Effects of Microcystin-LR on Metabolic Functions and Structure Succession of Sediment Bacterial Community under Anaerobic Conditions

 , ,
, ,
Abstract
:1. Introduction
2. Results
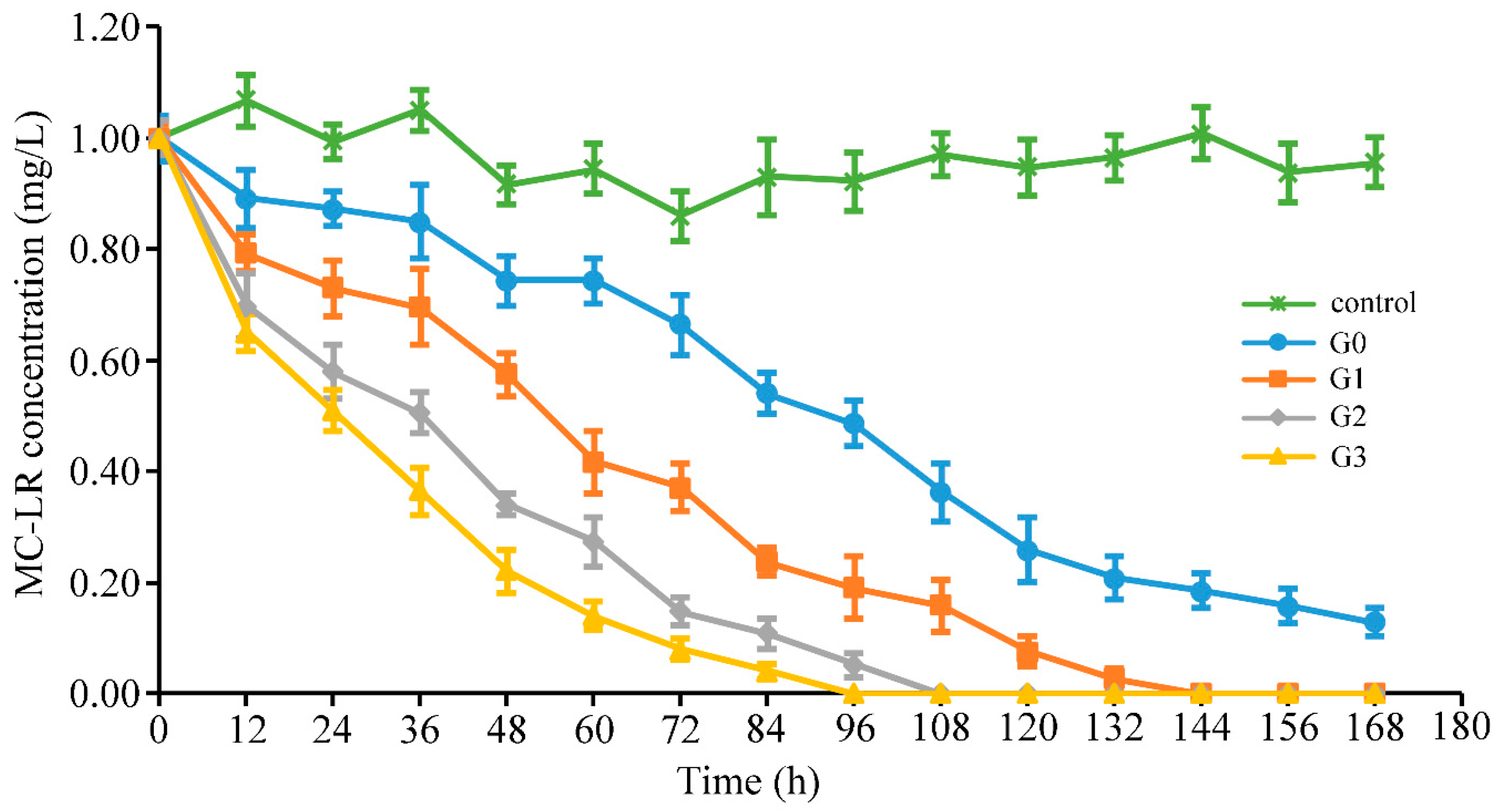
2.1. Effects on MC-Degrading Ability of Bacterial Community
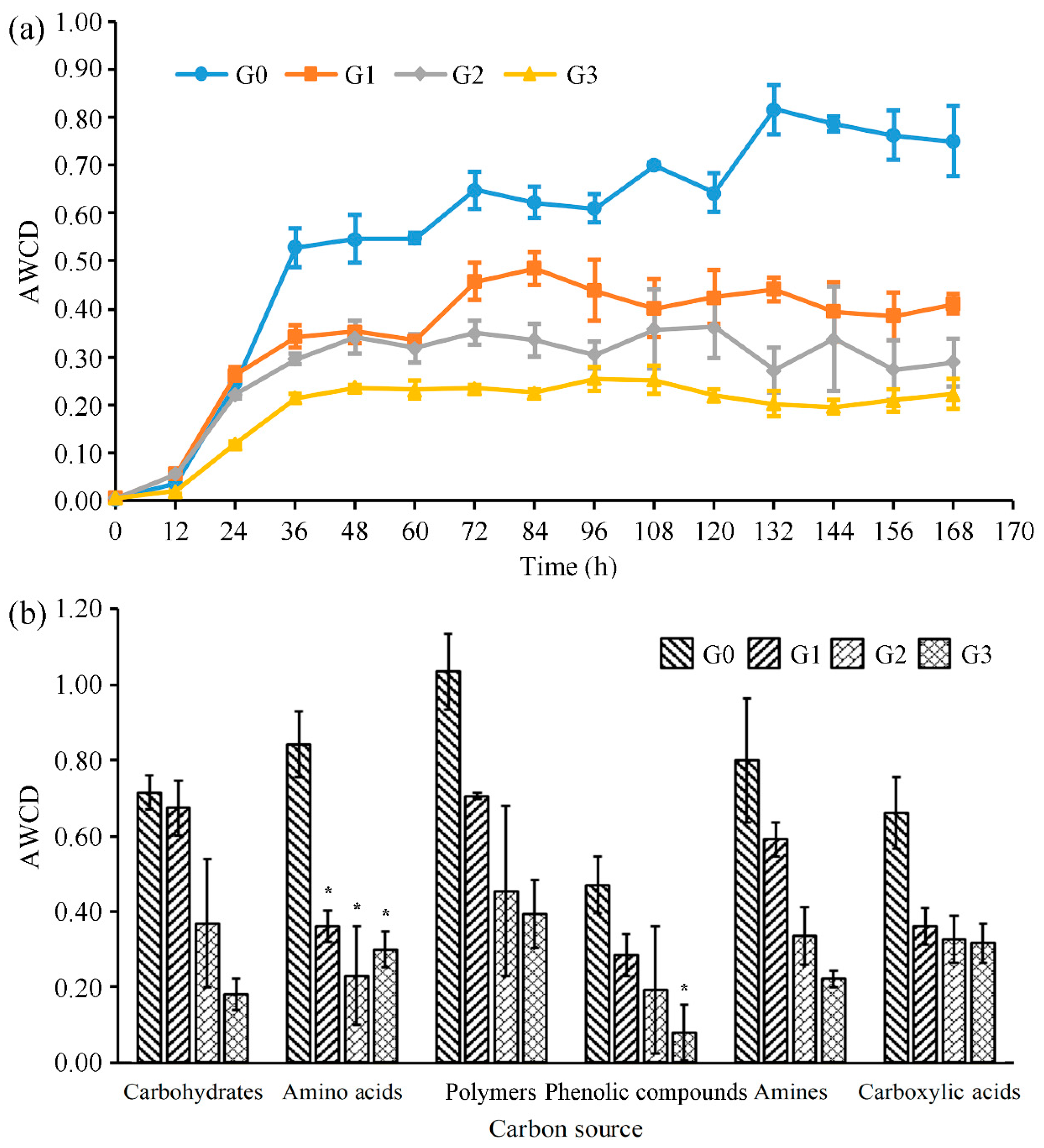
2.2. Effects on Metabolic Functions of Bacterial Community
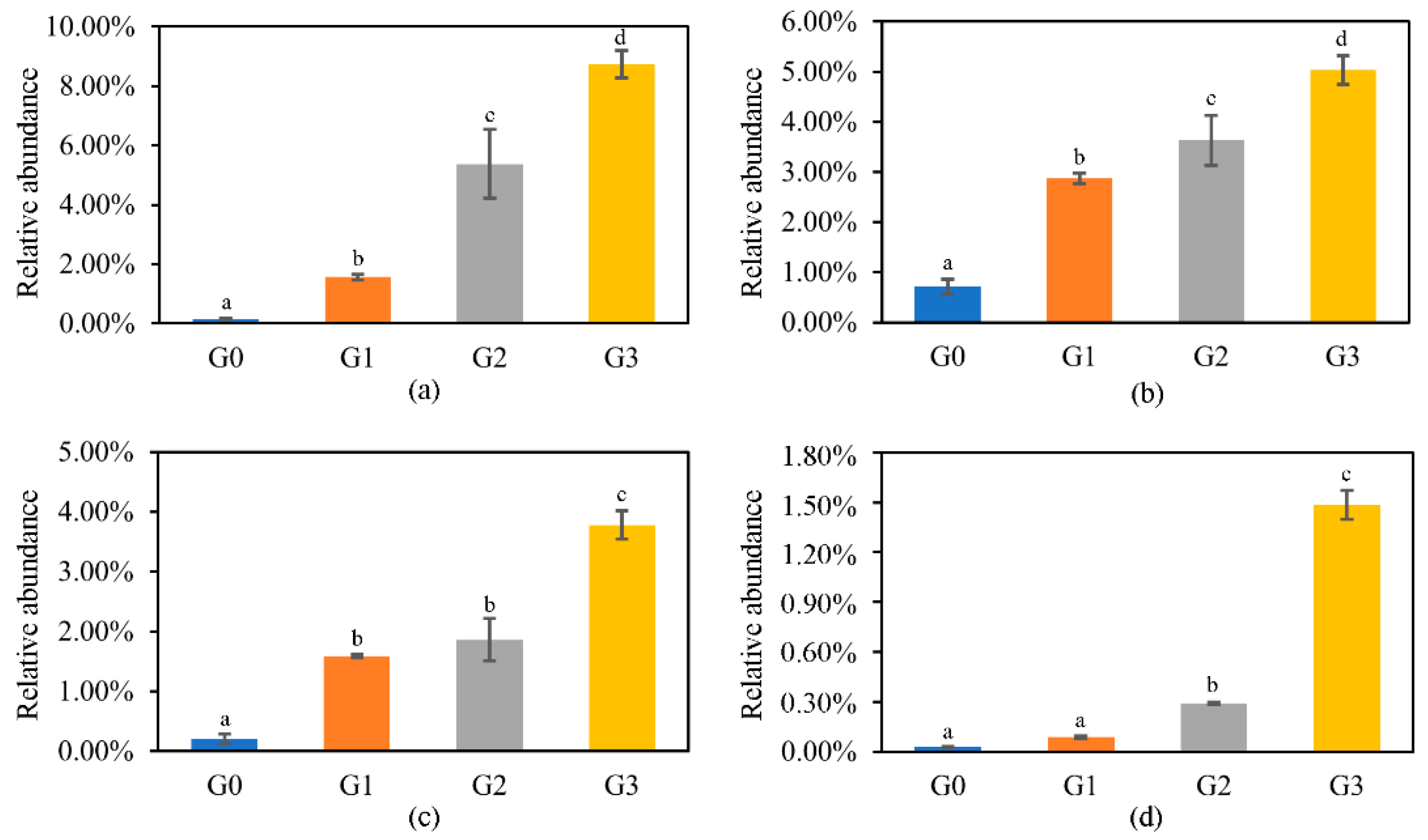
2.3. Effects on the Diversity and Composition of Bacterial Community
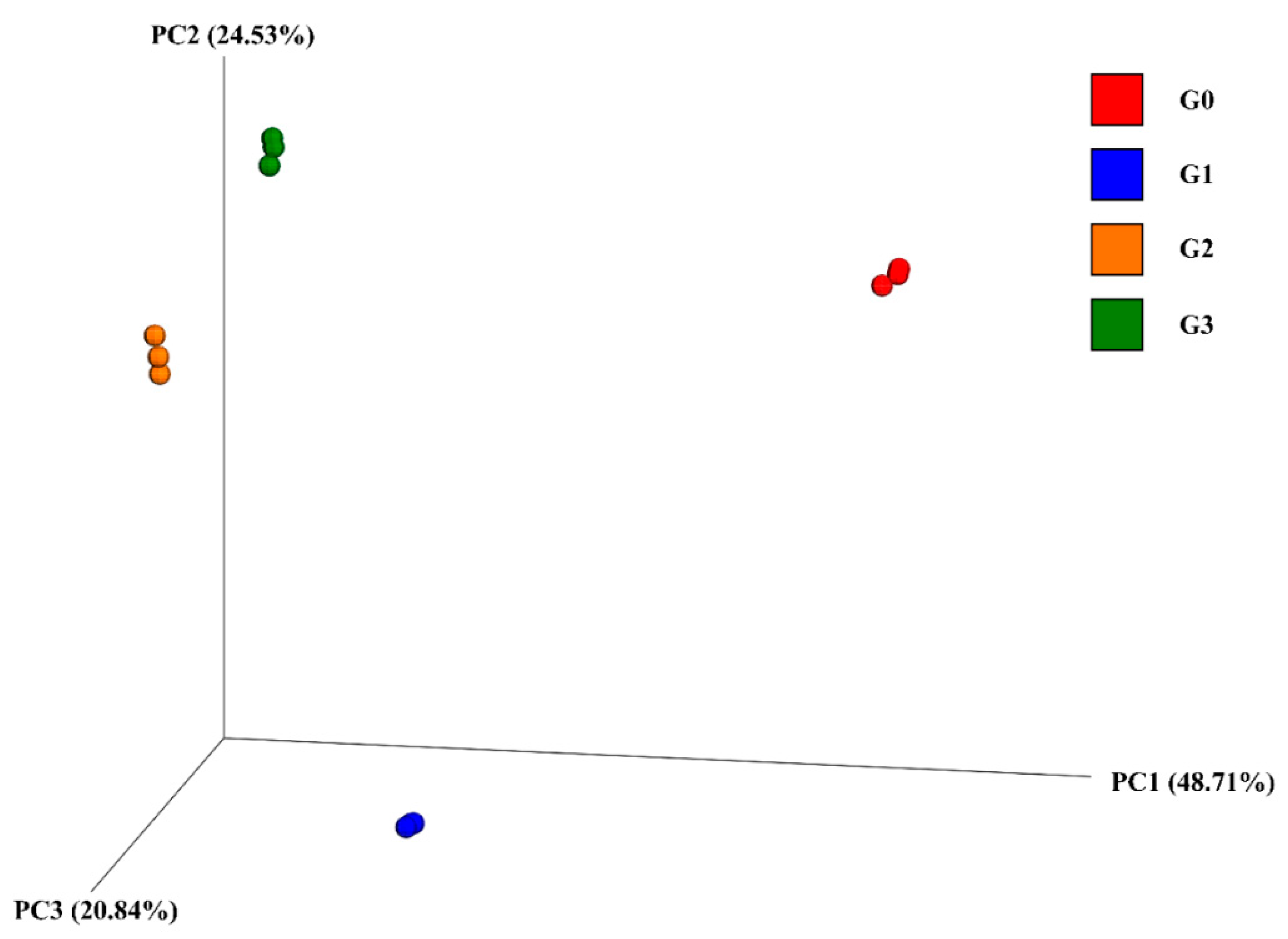
2.3.1. Diversity of the Bacterial Community
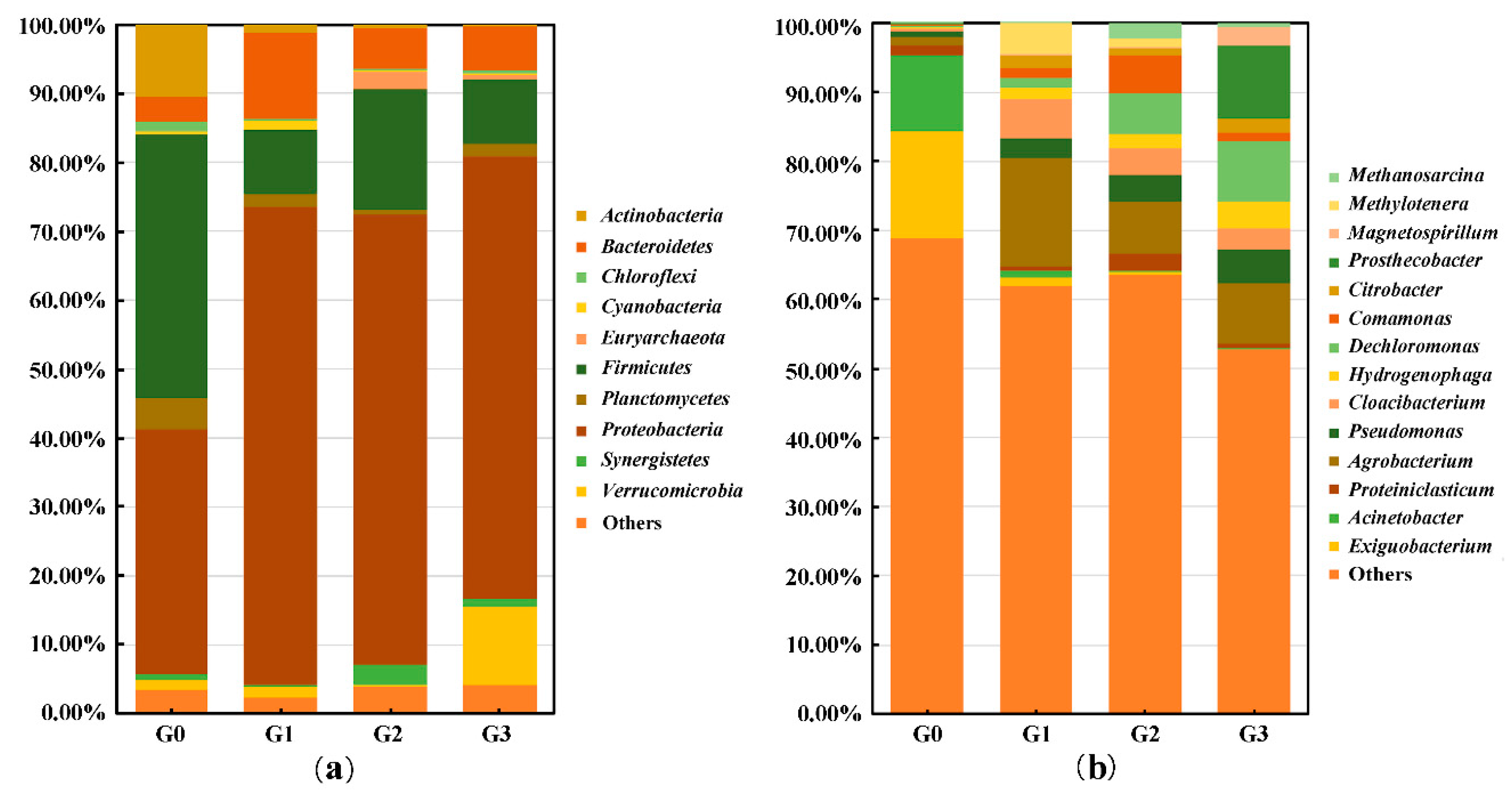
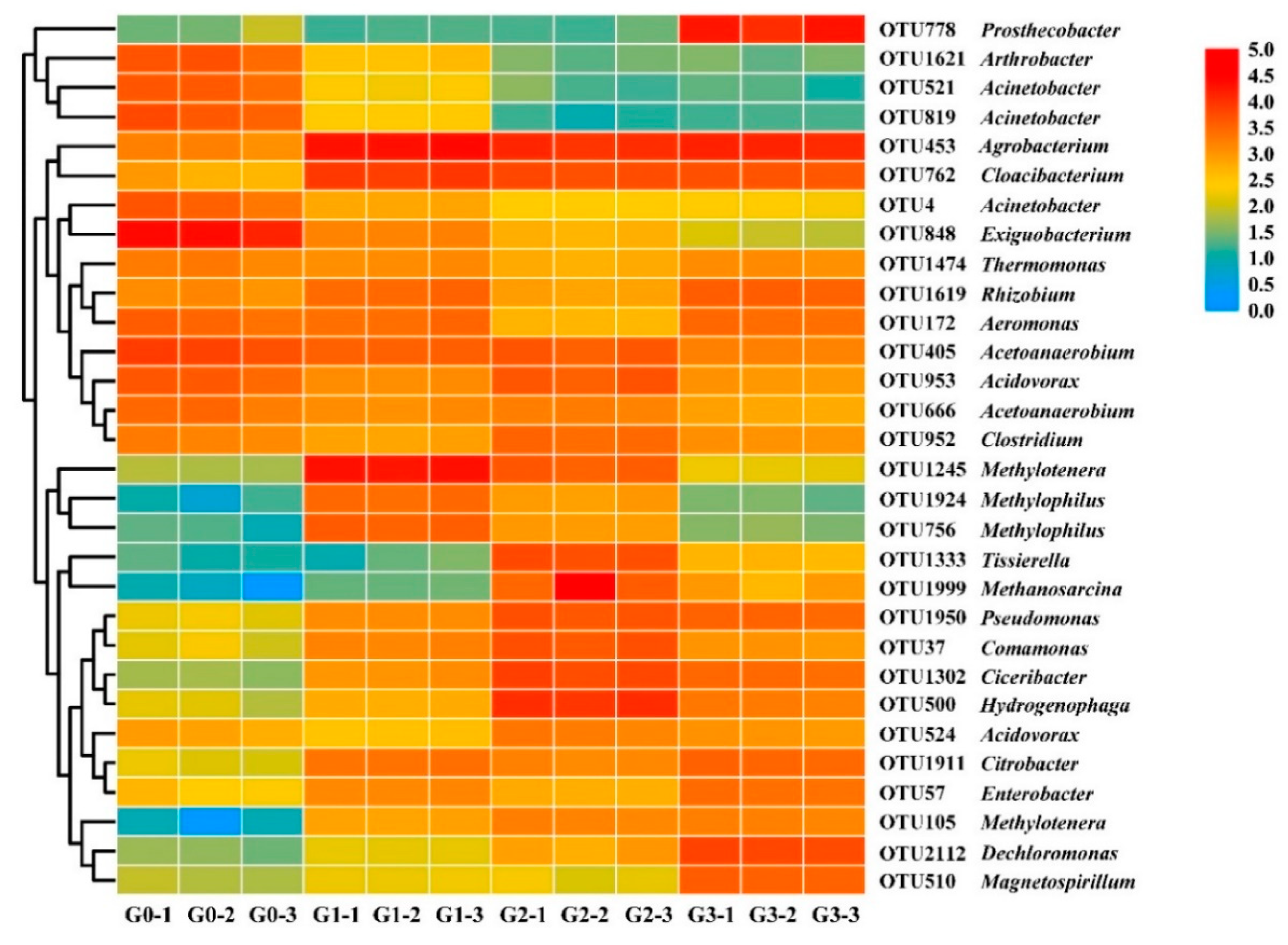
2.3.2. Changes in Bacterial Community Structure
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Sediment Samples and Reagents
5.2. Obtain of Bacterial Community and Experimental Setup
5.3. Detection of MC-LR
5.4. Metabolic Ability of Bacterial Community
5.5. DNA Extraction and Sequencing
5.6. Analysis of the Bacterial Community Structure
5.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.H.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Otten, T.G. Blooms bite the hand that feeds them. Science 2013, 342, 433–434. [Google Scholar] [CrossRef] [PubMed]
- Massey, I.Y.; Zhang, X.; Yang, F. Importance of bacterial biodegradation and detoxification processes of microcystins for environmental health. J. Toxicol. Environ. Health B 2018, 21, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Zilliges, Y.; Kehr, J.C.; Meissner, S.; Ishida, K.; Mikkat, S.; Hagemann, M.; Kaplan, A.; Borner, T.; Dittmann, E. The cyanobacterial hepatotoxin microcystin binds to proteins and increases the fitness of Microcystis under oxidative stress conditions. PLoS ONE 2011, 6, e17615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dittmann, E.; Erhard, M.; Kaebernick, M.; Scheler, C.; Neilan, B.A.; von Dohren, H.; Borner, T. Altered expression of two light-dependent genes in a microcystin-lacking mutant of Microcystis aeruginosa PCC 7806. Microbiology 2001, 147, 3113–3119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schatz, D.; Keren, Y.; Vardi, A.; Sukenik, A.; Carmeli, S.; Borner, T.; Dittmann, E.; Kaplan, A. Towards clarification of the biological role of microcystins, a family of cyanobacterial toxins. Environ. Microbiol. 2007, 9, 965–970. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Rzymski, P. Programmed cell death-like and accompanying release of microcystin in freshwater bloom-forming cyanobacterium Microcystis: From identification to ecological relevance. Toxins 2019, 11, 12. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Massey, I.Y.; Guo, J.; Yang, S.; Pu, Y.P.; Zeng, W.M.; Tan, H.Z. Microcystin-LR degradation utilizing a novel effective indigenous bacterial community YFMCD1 from Lake Taihu. J. Toxicol. Environ. Health A 2018, 81, 184–193. [Google Scholar] [CrossRef]
- Chen, L.; Xie, P. Mechanisms of microcystin-induced cytotoxicity and apoptosis. Mini-Rev. Med. Chem. 2016, 16, 1018–1031. [Google Scholar] [CrossRef]
- Li, J.M.; Li, R.H.; Li, J. Current research scenario for microcystins biodegradation—A review on fundamental knowledge, application prospects and challenges. Sci. Total Environ. 2017, 595, 615–632. [Google Scholar] [CrossRef]
- Shimizu, K.; Sano, T.; Kubota, R.; Kobayashi, N.; Tahara, M.; Obama, T.; Sugimoto, N.; Nishimura, T.; Ikarashi, Y. Effects of the amino acid constituents of microcystin variants on cytotoxicity to primary cultured rat hepatocytes. Toxins 2014, 6, 168–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Giesy, J.P.; Xie, P. The dose makes the poison. Sci. Total Environ. 2018, 621, 649–653. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.Q.; Wang, C.B.; Tian, C.C.; Xiao, B.D.; Song, L.R. Evaluation of the potential of anoxic biodegradation of intracellular and dissolved microcystins in lake sediments. J. Hazard. Mater. 2015, 286, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Henao, E.; Rzymski, P.; Waters, M.N. A review on the study of cyanotoxins in paleolimnological research: Current knowledge and future needs. Toxins 2019, 12, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunberg, A.K.; Blomqvist, P. Benthic overwintering of Microcystis colonies under different environmental conditions. J. Plankton Res. 2002, 24, 1247–1252. [Google Scholar] [CrossRef]
- Song, H.; Coggins, L.X.; Reichwaldt, E.S.; Ghadouani, A. The importance of lake sediments as a pathway for microcystin dynamics in shallow eutrophic lakes. Toxins 2015, 7, 900–918. [Google Scholar] [CrossRef] [PubMed]
- Lezcano, M.A.; Velazquez, D.; Quesada, A.; El-Shehawy, R. Diversity and temporal shifts of the bacterial community associated with a toxic cyanobacterial bloom: An interplay between microcystin producers and degraders. Water Res. 2017, 125, 52–61. [Google Scholar] [CrossRef]
- Su, X.M.; Steinman, A.D.; Tang, X.M.; Xue, Q.J.; Zhao, Y.Y.; Xie, L.Q. Response of bacterial communities to cyanobacterial harmful algal blooms in Lake Taihu, China. Harmful Algae 2017, 68, 168–177. [Google Scholar] [CrossRef]
- Parulekar, N.N.; Kolekar, P.; Jenkins, A.; Kleiven, S.; Utkilen, H.; Johansen, A.; Sawant, S.; Kulkarni-Kale, U.; Kale, M.; Saebo, M. Characterization of bacterial community associated with phytoplankton bloom in a eutrophic lake in South Norway using 16S rRNA gene amplicon sequence analysis. PLoS ONE 2017, 12, e0173408. [Google Scholar] [CrossRef]
- Chen, W.; Song, L.R.; Peng, L.; Wan, N.; Zhang, X.M.; Gan, N.Q. Reduction in microcystin concentrations in large and shallow lakes: Water and sediment-interface contributions. Water Res. 2008, 42, 763–773. [Google Scholar] [CrossRef]
- Zastepa, A.; Pick, F.R.; Blais, J.M.; Saleem, A. Analysis of intracellular and extracellular microcystin variants in sediments and pore waters by accelerated solvent extraction and high performance liquid chromatography-tandem mass spectrometry. Anal. Chim. Acta 2015, 872, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Maerki, M.; Muller, B.; Dinkel, C.; Wehrli, B. Mineralization pathways in lake sediments with different oxygen and organic carbon supply. Limnol. Oceanogr. 2009, 54, 428–438. [Google Scholar] [CrossRef] [Green Version]
- Yuan, X.Z.; Shi, X.S.; Zhang, D.L.; Qiu, Y.L.; Guo, R.B.; Wang, L.S. Biogas production and microcystin biodegradation in anaerobic digestion of blue algae. Energ. Environ. Sci. 2011, 4, 1511–1515. [Google Scholar] [CrossRef]
- Conrad, R.; Chan, O.C.; Claus, P.; Casper, P. Characterization of methanogenic Archaea and stable isotope fractionation during methane production in the profundal sediment of an oligotrophic lake (Lake Stechlin, Germany). Limnol. Oceanogr. 2007, 52, 1393–1406. [Google Scholar] [CrossRef]
- Chen, X.G.; Yang, X.; Yang, L.L.; Xiao, B.D.; Wu, X.Q.; Wang, J.T.; Wan, H.G. An effective pathway for the removal of microcystin LR via anoxic biodegradation in lake sediments. Water Res. 2010, 44, 1884–1892. [Google Scholar] [CrossRef] [PubMed]
- Holst, T.; Jorgensen, N.O.G.; Jorgensen, C.; Johansen, A. Degradation of microcystin in sediments at oxic and anoxic, denitrifying conditions. Water Res. 2003, 37, 4748–4760. [Google Scholar] [CrossRef]
- Wu, Y.C.; Cai, P.; Jing, X.X.; Niu, X.K.; Ji, D.D.; Ashry, N.M.; Gao, C.H.; Huang, Q.Y. Soil biofilm formation enhances microbial community diversity and metabolic activity. Environ. Int. 2019, 132, 105116. [Google Scholar] [CrossRef]
- Cao, Q.; Steinman, A.D.; Su, X.; Xie, L. Effects of microcystins contamination on soil enzyme activities and microbial community in two typical lakeside soils. Environ. Pollut. 2017, 231, 134–142. [Google Scholar] [CrossRef]
- Garland, J.L. Analysis and interpretation of community-level physiological profiles in microbial ecology. FEMS Microbiol. Ecol. 1997, 24, 289–300. [Google Scholar] [CrossRef]
- Preston-Mafham, J.; Boddy, L.; Randerson, P.F. Analysis of microbial community functional diversity using sole-carbon-source utilisation profiles—A critique. FEMS Microbiol. Ecol. 2002, 42, 1–14. [Google Scholar]
- Ding, Q.; Liu, K.Y.; Xu, K.; Sun, R.L.; Zhang, J.; Yin, L.H.; Pu, Y.P. Further understanding of degradation pathways of microcystin-LR by an indigenous Sphingopyxis sp. in environmentally relevant pollution concentrations. Toxins 2018, 10, 536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallego, S.; Devers-Lamrani, M.; Rousidou, K.; Karpouzas, D.G.; Martin-Laurent, F. Assessment of the effects of oxamyl on the bacterial community of an agricultural soil exhibiting enhanced biodegradation. Sci. Total Environ. 2019, 651, 1189–1198. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Zhou, Y.L.; Sun, R.L.; Wei, H.Y.; Li, Y.H.; Yin, L.H.; Pu, Y.P. Biodegradation of microcystin-LR and-RR by a novel microcystin-degrading bacterium isolated from Lake Taihu. Biodegradation 2014, 25, 447–457. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Feng, H.; Li, X.; Yi, X.; Guo, J.; Clara, T.; Yang, F. Anaerobic degradation of microcystin-LR by an indigenous bacterial Enterobacter sp. YF3. J. Toxicol. Environ. Health A 2019, 82, 1120–1128. [Google Scholar] [CrossRef]
- Bao, Z.; Wu, Y. Biodegradation of microcystin-LR by an amino acid-degrading anaerobic bacterium. Desalin. Water Treat. 2014, 57, 870–880. [Google Scholar] [CrossRef]
- Zhu, F.P.; Han, Z.L.; Duan, J.L.; Shi, X.S.; Wang, T.T.; Sheng, G.P.; Wang, S.G.; Yuan, X.Z. A novel pathway for the anaerobic biotransformation of microcystin-LR using enrichment cultures. Environ. Pollut. 2019, 247, 1064–1070. [Google Scholar] [CrossRef]
- Guo, P.P.; Zhu, L.S.; Wang, J.H.; Wang, J.; Liu, T. Effects of alkyl-imidazolium ionic liquid [Omim]Cl on the functional diversity of soil microbial communities. Environ. Sci. Pollut. R 2015, 22, 9059–9066. [Google Scholar] [CrossRef]
- Lear, G.; Bellamy, J.; Case, B.S.; Lee, J.E.; Buckley, H.L. Fine-scale spatial patterns in bacterial community composition and function within freshwater ponds. ISME J. 2014, 8, 1715–1726. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Lu, Q.Q.; Ding, Q.; Yin, L.H.; Pu, Y.P. A novel and native microcystin-degrading bacterium of Sphingopyxis sp isolated from Lake Taihu. Int. J. Environ. Res. Public Health 2017, 14, 1187. [Google Scholar] [CrossRef] [Green Version]
- Fang, H.; Lian, J.J.; Wang, H.F.; Cai, L.; Yu, Y.L. Exploring bacterial community structure and function associated with atrazine biodegradation in repeatedly treated soils. J. Hazard. Mater. 2015, 286, 457–465. [Google Scholar] [CrossRef]
- Ma, G.X.; Pei, H.Y.; Hu, W.R.; Xu, X.C.; Ma, C.X.; Li, X.Q. The removal of cyanobacteria and their metabolites through anoxic biodegradation in drinking water sludge. Bioresour. Technol. 2014, 165, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.X.; Pei, H.Y.; Hu, W.R.; Xu, X.C.; Ma, C.X.; Pei, R.T. Effects of glucose on microcystin-LR removal and the bacterial community composition through anoxic biodegradation in drinking water sludge. Environ. Technol. 2016, 37, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, R.; O’Connor, S.M.; Chan, E.; Coates, J.D. Anaerobic degradation of benzene, toluene, ethylbenzene, and xylene compounds by Dechloromonas strain RCB. Appl. Environ. Microb. 2005, 71, 8649–8655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, R.; Coates, J.D. Hydroxylation and carboxylation-two crucial steps of anaerobic benzene degradation by Dechloromonas strain RCB. Appl. Environ. Microb. 2005, 71, 5427–5432. [Google Scholar] [CrossRef] [Green Version]
- Salinero, K.K.; Keller, K.; Feil, W.S.; Feil, H.; Trong, S.; Di Bartolo, G.; Lapidus, A. Metabolic analysis of the soil microbe Dechloromonas aromatica str. RCB: Indications of a surprisingly complex life-style and cryptic anaerobic pathways for aromatic degradation. BMC Genom. 2009, 10. [Google Scholar] [CrossRef] [Green Version]
- Takenaka, S.; Watanabe, M.F. Microcystin LR degradation by Pseudomonas aeruginosa alkaline protease. Chemosphere 1997, 34, 749–757. [Google Scholar] [CrossRef]
- Kang, Y.H.; Park, C.S.; Han, M.S. Pseudomonas aeruginosa UCBPP-PA14 a useful bacterium capable of lysing Microcystis aeruginosa cells and degrading microcystins. J. Appl. Phycol. 2012, 24, 1517–1525. [Google Scholar] [CrossRef]
- Lemes, G.A.F.; Kist, L.W.; Bogo, M.R.; Yunes, J.S. Biodegradation of [D-Leu(1)] microcystin-LR by a bacterium isolated from sediment of Patos Lagoon estuary, Brazil. J. Venom. Anim Toxins 2015, 21. [Google Scholar] [CrossRef] [Green Version]
- Magic-Knezev, A.; Wullings, B.; Van der Kooij, D. Polaromonas and Hydrogenophaga species are the predominant bacteria cultured from granular activated carbon filters in water treatment. J. Appl. Microbiol. 2009, 107, 1457–1467. [Google Scholar] [CrossRef]
- Meyer-Cifuentes, I.; Fiedler, S.; Muller, J.A.; Kappelmeyer, U.; Mausezahl, I.; Heipieper, H.J. Draft genome sequence of Magnetospirillum sp. strain 15-1, a denitrifying toluene degrader isolated from a planted fixed-bed reactor. Microbiol. Resour. Ann. 2017, 5, e00764-17. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.X.; Hu, C.H.; Guo, L.Z.; Li, K.R.; Yu, H. Isolation of a 3-hydroxypyridine degrading bacterium, Agrobacterium sp. DW-1, and its proposed degradation pathway. AMB Express 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.H.; Zhang, C.; Yan, Y.C. Biodegradation of methyl parathion and p-nitrophenol by a newly isolated Agrobacterium sp strain Yw12. Biodegradation 2012, 23, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.N.; Liu, Z.; Xu, P. Biodegradation of nicotine by a newly isolated Agrobacterium sp strain S33. J. Appl. Microbiol. 2009, 107, 838–847. [Google Scholar] [CrossRef] [PubMed]
- Struthers, J.K.; Jayachandran, K.; Moorman, T.B. Biodegradation of atrazine by Agrobacterium radiobacter J14a and use of this strain in bioremediation of contaminated soil. Appl. Environ. Microb. 1998, 64, 3368–3375. [Google Scholar] [CrossRef] [Green Version]
- Misson, B.; Sabarta, M.; Amblard, C.; Latour, D. Benthic survival of Microcystis: Long-term viability and ability to transcribe microcystin genes. Harmful Algae 2012, 13, 20–25. [Google Scholar] [CrossRef]
- Bourne, D.G.; Jones, G.J.; Blakeley, R.L.; Jones, A.; Negri, A.P.; Riddles, P. Enzymatic pathway for the bacterial degradation of the cyanobacterial cyclic peptide toxin microcystin LR. Appl. Environ. Microbiol. 1996, 62, 4086–4094. [Google Scholar] [CrossRef] [Green Version]
- Bourne, D.G.; Riddles, P.; Jones, G.J.; Smith, W.; Blakeley, R.L. Characterisation of a gene cluster involved in bacterial degradation of the cyanobacterial toxin microcystin LR. Environ. Toxicol 2001, 16, 523–534. [Google Scholar] [CrossRef]
- Dziga, D.; Tokodi, N.; Backovic, D.D.; Kokocinski, M.; Antosiak, A.; Puchalski, J.; Strzalka, W.; Madej, M.; Meriluoto, J.; Svircev, Z. The Effect of a combined hydrogen peroxide-MlrA treatment on the phytoplankton community and microcystin concentrations in a mesocosm experiment in Lake Ludos. Toxins 2019, 11, 725. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, L. Enzymatic pathway for MCLR degradation by bacterium CJ5. Master’s Thesis, Wuhan University of Technology, Wuhan, China, 2014. [Google Scholar]
- Tang, X.; Krausfeldt, L.E.; Shao, K.; LeCleir, G.R.; Stough, J.M.A.; Gao, G.; Boyer, G.L.; Zhang, Y.; Paerl, H.W.; Qin, B.; et al. Seasonal gene expression and the ecophysiological implications of toxic Microcystis aeruginosa blooms in Lake Taihu. Environ. Sci. Technol. 2018, 52, 11049–11059. [Google Scholar] [CrossRef]
- Chiodini, R.J.; Dowd, S.E.; Chamberlin, W.M.; Galandiuk, S.; Davis, B.; Glassing, A. Microbial population differentials between mucosal and submucosal intestinal tissues in advanced crohn’s disease of the ileum. PLoS ONE 2015, 10, e0134382. [Google Scholar] [CrossRef]
- Glassing, A.; Dowd, S.E.; Galandiuk, S.; Davis, B.; Jorden, J.R.; Chiodini, R.J. Changes in 16s RNA gene microbial community profiling by concentration of prokaryotic DNA. J. Microbiol. Methods 2015, 119, 239–242. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | OTU Number | Shannon | Simpson | Pielou | Chao1 | ACE | Goods_Coverage |
|---|---|---|---|---|---|---|---|
| G0 | 1369 ± 31.19a | 7.22 ± 0.36a | 0.97 ± 0.01 | 0.69 ± 0.04a | 1498.62 ± 16.88a | 1470.41 ± 31.44a | 99.91% ± 0.00%a |
| G1 | 1045 ± 11.36b | 6.34 ± 0.06b | 0.95 ± 0.00 | 0.63 ± 0.00b | 1235.11 ± 59.95b | 1205.59 ± 26.19b | 99.87% ± 0.02%b |
| G2 | 844 ± 33.78c | 6.38 ± 0.35b | 0.96 ± 0.02 | 0.66 ± 0.03 | 1089.2 ± 85.93c | 1043.07 ± 58.28c | 99.89% ± 0.02% |
| G3 | 1025.33 ± 25.01b | 6.72 ± 0.08b | 0.97 ± 0.00 | 0.67 ± 0.01 | 1223.26 ± 36.18b | 1186.05 ± 41.49b | 99.88% ± 0.00% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, Q.; Liu, K.; Song, Z.; Sun, R.; Zhang, J.; Yin, L.; Pu, Y. Effects of Microcystin-LR on Metabolic Functions and Structure Succession of Sediment Bacterial Community under Anaerobic Conditions. Toxins 2020, 12, 183. https://doi.org/10.3390/toxins12030183
Ding Q, Liu K, Song Z, Sun R, Zhang J, Yin L, Pu Y. Effects of Microcystin-LR on Metabolic Functions and Structure Succession of Sediment Bacterial Community under Anaerobic Conditions. Toxins. 2020; 12(3):183. https://doi.org/10.3390/toxins12030183
Chicago/Turabian StyleDing, Qin, Kaiyan Liu, Zhiquan Song, Rongli Sun, Juan Zhang, Lihong Yin, and Yuepu Pu. 2020. "Effects of Microcystin-LR on Metabolic Functions and Structure Succession of Sediment Bacterial Community under Anaerobic Conditions" Toxins 12, no. 3: 183. https://doi.org/10.3390/toxins12030183
APA StyleDing, Q., Liu, K., Song, Z., Sun, R., Zhang, J., Yin, L., & Pu, Y. (2020). Effects of Microcystin-LR on Metabolic Functions and Structure Succession of Sediment Bacterial Community under Anaerobic Conditions. Toxins, 12(3), 183. https://doi.org/10.3390/toxins12030183