Fully Human Monoclonal Antibodies Effectively Neutralizing Botulinum Neurotoxin Serotype B

 , , ,
, , ,
Abstract
:
1. Introduction
2. Results
2.1. Immunization with Tetravalent Botulinum Toxoid Vaccine
2.2. Preparation of HuMAbs
2.3. Binding Specificity of HuMAbs
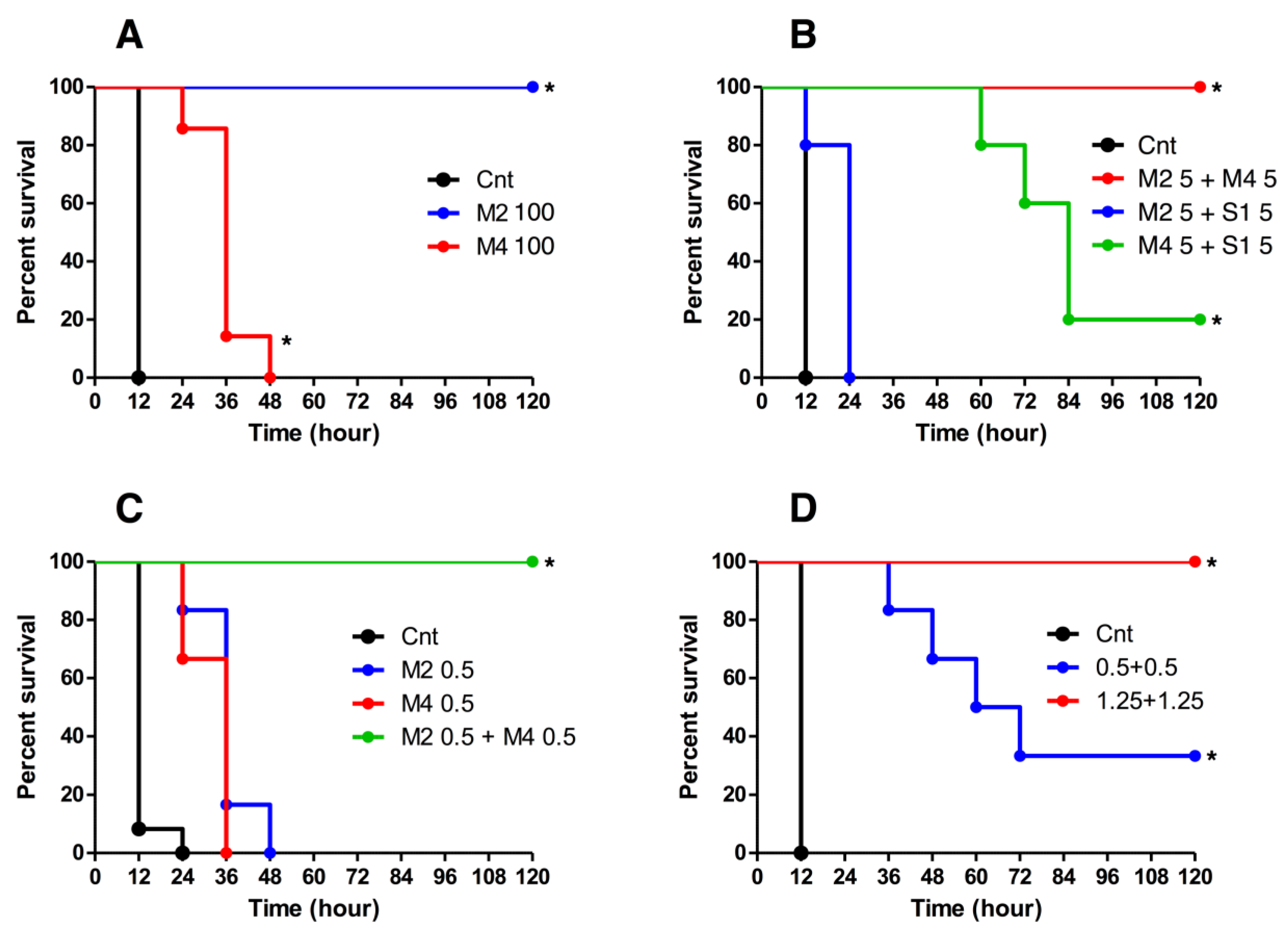
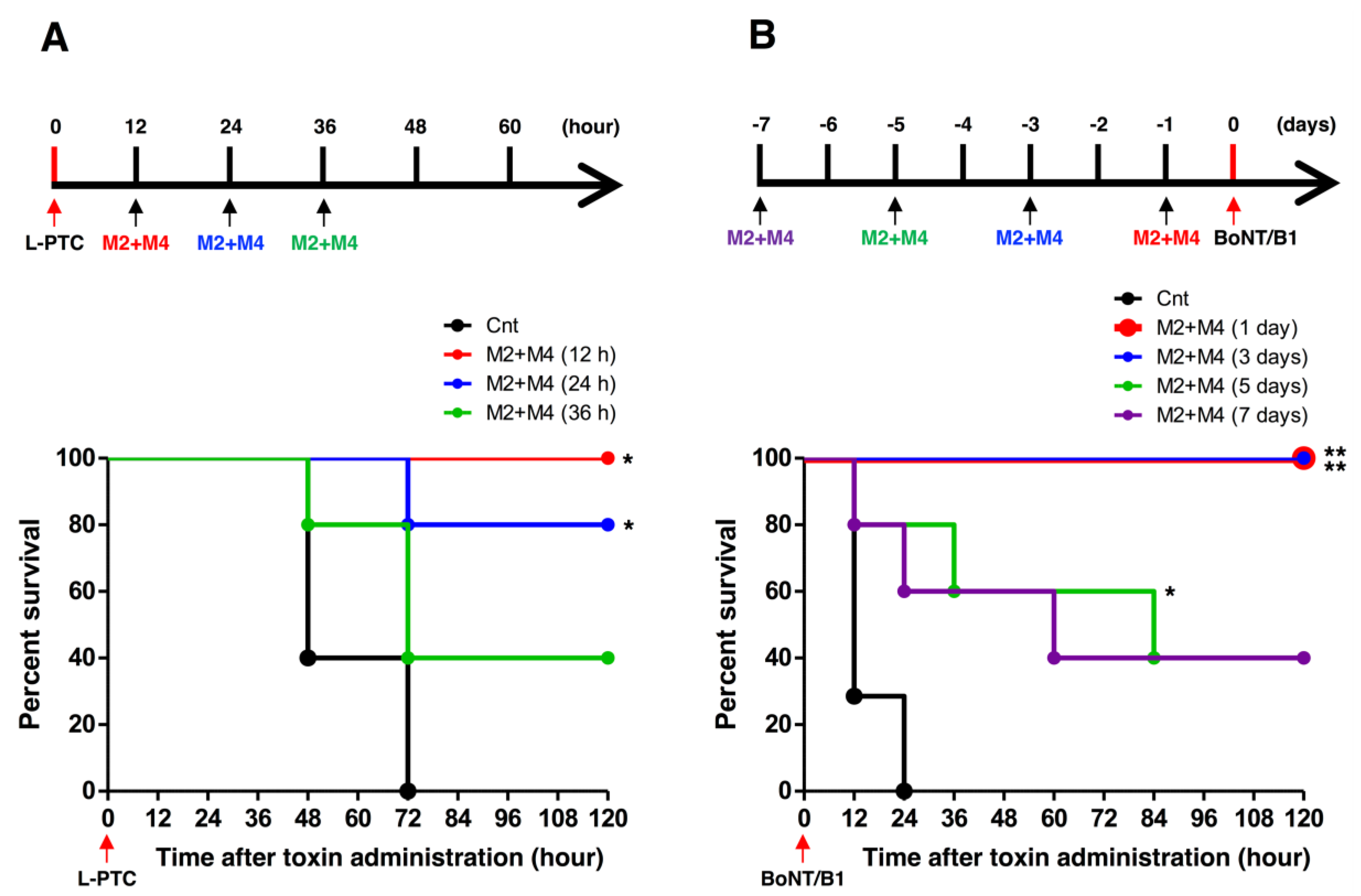
2.4. Neutralization Activity of HuMAbs
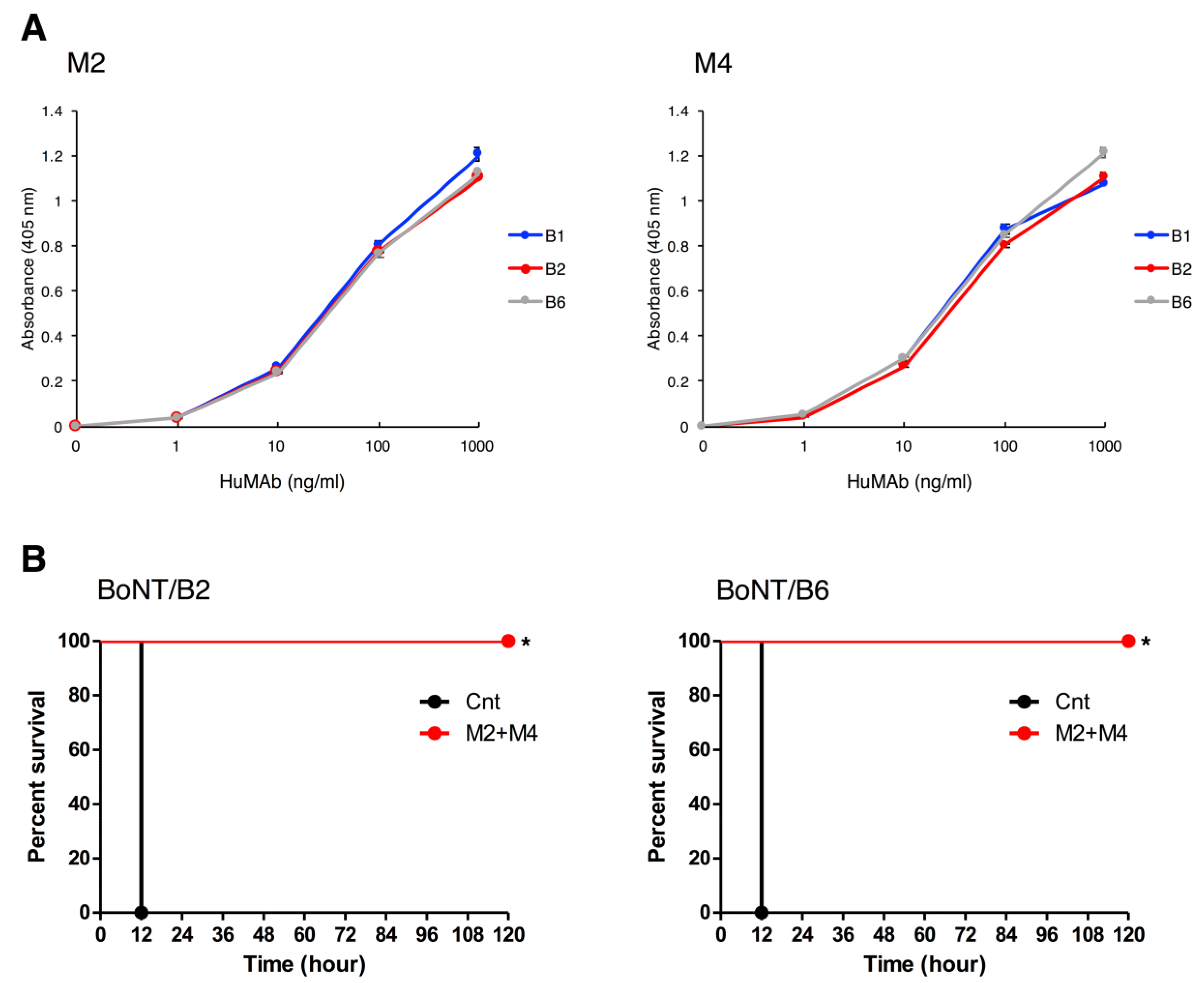
2.5. Binding and Neutralization Activity of HuMAbs against Subtypes BoNT/B2 and BoNT/B6
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Ethics Statement
5.2. Preparation of BoNT/A and BoNT/B
5.3. Inoculation of Botulinum Toxoid Vaccine
5.4. Preparation of HuMAbs
5.5. Preparation of Recombinant Proteins
5.6. Binding Assay Using ELISA
5.7. Competition ELISA
5.8. Sequencing of HuMAb Variable Region Gene Segments
5.9. Neutralization Test (Mouse Bioassay)
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schiavo, G.; Matteoli, M.; Montecucco, C. Neurotoxins affecting neuroexocytosis. Physiol. Rev. 2000, 80, 717–766. [Google Scholar] [CrossRef]
- Rossetto, O.; Pirazzini, M.; Montecucco, C. Botulinum neurotoxins: Genetic, structural and mechanistic insights. Nat. Rev. Genet. 2014, 12, 535–549. [Google Scholar] [CrossRef] [PubMed]
- Froude, J.W.; Stiles, B.G.; Pelat, T.; Thullier, P. Antibodies for biodefense. mAbs 2011, 3, 517–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, K.; Kohda, T.; Shibata, Y.; Tsukamoto, K.; Arimitsu, H.; Hayashi, M.; Mukamoto, M.; Sasakawa, N.; Kozaki, S. Unique Biological Activity of Botulinum D/C Mosaic Neurotoxin in Murine Species. Infect. Immun. 2012, 80, 2886–2893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Berntsson, R.; Tepp, W.H.; Tao, L.; Johnson, E.A.; Stenmark, P.; Dong, M. Structural basis for the unique ganglioside and cell membrane recognition mechanism of botulinum neurotoxin DC. Nat. Commun. 2017, 8, 1637. [Google Scholar] [CrossRef]
- Barash, J.R.; Arnon, S.S. A Novel Strain of Clostridium botulinum That Produces Type B and Type H Botulinum Toxins. J. Infect. Dis. 2013, 209, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Dover, N.; Barash, J.R.; Hill, K.; Xie, G.; Arnon, S.S. Molecular Characterization of a Novel Botulinum Neurotoxin Type H Gene. J. Infect. Dis. 2013, 209, 192–202. [Google Scholar] [CrossRef]
- Maslanka, S.E.; Lúquez, C.; Dykes, J.K.; Tepp, W.H.; Pier, C.L.; Pellett, S.; Raphael, B.; Kalb, S.R.; Barr, J.R.; Rao, A.; et al. A Novel Botulinum Neurotoxin, Previously Reported as Serotype H, Has a Hybrid-Like Structure With Regions of Similarity to the Structures of Serotypes A and F and Is Neutralized With Serotype A Antitoxin. J. Infect. Dis. 2015, 213, 379–385. [Google Scholar] [CrossRef]
- Fan, Y.; Barash, J.R.; Lou, J.; Conrad, F.; Marks, J.D.; Arnon, S.S. Immunological Characterization and Neutralizing Ability of Monoclonal Antibodies Directed Against Botulinum Neurotoxin Type, H. J. Infect. Dis. 2016, 213, 1606–1614. [Google Scholar] [CrossRef] [Green Version]
- Yao, G.; Lam, K.-H.; Perry, K.; Weisemann, J.; Rummel, A.; Jin, R. Crystal Structure of the Receptor-Binding Domain of Botulinum Neurotoxin Type HA, Also Known as Type FA or H. Toxins 2017, 9, 93. [Google Scholar] [CrossRef] [Green Version]
- Lam, K.-H.; Sikorra, S.; Weisemann, J.; Maatsch, H.; Perry, K.; Rummel, A.; Binz, T.; Jin, R. Structural and biochemical characterization of the protease domain of the mosaic botulinum neurotoxin type HA. Pathog. Dis. 2018, 76, 76. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Barash, J.R.; Conrad, F.; Lou, J.; Tam, C.; Cheng, L.W.; Arnon, S.S.; Marks, J.D. The Novel Clostridial Neurotoxin Produced by Strain IBCA10-7060 Is Immunologically Equivalent to BoNT/HA. Toxins 2019, 12, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Masuyer, G.; Zhang, J.; Shen, Y.; Lundin, D.; Henriksson, L.; Miyashita, S.-I.; Martínez-Carranza, M.; Dong, M.; Stenmark, P. Identification and characterization of a novel botulinum neurotoxin. Nat. Commun. 2017, 8, 14130. [Google Scholar] [CrossRef] [PubMed]
- Zornetta, I.; Tehran, D.A.; Arrigoni, G.; Anniballi, F.; Bano, L.; Leka, O.; Zanotti, G.; Binz, T.; Montecucco, C. The first non Clostridial botulinum-like toxin cleaves VAMP within the juxtamembrane domain. Sci. Rep. 2016, 6, 30257. [Google Scholar] [CrossRef]
- Brunt, J.; Carter, A.T.A.T.; Stringer, S.C.S.C.; Peck, M. Identification of a novel botulinum neurotoxin gene cluster in Enterococcus. FEBS Lett. 2018, 592, 310–317. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Lebreton, F.; Mansfield, M.J.; Miyashita, S.-I.; Zhang, J.; Schwartzman, J.A.; Tao, L.; Masuyer, G.; Martínez-Carranza, M.; Stenmark, P.; et al. Identification of a Botulinum Neurotoxin-like Toxin in a Commensal Strain of Enterococcus faecium. Cell Host Microbe 2018, 23, 169–176.e6. [Google Scholar] [CrossRef] [Green Version]
- Tehran, D.A.; Pirazzini, M. Novel Botulinum Neurotoxins: Exploring Underneath the Iceberg Tip. Toxins 2018, 10, 190. [Google Scholar] [CrossRef] [Green Version]
- Arnon, S.S.; Schechter, R.; Inglesby, T.V.; Henderson, D.A.; Bartlett, J.G.; Ascher, M.S.; Eitzen, E.; Fine, A.D.; Hauer, J.; Layton, M.; et al. Botulinum toxin as a biological weapon: Medical and public health management. JAMA 2001, 285, 1059–1070. [Google Scholar] [CrossRef]
- Matsumura, T.; Sugawara, Y.; Yutani, M.; Amatsu, S.; Yagita, H.; Kohda, T.; Fukuoka, S.-I.; Nakamura, Y.; Fukuda, S.; Hase, K.; et al. Botulinum toxin A complex exploits intestinal M cells to enter the host and exert neurotoxicity. Nat. Commun. 2015, 6, 6255. [Google Scholar] [CrossRef] [Green Version]
- Dong, M. SV2 Is the Protein Receptor for Botulinum Neurotoxin, A. Science 2006, 312, 592–596. [Google Scholar] [CrossRef]
- Dong, M.; Liu, H.; Tepp, W.H.; Johnson, E.A.; Janz, R.; Chapman, E.R. Glycosylated SV2A and SV2B Mediate the Entry of Botulinum Neurotoxin E into Neurons. Mol. Boil. Cell 2008, 19, 5226–5237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Z.; Chen, C.; Barbieri, J.T.; Kim, J.-J.P.; Baldwin, M.R. Glycosylated SV2 and Gangliosides as Dual Receptors for Botulinum Neurotoxin Serotype, F. Biochemistry 2009, 48, 5631–5641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishiki, T.; Kamata, Y.; Nemoto, Y.; Omori, A.; Ito, T.; Takahashi, M.; Kozaki, S. Identification of protein receptor for Clostridium botulinum type B neurotoxin in rat brain synaptosomes. J. Boil. Chem. 1994, 269, 10498–10503. [Google Scholar]
- Nishiki, T.-I.; Tokuyama, Y.; Kamata, Y.; Nemoto, Y.; Yoshida, A.; Sato, K.; Sekiguchi, M.; Takahashi, M.; Kozaki, S. The high-affinity binding ofClostridium botulinumtype B neurotoxin to synaptotagmin II associated with gangliosides GT1b/GD1a. FEBS Lett. 1996, 378, 253–257. [Google Scholar] [CrossRef] [Green Version]
- Hoch, D.H.; Romero-Mira, M.; Ehrlich, B.E.; Finkelstein, A.; Dasgupta, B.R.; Simpson, L.L. Channels formed by botulinum, tetanus, and diphtheria toxins in planar lipid bilayers: Relevance to translocation of proteins across membranes. Proc. Natl. Acad. Sci. USA 1985, 82, 1692–1696. [Google Scholar] [CrossRef] [Green Version]
- Koriazova, L.K.; Montal, M. Translocation of botulinum neurotoxin light chain protease through the heavy chain channel. Nat. Genet. 2003, 10, 13–18. [Google Scholar] [CrossRef]
- Pirazzini, M.; Tehran, D.A.; Zanetti, G.; Rossetto, O.; Montecucco, C. Hsp90 and Thioredoxin-Thioredoxin Reductase enable the catalytic activity of Clostridial neurotoxins inside nerve terminals. Toxicon 2018, 147, 32–37. [Google Scholar] [CrossRef]
- Arnon, S.S.; Schechter, R.; Maslanka, S.E.; Jewell, N.P.; Hatheway, C.L. Human Botulism Immune Globulin for the Treatment of Infant Botulism. N. Engl. J. Med. 2006, 354, 462–471. [Google Scholar] [CrossRef]
- Chen, C.; Wang, S.; Wang, H.; Mao, X.; Zhang, T.; Ji, G.; Shi, X.; Xia, T.; Lu, W.; Zhang, D.; et al. Potent Neutralization of Botulinum Neurotoxin/B by Synergistic Action of Antibodies Recognizing Protein and Ganglioside Receptor Binding Domain. PLoS ONE 2012, 7, e43845. [Google Scholar] [CrossRef]
- Nayak, S.U.; Griffiss, J.M.; McKenzie, R.; Fuchs, E.J.; Jurao, R.A.; An, A.T.; Ahene, A.; Tomic, M.; Hendrix, C.W.; Zenilman, J.M. Safety and Pharmacokinetics of XOMA 3AB, a Novel Mixture of Three Monoclonal Antibodies against Botulinum Toxin A. Antimicrob. Agents Chemother. 2014, 58, 5047–5053. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Dong, J.; Lou, J.; Wen, W.; Conrad, F.; Geren, I.N.; Garcia-Rodriguez, C.; Smith, T.J.; Smith, L.A.; Ho, M.; et al. Monoclonal Antibodies that Inhibit the Proteolytic Activity of Botulinum Neurotoxin Serotype/B. Toxins 2015, 7, 3405–3423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Y.; Garcia-Rodriguez, C.; Lou, J.; Wen, W.; Conrad, F.; Zhai, W.; Smith, T.J.; Smith, L.A.; Marks, J.D. A three monoclonal antibody combination potently neutralizes multiple botulinum neurotoxin serotype F subtypes. PLoS ONE 2017, 12, e0174187. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rodriguez, C.; Razai, A.; Geren, I.N.; Lou, J.; Conrad, F.; Wen, W.-H.; Farr-Jones, S.; Smith, T.J.; Brown, J.L.; Skerry, J.C.; et al. A Three Monoclonal Antibody Combination Potently Neutralizes Multiple Botulinum Neurotoxin Serotype E Subtypes. Toxins 2018, 10, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snow, D.M.; Riling, K.; Kimbler, A.; Espinoza, Y.; Wong, D.; Pham, K.; Martinez, Z.; Kraus, C.N.; Conrad, F.; Garcia-Rodriguez, C.; et al. Safety and Pharmacokinetics of a Four Monoclonal Antibody Combination against Botulinum C and D Neurotoxins. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomic, M.T.; Espinoza, Y.; Martinez, Z.; Pham, K.; Cobb, R.; Snow, D.M.; Earnhart, C.G.; Pals, T.; Syar, E.S.; Niemuth, N.; et al. Monoclonal Antibody Combinations Prevent Serotype A and Serotype B Inhalational Botulism in a Guinea Pig Model. Toxins 2019, 11, 208. [Google Scholar] [CrossRef] [Green Version]
- Espinoza, Y.; Wong, D.; Ahene, A.; Der, K.; Martinez, Z.; Pham, J.; Cobb, R.; Farr-Jones, S.; Marks, J.D.; Tomic, M.T. Pharmacokinetics of Human Recombinant Anti-Botulinum Toxin Antibodies in Rats. Toxins 2019, 11, 345. [Google Scholar] [CrossRef] [Green Version]
- Kubota-Koketsu, R.; Mizuta, H.; Oshita, M.; Ideno, S.; Yunoki, M.; Kuhara, M.; Yamamoto, N.; Okuno, Y.; Ikuta, K. Broad neutralizing human monoclonal antibodies against influenza virus from vaccinated healthy donors. Biochem. Biophys. Res. Commun. 2009, 387, 180–185. [Google Scholar] [CrossRef]
- Pan, Y.; Sasaki, T.; Kubota-Koketsu, R.; Inoue, Y.; Yasugi, M.; Yamashita, A.; Ramadhany, R.; Arai, Y.; Du, A.; Boonsathorn, N.; et al. Human monoclonal antibodies derived from a patient infected with 2009 pandemic influenza A virus broadly cross-neutralize group 1 influenza viruses. Biochem. Biophys. Res. Commun. 2014, 450, 42–48. [Google Scholar] [CrossRef] [Green Version]
- Misaki, R.; Fukura, N.; Kajiura, H.; Yasugi, M.; Kubota-Koketsu, R.; Sasaki, T.; Momota, M.; Ono, K.-I.; Ohashi, T.; Ikuta, K.; et al. Recombinant production and characterization of human anti-influenza virus monoclonal antibodies identified from hybridomas fused with human lymphocytes. Biologicals 2016, 44, 394–402. [Google Scholar] [CrossRef] [Green Version]
- Torii, Y.; Tokumaru, Y.; Kawaguchi, S.; Izumi, N.; Maruyama, S.; Mukamoto, M.; Kozaki, S.; Takahashi, M. Production and immunogenic efficacy of botulinum tetravalent (A., B., E., F) toxoid. Vaccine 2002, 20, 2556–2561. [Google Scholar] [CrossRef]
- Nowakowski, A.; Wang, C.; Powers, D.B.; Amersdorfer, P.; Smith, T.J.; Montgomery, V.A.; Sheridan, R.; Blake, R.; Smith, L.A.; Marks, J.D. Potent neutralization of botulinum neurotoxin by recombinant oligoclonal antibody. Proc. Natl. Acad. Sci. USA 2002, 99, 11346–11350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adekar, S.P.; Takahashi, T.; Jones, R.M.; Al-Saleem, F.H.; Ancharski, D.M.; Root, M.J.; Kapadnis, B.P.; Simpson, L.L.; Dessain, S.K. Neutralization of Botulinum Neurotoxin by a Human Monoclonal Antibody Specific for the Catalytic Light Chain. PLoS ONE 2008, 3, e3023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diamant, E.; Lachmi, B.-E.; Keren, A.; Barnea, A.; Marcus, H.; Cohen, S.; Ben David, A.; Zichel, R. Evaluating the Synergistic Neutralizing Effect of Anti-Botulinum Oligoclonal Antibody Preparations. PLoS ONE 2014, 9, e87089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujinaga, Y.; Sugawara, Y.; Matsumura, T. Uptake of botulinum neurotoxin in the intestine. Curr. Top. Microbiol. Immunol. 2013, 364, 45–59. [Google Scholar]
- Kozaki, S.; Kamata, Y.; Nishiki, T.-I.; Kakinuma, H.; Maruyama, H.; Takahashi, H.; Karasawa, T.; Yamakawa, K.; Nakamura, S. Characterization of Clostridium botulinum Type B Neurotoxin Associated with Infant Botulism in Japan. Infect. Immun. 1998, 66, 4811–4816. [Google Scholar] [CrossRef] [Green Version]
- Umeda, K.; Seto, Y.; Kohda, T.; Mukamoto, M.; Kozaki, S. Genetic Characterization of Clostridium botulinum Associated with Type B Infant Botulism in Japan. J. Clin. Microbiol. 2009, 47, 2720–2728. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.-H.; Kim, K.-S.; Kim, H.-W.; Jeong, S.T.; Huh, G.H.; Kim, J.C.; Jung, H.H. Isolation and characterization of a neutralizing antibody specific to internalization domain of Clostridium botulinum neurotoxin type B. Toxicon 2004, 44, 19–25. [Google Scholar] [CrossRef]
- Cheng, L.W.; Henderson, T.D.; Lam, T.I.; Stanker, L.H. Use of Monoclonal Antibodies in the Sensitive Detection and Neutralization of Botulinum Neurotoxin Serotype B. Toxins 2015, 7, 5068–5078. [Google Scholar] [CrossRef]
- Rasetti-Escargueil, C.; Avril, A.; Chahboun, S.; Tierney, R.; Bak, N.; Miethe, S.; Mazuet, C.; Popoff, M.R.; Thullier, P.; Hust, M.; et al. Development of human-like scFv-Fc antibodies neutralizing Botulinum toxin serotype B. mAbs 2015, 7, 1161–1177. [Google Scholar] [CrossRef]
- Miethe, S.; Mazuet, C.; Liu, Y.; Tierney, R.; Rasetti-Escargueil, C.; Avril, A.; Frenzel, A.; Thullier, P.; Pelat, T.; Urbain, R.; et al. Development of Germline-Humanized Antibodies Neutralizing Botulinum Neurotoxin A and B. PLoS ONE 2016, 11, e0161446. [Google Scholar] [CrossRef]
- Garcia-Rodriguez, C.; Geren, I.N.; Lou, J.; Conrad, F.; Forsyth, C.; Wen, W.; Chakraborti, S.; Zao, H.; Manzanarez, G.; Smith, T.J.; et al. Response re: ’Neutralizing human monoclonal antibodies binding multiple serotypes of botulinum neurotoxin’. Protein Eng. Des. Sel. 2011, 24, 633–634. [Google Scholar] [CrossRef]
- Adekar, S.P.; Jones, R.M.; Elias, M.; Al-Saleem, F.H.; Root, M.J.; Simpson, L.L.; Dessain, S. Hybridoma populations enriched for affinity-matured human IgGs yield high-affinity antibodies specific for botulinum neurotoxins. J. Immunol. Methods 2008, 333, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Garcia-Rodriguez, C.; Manzanarez, G.; Silberg, M.; Conrad, F.; Bettencourt, J.; Pan, X.; Breece, T.; To, R.; Li, M.; et al. Engineered domain-based assays to identify individual antibodies in oligoclonal combinations targeting the same protein. Anal. Biochem. 2012, 430, 141–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakaguchi, G. Clostridium botulinum toxins. Pharmacol. Ther. 1982, 19, 165–194. [Google Scholar] [CrossRef]
- Adekar, S.P.; Segan, A.T.; Chen, C.; Bermudez, R.; Elias, M.D.; Selling, B.H.; Kapadnis, B.P.; Simpson, L.L.; Simon, P.M.; Dessain, S.K. Enhanced Neutralization Potency of Botulinum Neurotoxin Antibodies Using a Red Blood Cell-Targeting Fusion Protein. PLoS ONE 2011, 6, 17491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, R.; Zhao, H.; Al-Saleem, F.H.; Ubaid, A.S.; Puligedda, R.D.; Segan, A.T.; Lindorfer, M.A.; Bermudez, R.; Elias, M.; Adekar, S.P.; et al. Mechanisms of enhanced neutralization of botulinum neurotoxin by monoclonal antibodies conjugated to antibodies specific for the erythrocyte complement receptor. Mol. Immunol. 2013, 57, 247–254. [Google Scholar] [CrossRef]
- Suzuki, T.; Ishii-Watabe, A.; Tada, M.; Kobayashi, T.; Kanayasu-Toyoda, T.; Kawanishi, T.; Yamaguchi, T. Importance of Neonatal FcR in Regulating the Serum Half-Life of Therapeutic Proteins Containing the Fc Domain of Human IgG1: A Comparative Study of the Affinity of Monoclonal Antibodies and Fc-Fusion Proteins to Human Neonatal FcR. J. Immunol. 2010, 184, 1968–1976. [Google Scholar] [CrossRef]
- Hill, K.K.; Smith, T.J.; Helma, C.H.; Ticknor, L.; Foley, B.; Svensson, R.T.; Brown, J.L.; Johnson, E.A.; Smith, L.A.; Okinaka, R.T.; et al. Genetic Diversity among Botulinum Neurotoxin-Producing Clostridial Strains. J. Bacteriol. 2006, 189, 818–832. [Google Scholar] [CrossRef] [Green Version]
- Kalb, S.R.; Baudys, J.; Rees, J.C.; Smith, T.J.; Smith, L.A.; Helma, C.H.; Hill, K.; Kull, S.; Kirchner, S.; Dorner, M.B.; et al. De novo subtype and strain identification of botulinum neurotoxin type B through toxin proteomics. Anal. Bioanal. Chem. 2012, 403, 215–226. [Google Scholar] [CrossRef] [Green Version]
- Peck, M.; Smith, T.J.; Anniballi, F.; Austin, J.W.; Bano, L.; Bradshaw, M.; Cuervo, P.; Cheng, L.W.; Derman, Y.; Dorner, B.G.; et al. Historical Perspectives and Guidelines for Botulinum Neurotoxin Subtype Nomenclature. Toxins 2017, 9, 38. [Google Scholar] [CrossRef]
- Beers, W.H.; Reich, E. Isolation and characterization of Clostridium botulinum type B toxin. J. Boil. Chem. 1969, 244, 4473–4479. [Google Scholar]
- Kohda, T.; Hosomi, K.; Kozaki, S.; Nakamura, K.; Torii, Y.; Mukamoto, M. Characterization of the functional activity of botulinum neurotoxin subtype B6. Microbiol. Immunol. 2017, 61, 482–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohda, T.; Ihara, H.; Seto, Y.; Tsutsuki, H.; Mukamoto, M.; Kozaki, S. Differential contribution of the residues in C-terminal half of the heavy chain of botulinum neurotoxin type B to its binding to the ganglioside GT1b and the synaptotagmin 2/GT1b complex. Microb. Pathog. 2007, 42, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Arimitsu, H.; Inoue, K.; Sakaguchi, Y.; Lee, J.; Fujinaga, Y.; Watanabe, T.; Ohyama, T.; Hirst, R.; Oguma, K. Purification of Fully Activated Clostridium botulinum Serotype B Toxin for Treatment of Patients with Dystonia. Infect. Immun. 2003, 71, 1599–1603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reed, L.; Muench, H. A simple method of estimating fifty per cent endpoints12. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Kondo, H.; Shimizu, T.; Kubonoya, M.; Izumi, N.; Takahashi, M.; Sakaguchi, G. Titration of botulinum toxins for lethal toxicity by intravenous injection into mice. Jpn. J. Med Sci. Boil. 1984, 37, 131–135. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Volunteer Number | Number of Vaccinations | Days after Last Immunization | ELISA Titer (log2) BoNT/A1 | ELISA Titer (log2) BoNT/B1 |
|---|---|---|---|---|
| Bkf002 | 5 | 9 | 14 | 13 |
| Bkf002 | 5 | 18 | 13 | 15 |
| Bkf003 | 4 | 9 | 13 | 12 |
| Bkf003 | 4 | 18 | 13 | 14 |
| Volunteer Number | Days after Last Immunization | Clone No. | Isotype |
|---|---|---|---|
| Bkf002 | 9 | M1 | IgG * |
| 9 | M2 | IgG1 | |
| 9 | M3 | IgM | |
| 9 | M4 | IgG1 | |
| 9 | M5 | IgM | |
| 9 | M6 | IgM | |
| 18 | M7 | IgA | |
| Bkf003 | 9 | S1 | IgG1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsumura, T.; Amatsu, S.; Misaki, R.; Yutani, M.; Du, A.; Kohda, T.; Fujiyama, K.; Ikuta, K.; Fujinaga, Y. Fully Human Monoclonal Antibodies Effectively Neutralizing Botulinum Neurotoxin Serotype B. Toxins 2020, 12, 302. https://doi.org/10.3390/toxins12050302
Matsumura T, Amatsu S, Misaki R, Yutani M, Du A, Kohda T, Fujiyama K, Ikuta K, Fujinaga Y. Fully Human Monoclonal Antibodies Effectively Neutralizing Botulinum Neurotoxin Serotype B. Toxins. 2020; 12(5):302. https://doi.org/10.3390/toxins12050302
Chicago/Turabian StyleMatsumura, Takuhiro, Sho Amatsu, Ryo Misaki, Masahiro Yutani, Anariwa Du, Tomoko Kohda, Kazuhito Fujiyama, Kazuyoshi Ikuta, and Yukako Fujinaga. 2020. "Fully Human Monoclonal Antibodies Effectively Neutralizing Botulinum Neurotoxin Serotype B" Toxins 12, no. 5: 302. https://doi.org/10.3390/toxins12050302
APA StyleMatsumura, T., Amatsu, S., Misaki, R., Yutani, M., Du, A., Kohda, T., Fujiyama, K., Ikuta, K., & Fujinaga, Y. (2020). Fully Human Monoclonal Antibodies Effectively Neutralizing Botulinum Neurotoxin Serotype B. Toxins, 12(5), 302. https://doi.org/10.3390/toxins12050302