch. Pharmacal. Res. 2006, 29, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Wiese, J.; Wenzel-Storjohann, A.; Malien, S.; Schmaljohann, R.; Imhoff, J.F. Engyodontochones, antibiotic polyketides from the marine fungus Engyodontium album strain LF069. Chem. Eur. J. 2016, 22, 7452–7462. [Google Scholar] [CrossRef]
- Huang, R.; Wang, T.; Xie, X.S.; Ma, K.X.; Fang, X.W.; Wu, S.H. Secondary metabolites from an endophytic fungus Nigrospora sp. Chem. Nat. Compd. 2016, 52, 697–699. [Google Scholar] [CrossRef]
- Zaidi, J.H.; Naeem, F.; Iqbal, R.; Choudhary, M.I.; Khan, K.M.; Perveen, S.; Shah, S.T.A.; Hayat, S.; Voelter, Z. Synthesis and bioactivities of naturally occurring anthraquinones: Isochrysophanol, isozyganein, ω-hydroxyisochrysophanol and morindaparvin. Naturforsch. B 2001, 56, 689–696. [Google Scholar] [CrossRef]
- Wang, W.; Liao, Y.; Tang, C.; Huang, X.; Luo, Z.; Chen, J.; Cai, P. Cytotoxic and antibacterial compounds from the coral-derived fungus Aspergillus tritici SP2-8-1. Mar. Drugs 2017, 15, 348. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.; Kim, S.; Li, W.; Bang, S.; Lee, H.; Lee, H.J.; Noh, R.Y.; Park, J.E.; Bang, W.Y.; Shim, S.H. Bioactive secondary metabolites produced by an endophytic fungus Gaeumannomyces sp. JS0464 from a maritime halophyte Phragmites communis. J. Antibiot. 2017, 70, 737–742. [Google Scholar] [CrossRef]
- Ayer, W.A.; Trifonov, L.S. Anthraquinones and a 10-hydroxyanthrone from Phialophora alba. J. Nat. Prod. 1994, 57, 317–319. [Google Scholar] [CrossRef]
- Gill, M.; Morgan, P.M. New fungal anthraquinones. Arkivoc 2001, 7, 145–156. [Google Scholar]
- Yang, Y.; Yan, Y.M.; Wei, W.; Luo, J.; Zhang, L.S.; Zhou, X.J.; Wanga, P.C.; Yang, Y.X.; Cheng, Y.X. Anthraquinone derivatives from Rumex plants and endophytic Aspergillus fumigatus and their effects on diabetic nephropathy. Bioorg. Med. Chem. Lett. 2013, 23, 3905–3909. [Google Scholar] [CrossRef] [Green Version]
- Na, M.; Jin, W.Y.; Min, B.S.; Ahn, J.S.; Bae, K.H. Protein tyrosine phosphatase 1B inhibitory activity of anthraquinones and stilbenes. Nat. Prod. Sci. 2008, 14, 143–146. [Google Scholar]
- Zhang, Y.; Jia, A.; Chen, H.; Wang, M.; Ding, G.; Sun, L.; Dai, M. Anthraquinones from the saline-alkali plant endophytic fungus Eurotium rubrum. J. Antibiot. 2017, 70, 1138–1141. [Google Scholar] [CrossRef] [PubMed]
- Anke, H.; Kolthoum, I.; Laatsch, H. Metabolic products of microorganisms. 192. The anthraquinones of the Aspergillus glaucus group. II. Biological activity. Arch. Microbiol. 1980, 126, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Huo, J.; Kurtán, T.; Mándi, A.; Antus, S.; Tang, H.; Draeger, S.; Schulz, B.; Hussain, H.; Khron, K.; et al. Structural and stereochemical studies of hydroxyanthraquinone derivatives from the endophytic fungus Coniothyrium sp. Chirality 2013, 25, 141–148. [Google Scholar] [CrossRef] [Green Version]
- Qin, S.D.; Wang, Y.; Wang, W.; Zhu, W.M. Anti-H1N1-virus secondary metabolites from mangrove-derived aciduric fungus Penicillium sp. OUCMDZ-4736. Chin. J. Mar. Drugs 2016, 35, 21–28. [Google Scholar]
- Jin, Y.; Qin, S.; Gao, H.; Zhu, G.; Wang, W.; Zhu, W.; Wang, Y. An anti-HBV anthraquinone from aciduric fungus Penicillium sp. OUCMDZ-4736 under low pH stress. Extremophiles 2018, 22, 39–45. [Google Scholar] [CrossRef]
- Anisha, C.; Sachidanandan, P.; Radhakrishnan, E.K. Endophytic Paraconiothyrium sp. from Zingiber officinale Rosc. displays broad-spectrum antimicrobial activity by production of danthron. Curr. Microbiol. 2018, 75, 343–352. [Google Scholar] [CrossRef]
- Awad, N.E.; Kassem, H.A.; Hamed, M.A.; El-Naggar, M.A.A.; El-Feky, A.M.M. Bioassays guided isolation of compounds from Chaetomium globosum. J. Mycol. Med. 2014, 24, e35–e42. [Google Scholar] [CrossRef]
- Lu, H.F.; Wang, H.L.; Chuang, Y.Y.; Tang, Y.J.; Yang, J.S.; Ma, Y.S.; Chiang, J.H.; Lu, C.C.; Yang, J.L.; Lai, T.Y.; et al. Danthron induced apoptosis through mitochondria- and caspase-3-dependent pathways in human brain Glioblastoma multiforms GBM 8401 cells. Neurochem. Res. 2009, 35, 390–398. [Google Scholar] [CrossRef]
- Khan, N.; Afroz, F.; Begum, M.N.; Rony, S.R.; Sharmin, S.; Moni, F.; Choudhury, M.; Hasand, K.; Shahaa, M.; Sohrab, M.H. Endophytic Fusarium solani: A rich source of cytotoxic and antimicrobial napthaquinone and aza-anthraquinone derivatives. Toxicol. Rep. 2018, 5, 970–976. [Google Scholar] [CrossRef]
- Masi, M.; Nocera, P.; Zonno, M.C.; Tuzi, A.; Pescitelli, G.; Cimmino, A.; Boari, A.; Infantino, A.; Vurro, M.; Evidente, A. Lentiquinones A, B, and C, phytotoxic anthraquinone derivatives isolated from Ascochyta lentis, a pathogen of lentil. J. Nat. Prod. 2018, 81, 2700–2709. [Google Scholar] [CrossRef] [PubMed]
- Andolfi, A.; Cimmino, A.; Villegas-Fernandez, A.M.; Tuzi, A.; Santini, A.; Melck, D.; Rubiales, D.; Evidente, A. Lentisone, a new phytotoxic anthraquinone produced by Ascochyta lentis, the causal agent of Ascochyta Blight in Lens culinaris. J. Agric. Food Chem. 2013, 61, 7301–7308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.; Wang, F.; Luo, M.; Chen, Y.; Song, Y.; Zhang, W.; Zhang, S.; Ju, J. Halogenated anthraquinones from the marine-derived fungus Aspergillus sp. SCSIO F063. J. Nat. Prod. 2012, 75, 1346–1352. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Zhang, X.; Zheng, H.; Zheng, Z.; Nong, X.; Liang, X.; Ma, X.; Qi, S. Novel anthraquinone derivatives as inhibitors of protein tyrosine phosphatases and indoleamine 2,3-dioxygenase 1 from the deep-sea derived fungus Alternaria tenuissima DFFSCS013. Org. Chem. Front. 2019, 6, 3252–3258. [Google Scholar] [CrossRef]
- Padhi, S.; Masi, M.; Cimmino, A.; Tuzi, A.; Jena, S.; Tayung, K.; Evidente, A. Funiculosone, a substituted dihydroxanthene-1,9-dione with two of its analogues produced by an endolichenic fungus Talaromyces funiculosus and their antimicrobial activity. Phytochemistry 2019, 157, 175–183. [Google Scholar] [CrossRef]
- Masi, M.; Reveglia, P.; Baaijens-Billones, R.; Górecki, M.; Pescitelli, G.; Savocchia, S.; Evidente, A. Phytotoxic metabolites from three Neofusicoccum species causal agents of Botryosphaeria dieback in australia, luteopyroxin, neoanthraquinone, and luteoxepinone, a disubstituted furo-α-pyrone, a hexasubstituted anthraquinone, and a trisubstituted oxepi-2-one from Neofusicoccum luteum. J. Nat. Prod. 2020, 83, 453–460. [Google Scholar]
- Bien, H.S.; Stawitz, J.; Wunderlich, K. Anthraquinone dyes and intermediates. In Ullmann’s Encyclopedia of Industrial Chemistry; Wiley-VCH Verlag: Weinheim, Germany, 2000. [Google Scholar]
- Vankar, P.S.; Shanker, R.; Mahanta, D.; Tiwari, S.C. Ecofriendly sonicator dyeing of cotton with Rubia cordifolia Linn using biomordant. Dyes Pigments 2008, 76, 207–212. [Google Scholar] [CrossRef]
- Akilandeswari, P.; Pradeep, B.V. Exploration of industrially important pigments from soil fungi. Appl. Microbiol. Biotechnol. 2016, 100, 1631–1643. [Google Scholar] [CrossRef]
- Guinot, P.; Rogé, A.; Gargadennec, A.; Garcia, M.; Dupont, D.; Lecoeur, E.; Candeliera, L.; Andary, C. Dyeing plants screening: An approach to combine past heritage and present development. Color Technol. 2006, 122, 93–101. [Google Scholar] [CrossRef]
- Moiseeva, Z.Z. Means for preparing and using nitro derivatives of anthraquinone in dye synthesis. Organicheskie Poluprodukty i Krasiteli 1969, 4, 70–79. (In Russian) [Google Scholar]
- Salabert, J.; Sebastián, R.M.; Vallribera, A. Anthraquinone dyes for superhydrophobic cotton. Chem. Commun. 2015, 51, 14251–14254. [Google Scholar] [CrossRef] [PubMed]
- Benkhaya, S.; Mrabet, S.; El Harfi, A. A review on classifications, recent synthesis and applications of textile dyes. Inorg. Chem. Commun. 2020, 115, 107891. [Google Scholar] [CrossRef]
- Ledakowicz, S.; Solecka, M.; Zylla, R. Biodegradation, decolourisation and detoxification of textile wastewater enhanced by advanced oxidation processes. J. Biotechnol. 2001, 89, 175–184. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Anthraquinone | Fungus | Biological Activity | Reference |
|---|---|---|---|
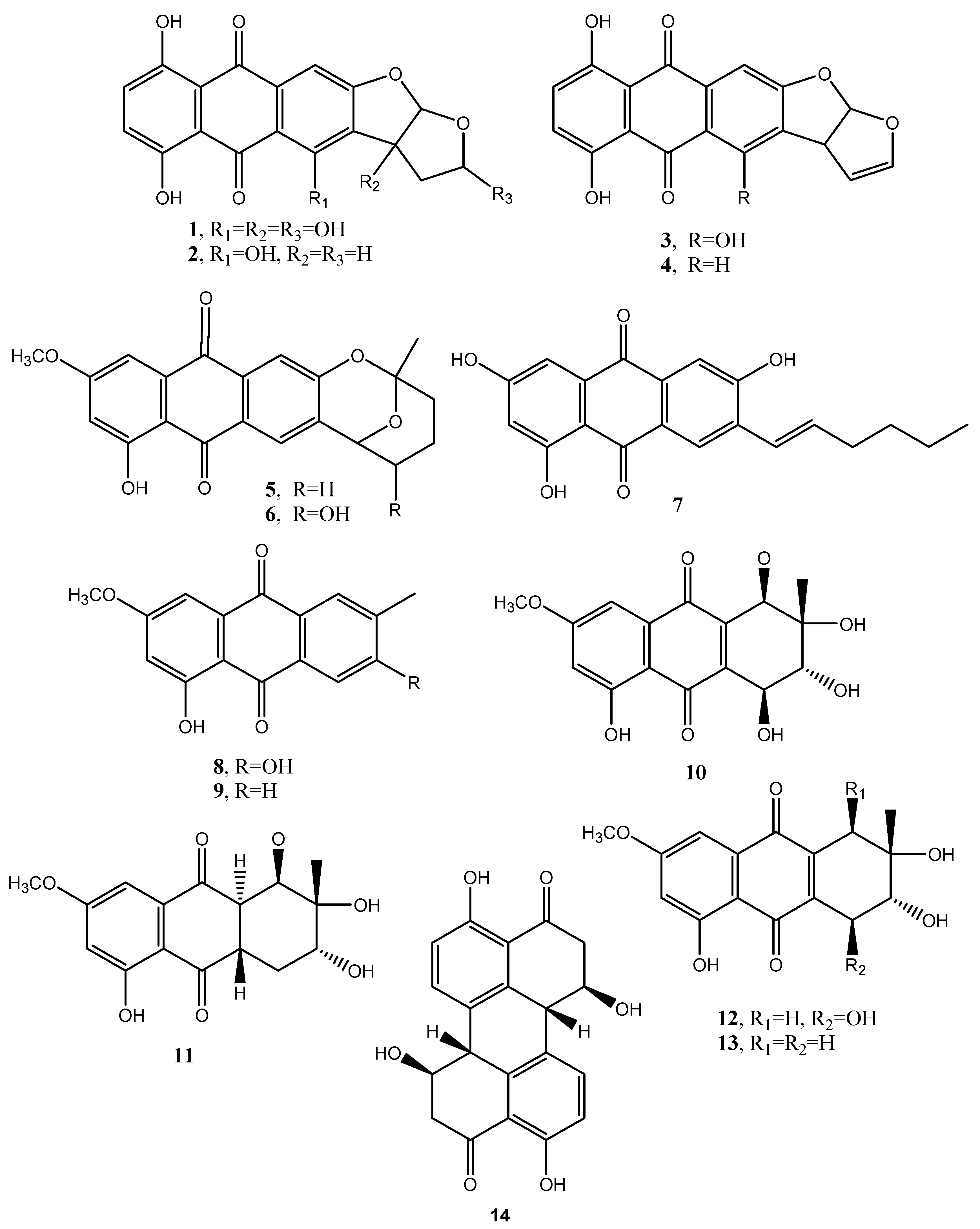
| Dothistromin (1, Figure 2) | Dothistroma pini | Phytotoxic | [13] |
| Bisdeoxydothistromin (2, Figure 2) | “ 1 | No activity | [14] |
| Bisdeoxydehydrodothistromin (3, Figure 2) | “ | “ | “ |
| 6-Deoxyversicolorin C (4, Figure 2) | “ | “ | “ |
| Averufin (5, Figure 2) | “ | “ | “ |
| Nidurufin (6, Figure 2) | “ | “ | “ |
| Averythrin (7, Figure 2) | Dothistroma pini Aspergillus versicolor | “ | “ [15] |
| Macrosporin (8, Figure 2) | Alternaria porri, Alternaria solani, Alternaria cucumerina, Diaporthe angelicae, Stemphyfium botryosum | Phytotoxic | [16,17,20] |
| 6-Methylxanthopurpurin 3 methyl eter (9, Figure 2) | Alternaria bataticola Alternaria solani | “ | “ |
| Alternasolanol A (10, Figure 2) | D. angelicae | ” | [17] |
| Alternasolanol J (11, Figure 2) | “ | “ | “ |
| Stemphylin (12, Figure 2) | S. botryosum | Phytotoxic and weak antitumor | [20] |
| Dactylariol (13, Figure 2) | “ | Phytotoxic and in vitro anticancer | “ |
| Stemphyperylenol (14, Figure 2) | Stemphylium botryosum | Weak phytotoxic | [21] |
| Alterporriol (15, Figure 3) | “ | No activity | “ |
| Stemphylenol A (16, Figure 3) | “ | “ | “ |
| Stemphylenol B (17, Figure 3) | “ | “ | “ |
| Rugulosin (18, Figure 3) | Cryphonectria parasitica Hormonema dematioides | Phytotoxic Insecticidal | [22,24] |
| Skyrin (19, Figure 3) | ” | Phytotoxic | ” |
| Crysophanol (20, Figure 3) | “ | “ | “ |
| Emodin (21, Figure 3) | C. parasitica, Pyrenophora tritici-repentis, Gliocladium sp. T31, Aspercgillus glaucus, H. dematioides, Aspergillus fumigatus, Phoma foevata | Phytotoxic Mycotoxic Anticancer and inhibition in vitro DNA-dependent RNA polymerase | [22,24,25,29,45,59,70] |
| 2-Chloroemodin (22, Figure 3) | Aspergillus fumigatus | No activity | [25] |
| Physcion (23, Figure 3) | “ Microsporum sp. | Anticancer | “ [26] |
| Catenarin (24, Figure 3) | Drechslera teres, Drechslera graminea, Drechslera tritici-repentis, Drechslera phlei, Drechslera dictyoides, Drechslera avenae Aspergillus cristatus | Antibiotic against Gram+ Phytotoxic Anticancer and inhibition in vitro DNA-dependent RNA polymerase | [28,29,70] |
| Helminthosporin (25, Figure 3) | D. avenae, Bipolaris sorokiniana | No activity | [28.29] |
| Cynodontin (26, Figure 3) | “ | No activity | “ |
| Cytoskyrin A (27, Figure 3) | Cytospora sp. CR200 | BIA | [30,31] |
| Cytoskyrin B (28, Figure 3) | “ | BIA | “ |
| Luteoskyrin (29, Figure 3) | “ | No activity | “ |
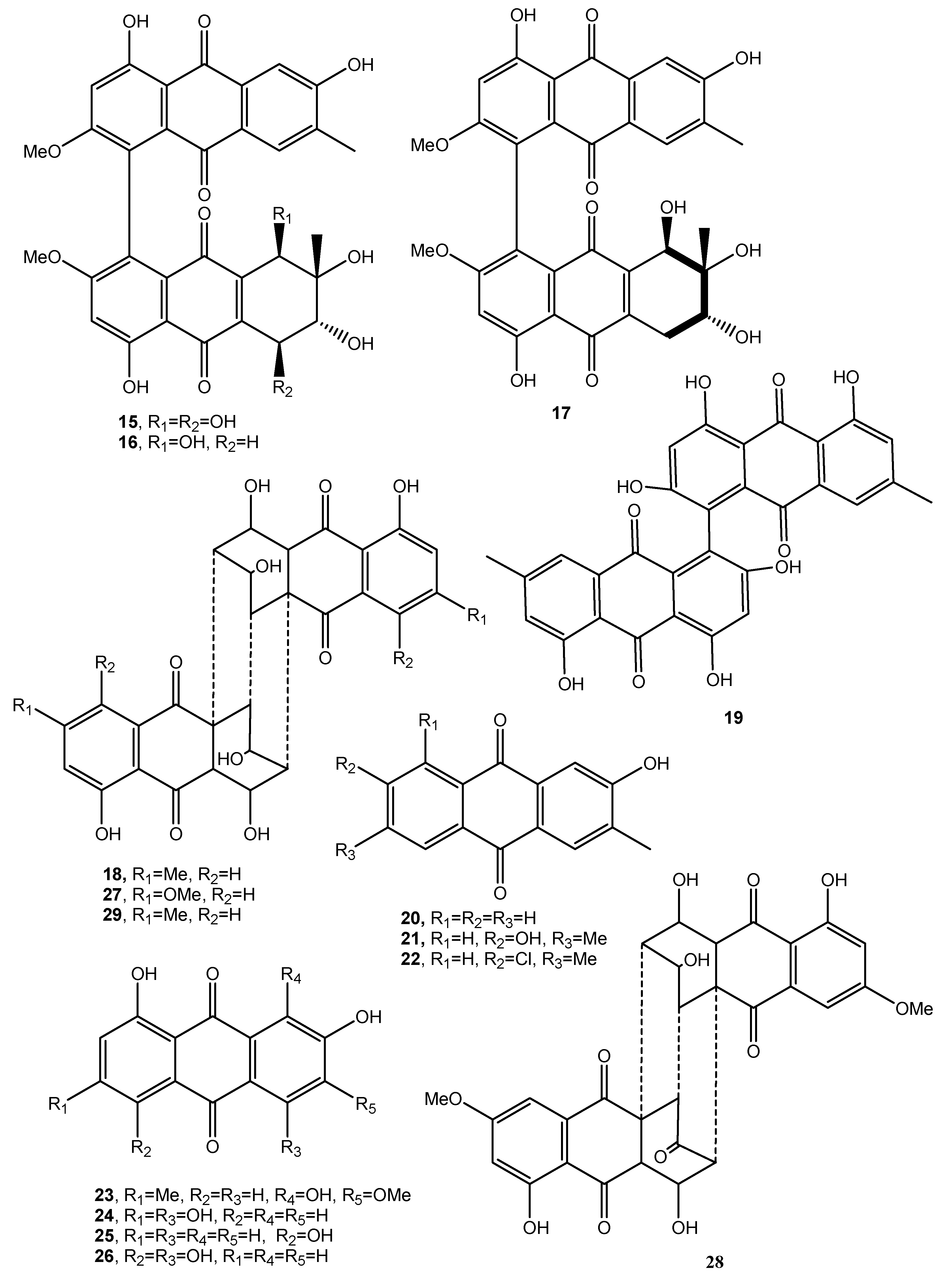
| Dendryol A (30, Figure 4) | Dendryphiella sp. | Phytotoxic | [35] |
| Dendryol B (31, Figure 4) | “ | “ | “ |
| Dendryol C (32, Figure 4) | “ | “ | “ |
| Dendryol D (33, Figure 4) | “ | “ | “ |
| Rubellin A (34, Figure 4) | Ramularia collo-cygni | Increased photodynamic oxygen activation | [37,38] |
| Rubellin B (35, Figure 4) | Phytotoxic, antibiotic, antiproliferative, and cytotoxic | [38,39] | |
| Rubellin C (36, Figure 4) | “ | “ | “ |
| Rubellin D (37, Figure 4) | “ | “ | “ |
| Rubellin E (38, Figure 4) | “ | “ | “ |
| Rubellin F (39, Figure 4) | “ | No activity | “ |
| Uridinetubellins I (40, Figure 4) | Ramularia uredinicola Ramularia collo-cygni | The photodynamic action toward three mammalian cell lines | [40] |
| Uridinetubellins II, (41, Figure 4) | “ | “ | “ |
| Caeruleoramularin (42, Figure 4) | “ | No activity | “ |
| 1-Hydroxy-3-methyl-anthraquinone (43, Figure 4) | Trichoderma harzianum | No activity | [41] |
| 1,8-Dihydroxy-3-methyl-anthraquinone (44, Figure 4) | “ | “ | “ |
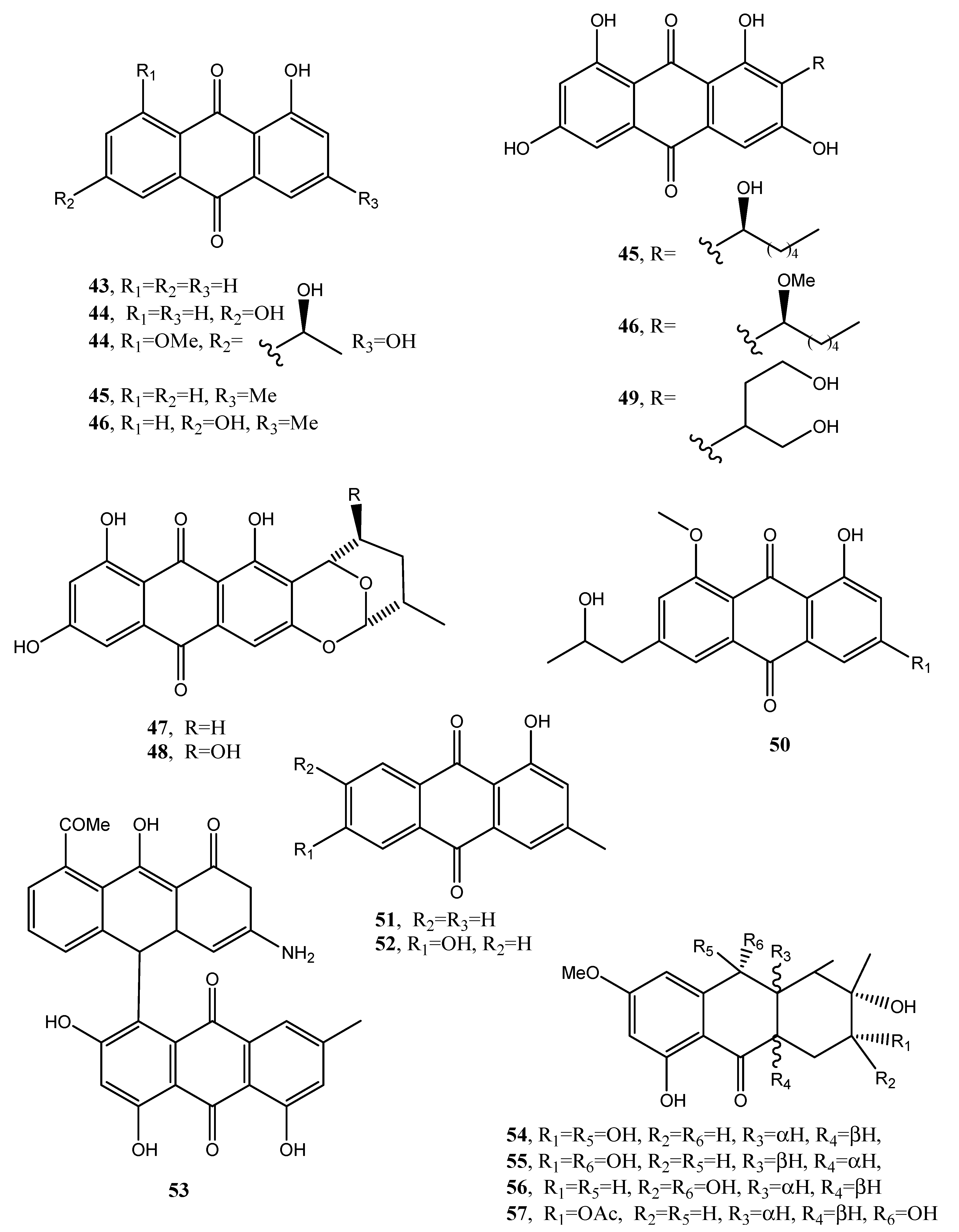
| Averantin (45, Figure 5) | Aspergillus versicolor | Antibiotic, Cytotoxic | [42,45,48] |
| Methyl-averantin (46, Figure 5) | “ | Cytotoxic | “ |
| Averufin (47, Figure 5) | “ | No activity | “ |
| Nidurufin (48, Figure 5) | “ | Antibiotic and cytotoxic | “ |
| Versiconol (49, Figure 5) | “ | No activity | “ |
| Isorhodoptilometrin-1-methyl ether (50, Figure 5) | “ | Antibiotic and mild anticancer | [43] |
| Pachybasin (51, Figure 5) | P. foevata, Coniothyrium sp., Ascochyta lentis | Weak antibiotic Antibiotic | [45,71,78] |
| Phomarin (52, Figure 5) | “ | Antifungal activity Antibiotic | [45,71,78] |
| Anhydropseudophlegmacin-9,10-quinone-3′-amino-8′-O-methyl ether (53, Figure 5) | Phoma herbarum | Phytotoxic | [46] |
| Tetrahydroaltersolanol C (54, Figure 5) | Alternaria sp. | Antiviral | [47] |
| Tetrahydroaltersolanol D (55, Figure 5) | “ | No activity | “ |
| Tetrahydroaltersolanol E (56, Figure 5) | “ | “ | “ |
| Tetrahydroaltersolanol F (57, Figure 5) | “ | “ | “ |
| Dihydroaltersolanol A (58, Figure 6) | “ | “ | “ |
| Alterporriol N (59, Figure 6) | “ | “ | “ |
| Alterporriol O (60, Figure 6) | “ | “ | “ |
| Alterporriol P (61, Figure 5) | “ | Cytotoxic | “ |
| Alterporriol Q (62, Figure 6) | “ | Antiviral | “ |
| Alterporriol R (63, Figure 6) | “ | No activity | “ |
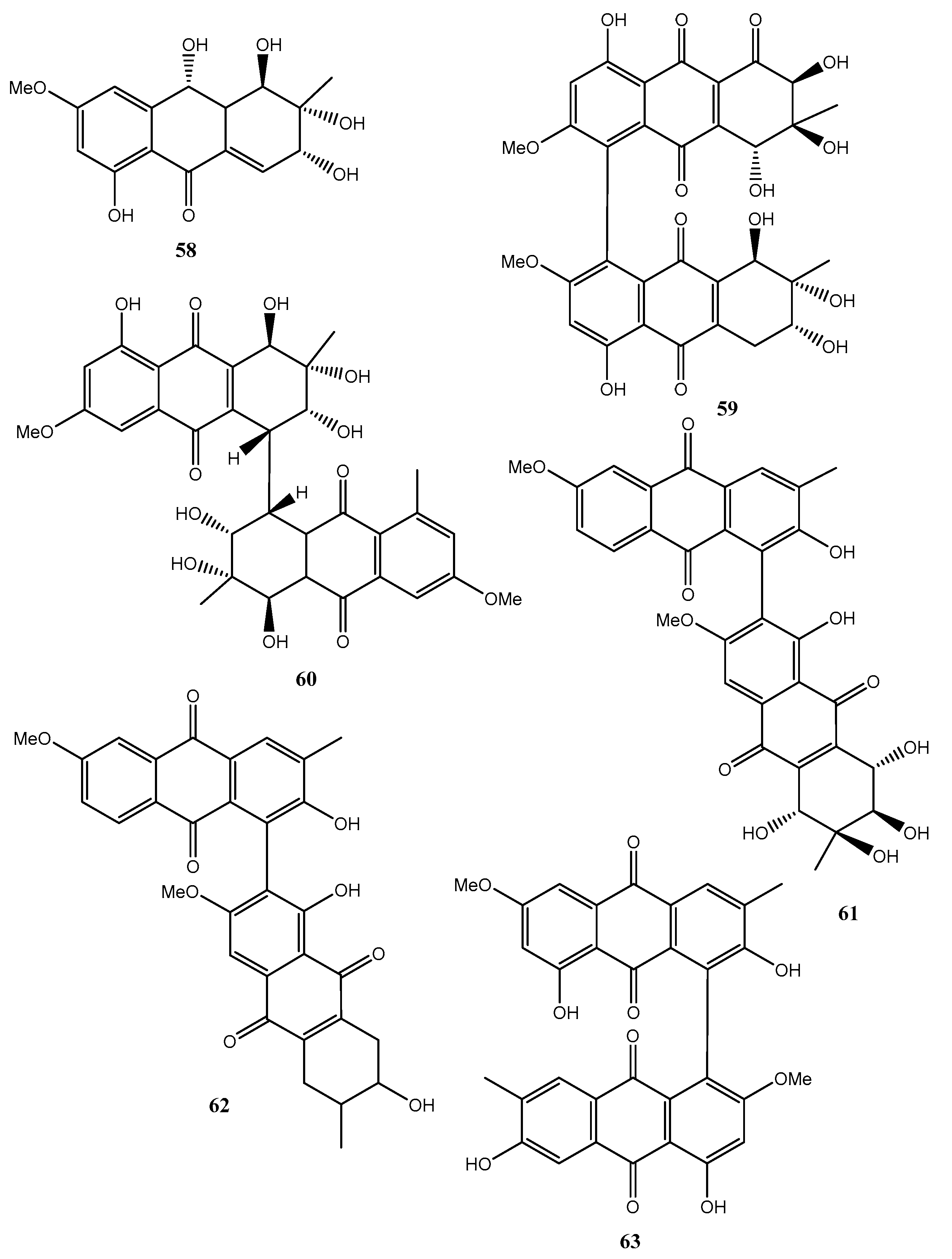
| Holoroquinone (64, Figure 7) | Halorosellinia sp. | Antitumor | [48] |
| Torrubiellin A (65, Figure 7) | Torrubiella sp. BCC 28517 | Moderate antimalarial, antifungal, antibacterial, cytotoxic | [49] |
| Torrubiellin A (66, Figure 7) | “ | Antimalarial, antifungal, antibacterial, cytotoxic | “ |
| Acremoxanthone C (67, Figure 7) | Hypocreales sp. MSX 17022 | Moderate cytotoxic | [50] |
| Acremoxanthone D (68, Figure 7) | “ | Moderate cytotoxic, and moderate 20S proteosome inhibition | “ |
| Penicillanthranin A (69, Figure 7) | Penicillium citrinum PSU-F51 | Moderate antibacterial and mild cytotoxic | [51] |
| Penicillanthranin B (70, Figure 7) | “ | No activity | “ |
| Auxarthrol C (71, Figure 7) | Stemphylium sp. 33231 | “ | [52] |
| Macrosporin 2-O-(6′-acetyl)-α-d-glucopyranoside (72, Figure 7) | “ | “ | “ |
| 2-O-Acetylaltersolanol B (73, Figure 8) | “ | “ | “ |
| 2-O-Acetylaltersolanol L (74, Figure 8) | “ | “ | |
| Alterporriols T (75 Figure 8) | “ | “ | “ |
| Alterporriols U (76 Figure 8) | “ | “ | “ |
| Alterporriols V (77 Figure 8) | “ | “ | “ |
| Alterporriols W (78 Figure 8) | “ | Weak antibacterial and moderate zootoxic | “ |
| 6,8,1′-Tri-O-methyl averantin (79, Figure 8) | Penicillium purpurogenum Endophytic fungus ZSUH-36 | Zootoxic and antifungal | [53,54] |
| 6,8-Di-O-methyl averufnin (80, Figure 8) | Penicillium purpurogenum Endophytic fungus ZSUH-36 Aspergillus versicolor | No activity | [53,54,56] |
| 6,8-Di-O-methyl averufanin (81, Figure 8) | “ | Antibiotic and zootoxic | [53,54,56] |
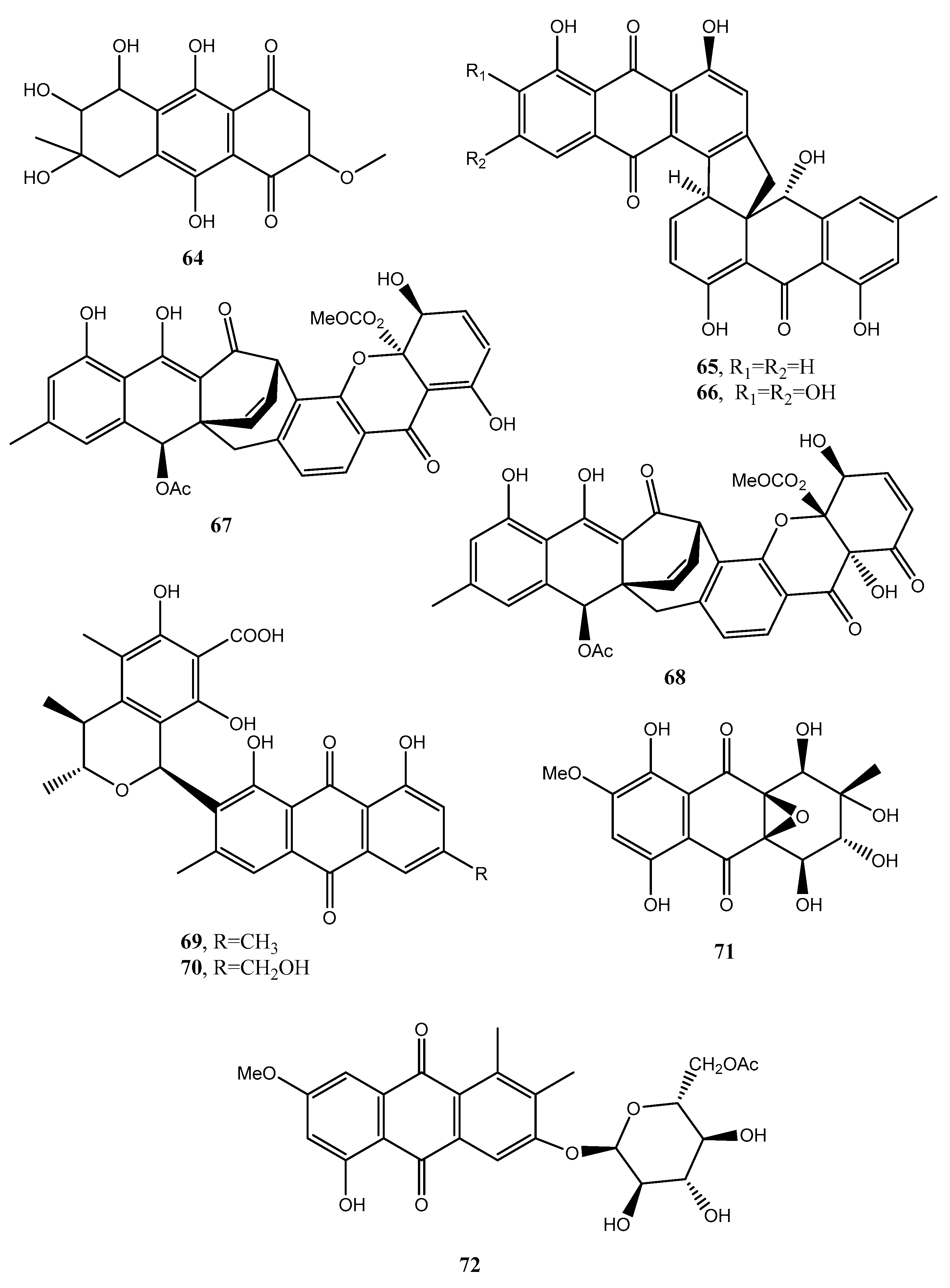
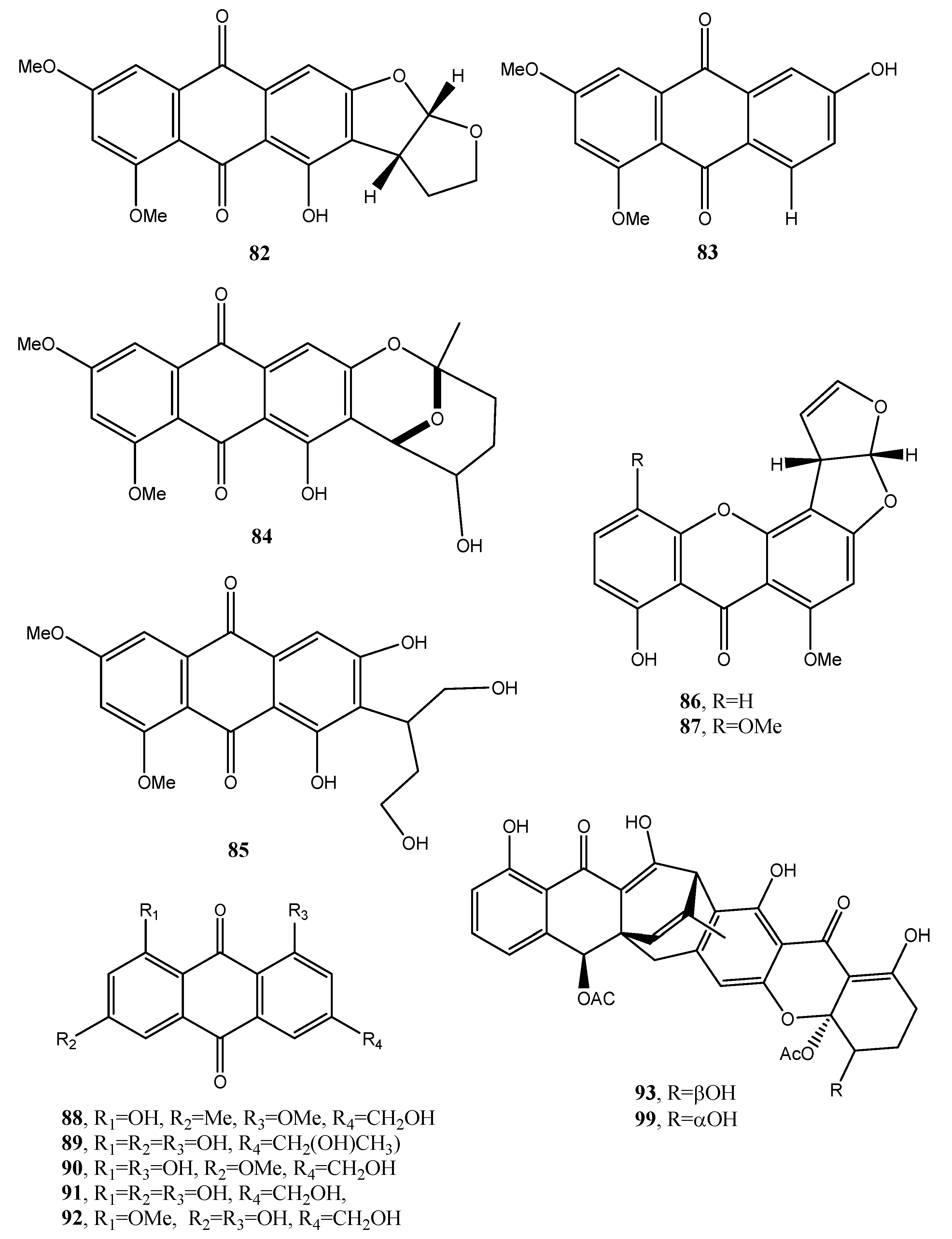
| Aversin (82, Figure 9) | “ | Antifungal | [53,55,56] |
| 1,3-Dihydroxy-6,8-dimethoxy-9,10-anthraquinone (83, Figure 9) | Penicillium purpurogenum | No activity | [53] |
| 6,8-Di-O-methylnidurufin (84, Figure 9) | Penicillium purpurogenum Endophytic fungus ZSUH-36 Aspergillus versicolor | Antifungal and phytotoxic | [53,54,56] |
| 6,8-Di-O-methyl versiconol (85, Figure 9) | Penicillium purpurogenum Endophytic fungus ZSUH-36 | Antifungal and phytotoxic | [53,55] |
| 5-Methyoxysterigmatocystin (86, Figure 9) | “ | Zootoxic | [53] |
| Sterigmatocystin (87, Figure 9) | Penicillium purpurogenum | No activity | [55] |
| Questin (88, Figure 9) | Aspergillus sp. YL-6, Polygonum cuspidatum | Allelopathy | [57,58] |
| Isorhodoptilometrin (89, Figure 9) | Aspergillus sp. YL-6, Gliocladium sp. T31 | Alleopathy | [57,59] |
| Fallacinol (90, Figure 9) | Polygonum cuspidatum | No activity | [58] |
| Citreorosein (91, Figure 9) | Polygonum cuspidatum Gliocladium sp. T31 | “ | [58,59] |
| Questinol (92, Figure 9) | Gliocladium sp. T31 | “ | [58] |
| Engyodontochone A (93, Figure 9) | Engyodontium album | Antibiotic | [60] |
| Engyodontochone B (94, Figure 10) | “ | No activity | “ |
| Engyodontochone C (95, Figure 10) | “ | Antibiotic | “ |
| Engyodontochone D (96, Figure 10) | “ | “ | “ |
| Engyodontochone E (97, Figure 10) | “ | “ | “ |
| Engyodontochone F (98, Figure 10) | “ | “ | “ |
| Betacolin-like compound (99, Figure 9) | “ | Antibiotic | “ |
| JBIR-99 (100, Figure 10) | “ | No activity | “ |
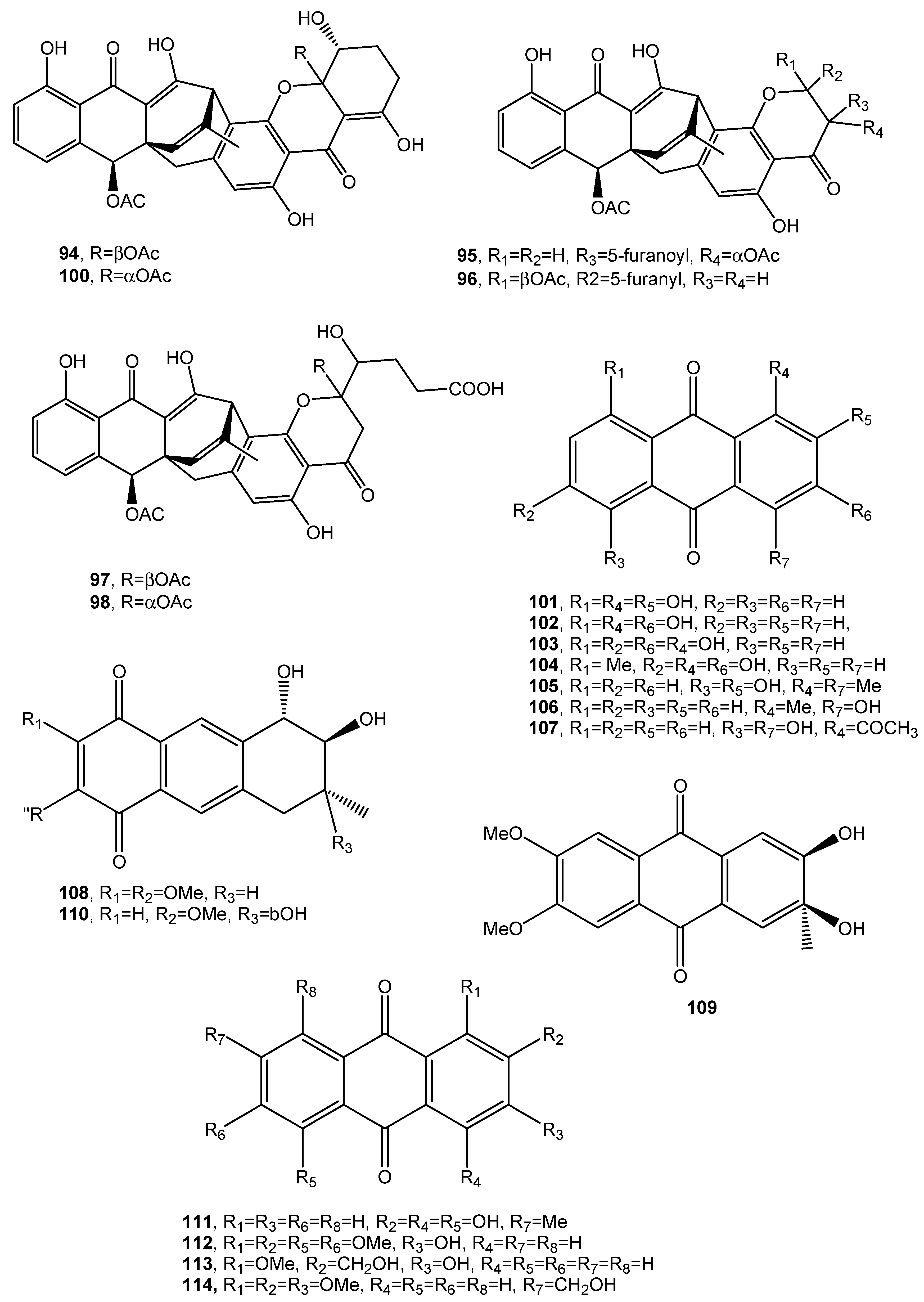
| 1,2,8-Trihydroxyanthraquinone (101, Figure 10) | Nigrospora sp. | “ | [61] |
| 1,3,8-Trihydroxyanthraquinone (102, Figure 10) | “ | Antifungal | “ |
| 1,3,6-trihydroxy-8-methylanthraquinone (103, Figure 10) | “ | “ | “ |
| Rheoemodin (104, Figure 10 | “ | Antimicrobial | “ |
| Aloesaponarin II (105, Figure 10) | “ | Antifungal | “ |
| Isozyganein (106, Figure 10) | “ | Antioxidant | “ |
| 1-Acetyl-4,5-dihydroxy-anthraquinone (107, Figure 10) | “ | No activity | |
| Aspetritone A (108, Figure 10) | Aspergillus tritici | Strong antibiotic and cytotoxic activity | [63] |
| Aspetritone B (109, Figure 10) | “ | No activity | “ |
| Bostrocyn (110, Figure 10) | “ | “ | “ |
| Compound 111 (Figure 10) | “ | “ | “ |
| Compound 112 (Figure 10) | “ | “ | “ |
| Compound 113 (Figure 10) | “ | “ | “ |
| Compound 114 (Figure 11) | “ | “ | “ |
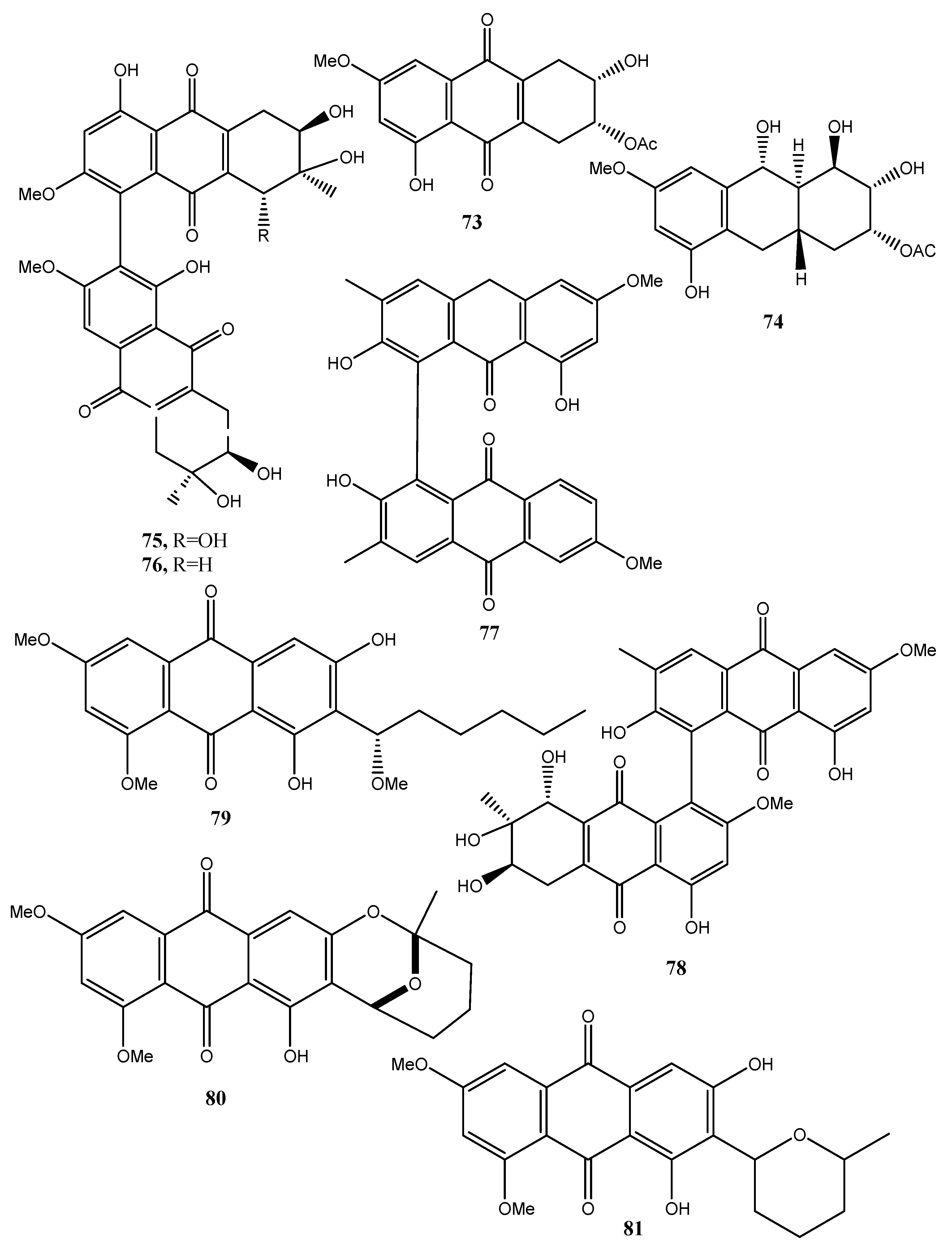
| 1-O-methyl-6-O-(α-d-ribofuranosyl)-emodin (115, Figure 11) | Gaeumannomyces sp. | Anti-inflammatory Reduction of NO production by LPS- | [64] |
| 1-O-Methylemodin (116, Figure 11) | Gaeumannomyces sp. Phialophora alba | Anti-inflammatory Reduction of NO production by LPS Growth inhibition of Phellinus tremulae Inhibition of the secretion of IL-625 Protein tyrosine phosphatase 1B inhibition | [64,65,66,67,68] |
| 5-Chloro-6,8-dihydroxy-1-methoxy-3-methylanthraquinone (117, Figure 11) | Phialophora alba | No activity | [65] |
| 7-Chloro-6,8-dihydroxy-1-methoxy-3-methylanthraquinone (118, Figure 11) | “ | “ | “ |
| 5-Chloro-6,8,10-trihydroxy-1-methoxy-3-methyl-9(10H) anthracenone (119, Figure 11) | “ | “ | “ |
| 5-chloro-8,10-dihydroxy-l,6-dimethoxy-3-metbyl-9(10H)-anthracenone (120, Figure 11) | “ | “ | “ |
| Rubrumol (121, Figure 11) | Eurotium rubrum | Activity when tested on Topo I | [69] |
| Rubrocristin (122, Figure 11) | “ Aspergillus glaucus | No Activity | “ |
| 2-Methyleurotinone (123, Figure 11) | Eurotium rubrum | “ | “ |
| Conyothyrinone A (124, Figure 11) | Eurotium rubrum Coniothyrium sp. | Antifungal activity | [69,70,71] |
| Erythroglaucin (125, Figure 11) | Aspergillus glaucus | No activity | [70] |
| Physcion-9-anthrone (126, Figure 11) | “ | “ | “ |
| Viocristin (127 Figure 11) | “ | Antibacterial activity Anticancer activity | “ |
| Isoviocristin (128, Figure 11) | “ | Antibacterial activity | “ |
| Conyothyrinone B (129, Figure 11) | Coniothyrium sp. | Antimicrobial activity | [71] |
| Conyothyrinone C (130, Figure 11) | “ | “ | “ |
| Conyothyrinone D (131, Figure 11) | “ | “ | “ |
| 1,7-Dihydroxy-3-methyl-9,10-anthraquinone (132, Figure 11) | “ | Antimicrobial activity and strong antibacterial activity | “ |
| 1-Hydroxy-3-hydroxymethyl-9,10-anthraquinone (133, Figure 11) | “ | Antimicrobial | “ |
| (–)-2′R-1-hydroxyisorhodoptilometrin (134, Figure 11) | Penicillium sp. OUCMDZ-4736 | Anti-hepatitis B virus | [72,73] |
| Methyl 3,4,8-trihydroxy-6-methyl-9-oxo-9H-xanthene-1-carboxylate. (135, Figure 11) | “ | No activity | [72] |
| Methyl 6,8-dihydroxy-3-methyl-9-oxo-9H-xanthene-1-carboxylate (136, Figure 11) | “ | “ | “ |
| Danthron (137, Figure 11) | Paraconiothyrium sp. | Antibacterial, antifungal and anticancer | [74] |
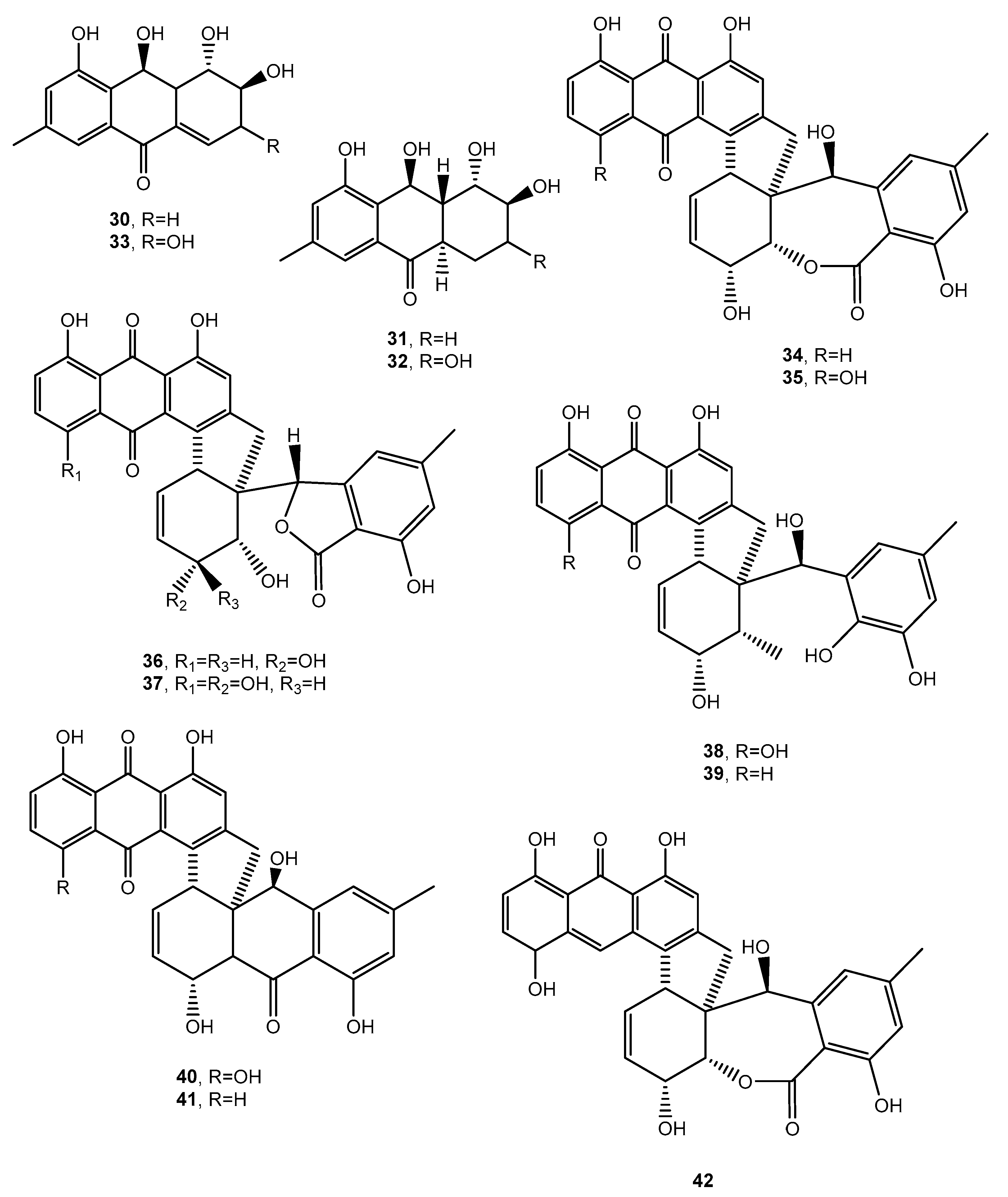
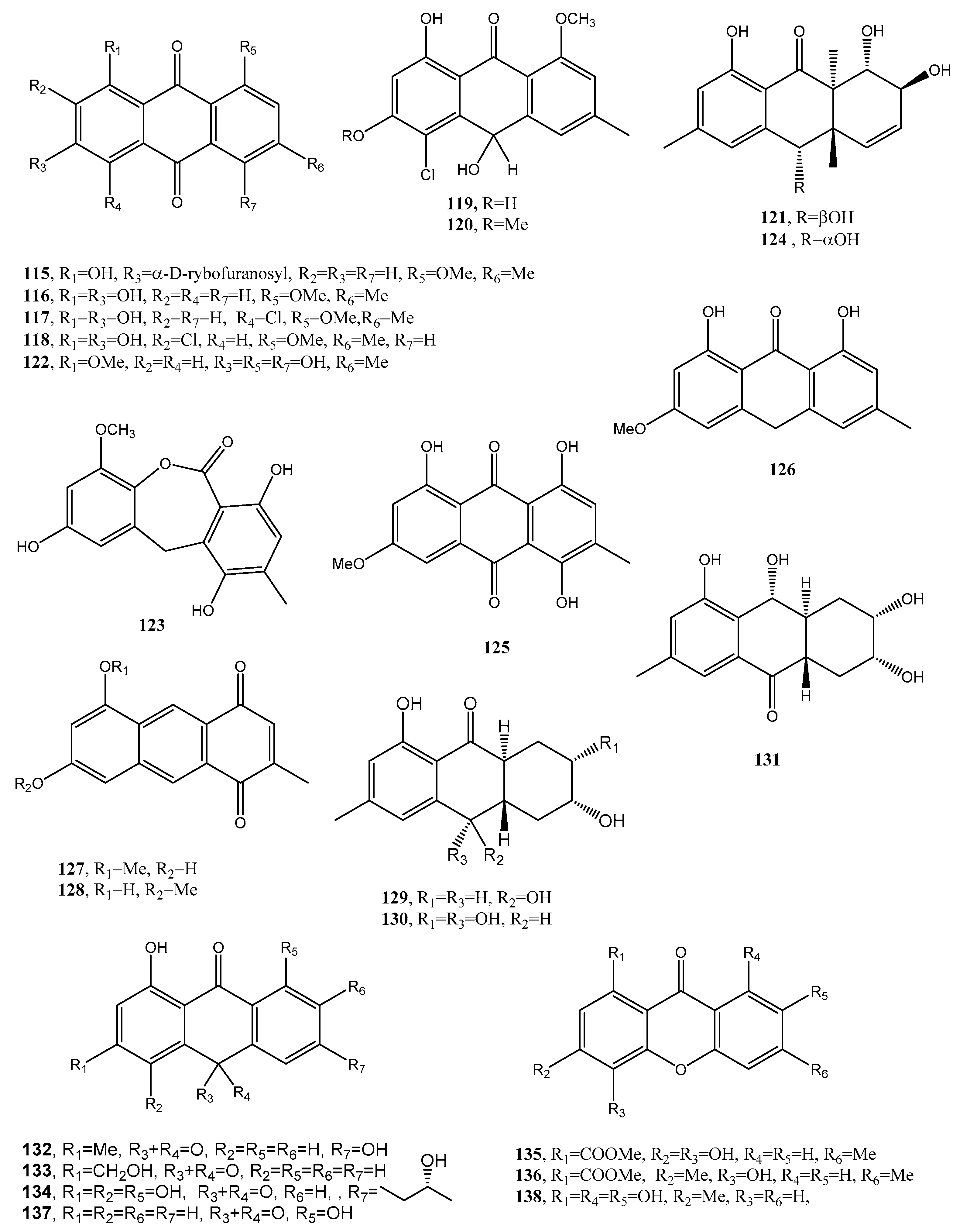
| Bostrycoidin (138, Figure 12) | Fusarium solani | Antimicrobial and anticancer | [77] |
| Lentiquinones A (139, Figure 12) | Ascochyta lentis | Phytotoxic and antimicrobial | [78] |
| Lentiquinones B (140, Figure 12) | “ | “ | “ |
| Lentiquinones C (141, Figure 12) | “ | “ | “ |
| Lentisone (142, Figure 12) | “ | “ | “ |
| ω-Hydroxypachybasin (143, Figure 12 | “ | “ | “ |
| 1,7-Dihydroxy-3-methylanthracene-9,10-dione (144, Figure 12) | “ | “ | “ |
| Anthraquinone dimer (145, Figure 12) | Aspergillus versicolor | Selective antibacterial | [15] |
| Anthraquinone dimer (146, Figure 12) | “ | “ | “ |
| 1′-O-Methylaverantin (147, Figure 12) | “ | No activity | “ |
| Averantin (148, Figure 12) | “ Aspergillus sp. SCSIO F063 | “ | “ [80] |
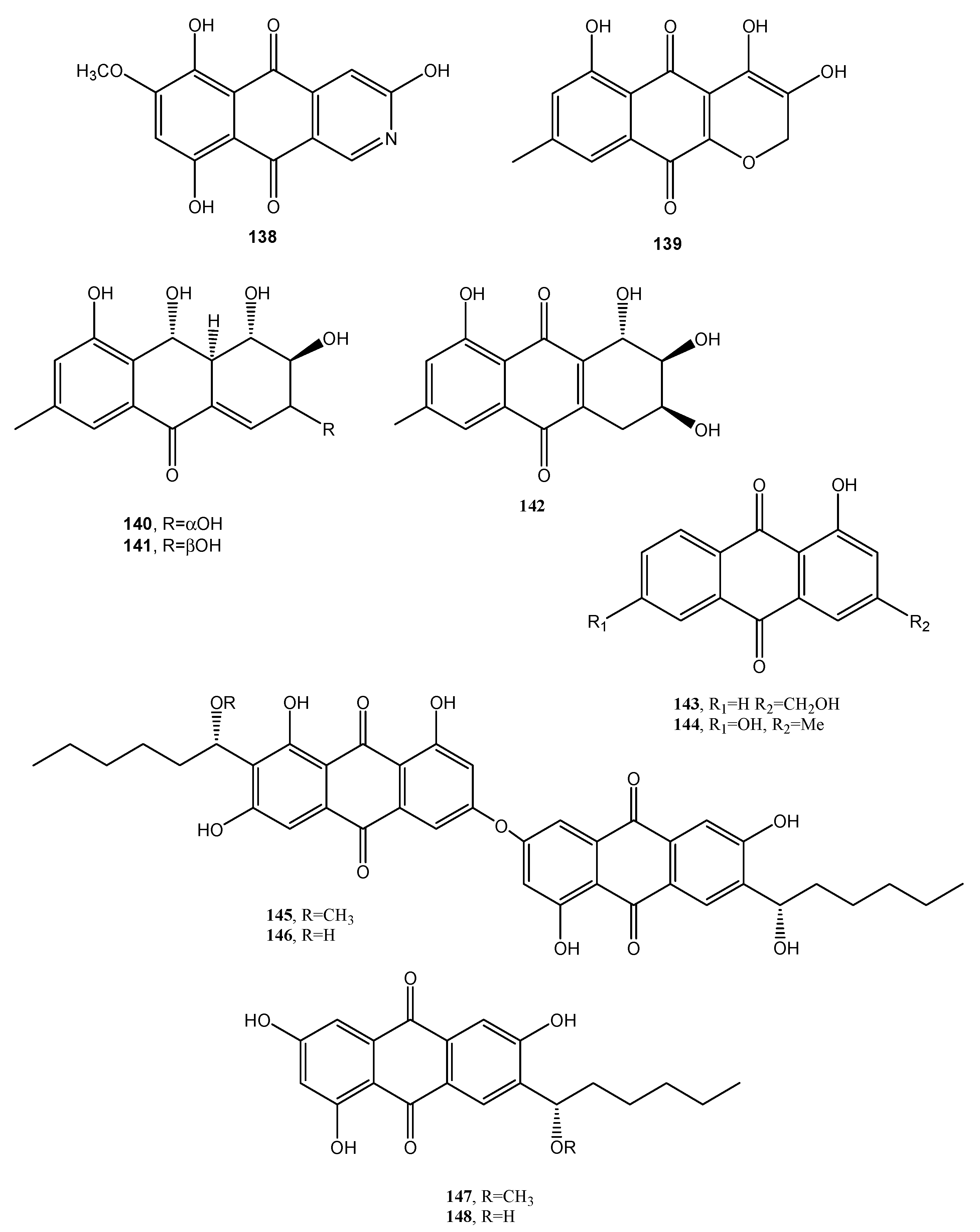
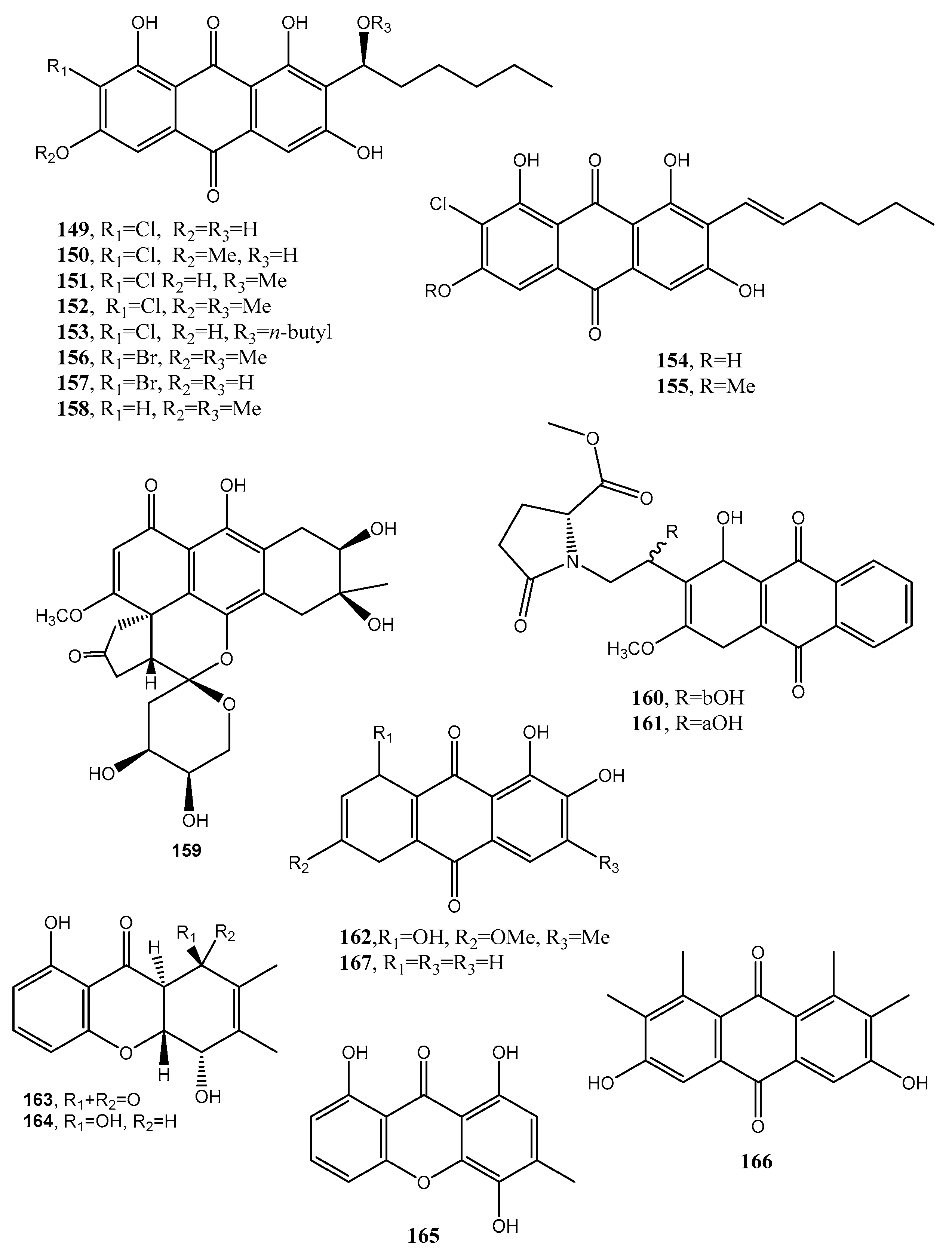
| (1′S)-7-Chloroaverantin (149, Figure 13) | Aspergillus sp. SCSIO F063 | “ | [80] |
| (1′S)-6-O-Methyl-7-chloroaverantin (150 Figure 13) | “ | “ | |
| (1′S)-1′-O-Methyl-7-chloroaverantin (151, Figure 13) | “ | “ | “ |
| (1′S)-6,1′-O,O-Dimethyl-7-chloroaverantin (152, Figure 13) | “ | “ | “ |
| (1′S)-7-Chloroaverantin-1′-butyl ether (153, Figure 13) | “ | “ | “ |
| 7-Chloroaverythrin (154, Figure 13) | “ | “ | “ |
| 6-O-Methyl-7-chloroaverythrin (155, Figure 13) | “ | Anticancer | “ |
| (1′S)-6,1′-O,O-Dimethyl-7-bromoaverantin (156, Figure 13) | “ | No activity | “ |
| and (1′S)-6-O-Methyl-7-bromoaverantinone (157, Figure 13) | “ | “ | “ |
| (1′S)-6,1′-O,O-Dimethylaverantin (158, Figure 13) | “ | “ | “ |
| Anthrininone A (159, Figure 13) | Alternaria tenuissima | Inhibition activity against indoleamine 2,3-dioxygenase and stimulate intracellular calcium levels | [81] |
| Anthrininone B (160, Figure 13) | “ | Inhibition activity against indoleamine 2,3-dioxygenase and against different protein tyrosine phosphatases | “ |
| Anthrininone C (161, Figure 13) | “ | “ | “ |
| 6-O-Methylalaternin (162, Figure 12 | “ | “ | “ |
| Funiculosone (163, Figure 13) | Talaromyces funiculosus | Antimicrobial | [82] |
| Mangrovamide J (164 Figure 13) | “ | “ | “ |
| Ravenelin (165 Figure 13) | “ | “ | “ |
| Neoanthraquinone (166, Figure 13) | Neofusicoccum luteum | Phytotoxic | [83] |
| Alizarin (167, Figure 13) | Rubia tinctorum | Dye | [84] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masi, M.; Evidente, A. Fungal Bioactive Anthraquinones and Analogues. Toxins 2020, 12, 714. https://doi.org/10.3390/toxins12110714
Masi M, Evidente A. Fungal Bioactive Anthraquinones and Analogues. Toxins. 2020; 12(11):714. https://doi.org/10.3390/toxins12110714
Chicago/Turabian StyleMasi, Marco, and Antonio Evidente. 2020. "Fungal Bioactive Anthraquinones and Analogues" Toxins 12, no. 11: 714. https://doi.org/10.3390/toxins12110714
APA StyleMasi, M., & Evidente, A. (2020). Fungal Bioactive Anthraquinones and Analogues. Toxins, 12(11), 714. https://doi.org/10.3390/toxins12110714