An Appetite for Destruction: Detecting Prey-Selective Binding of α-Neurotoxins in the Venom of Afro-Asian Elapids

 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
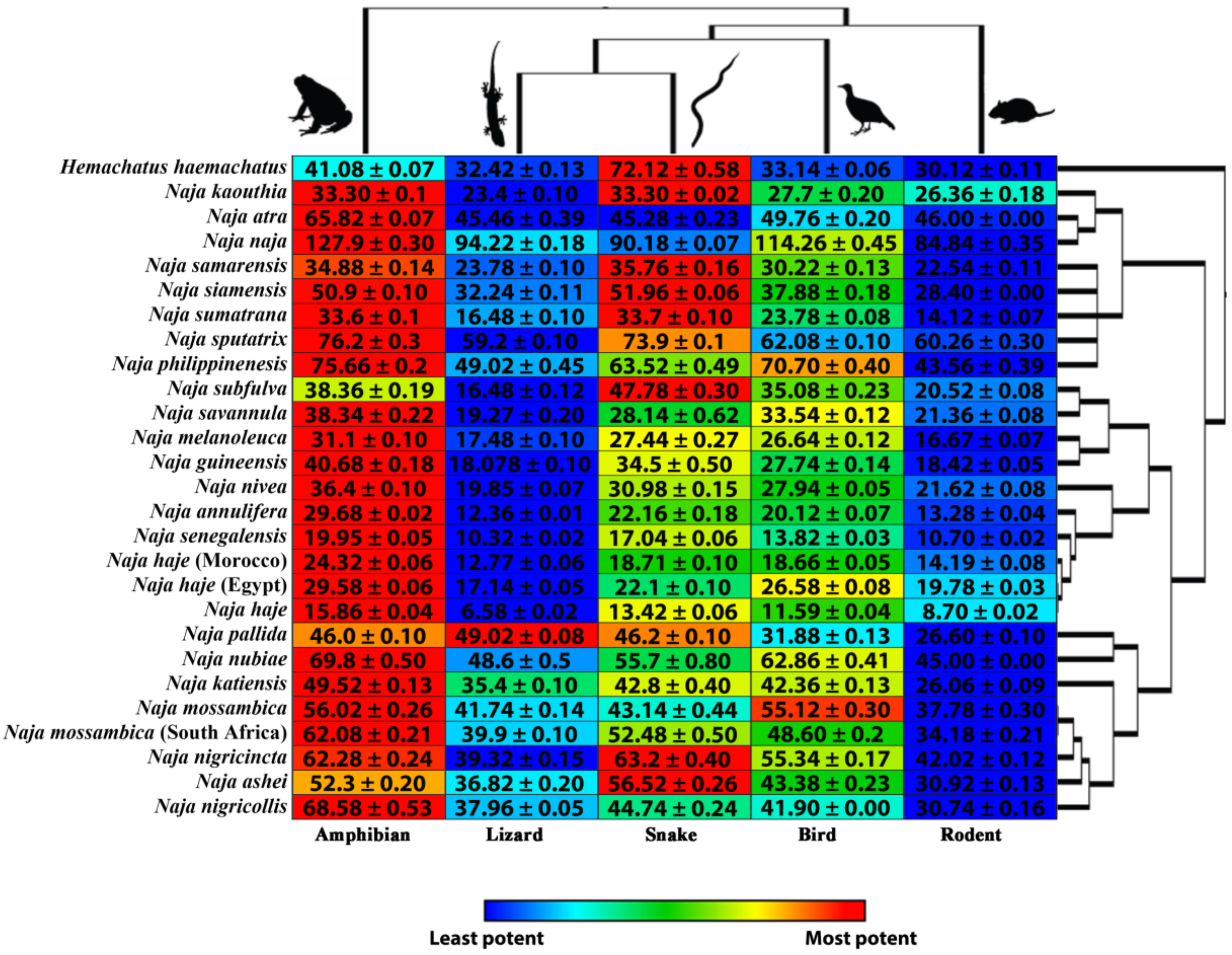
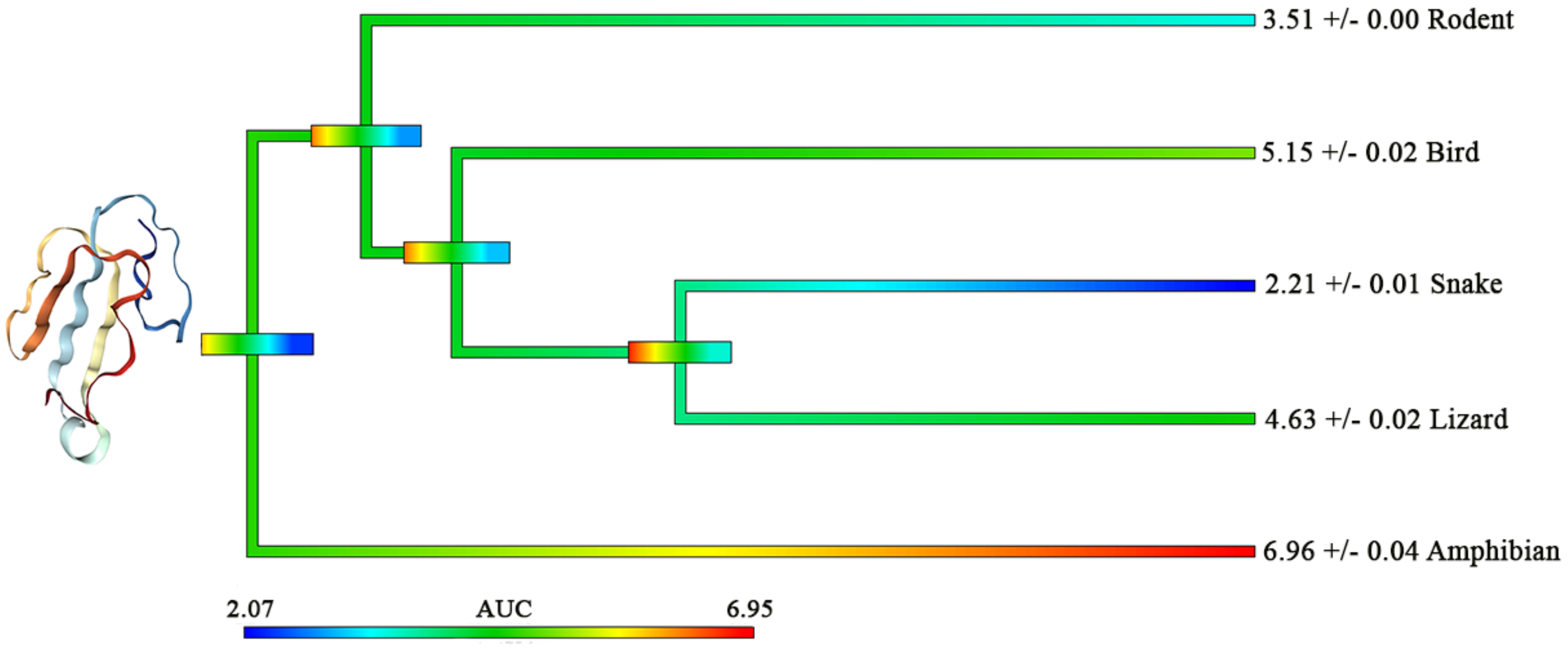
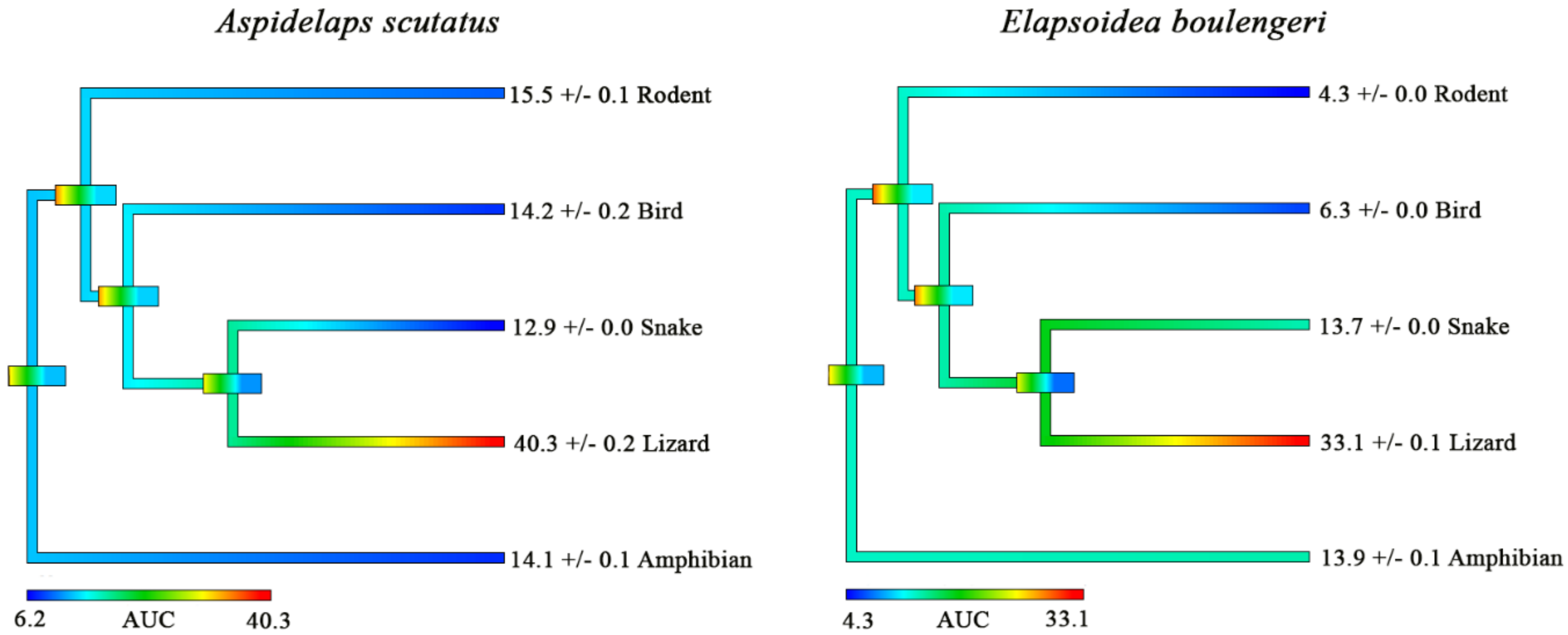
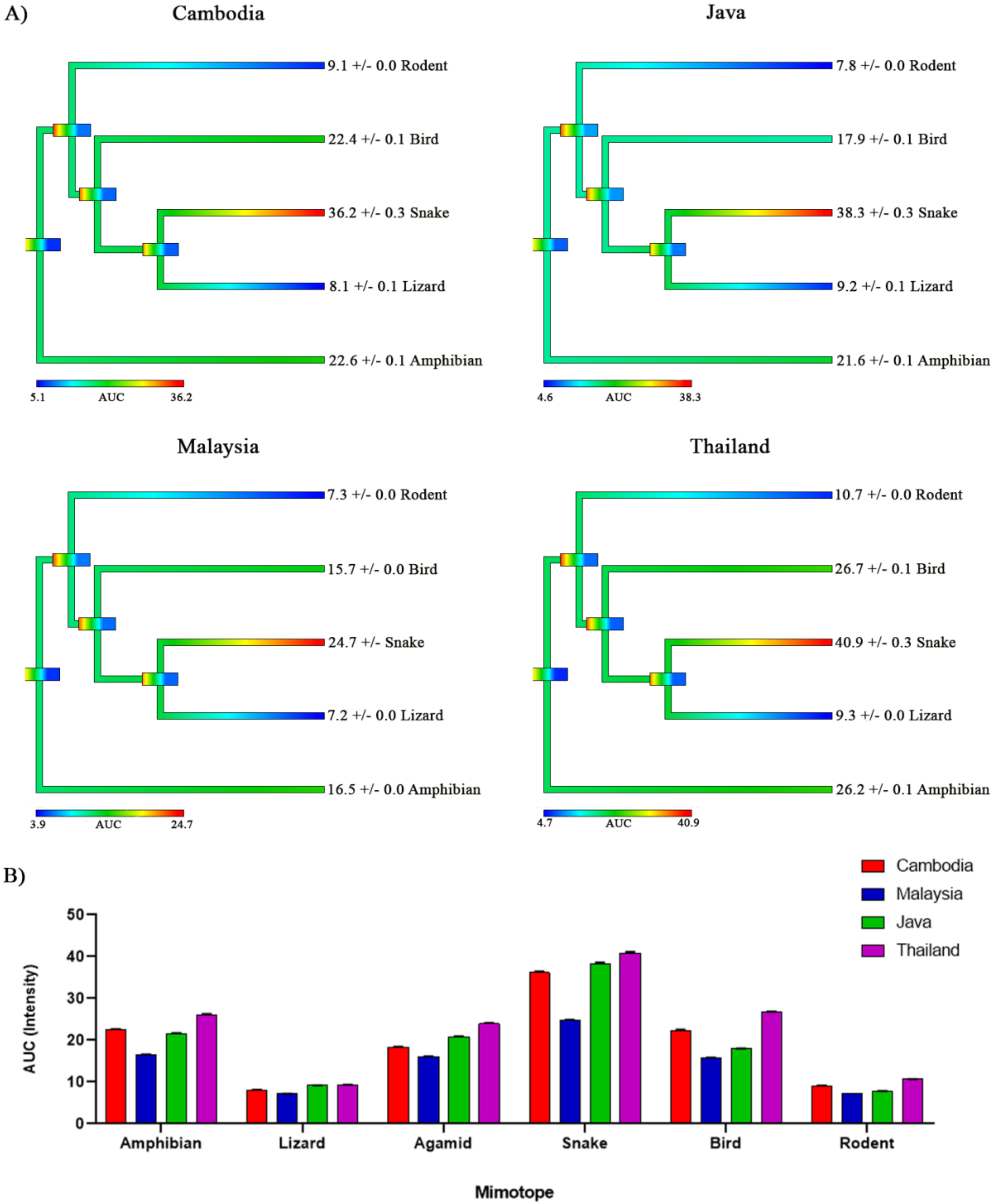
2. Results and Discussion
3. Materials and Methods
3.1. Venom Collection and Preparation
3.2. Mimotope Production and Preparation
3.3. Biolayer Interferometry (BLI)
3.4. Data Processing and Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Van Valen, L. A new evolutionary law. Evol. Theory 1973, 1, 1–30. [Google Scholar]
- Sousa, L.; Zdenek, C.; Dobson, J.; Coimbra, F.; Gillett, A.; Del-Rei, T.; Chalkidis, H.; Sant’Anna, S.; Teixeira-da-Rocha, M.; Grego, K.; et al. Coagulotoxicity of Bothrops (lancehead pit-vipers) venoms from Brazil: Differential biochemistry and antivenom efficacy resulting from prey-driven venom variation. Toxins 2018, 10, 411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youngman, N.J.; Zdenek, C.N.; Dobson, J.S.; Bittenbinder, M.A.; Gillett, A.; Hamilton, B.; Dunstan, N.; Allen, L.; Veary, A.; Veary, E.; et al. Mud in the blood: Novel potent anticoagulant coagulotoxicity in the venoms of the Australian elapid snake genus Denisonia (mud adders) and relative antivenom efficacy. Toxicol. Lett. 2019, 302, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barlow, A.; Pook, C.E.; Harrison, R.A.; Wüster, W. Coevolution of diet and prey-specific venom activity supports the role of selection in snake venom evolution. Proc. R. Soc. B Biol. Sci. 2009, 276, 2443–2449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Silva, N.J.; Aird, S.D. Prey specificity, comparative lethality and compositional differences of coral snake venoms. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2001, 128, 425–456. [Google Scholar]
- Dutertre, S.; Jin, A.-H.; Vetter, I.; Hamilton, B.; Sunagar, K.; Lavergne, V.; Dutertre, V.; Fry, B.G.; Antunes, A.; Venter, D.J.; et al. Evolution of separate predation-and defence-evoked venoms in carnivorous cone snails. Nat. Commun. 2014, 5, 3521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, M.E. Agatoxins: Ion channel specific toxins from the American funnel web spider, Agelenopsis aperta. Toxicon 2004, 43, 509–525. [Google Scholar] [CrossRef]
- Fainzilber, M.; Gordon, D.; Hasson, A.; Spira, M.E.; Zlotkin, E. Mollusc-specific toxins from the venom of Conus textile neovicarius. Eur. J. Biochem. 1991, 202, 589–595. [Google Scholar] [CrossRef]
- Heyborne, W.H.; Mackessy, S.P. Identification and characterization of a taxon-specific three-finger toxin from the venom of the Green Vinesnake (Oxybelis fulgidus; family Colubridae). Biochimie 2013, 95, 1923–1932. [Google Scholar] [CrossRef]
- Modahl, C.M.; Mrinalini; Frietze, S.; Mackessy, S.P. Adaptive evolution of distinct prey-specific toxin genes in rear-fanged snake venom. Proc. R. Soc. B Biol. Sci. 2018, 285, 20181003. [Google Scholar] [CrossRef]
- Pawlak, J.; Mackessy, S.P.; Fry, B.G.; Bhatia, M.; Mourier, G.; Fruchart-Gaillard, C.; Servent, D.; Ménez, R.; Stura, E.; Ménez, A.; et al. Denmotoxin, a three-finger toxin from the colubrid snake Boiga dendrophila (Mangrove Catsnake) with bird-specific activity. J. Biol. Chem. 2006, 281, 29030–29041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawlak, J.; Mackessy, S.P.; Sixberry, N.M.; Stura, E.A.; Le Du, M.H.; Ménez, R.; Foo, C.S.; Ménez, A.; Nirthanan, S.; Kini, R.M.; et al. Irditoxin, a novel covalently linked heterodimeric three-finger toxin with high taxon-specific neurotoxicity. FASEB J. 2009, 23, 534–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, J.; Agwa, A.J.; Szanto, T.G.; Csóti, A.; Panyi, G.; Schroeder, C.I.; Walker, A.A.; King, G.F. Weaponisation ‘on the fly’: Convergent recruitment of knottin and defensin peptide scaffolds into the venom of predatory assassin flies. Insect Biochem. Mol. Biol. 2020, 118, 103310. [Google Scholar] [CrossRef] [PubMed]
- Dashevsky, D.; Fry, B.G. Ancient diversification of three-finger toxins in Micrurus coral snakes. J. Mol. Evol. 2018, 86, 58–67. [Google Scholar] [CrossRef]
- Kordiš, D.; Gubenšek, F. Adaptive evolution of animal toxin multigene families. Gene 2000, 261, 43–52. [Google Scholar] [CrossRef]
- Lynch, V.J. Inventing an arsenal: Adaptive evolution and neofunctionalization of snake venom phospholipase A 2 genes. BMC Evol. Biol. 2007, 7, 2. [Google Scholar] [CrossRef] [Green Version]
- Kini, R.M.; Evans, H.J. A model to explain the pharmacological effects of snake venom phospholipases A2. Toxicon 1989, 27, 613–635. [Google Scholar] [CrossRef]
- Jackson, T.; Sunagar, K.; Undheim, E.; Koludarov, I.; Chan, A.; Sanders, K.; Ali, S.; Hendrikx, I.; Dunstan, N.; Fry, B.G.; et al. Venom down under: Dynamic evolution of Australian elapid snake toxins. Toxins 2013, 5, 2621–2655. [Google Scholar] [CrossRef] [Green Version]
- Fry, B.G.; Sunagar, K.; Casewell, N.R.; Kochva, E.; Roelants, K.; Schieb, H.; Wüster, W.; Vidal, N.; Young, B.; Burbink, F.; et al. The origin and evolution of the Toxicofera reptile venom system. In Venomous Reptiles and Their Toxins: Evolution, Pathophysiology and Biodiscovery; Fry, B.G., Ed.; Oxford University Press: New York, NY, USA, 2015; pp. 15–21. [Google Scholar]
- Kini, R.M. Excitement ahead: Structure, function and mechanism of snake venom phospholipase A2 enzymes. Toxicon 2003, 42, 827–840. [Google Scholar] [CrossRef]
- Chang, C.C.; Lee, J.D. Crotoxin, the neurotoxin of South American rattlesnake venom, is a presynaptic toxin acting like β-bungarotoxin. Naunyn-Schmiedeberg’s Arch. Pharmacol. 1977, 296, 159–168. [Google Scholar] [CrossRef]
- Chang, C.C.; Lee, J.D.; Eaker, D.; Fohlman, J. The presynaptic neuromuscular blocking action of taipoxin. A comparison with β-bungarotoxin and crotoxin. Toxicon 1977, 15, 571–576. [Google Scholar] [CrossRef]
- Su, M.; Chang, C. Presynaptic effects of snake venom toxins which have phospholipase A2 activity (β-bungarotoxin, taipoxin, crotoxin). Toxicon 1984, 22, 631–640. [Google Scholar] [CrossRef]
- Hart, A.J.; Isbister, G.K.; O’Donnell, P.; Williamson, N.A.; Hodgson, W.C. Species differences in the neuromuscular activity of post-synaptic neurotoxins from two Australian black snakes (Pseudechis porphyriacus and Pseudechis colletti). Toxicol. Lett. 2013, 219, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Modahl, C.M.; Mukherjee, A.K.; Mackessy, S.P. An analysis of venom ontogeny and prey-specific toxicity in the Monocled Cobra (Naja kaouthia). Toxicon 2016, 119, 8–20. [Google Scholar] [CrossRef]
- Barber, C.M.; Isbister, G.K.; Hodgson, W.C. Alpha neurotoxins. Toxicon 2013, 66, 47–58. [Google Scholar] [CrossRef]
- Nirthanan, S.; Gwee, M.C. Three-finger α-neurotoxins and the nicotinic acetylcholine receptor, forty years on. J. Pharmacol. Sci. 2004, 94, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Fry, B.G.; Lumsden, N.G.; Wüster, W.; Wickramaratna, J.C.; Hodgson, W.C.; Kini, R.M. Isolation of a neurotoxin (α-colubritoxin) from a nonvenomous colubrid: Evidence for early origin of venom in snakes. J. Mol. Evol. 2003, 57, 446–452. [Google Scholar] [CrossRef]
- Chang, C. The action of snake venoms on nerve and muscle. In Snake Venoms; Lee, C.Y., Ed.; Springer: Heidelberg, Germany, 1979; Volume 52, pp. 309–376. [Google Scholar]
- Servent, D.; Ménez, A. Snake neurotoxins that interact with nicotinic acetylcholine receptors. In Handbook of Neurotoxicology; Massaro, E.J., Ed.; Humana Press: Totowa, NJ, USA, 2002; Volume 1, pp. 385–425. [Google Scholar]
- Mackessy, S.P.; Sixberry, N.M.; Heyborne, W.H.; Fritts, T. Venom of the Brown Treesnake, Boiga irregularis: Ontogenetic shifts and taxa-specific toxicity. Toxicon 2006, 47, 537–548. [Google Scholar] [CrossRef]
- Zdenek, C.N.; Harris, R.J.; Kuruppu, S.; Youngman, N.J.; Dobson, J.S.; Debono, J.; Khan, M.; Smith, I.; Yarski, M.; Harrich, D.; et al. A Taxon-Specific and High-Throughput Method for Measuring Ligand Binding to Nicotinic Acetylcholine Receptors. Toxins 2019, 11, 600. [Google Scholar] [CrossRef] [Green Version]
- Layloo, I.; Smith, C.; Maritz, B. Diet and feeding in the Cape Cobra, Naja nivea. Afr. J. Herpetol. 2017, 66, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Luiselli, L. The ghost of a recent invasion in the reduced feeding rates of spitting cobras during the dry season in a rainforest region of tropical Africa? Acta Oecologica 2001, 22, 311–314. [Google Scholar] [CrossRef]
- Luiselli, L.; Angelici, F.M.; Akani, G.C. Comparative feeding strategies and dietary plasticity of the sympatric cobras Naja melanoleuca and Naja nigricollis in three diverging Afrotropical habitats. Can. J. Zool. 2002, 80, 55–63. [Google Scholar] [CrossRef]
- Maritz, B.; Alexander, G.J.; Maritz, R.A. The underappreciated extent of cannibalism and ophiophagy in African cobras. Ecology 2019, 100, e02522. [Google Scholar] [CrossRef] [PubMed]
- Shine, R.; Branch, W.; Webb, J.; Harlow, P.S.; Shine, T.; Keogh, J.S. Ecology of cobras from southern Africa. J. Zool. 2007, 272, 183–193. [Google Scholar] [CrossRef]
- Kgaditse, M.M. The evolution and diversification of diet in elapids. Master’s Thesis, University of the Witwatersrand, Johannesburg, South Africa, 22 July 2016. [Google Scholar]
- Lee, M.S.; Sanders, K.L.; King, B.; Palci, A. Diversification rates and phenotypic evolution in venomous snakes (Elapidae). R. Soc. Open Sci. 2016, 3, 150277. [Google Scholar] [CrossRef] [Green Version]
- Wüster, W.; Chirio, L.; Trape, J.-F.; Ineich, I.; Jackson, K.; Greenbaum, E.; Barron, C.; Kusamba, C.; Nagy, Z.T.; Storey, R.; et al. Integration of nuclear and mitochondrial gene sequences and morphology reveals unexpected diversity in the forest cobra (Naja melanoleuca) species complex in Central and West Africa (Serpentes: Elapidae). Zootaxa 2018, 4455, 68–98. [Google Scholar] [CrossRef]
- Wüster, W.; Crookes, S.; Ineich, I.; Mané, Y.; Pook, C.E.; Trape, J.-F.; Broadley, D.G. The phylogeny of cobras inferred from mitochondrial DNA sequences: Evolution of venom spitting and the phylogeography of the African spitting cobras (Serpentes: Elapidae: Naja nigricollis complex). Mol. Phylogenetics Evol. 2007, 45, 437–453. [Google Scholar] [CrossRef]
- Bittenbinder, M.A.; Zdenek, C.N.; Youngman, N.J.; Dobson, J.S.; Naude, A.; Vonk, F.J.; Fry, B.G. Coagulotoxic cobras: Clinical implications of strong anticoagulant actions of African spitting Naja Venoms that are not neutralised by antivenom but are by LY315920 (Varespladib). Toxins 2018, 10, 516. [Google Scholar] [CrossRef] [Green Version]
- Bhat, M.K.; Gowda, T.V. Purification and characterization of a myotoxic phospholipase A2 from Indian cobra (Naja naja naja) venom. Toxicon 1989, 27, 861–873. [Google Scholar] [CrossRef]
- Lyons, K.; Dugon, M.M.; Healy, K. Diet breadth mediates the prey specificity of venom potency in snakes. Toxins 2020, 12, 74. [Google Scholar] [CrossRef] [Green Version]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The protein data bank. Nucleic Acids Res 2000, 235–242. Available online: http://www.rcsb.org/. [CrossRef] [PubMed] [Green Version]
- Walkinshaw, M.D.; Saenger, W.; Maelicke, A. Three-dimensional structure of the” long” neurotoxin from cobra venom. Proc. Natl. Acad. Sci. USA 1980, 77, 2400–2404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shine, R.; Haagner, G.V.; Branch, W.R.; Harlow, P.S.; Webb, J.K. Natural history of the African shieldnose snake Aspidelaps scutatus (Serpentes, Elapidae). J. Herpetol. 1996, 30, 361–366. [Google Scholar] [CrossRef] [Green Version]
- Chanhome, L.; Jintakune, P.; Wilde, H.; Cox, M.J. Venomous snake husbandry in Thailand. Wilderness Environ. Med. 2001, 12, 17–23. [Google Scholar] [CrossRef] [Green Version]
- O’Shea, M. Venomous Snakes of the World; Princeton University Press: Princeton, NJ, USA, 2005; pp. 96–97. [Google Scholar]
- Fry, B.; Hendrikx, I.; Rowley, P.; Jackson, T.; Van der Ploeg, H.; Johnson, R.; Sasa, M.; Dunstan, N.; Barve, S.; Lock, B.; et al. Maintaining venomous reptile collections. In Venomous Reptiles and Their Toxins: Evolution, Pathophysiology and Biodiscovery; Fry, B.G., Ed.; Oxford University Press: New York, NY, USA, 2015; p. 89. [Google Scholar]
- Vonk, F.J.; Casewell, N.R.; Henkel, C.V.; Heimberg, A.M.; Jansen, H.J.; McCleary, R.J.; Kerkkamp, H.M.; Vos, R.A.; Guerreiro, I.; Calvete, J.J.; et al. The king cobra genome reveals dynamic gene evolution and adaptation in the snake venom system. Proc. Natl. Acad. Sci. USA 2013, 110, 20651–20656. [Google Scholar] [CrossRef] [Green Version]
- Panagides, N.; Jackson, T.; Ikonomopoulou, M.; Arbuckle, K.; Pretzler, R.; Yang, D.; Ali, S.; Koludarov, I.; Dobson, J.; Sanker, B.; et al. How the cobra got its flesh-eating venom: Cytotoxicity as a defensive innovation and its co-evolution with hooding, aposematic marking, and spitting. Toxins 2017, 9, 103. [Google Scholar] [CrossRef] [Green Version]
- Bracci, L.; Lozzi, L.; Lelli, B.; Pini, A.; Neri, P. Mimotopes of the nicotinic receptor binding site selected by a combinatorial peptide library. Biochemistry 2001, 40, 6611–6619. [Google Scholar] [CrossRef]
- Bracci, L.; Lozzi, L.; Pini, A.; Lelli, B.; Falciani, C.; Niccolai, N.; Bernini, A.; Spreafico, A.; Soldani, P.; Neri, P.; et al. A branched peptide mimotope of the nicotinic receptor binding site is a potent synthetic antidote against the snake neurotoxin α-bungarotoxin. Biochemistry 2002, 41, 10194–10199. [Google Scholar] [CrossRef]
- Chiappinelli, V.A.; Weaver, W.R.; McLane, K.E.; Conti-Fine, B.M.; Fiordalisi, J.J.; Grant, G.A. Binding of native κ-neurotoxins and site-directed mutants to nicotinic acetylcholine receptors. Toxicon 1996, 34, 1243–1256. [Google Scholar] [CrossRef]
- Katchalski-Katzir, E.; Kasher, R.; Balass, M.; Scherf, T.; Harel, M.; Fridkin, M.; Sussman, J.L.; Fuchs, S. Design and synthesis of peptides that bind α-bungarotoxin with high affinity and mimic the three-dimensional structure of the binding-site of acetylcholine receptor. Biophys. Chem. 2002, 100, 293–305. [Google Scholar] [CrossRef]
- McLane, K.E.; Wu, X.; Conti-Tronconi, B.M. An α-bungarotoxin-binding sequence on the Torpedo nicotinic acetylcholine receptor α-subunit: Conservative amino acid substitutions reveal side-chain specific interactions. Biochemistry 1994, 33, 2576–2585. [Google Scholar] [CrossRef] [PubMed]
- McLane, K.E.; Wu, X.; Diethelm, B.; Conti-Tronconi, B.M. Structural determinants of α-bungarotoxin binding to the sequence segment 181-200 of the muscle nicotinic acetylcholine receptor. α-subunit: Effects of cysteine/cystine modification and species-specific amino acid substitutions. Biochemistry 1991, 30, 4925–4934. [Google Scholar] [CrossRef] [PubMed]
- Tzartos, S.; Remoundos, M.S. Fine localization of the major alpha-bungarotoxin binding site to residues alpha 189-195 of the Torpedo acetylcholine receptor. Residues 189, 190, and 195 are indispensable for binding. J. Biol. Chem. 1990, 265, 21462–21467. [Google Scholar] [PubMed]
- Testai, F.D.; Venera, G.D.; Peña, C.; de Jiménez Bonino, M.J.B. Histidine 186 of the nicotinic acetylcholine receptor α subunit requires the presence of the 192–193 disulfide bridge to interact with α-bungarotoxin. Neurochem. Int. 2000, 36, 27–33. [Google Scholar] [CrossRef]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of phylogenetics and evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef] [Green Version]
- Revell, L.J. phytools: An R package for phylogenetic comparative biology (and other things). Methods Ecol. Evol. 2012, 3, 217–223. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harris, R.J.; Zdenek, C.N.; Harrich, D.; Frank, N.; Fry, B.G. An Appetite for Destruction: Detecting Prey-Selective Binding of α-Neurotoxins in the Venom of Afro-Asian Elapids. Toxins 2020, 12, 205. https://doi.org/10.3390/toxins12030205
Harris RJ, Zdenek CN, Harrich D, Frank N, Fry BG. An Appetite for Destruction: Detecting Prey-Selective Binding of α-Neurotoxins in the Venom of Afro-Asian Elapids. Toxins. 2020; 12(3):205. https://doi.org/10.3390/toxins12030205
Chicago/Turabian StyleHarris, Richard J., Christina N. Zdenek, David Harrich, Nathaniel Frank, and Bryan G. Fry. 2020. "An Appetite for Destruction: Detecting Prey-Selective Binding of α-Neurotoxins in the Venom of Afro-Asian Elapids" Toxins 12, no. 3: 205. https://doi.org/10.3390/toxins12030205
APA StyleHarris, R. J., Zdenek, C. N., Harrich, D., Frank, N., & Fry, B. G. (2020). An Appetite for Destruction: Detecting Prey-Selective Binding of α-Neurotoxins in the Venom of Afro-Asian Elapids. Toxins, 12(3), 205. https://doi.org/10.3390/toxins12030205