Microcystin-LR Does Not Alter Cell Survival and Intracellular Signaling in Human Bronchial Epithelial Cells



Abstract
:1. Introduction
2. Results
2.1. MC-LR Uptake and The Expression of Genes Encoding OATPs
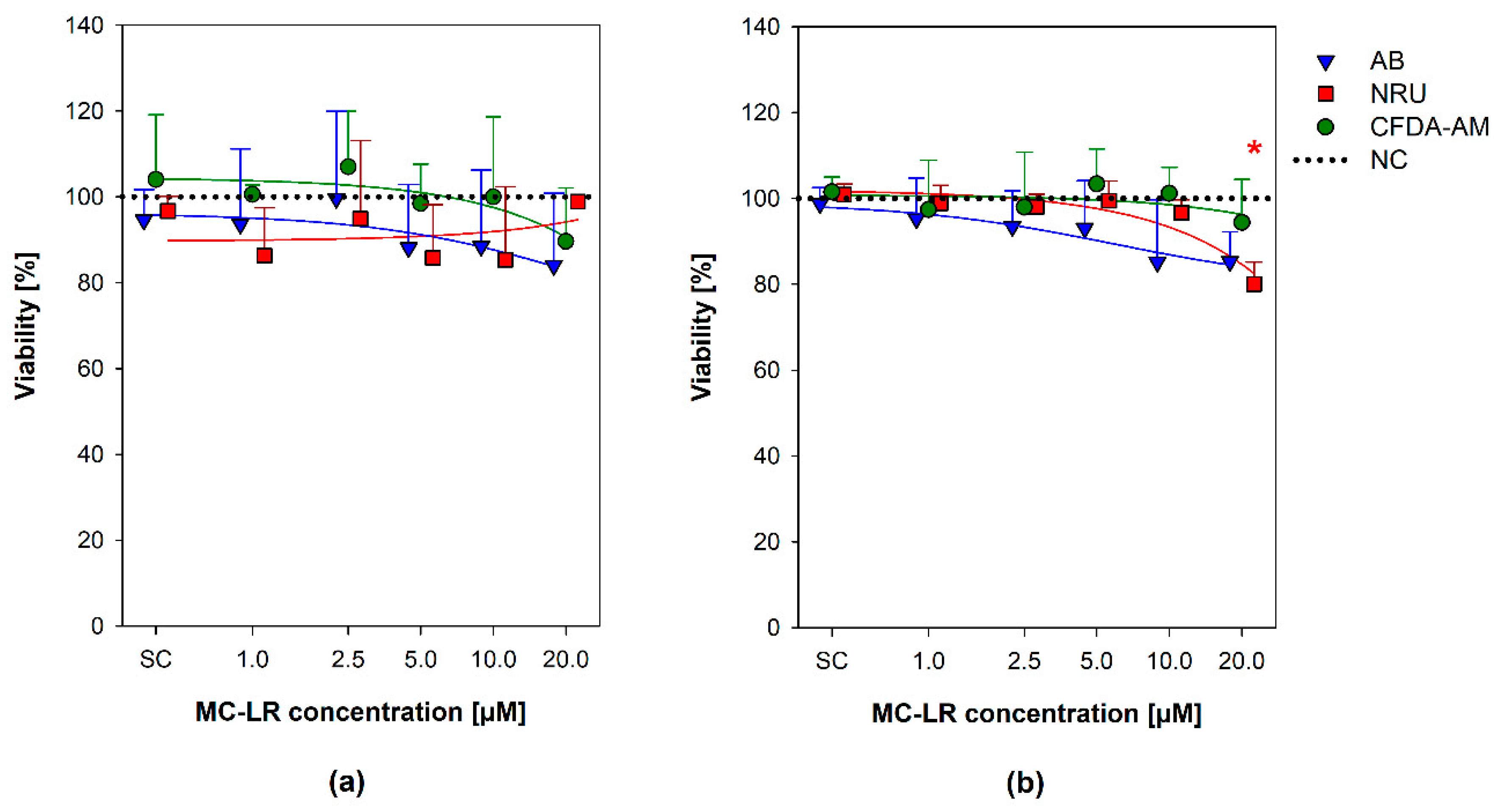
2.2. Viability Assays
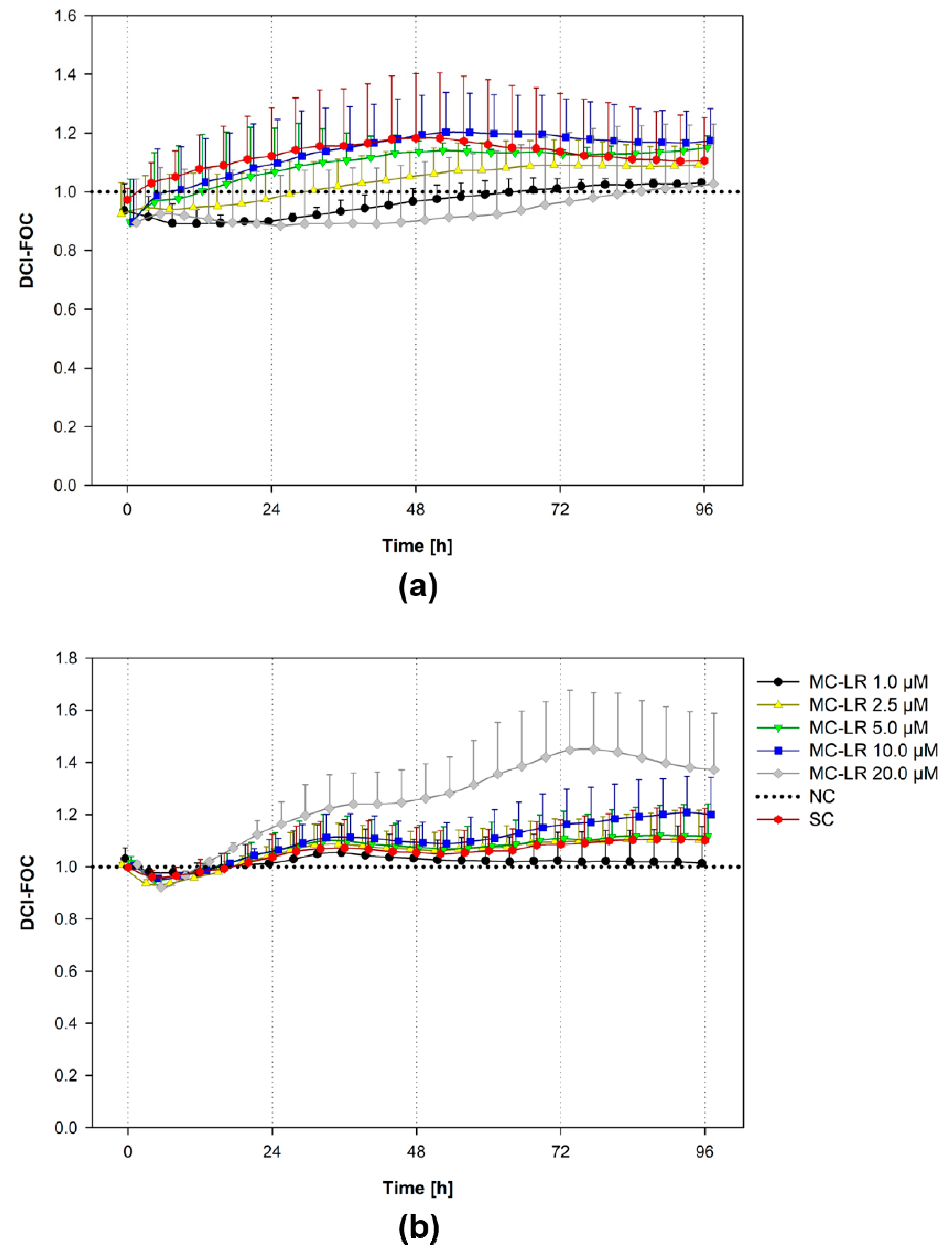
2.3. Real-Time Cell Analysis
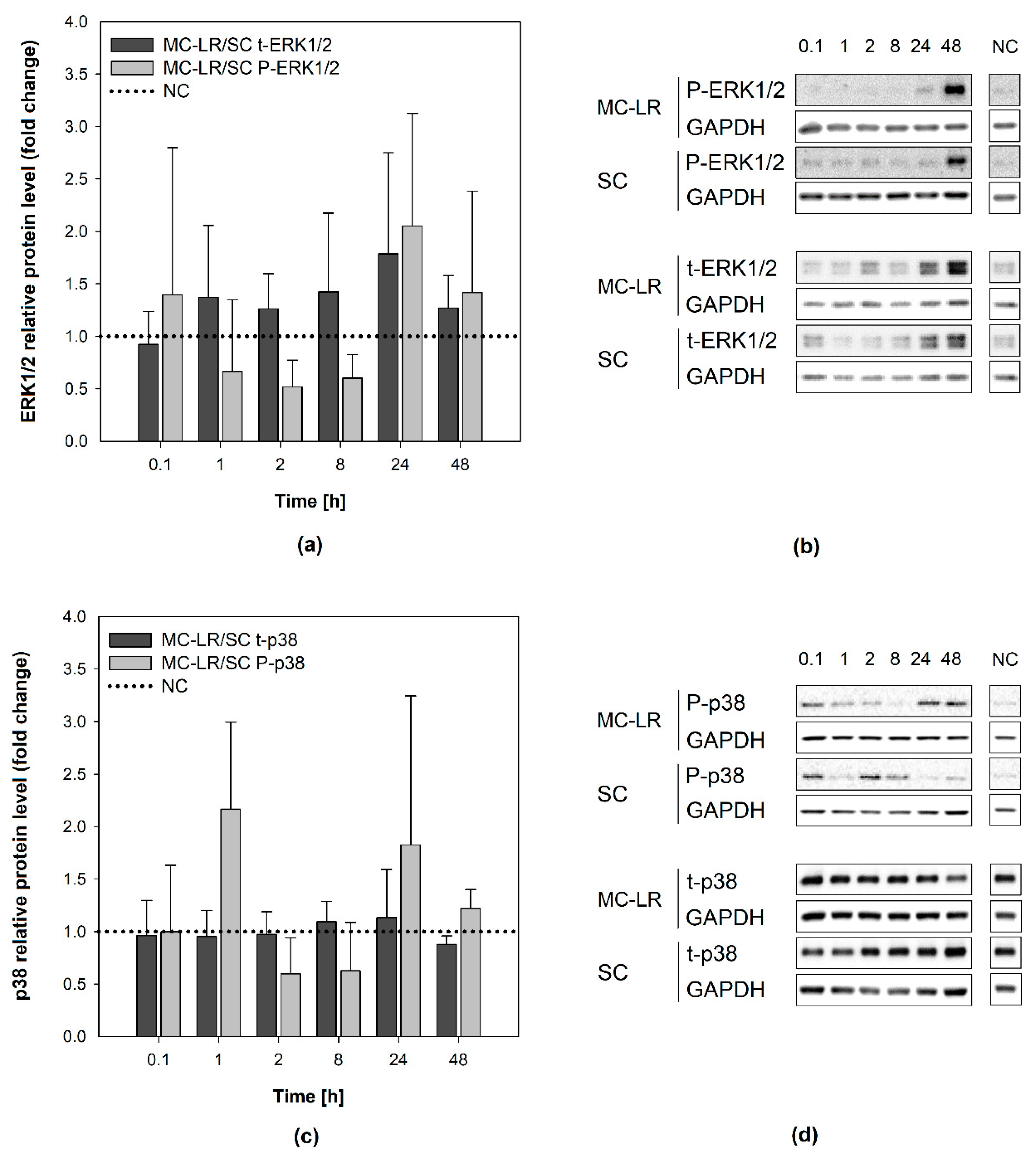
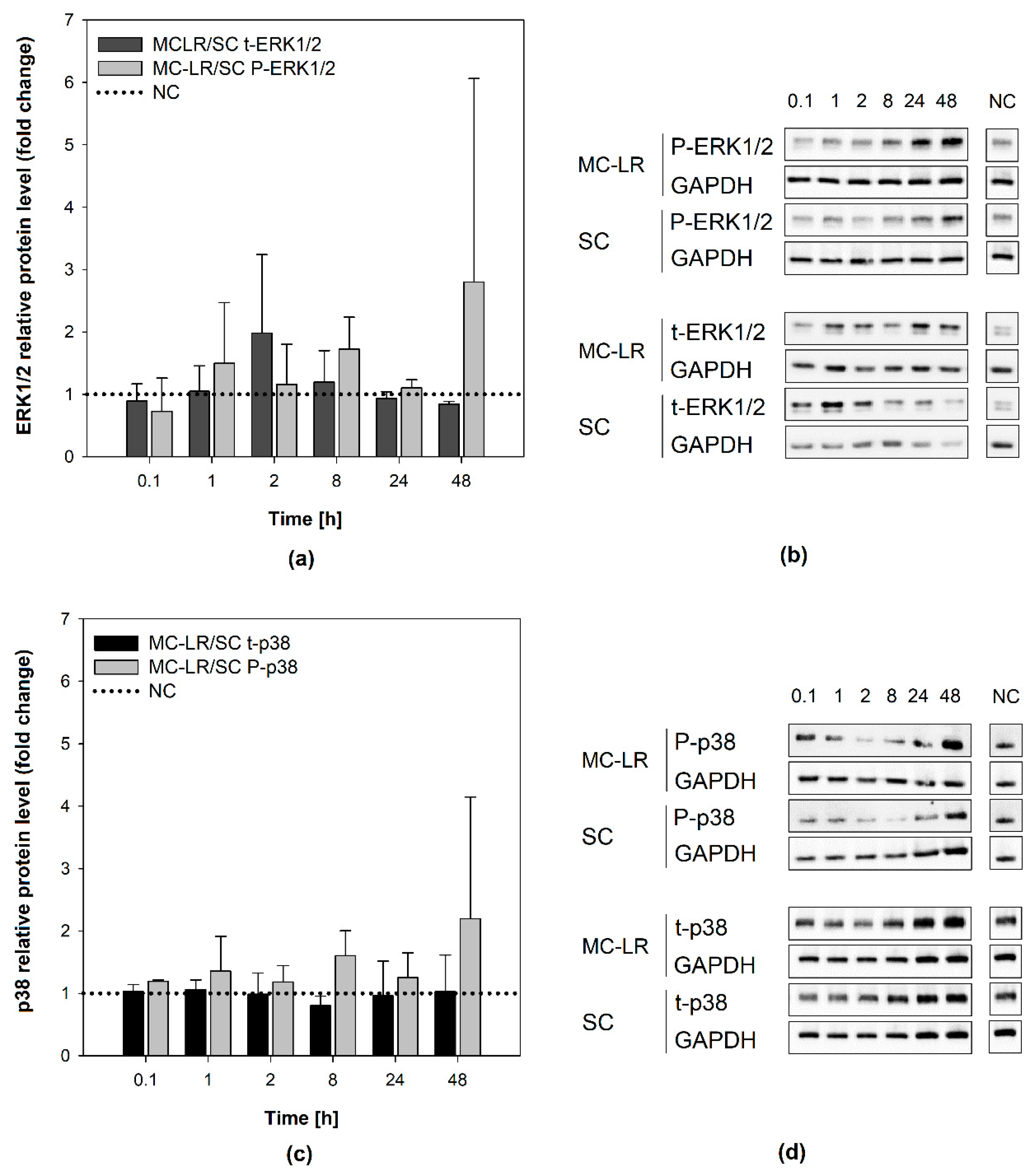
2.4. Evaluation of MAPKs Activity
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Cell Cultures
5.2. Experimental Design
5.3. Impedimetric Real-Time Cell Analysis (RTCA)
5.4. Cytotoxicity Assays
5.5. Reverse-Transcription Polymerase Chain Reaction (RT-PCR)
5.6. Western Blotting
5.7. Data Evaluation and Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lee, J.; Lee, S.; Jiang, X. Cyanobacterial toxins in freshwater and food: Important sources of exposure to humans. Annu. Rev. Food Sci. Technol. 2017, 8, 281–304. [Google Scholar] [CrossRef]
- Codd, G.A.; Morrison, L.F.; Metcalf, J.S. Cyanobacterial toxins: Risk management for health protection. Toxicol. Appl. Pharmacol. 2005, 203, 264–272. [Google Scholar] [CrossRef]
- Paerl, H.W.; Otten, T.G. Harmful cyanobacterial blooms: Causes, consequences, and controls. Microb. Ecol. 2013, 65, 995–1010. [Google Scholar] [CrossRef]
- Buratti, F.M.; Manganelli, M.; Vichi, S.; Stefanelli, M.; Scardala, S.; Testai, E.; Funari, E. Cyanotoxins: Producing organisms, occurrence, toxicity, mechanism of action and human health toxicological risk evaluation. Arch. Toxicol. 2017, 91, 1049–1130. [Google Scholar] [CrossRef]
- Grosse, Y.; Baan, R.; Straif, K.; Secretan, B.; El Ghissassi, F.; Cogliano, V.; Cantor, K.P.; Falconer, I.R.; Levallois, P.; Verger, P.; et al. Carcinogenicity of nitrate, nitrite, and cyanobacterial peptide toxins. Lancet Oncol. 2006, 7, 628–629. [Google Scholar] [CrossRef] [Green Version]
- Giannuzzi, L.; Krock, B.; Minaglia, M.C.C.; Rosso, L.; Houghton, C.; Sedan, D.; Malanga, G.; Espinosa, M.; Andrinolo, D.; Hernando, M. Growth, toxin production, active oxygen species and catalase activity of Microcystis aeruginosa (Cyanophyceae) exposed to temperature stress. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2016, 189, 22–30. [Google Scholar] [CrossRef] [Green Version]
- Visser, P.M.; Verspagen, J.M.H.; Sandrini, G.; Stal, L.J.; Matthijs, H.C.P.; Davis, T.W.; Paerl, H.W.; Huisman, J. How rising CO2 and global warming may stimulate harmful cyanobacterial blooms. Harmful Algae 2016, 54, 145–159. [Google Scholar] [CrossRef] [Green Version]
- Carmichael, W.W. Health effects of toxin-producing cyanobacteria: “The CyanoHABs”. Hum. Ecol. Risk Assess. An. Int. J. 2001, 7, 1393–1407. [Google Scholar] [CrossRef]
- Zhou, M.; Tu, W.W.; Xu, J. Mechanisms of microcystin-LR-induced cytoskeletal disruption in animal cells. Toxicon 2015, 101, 92–100. [Google Scholar] [CrossRef]
- Wiśniewska, K.; Lewandowska, A.U.; Śliwińska-Wilczewska, S. The importance of cyanobacteria and microalgae present in aerosols to human health and the environment—Review study. Environ. Int. 2019, 131, 104964. [Google Scholar] [CrossRef]
- Facciponte, D.N.; Bough, M.W.; Seidler, D.; Carroll, J.L.; Ashare, A.; Andrew, A.S.; Tsongalis, G.J.; Vaickus, L.J.; Henegan, P.L.; Butt, T.H.; et al. Identifying aerosolized cyanobacteria in the human respiratory tract: A proposed mechanism for cyanotoxin-associated diseases. Sci. Total Environ. 2018, 645, 1003–1013. [Google Scholar] [CrossRef]
- Lewandowska, A.U.; Śliwińska-Wilczewska, S.; Woźniczka, D. Identification of cyanobacteria and microalgae in aerosols of various sizes in the air over the Southern Baltic Sea. Mar. Pollut. Bull. 2017, 125, 30–38. [Google Scholar] [CrossRef]
- May, N.W.; Olson, N.E.; Panas, M.; Axson, J.L.; Tirella, P.S.; Kirpes, R.M.; Craig, R.L.; Gunsch, M.J.; Laskin, A.; Ault, A.P.; et al. Aerosol emissions from Great lakes harmful algal blooms. Environ. Sci. Technol. 2018, 52, 397–405. [Google Scholar] [CrossRef]
- Backer, L.C.; McNeel, S.V.; Barber, T.; Kirkpatrick, B.; Williams, C.; Irvin, M.; Zhou, Y.; Johnson, T.B.; Nierenberg, K.; Aubel, M.; et al. Recreational exposure to microcystins during algal blooms in two California lakes. Toxicon 2010, 55, 909–921. [Google Scholar] [CrossRef]
- Scherer, P.I.; Raeder, U.; Geist, J.; Zwirglmaier, K. Influence of temperature, mixing, and addition of microcystin-LR on microcystin gene expression in Microcystis aeruginosa. Microbiologyopen 2017, 6, 1–10. [Google Scholar] [CrossRef]
- Nishiwaki-Matsushima, R.; Ohta, T.; Nishiwaki, S.; Suganuma, M.; Kohyama, K.; Ishikawa, T.; Carmichael, W.W.; Fujiki, H. Liver tumor promotion by the cyanobacterial cyclic peptide toxin microcystin-LR. J. Cancer Res. Clin. Oncol. 1992, 118, 420–424. [Google Scholar] [CrossRef]
- Svirčev, Z.; Drobac, D.; Tokodi, N.; Mijović, B.; Codd, G.A.; Meriluoto, J. Toxicology of microcystins with reference to cases of human intoxications and epidemiological investigations of exposures to cyanobacteria and cyanotoxins. Arch. Toxicol. 2017, 91, 621–650. [Google Scholar] [CrossRef]
- McLellan, N.L.; Manderville, R.A. Toxic mechanisms of microcystins in mammals. Toxicol. Res. 2017, 6, 391–405. [Google Scholar] [CrossRef] [Green Version]
- Bouaïcha, N.; Miles, C.O.; Beach, D.G.; Labidi, Z.; Djabri, A.; Benayache, N.Y.; Nguyen-Quang, T. Structural diversity, characterization and toxicology of microcystins. Toxins 2019, 11, 714. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Huang, H.; Xue, L.; Zhuang, D. Effects of MC-LR on ROS level in human bronchial epithelia cells and Chinese hamster ovary cells. Life Sci. J. 2015, 12, 170–173. [Google Scholar]
- Fischer, A.; Hoeger, S.J.; Stemmer, K.; Feurstein, D.J.; Knobeloch, D.; Nussler, A.; Dietrich, D.R. The role of organic anion transporting polypeptides (OATPs/SLCOs) in the toxicity of different microcystin congeners in vitro: A comparison of primary human hepatocytes and OATP-transfected HEK293 cells. Toxicol. Appl. Pharmacol. 2010, 245, 9–20. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Liu, Y.L.; Conklin, A.; Westrick, J.; Weavers, L.K.; Dionysiou, D.D.; Lenhart, J.J.; Mouser, P.J.; Szlag, D.; Walker, H.W. Toxic cyanobacteria and drinking water: Impacts, detection, and treatment. Harmful Algae 2016, 54, 174–193. [Google Scholar] [CrossRef]
- International Agency for Research on Cancer (IARC). IARC Monographs on the Evaluation of Carcinogenic Risks to Humans. Ingested Nitrate and Nitrite, and Cyanobacterial Peptide Toxins; International Agency for Research on Cancer (IARC): Lyon, France, 2010; Volume 94, ISBN 978-92-832-1294-2. [Google Scholar]
- Liu, J.; Wang, B.; Huang, P.; Wang, H.; Xu, K.; Wang, X.; Xu, L.; Guo, Z. Microcystin-LR promotes cell proliferation in the mice liver by activating Akt and p38/ERK/JNK cascades. Chemosphere 2016, 163, 14–21. [Google Scholar] [CrossRef]
- Wang, H.; Xu, K.; Wang, B.; Liu, J.; Wang, X.; Xing, M.; Huang, P.; Guo, Z.; Xu, L. Microcystin-LR induces a wide variety of biochemical changes in the A549 human non-small cell lung cancer cell line: Roles for protein phosphatase 2A and its substrates. Environ. Toxicol. 2017, 32, 1065–1078. [Google Scholar] [CrossRef]
- Zegura, B. An Overview of the mechanisms of Microcystin-LR genotoxicity and potential carcinogenicity. Mini Rev. Med. Chem. 2016, 16, 1042–1062. [Google Scholar] [CrossRef]
- Zhang, X.; Xie, P.; Zhang, X.; Zhou, W.; Zhao, S.; Zhao, Y.; Cai, Y. Toxic effects of microcystin-LR on the HepG2 cell line under hypoxic and normoxic conditions. J. Appl. Toxicol. 2013, 33, 1180–1186. [Google Scholar] [CrossRef]
- Žegura, B.; Sedmak, B.; Filipič, M. Microcystin-LR induces oxidative DNA damage in human hepatoma cell line HepG2. Toxicon 2003, 41, 41–48. [Google Scholar] [CrossRef]
- Li, Y.; Li, J.; Huang, H.; Yang, M.; Zhuang, D.; Cheng, X.; Zhang, H.; Fu, X. Microcystin-LR induces mitochondria-mediated apoptosis in human bronchial epithelial cells. Exp. Ther. Med. 2016, 12, 633–640. [Google Scholar] [CrossRef] [Green Version]
- Lone, Y.; Bhide, M.; Koiri, R.K. Microcystin-LR induced immunotoxicity in mammals. J. Toxicol. 2016, 2016, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Adamovsky, O.; Moosova, Z.; Pekarova, M.; Basu, A.; Babica, P.; Svihalkova Sindlerova, L.; Kubala, L.; Blaha, L. Immunomodulatory potency of microcystin, an important water-polluting cyanobacterial toxin. Environ. Sci. Technol. 2015, 49, 12457–12464. [Google Scholar] [CrossRef]
- Chen, L.; Chen, J.; Zhang, X.; Xie, P. A review of reproductive toxicity of microcystins. J. Hazard. Mater. 2016, 301, 381–399. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Yan, W.; Liu, C.; Li, L.; Yu, L.; Zhao, S.; Li, G. Microcystin-LR exposure induces developmental neurotoxicity in zebrafish embryo. Environ. Pollut. 2016, 213, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Stebbing, J.; Lit, L.C.; Zhang, H.; Darrington, R.S.; Melaiu, O.; Rudraraju, B.; Giamas, G. The regulatory roles of phosphatases in cancer. Oncogene 2014, 33, 939–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez Hernández, J.; López-Rodas, V.; Costas, E. Microcystins from tap water could be a risk factor for liver and colorectal cancer: A risk intensified by global change. Med. Hypotheses 2009, 72, 539–540. [Google Scholar] [CrossRef]
- Kubickova, B.; Babica, P.; Hilscherová, K.; Šindlerová, L. Effects of cyanobacterial toxins on the human gastrointestinal tract and the mucosal innate immune system. Environ. Sci. Eur. 2019, 31. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, G.M.C.; Oliveira, V.R.; Soares, R.M.; Azevedo, S.M.F.O.; Lima, L.M.; Barreiro, E.J.; dos Santos Valenca, S.; Saldiva, P.H.N.; Faffe, D.S.; Zin, W.A. Can LASSBio 596 and dexamethasone treat acute lung and liver inflammation induced by microcystin-LR? Toxicon 2010, 56, 604–612. [Google Scholar] [CrossRef]
- Gupta, N.; Pant, S.C.; Vijayaraghavan, R.; Rao, P.V.L. Comparative toxicity evaluation of cyanobacterial cyclic peptide toxin microcystin variants (LR, RR, YR) in mice. Toxicology 2003, 188, 285–296. [Google Scholar] [CrossRef]
- Soares, R.M.; Cagido, V.R.; Ferraro, R.B.; Meyer-Fernandes, J.R.; Rocco, P.R.M.; Zin, W.A.; Azevedo, S.M.F.O. Effects of microcystin-LR on mouse lungs. Toxicon 2007, 50, 330–338. [Google Scholar] [CrossRef]
- Zhao, S.; Sun, H.; Yan, W.; Xu, D.; Shen, T. A proteomic study of the pulmonary injury induced by microcystin-LR in mice. Toxicon 2018, 150, 304–314. [Google Scholar] [CrossRef]
- Carvalho, G.M.C.; Oliveira, V.R.; Casquilho, N.V.; Araujo, A.C.P.; Soares, R.M.; Azevedo, S.M.F.O.; Pires, K.M.P.; Valença, S.S.; Zin, W.A. Pulmonary and hepatic injury after sub-chronic exposure to sublethal doses of microcystin-LR. Toxicon 2016, 112, 51–58. [Google Scholar] [CrossRef]
- Oliveira, V.R.; Mancin, V.G.L.; Pinto, E.F.; Soares, R.M.; Azevedo, S.M.F.O.; Macchione, M.; Carvalho, A.R.; Zin, W.A. Repeated intranasal exposure to microcystin-LR affects lungs but not nasal epithelium in mice. Toxicon 2015, 104, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xu, L.; Zhou, W.; Zhao, Q.; Wang, Y. Chronic exposure to microcystin-LR affected mitochondrial DNA maintenance and caused pathological changes of lung tissue in mice. Environ. Pollut. 2016, 210, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Benson, M.J.; Hutt, A.J.; Rein, K.; Boggs, E.S.; Barr, B.E.; Fleming, E.L. The toxicity of microcystin LR in mice following 7 days of inhalation exposure. Toxicon 2005, 45, 691–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Gu, S.; Yin, X.; Yuan, M.; Xiang, Z.; Li, Z.; Cao, H.; Meng, X.; Hu, K.; Han, X. The toxic effects of microcystin-LR on mouse lungs and alveolar type II epithelial cells. Toxicon 2016, 115, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.J.; Chien, M.S.; Wu, F.J.; Chou, H.N.; Lee, S.J. Inhibition of embryonic development by microcystin-LR in zebrafish, Danio Rerio. Toxicon 2005, 45, 303–308. [Google Scholar] [CrossRef]
- Overgaard, C.E.; Mitchell, L.A.; Koval, M. Roles for claudins in alveolar epithelial barrier function. Ann. N. Y. Acad. Sci. 2012, 1257, 167–174. [Google Scholar] [CrossRef]
- Repetto, G.; Peso, A.; Zurita, J.L. Neutral red uptake assay for the estimation of cell viability/cytotoxicity. Nat. Protoc. 2008, 3, 1125–1131. [Google Scholar] [CrossRef]
- Organisation for Economic Co-Operation and Development (OECD). Test No. 432: In Vitro 3T3 NRU Phototoxicity Test; OECD Guidelines for the Testing of Chemicals, Section 4; Organisation for Economic Co-Operation and Development (OECD): Paris, France, 2019; ISBN 9789264071162. [Google Scholar]
- Fischer, W.J.; Altheimer, S.; Cattori, V.; Meier, P.J.; Dietrich, D.R.; Hagenbuch, B. Organic anion transporting polypeptides expressed in liver and brain mediate uptake of microcystin. Toxicol. Appl. Pharmacol. 2005, 203, 257–263. [Google Scholar] [CrossRef] [Green Version]
- Bleasby, K.; Castle, J.C.; Roberts, C.J.; Cheng, C.; Bailey, W.J.; Sina, J.F.; Kulkarni, A.V.; Hafey, M.J.; Evers, R.; Johnson, J.M.; et al. Expression profiles of 50 xenobiotic transporter genes in humans and pre-clinical species: A resource for investigations into drug disposition. Xenobiotica 2006, 36, 963–988. [Google Scholar] [CrossRef]
- Tamai, I.; Nezu, J.; Uchino, H.; Sai, Y.; Oku, A.; Shimane, M.; Tsuji, A. molecular identification and characterization of novel members of the human organic anion transporter (OATP) family. Biochem. Biophys. Res. Commun. 2000, 273, 251–260. [Google Scholar] [CrossRef]
- Sakamoto, A.; Matsumaru, T.; Yamamura, N.; Uchida, Y.; Tachikawa, M.; Ohtsuki, S.; Terasaki, T. Quantitative expression of human drug transporter proteins in lung tissues: Analysis of regional, gender, and interindividual differences by liquid chromatography-tandem mass spectrometry. J. Pharm. Sci. 2013, 102, 3395–3406. [Google Scholar] [CrossRef] [PubMed]
- Kreft, M.E.; Jerman, U.D.; Lasič, E.; Hevir-Kene, N.; Rižner, T.L.; Peternel, L.; Kristan, K. The characterization of the human cell line Calu-3 under different culture conditions and its use as an optimized in vitro model to investigate bronchial epithelial function. Eur. J. Pharm. Sci. 2015, 69, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Endter, S.; Francombe, D.; Gumbleton, M.; Ehrhardt, C. RT-PCR analysis of ABC, SLC and SLCO drug transporters in human lung epithelial cell models. J. Pharm. Pharmacol. 2009, 61, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Raska, J.; Ctverackova, L.; Dydowiczova, A.; Sovadinova, I.; Blaha, L.; Babica, P. Tumor-Promoting cyanotoxin microcystin-LR does not induce procarcinogenic events in adult human liver stem cells. Toxicol. Appl. Pharmacol. 2018, 345, 103–113. [Google Scholar] [CrossRef]
- MacKintosh, C.; Beattie, K.A.; Klumpp, S.; Cohen, P.; Codd, G.A. Cyanobacterial microcystin-LR is a potent and specific inhibitor of protein phosphatases 1 and 2A from both mammals and higher plants. FEBS Lett. 1990, 264, 187–192. [Google Scholar] [CrossRef] [Green Version]
- Mikhailov, A.; Härmälä-Braskén, A.-S.; Hellman, J.; Meriluoto, J.; Eriksson, J.E. Identification of ATP-synthase as a novel intracellular target for microcystin-LR. Chem. Biol. Interact. 2003, 142, 223–237. [Google Scholar] [CrossRef]
- Chen, T.; Cui, J.; Liang, Y.; Xin, X.; Owen Young, D.; Chen, C.; Shen, P. Identification of human liver mitochondrial aldehyde dehydrogenase as a potential target for microcystin-LR. Toxicology 2006, 220, 71–80. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhang, L.; Shi, G. Proteasome as a molecular target of microcystin-LR. Toxins 2015, 7, 2221–2231. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Liu, C.; Fu, X.; Zhang, S.; Xin, Y.; Li, Y.; Xue, L.; Cheng, X.; Zhang, H. Microcystin-LR induced apoptosis in rat sertoli cells via the mitochondrial caspase-dependent pathway: Role of reactive oxygen species. Front. Physiol. 2016, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Takser, L.; Benachour, N.; Husk, B.; Cabana, H.; Gris, D. Cyanotoxins at low doses induce apoptosis and inflammatory effects in murine brain cells: Potential implications for neurodegenerative diseases. Toxicol. Rep. 2016, 3, 180–189. [Google Scholar] [CrossRef] [Green Version]
- Basu, A.; Dydowiczová, A.; Čtveráčková, L.; Jaša, L.; Trosko, J.E.; Bláha, L.; Babica, P. Assessment of hepatotoxic potential of cyanobacterial toxins using 3D In Vitro model of adult human liver stem cells. Environ. Sci. Technol. 2018, 52, 10078–10088. [Google Scholar] [CrossRef] [PubMed]
- Menezes, C.; Alverca, E.; Dias, E.; Sam-Bento, F.; Pereira, P. Involvement of endoplasmic reticulum and autophagy in microcystin-LR toxicity in Vero-E6 and HepG2 cell lines. Toxicol. Vitr. 2013, 27, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Chong, M.W.K.; Gu, K.D.; Lam, P.K.S.; Yang, M.; Fong, W.F. Study on the cytotoxicity of microcystin-LR on cultured cells. Chemosphere 2000, 41, 143–147. [Google Scholar] [CrossRef]
- Huguet, A.; Henri, J.; Petitpas, M.; Hogeveen, K.; Fessard, V. Comparative cytotoxicity, oxidative stress, and cytokine secretion induced by two cyanotoxin variants, microcystin-LR and-RR, in human intestinal Caco-2 cells. J. Biochem. Mol. Toxicol. 2013, 27, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Kubickova, B.; Laboha, P.; Hildebrandt, J.P.; Hilscherová, K.; Babica, P. Effects of cylindrospermopsin on cultured immortalized human airway epithelial cells. Chemosphere 2019, 220, 620–628. [Google Scholar] [CrossRef] [PubMed]
- Zihni, C.; Mills, C.; Matter, K.; Balda, M.S. Tight junctions: From simple barriers to multifunctional molecular gates. Nat. Rev. Mol. Cell Biol. 2016, 17, 564–580. [Google Scholar] [CrossRef]
- Zhou, Y.; Geng, X.; Chen, Y.; Shi, H.; Yang, Y.; Zhu, C.; Yu, G.; Tang, Z. Essential roles of Akt/Snail pathway in microcystin-LR-induced tight junction toxicity in Sertoli cell. Food Chem. Toxicol. 2018, 112, 290–298. [Google Scholar] [CrossRef]
- Upadhyay, S.; Palmberg, L. Air-Liquid interface: Relevant In Vitro models for investigating air pollutant-induced pulmonary toxicity. Toxicol. Sci. 2018, 164, 21–30. [Google Scholar] [CrossRef] [Green Version]
- Qian, F.; Deng, J.; Wang, G.; Ye, R.D.; Christman, J.W. Pivotal role of mitogen-activated protein kinase-activated protein kinase 2 in inflammatory pulmonary diseases. Curr. Protein Pept. Sci. 2016, 17, 332–342. [Google Scholar] [CrossRef]
- Sun, Y.; Yu, X.; Li, M.; Liu, J. P44/42 MAPK signal pathway-mediated hyperphosphorylation of paxillin and redistribution of E-cadherin was involved in microcystin-LR-reduced cellular adhesion in a human liver cell line. Chemosphere 2018, 200, 594–602. [Google Scholar] [CrossRef]
- Díez-Quijada, L.; Prieto, A.I.; Guzmán-Guillén, R.; Jos, A.; Cameán, A.M. Occurrence and toxicity of microcystin congeners other than MC-LR and MC-RR: A review. Food Chem. Toxicol. 2019, 125, 106–132. [Google Scholar] [CrossRef] [PubMed]
- Faassen, E.J.; Lürling, M. Occurrence of the microcystins MC-LW and MC-LF in dutch surface waters and their contribution to total microcystin toxicity. Mar. Drugs 2013, 11, 2643–2654. [Google Scholar] [CrossRef] [PubMed]
- Mantzouki, E.; Lürling, M.; Fastner, J.; de Senerpont Domis, L.; Wilk-Woźniak, E.; Koreivienė, J.; Seelen, L.; Teurlincx, S.; Verstijnen, Y.; Krztoń, W.; et al. Temperature effects explain continental scale distribution of cyanobacterial toxins. Toxins 2018, 10, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization (WHO). Guidelines for Safe Recreational Water Environments, 1st-Coasta ed.; World Health Organization (WHO): Geneva, Switzerland, 2003; ISBN 9241545801. [Google Scholar]
- United States Environmental Protection Agency (EPA). Recommended Human Health Recreational Ambient Water Quality Criteria or Swimming Advisories for Microcystins and Cylindrospermopsin; (EPA 822-R-19-001); United States Environmental Protection Agency (EPA): Washington, DC, USA, 2019.
- Pilotto, L.S.; Douglas, R.M.; Burch, M.D.; Cameron, S. Health effects of exposure to cyanobacteria (blue-green algae ) during recreational water-related activities. Aust. N. Z. J. Public Health 1997, 21, 562–566. [Google Scholar] [CrossRef] [Green Version]
- Stewart, I.; Webb, P.M.; Schluter, P.J.; Fleming, L.E.; Burns, J.W.; Gantar, M.; Backer, L.C.; Shaw, G.R. Epidemiology of recreational exposure to freshwater cyanobacteria—An international prospective cohort study. BMC Public Health 2006, 6, 93. [Google Scholar] [CrossRef] [Green Version]
- Yankaskas, J.R.; Haizlip, J.E.; Conrad, M.; Koval, D.; Lazarowski, E.; Paradiso, A.M.; Rinehart, C.A.; Sarkadi, B.; Schlegel, R.; Boucher, R.C. Papilloma-Virus immortalized tracheal epithelial cells retail a well-differentiated phenotype. Am. J. Physiol. 1993, 264, C1219–C1230. [Google Scholar] [CrossRef]
- Gruenert, D.C.; Willems, M.; Cassiman, J.J.; Frizzell, R.A. Established cell lines used in cystic fibrosis research. J. Cyst. Fibros. 2004, 3, 191–196. [Google Scholar] [CrossRef] [Green Version]
- Cozens, A.L.; Yezzi, M.J.; Kunzelmann, K.; Ohrui, T.; Chin, L.; Eng, K.; Finkbeiner, W.E.; Widdicombe, J.H.; Gruenert, D.C. CFTR expression and chloride secretion in polarized immortal human bronchial epithelial cells. Am. J. Respir. Cell Mol. Biol. 1994, 10, 38–47. [Google Scholar] [CrossRef]
- ACEA Biosciences. Calculation Principles of RTCA Software; ACEA Biosciences: San Diego, CA, USA, 2016. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MC-LR Administration | Respiratory-Related Symptoms | Reference |
|---|---|---|
| Intraperitoneal (acute) | Change in respiratory variables, increased lung impedance, pulmonary lesions, lung inflammation. | Carvalho et al., 2010 [37]; Gupta et al., 2003 [38]; Soares et al., 2007 [39] |
| Intratracheal instillation (acute) | Increased number alveolar septa collapsed areas, fractured alveolar walls, damage to F-actin, multiple inflammatory-related protein alterations. | Zhao et al., 2018 [40] |
| Intraperitoneal (20 days) | Impairment of respiratory mechanics, pulmonary parenchyma degradation, augmented contents of inflammatory mediators in lung tissue, a dose-dependent lung inflammatory response. | Carvalho et al., 2016 [41] |
| Intranasal (30 days) | Impairment of all respiratory mechanical components, pulmonary parenchyma damage marked by the augmented alveolar collapsed areas and the number of inflammatory cells | Oliveira et al., 2015 [42] |
| Intranasal (30 days) | Lung structure disorder, thickening of alveolar septa, aggregation of inflammatory cells (induction of oxidative stress, altered expression of inflammatory cytokines, etc.) | X. Li et al., 2016 [43] |
| Inhalation (7 days) | Degeneration and necrosis of nasal respiratory epithelium, neutrophilic inflammation | Benson et al., 2005 [44] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brózman, O.; Kubickova, B.; Babica, P.; Laboha, P. Microcystin-LR Does Not Alter Cell Survival and Intracellular Signaling in Human Bronchial Epithelial Cells. Toxins 2020, 12, 165. https://doi.org/10.3390/toxins12030165
Brózman O, Kubickova B, Babica P, Laboha P. Microcystin-LR Does Not Alter Cell Survival and Intracellular Signaling in Human Bronchial Epithelial Cells. Toxins. 2020; 12(3):165. https://doi.org/10.3390/toxins12030165
Chicago/Turabian StyleBrózman, Ondřej, Barbara Kubickova, Pavel Babica, and Petra Laboha. 2020. "Microcystin-LR Does Not Alter Cell Survival and Intracellular Signaling in Human Bronchial Epithelial Cells" Toxins 12, no. 3: 165. https://doi.org/10.3390/toxins12030165
APA StyleBrózman, O., Kubickova, B., Babica, P., & Laboha, P. (2020). Microcystin-LR Does Not Alter Cell Survival and Intracellular Signaling in Human Bronchial Epithelial Cells. Toxins, 12(3), 165. https://doi.org/10.3390/toxins12030165