Assessment of Ciguatera and Other Phycotoxin-Related Risks in Anaho Bay (Nuku Hiva Island, French Polynesia): Molecular, Toxicological, and Chemical Analyses of Passive Samplers

 ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
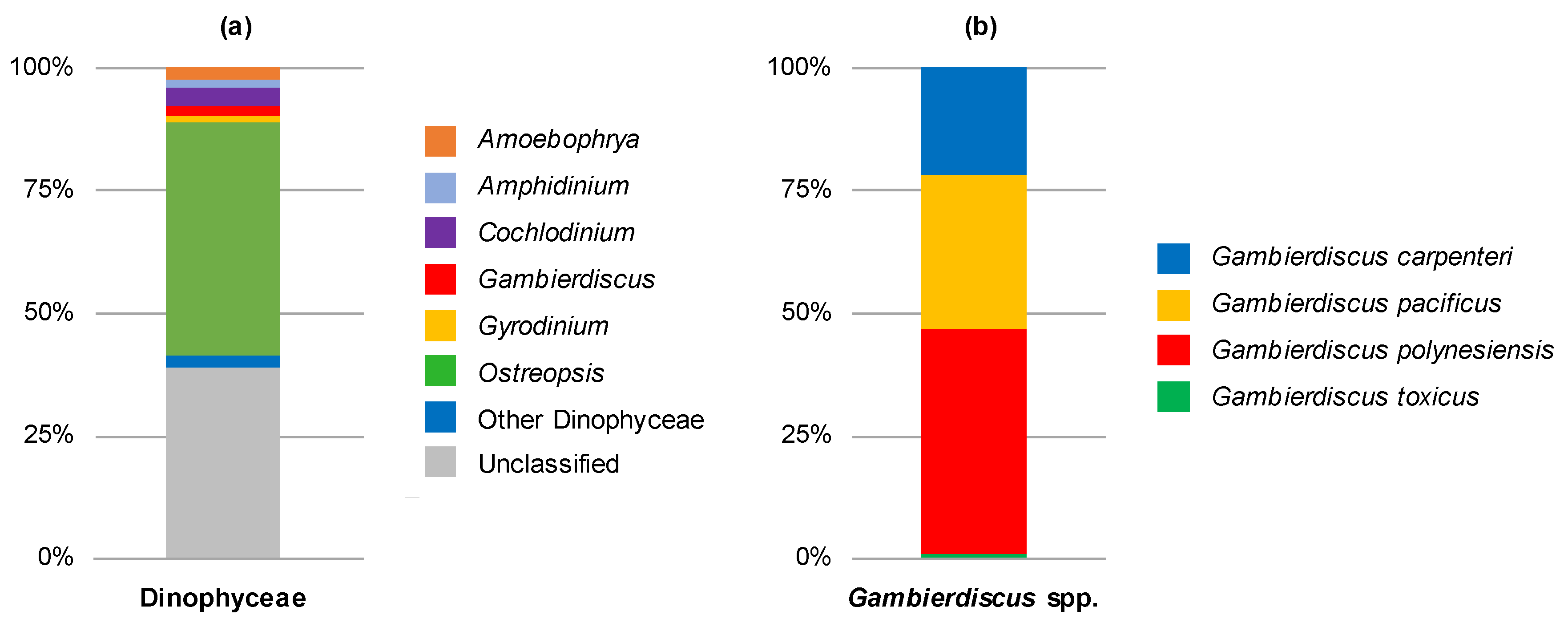
2.1. Molecular Analyses of Environmental Samples
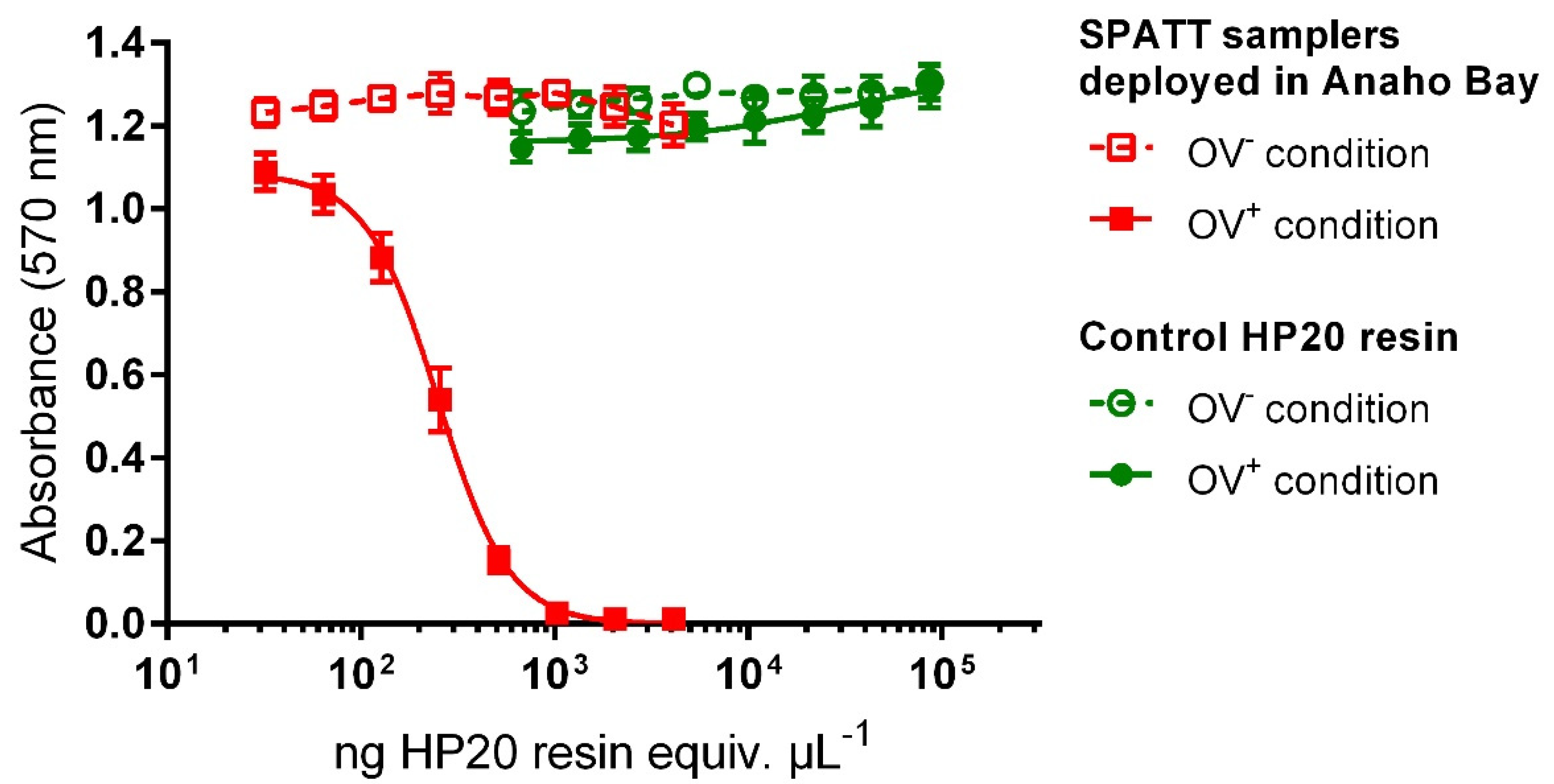
2.2. Evaluation of SPATT Samples Toxicity Using CBA-N2a
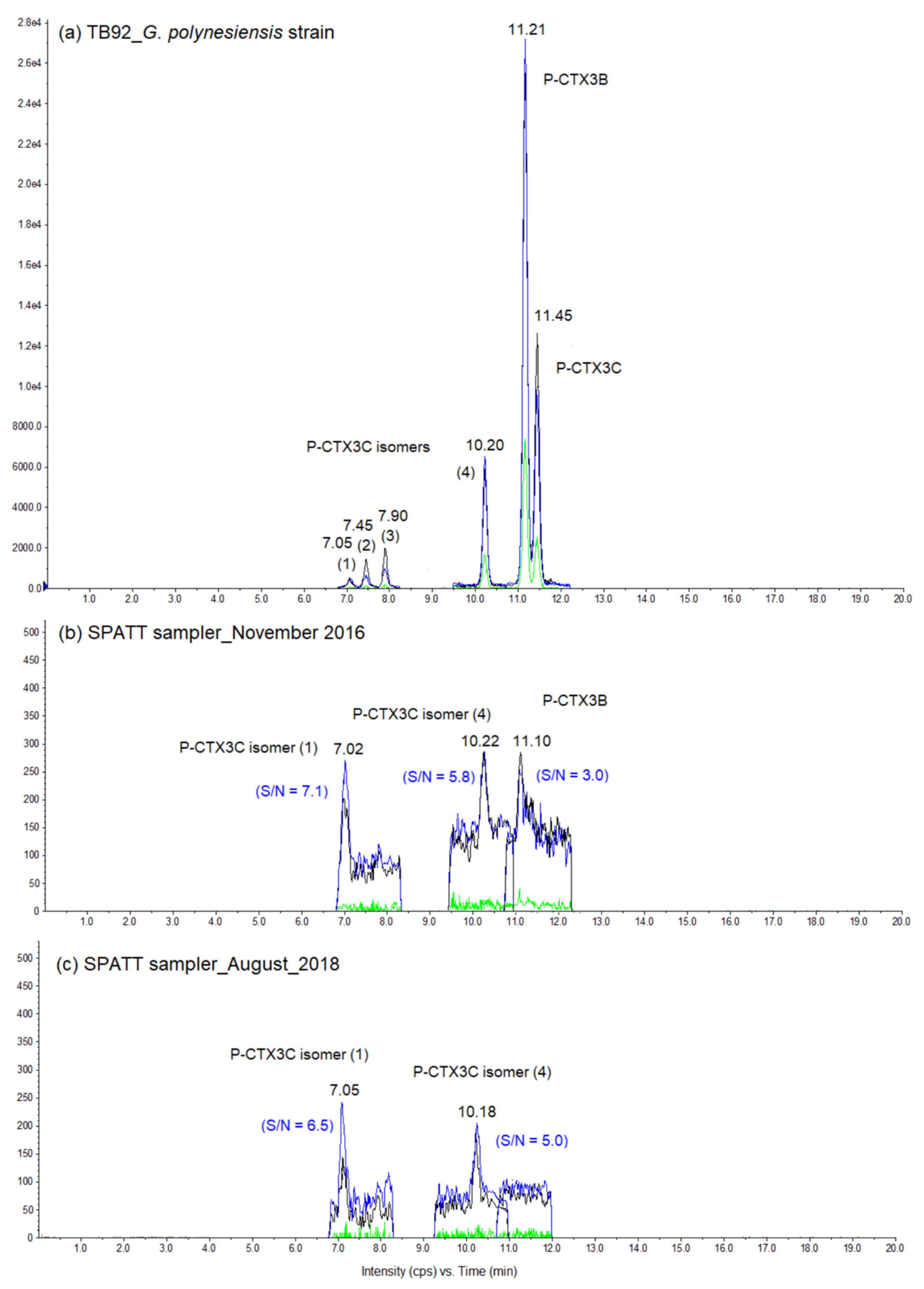
2.3. Characterization of Toxins Adsorbed by HP20 Resin Using LC-MS/MS
3. Discussion
3.1. Benthic Harmful Algae Populations and Toxin Profiles over Time in Anaho Bay
3.2. Effects of Deployment Time on the Efficacy of CTX Detection
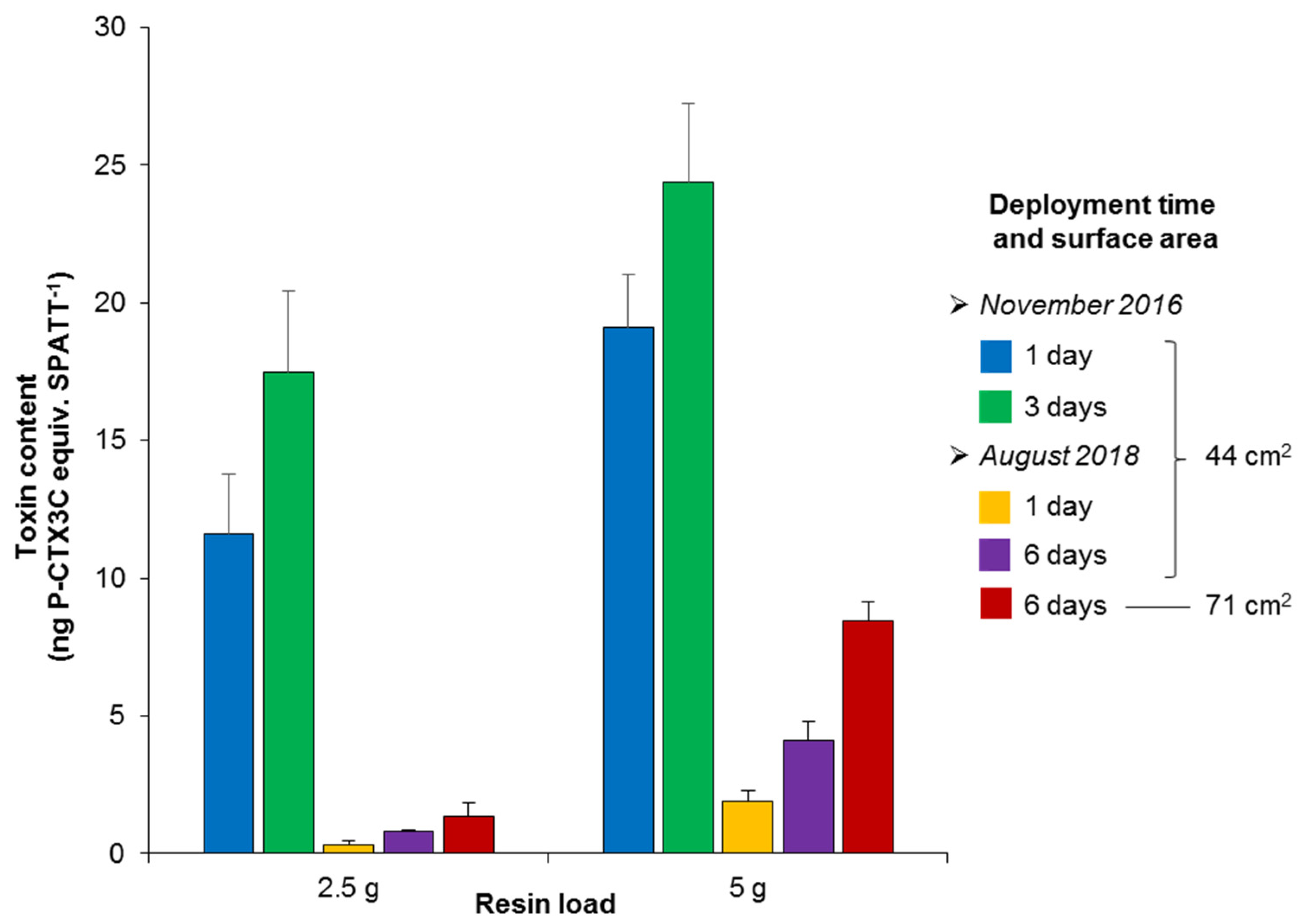
3.3. Effects of Resin Load on the Efficacy of CTX Detection
4. Conclusions
5. Materials and Methods
5.1. Environmental Analyses
5.2. HP20 Resin and SPATT Device Preparation
5.3. Field Deployments of SPATT Devices
5.4. Toxin Extraction from HP20 Resin
5.5. Neuroblastoma Cell-Based Assay
5.6. Liquid Chromatography Coupled with Tandem Mass Spectrometry Analyses
5.6.1. Detection Method for P-CTXs
5.6.2. Detection Method for OA and DTXs
5.6.3. Detection Methods for Other Toxins
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kudela, R.M.; Berdalet, E.; Bernard, S.; Burford, M.; Fernand, L.; Lu, S.; Roy, S.; Tester, P.A.; Usup, G.; Magnien, R.; et al. Harmful Algal Blooms: A Scientific Summary for Policy Makers; International Society for the Study of Harmful Algae (ISSHA) and Intergovernmental Oceanographic Commission (IOC) of UNESCO: Paris, France, 2015. [Google Scholar]
- Doucette, G.J.; Kudela, R.M. In situ and real-time identification of toxins and toxin-producing microorganisms in the environment. Compr. Anal. Chem. 2017, 78, 411–443. [Google Scholar] [CrossRef]
- Abraham, A.; Rambla-Alegre, M. Marine toxins analysis for consumer protection. Compr. Anal. Chem. 2017, 78, 343–378. [Google Scholar] [CrossRef]
- MacKenzie, L.; Beuzenberg, V.; Holland, P.; McNabb, P.; Selwood, A. Solid phase adsorption toxin tracking (SPATT): A new monitoring tool that simulates the biotoxin contamination of filter feeding bivalves. Toxicon 2004, 44, 901–918. [Google Scholar] [CrossRef] [PubMed]
- Kudela, R.M. Passive sampling for freshwater and marine algal toxins. Compr. Anal. Chem. 2017, 78, 379–409. [Google Scholar] [CrossRef]
- Roué, M.; Darius, H.; Chinain, M. Solid phase adsorption toxin tracking (SPATT) technology for the monitoring of aquatic toxins: A review. Toxins 2018, 10, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soliño, L.; Costa, P.R. Global impact of ciguatoxins and ciguatera fish poisoning on fish, fisheries and consumers. Environ. Res. 2020, 182, 109111. [Google Scholar] [CrossRef]
- Chinain, M.; Gatti, C.M.; Roué, M.; Darius, H.T. Ciguatera-causing dinoflagellates in the genera Gambierdiscus and Fukuyoa: Distribution, ecophysiology and toxicology. In Dinoflagellates: Morphology, Life History and Ecological Significance; Durvasula, S.R., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2020; in press. [Google Scholar]
- Parsons, M.L.; Aligizaki, K.; Dechraoui Bottein, M.Y.; Fraga, S.; Morton, S.L.; Penna, A.; Rhodes, L. Gambierdiscus and Ostreopsis: Reassessment of the state of knowledge of their taxonomy, geography, ecophysiology, and toxicology. Harmful Algae 2012, 14, 107–129. [Google Scholar] [CrossRef]
- Pérez-Arellano, J.-L.; Luzardo, O.P.; Pérez Brito, A.; Hernández Cabrera, M.; Zumbado, M.; Carranza, C.; Angel-Moreno, A.; Dickey, R.W.; Boada, L.D. Ciguatera fish poisoning, Canary Islands. Emerg. Infect. Dis. 2005, 11, 1981–1982. [Google Scholar] [CrossRef] [Green Version]
- Otero, P.; Pérez, S.; Alfonso, A.; Vale, C.; Rodríguez, P.; Gouveia, N.N.; Gouveia, N.; Delgado, J.; Vale, P.; Hirama, M.; et al. First toxin profile of ciguateric fish in Madeira arquipelago (Europe). Anal. Chem. 2010, 82, 6032–6039. [Google Scholar] [CrossRef]
- Roué, M.; Darius, H.T.; Viallon, J.; Ung, A.; Gatti, C.M.; Harwood, D.T.; Chinain, M. Application of solid phase adsorption toxin tracking (SPATT) devices for the field detection of Gambierdiscus toxins. Harmful Algae 2018, 71, 40–49. [Google Scholar] [CrossRef]
- Darius, H.T.; Ponton, D.; Revel, T.; Cruchet, P.; Ung, A.; Tchou Fouc, M.; Chinain, M. Ciguatera risk assessment in two toxic sites of French Polynesia using the receptor-binding assay. Toxicon 2007, 50, 612–626. [Google Scholar] [CrossRef] [PubMed]
- Darius, H.T.; Roué, M.; Sibat, M.; Viallon, J.; Gatti, C.M.; Vandersea, M.W.; Tester, P.A.; Litaker, R.W.; Amzil, Z.; Hess, P.; et al. Tectus niloticus (Tegulidae, Gastropod) as a novel vector of ciguatera poisoning: Detection of Pacific ciguatoxins in toxic samples from Nuku Hiva Island (French Polynesia). Toxins 2018, 10, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darius, H.T.; Roué, M.; Sibat, M.; Viallon, J.; Gatti, C.M.; Vandersea, M.W.; Tester, P.A.; Litaker, R.W.; Amzil, Z.; Hess, P.; et al. Toxicological investigations on the sea urchin Tripneustes gratilla (Toxopneustidae, Echinoid) from Anaho Bay (Nuku Hiva, French Polynesia): Evidence for the presence of Pacific ciguatoxins. Mar. Drugs 2018, 16, 122. [Google Scholar] [CrossRef] [Green Version]
- Chinain, M.; Darius, H.T.; Ung, A.; Cruchet, P.; Wang, Z.; Ponton, D.; Laurent, D.; Pauillac, S. Growth and toxin production in the ciguatera-causing dinoflagellate Gambierdiscus polynesiensis (Dinophyceae) in culture. Toxicon 2010, 56, 739–750. [Google Scholar] [CrossRef]
- Longo, S.; Sibat, M.; Viallon, J.; Darius, H.T.; Hess, P.; Chinain, M. Intraspecific variability in the toxin production and toxin profiles of in vitro cultures of Gambierdiscus polynesiensis (Dinophyceae) from French Polynesia. Toxins 2019, 11, 735. [Google Scholar] [CrossRef] [Green Version]
- Gatti, C.M.; Lonati, D.; Darius, H.T.; Zancan, A.; Roué, M.; Schicchi, A.; Locatelli, C.A.; Chinain, M. Tectus niloticus (Tegulidae, Gastropod) as a novel vector of ciguatera poisoning: Clinical characterization and follow-up of a mass poisoning event in Nuku Hiva Island (French Polynesia). Toxins 2018, 10, 102. [Google Scholar] [CrossRef] [Green Version]
- MacKenzie, L.A. In situ passive solid-phase adsorption of micro-algal biotoxins as a monitoring tool. Curr. Opin. Biotechnol. 2010, 21, 326–331. [Google Scholar] [CrossRef]
- Skinner, M.P.; Brewer, T.D.; Johnstone, R.; Fleming, L.E.; Lewis, R.J. Ciguatera fish poisoning in the Pacific Islands (1998 to 2008). PLOS Negl. Trop. D 2011, 5, e1416. [Google Scholar] [CrossRef]
- Lassus, P.; Chomérat, N.; Hess, P.; Nézan, E. Toxic and Harmful Microalgae of the World Ocean/Micro-Algues Toxiques et Nuisibles de L’océan Mondial; IOC Manuals and Guides, 68; International Society for the Study of Harmful Algae (ISSHA) and Intergovernmental Oceanographic Commission (IOC) of UNESCO: Copenhagen, Denmark, 2016. [Google Scholar]
- Zendong, Z.; Bertrand, S.; Herrenknecht, C.; Abadie, E.; Jauzein, C.; Lemée, R.; Gouriou, J.; Amzil, Z.; Hess, P. Passive sampling and high resolution mass spectrometry for chemical profiling of French coastal areas with a focus on marine biotoxins. Environ. Sci. Technol. 2016, 50, 8522–8529. [Google Scholar] [CrossRef]
- Chomérat, N.; Bilien, G.; Derrien, A.; Henry, K.; Ung, A.; Viallon, J.; Darius, H.T.; Mahana iti Gatti, C.; Roué, M.; Hervé, F.; et al. Ostreopsis lenticularis Y. Fukuyo (Dinophyceae, Gonyaulacales) from French Polynesia (South Pacific Ocean): A revisit of its morphology, molecular phylogeny and toxicity. Harmful Algae 2019, 84, 95–111. [Google Scholar] [CrossRef] [Green Version]
- Accoroni, S.; Totti, C. The toxic benthic dinoflagellates of the genus Ostreopsis in temperate areas: A review. Adv. Oceanogr. Limnol. 2016, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Fux, E.; Bire, R.; Hess, P. Comparative accumulation and composition of lipophilic marine biotoxins in passive samplers and in mussels (M. edulis) on the West Coast of Ireland. Harmful Algae 2009, 8, 523–537. [Google Scholar] [CrossRef]
- Pizarro, G.; Moroño, Á.; Paz, B.; Franco, M.J.; Pazos, Y.; Reguera, B. Evaluation of passive samplers as a monitoring tool for early warning of Dinophysis toxins in shellfish. Mar. Drugs 2013, 11, 3823–3845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Altares, M.; Casanova, A.; Bane, V.; Diogène, J.; Furey, A.; de la Iglesia, P. Confirmation of pinnatoxins and spirolides in shellfish and passive samplers from Catalonia (Spain) by liquid chromatography coupled with triple quadrupole and high-resolution hybrid tandem mass spectrometry. Mar. Drugs 2014, 12, 3706–3732. [Google Scholar] [CrossRef]
- Gibble, C.M.; Kudela, R.M. Detection of persistent microcystin toxins at the land-sea interface in Monterey Bay, California. Harmful Algae 2014, 39, 146–153. [Google Scholar] [CrossRef] [Green Version]
- Zendong, Z.; Kadiri, M.; Herrenknecht, C.; Nézan, E.; Mazzeo, A.; Hess, P. Algal toxin profiles in Nigerian coastal waters (Gulf of Guinea) using passive sampling and liquid chromatography coupled to mass spectrometry. Toxicon 2016, 114, 16–27. [Google Scholar] [CrossRef] [Green Version]
- Hattenrath-Lehmann, T.K.; Lusty, M.W.; Wallace, R.B.; Haynes, B.; Wang, Z.; Broadwater, M.; Deeds, J.R.; Morton, S.L.; Hastback, W.; Porter, L.; et al. Evaluation of rapid, early warning approaches to track shellfish toxins associated with Dinophysis and Alexandrium blooms. Mar. Drugs 2018, 16, 28. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, M.; Bane, V.; García-Altares, M.; van Pelt, F.N.A.M.; Furey, A.; O’Halloran, J. Assessment of emerging biotoxins (pinnatoxin G and spirolides) at Europe’s first marine reserve: Lough Hyne. Toxicon 2015, 108, 202–209. [Google Scholar] [CrossRef]
- Li, F.-L.; Li, Z.-X.; Guo, M.-M.; Wu, H.-Y.; Zhang, T.-T.; Song, C.-H. Investigation of diarrhetic shellfish toxins in Lingshan Bay, Yellow Sea, China, using solid-phase adsorption toxin tracking (SPATT). Food Addit. Contam. A 2016, 33, 1367–1373. [Google Scholar] [CrossRef]
- Zendong, Z.; Herrenknecht, C.; Abadie, E.; Brissard, C.; Tixier, C.; Mondeguer, F.; Séchet, V.; Amzil, Z.; Hess, P. Extended evaluation of polymeric and lipophilic sorbents for passive sampling of marine toxins. Toxicon 2014, 91, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Wood, S.A.; Holland, P.T.; MacKenzie, L. Development of solid phase adsorption toxin tracking (SPATT) for monitoring anatoxin-a and homoanatoxin-a in river water. Chemosphere 2011, 82, 888–894. [Google Scholar] [CrossRef] [PubMed]
- Fux, E.; Marcaillou, C.; Mondeguer, F.; Bire, R.; Hess, P. Field and mesocosm trials on passive sampling for the study of adsorption and desorption behaviour of lipophilic toxins with a focus on OA and DTX1. Harmful Algae 2008, 7, 574–583. [Google Scholar] [CrossRef] [Green Version]
- Lane, J.Q.; Roddam, C.M.; Langlois, G.W.; Kudela, R.M. Application of solid phase adsorption toxin tracking (SPATT) for field detection of the hydrophilic phycotoxins domoic acid and saxitoxin in coastal California. Limnol. Oceanogr. Meth. 2010, 8, 645–660. [Google Scholar] [CrossRef]
- Rodríguez, P.; Alfonso, A.; Turrell, E.; Lacaze, J.-P.; Botana, L.M. Study of solid phase adsorption of paralytic shellfish poisoning toxins (PSP) onto different resins. Harmful Algae 2011, 10, 447–455. [Google Scholar] [CrossRef]
- Vandersea, M.W.; Kibler, S.R.; Holland, W.C.; Tester, P.A.; Schultz, T.F.; Faust, M.A.; Holmes, M.J.; Chinain, M.; Wayne Litaker, R. Development of semi-quantitative PCR assays for the detection and enumeration of Gambierdiscus species (Gonyaulacales, Dinophyceae). J. Phycol. 2012, 48, 902–915. [Google Scholar] [CrossRef]
- Smith, K.F.; Biessy, L.; Argyle, P.A.; Trnski, T.; Halafihi, T.; Rhodes, L.L. Molecular identification of Gambierdiscus and Fukuyoa (Dinophyceae) from environmental samples. Mar. Drugs 2017, 15, 243. [Google Scholar] [CrossRef] [Green Version]
- Cox, M.P.; Peterson, D.A.; Biggs, P.J. SolexaQA: At-a-glance quality assessment of Illumina second-generation sequencing data. BMC Bioinform. 2010, 11, 485. [Google Scholar] [CrossRef] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.F.; Kohli, G.S.; Murray, S.A.; Rhodes, L.L. Assessment of the metabarcoding approach for community analysis of benthic-epiphytic dinoflagellates using mock communities. N. Z. J. Mar. Freshw. Res. 2017, 51, 555–576. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Sibat, M.; Herrenknecht, C.; Darius, H.T.; Roué, M.; Chinain, M.; Hess, P. Detection of Pacific ciguatoxins using liquid chromatography coupled to either low or high resolution mass spectrometry (LC-MS/MS). J. Chromatogr. A 2018, 1571, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Vial, J.; Jardy, A. Experimental comparison of the different approaches to estimate LOD and LOQ of an HPLC method. Anal. Chem. 1999, 71, 2672–2677. [Google Scholar] [CrossRef]
- Sanagi, M.M.; Ling, S.L.; Nasir, Z.; Hermawan, D.; Ibrahim, W.A.; Abu Naim, A. Comparison of signal-to-noise, blank determination, and linear regression methods for the estimation of detection and quantification limits for volatile organic compounds by gas chromatography. J. AOAC Int. 2009, 92, 1833–1838. [Google Scholar] [CrossRef] [Green Version]
- Zendong, Z.; Sibat, M.; Herrenknecht, C.; Hess, P.; McCarron, P. Relative molar response of lipophilic marine algal toxins in liquid chromatography/electrospray ionization mass spectrometry. Rapid Commun. Mass Spectrom. 2017, 31, 1453–1461. [Google Scholar] [CrossRef]
- Brissard, C.; Hervé, F.; Sibat, M.; Séchet, V.; Hess, P.; Amzil, Z.; Herrenknecht, C. Characterization of ovatoxin-h, a new ovatoxin analog, and evaluation of chromatographic columns for ovatoxin analysis and purification. J. Chromatogr. A 2015, 1388, 87–101. [Google Scholar] [CrossRef] [Green Version]
- Pisapia, F.; Sibat, M.; Herrenknecht, C.; Lhaute, K.; Gaiani, G.; Ferron, P.-J.; Fessard, V.; Fraga, S.; Nascimento, S.M.; Litaker, R.W.; et al. Maitotoxin-4, a novel MTX analog produced by Gambierdiscus excentricus. Mar. Drugs 2017, 15, 220. [Google Scholar] [CrossRef]
- Bormans, M.; Amzil, Z.; Mineaud, E.; Brient, L.; Savar, V.; Robert, E.; Lance, E. Demonstrated transfer of cyanobacteria and cyanotoxins along a freshwater-marine continuum in France. Harmful Algae 2019, 87, 101639. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
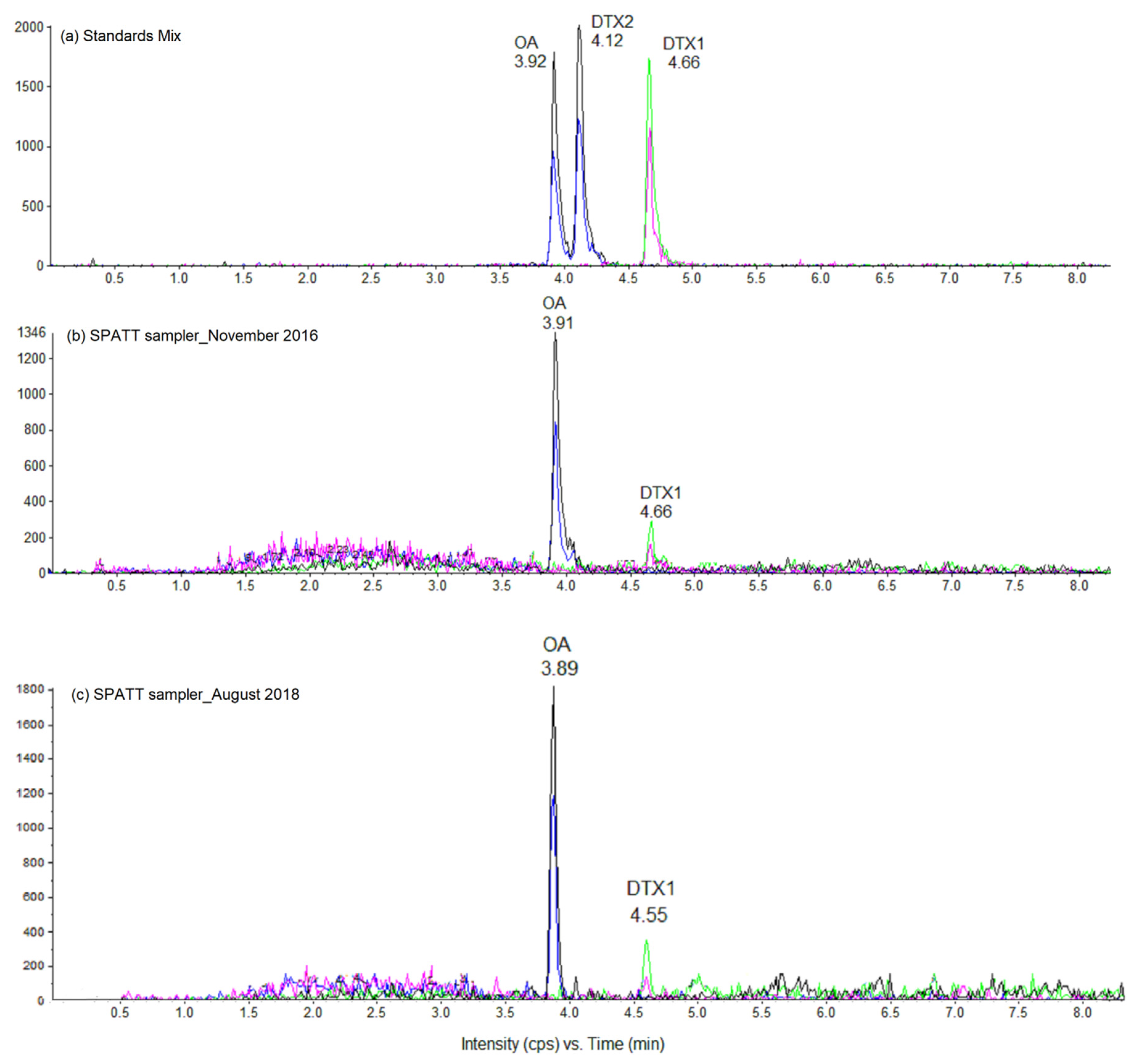{kind=link}
| Field Experiment | Dinoflagellate Genera Observed on Macroalgal Substrates | qPCR Analyses of Artificial WS Substrates | |
|---|---|---|---|
| Cell Density of Gambierdiscus spp. 2 | Gambierdiscus Species Identification | ||
| November 2016 1 | Gambierdiscus Ostreopsis Prorocentrum | ~2900 | G. polynesiensis (82%) G. carpenteri (17%) G. caribaeus (<1%) G. pacificus (<1%) G. toxicus (<1%) |
| August 2018 | Ostreopsis Gambierdiscus | ND | ND |
| Field Experiment | Deployment Time (days) | Resin Load (g) | Surface of Exposure (cm2) | EC50 1 (ng HP20 Resin Equiv. µL−1) | Toxin Content (ng P-CTX3C Equiv. g−1 HP20 Resin) |
|---|---|---|---|---|---|
| November 2016 | 1 | 2.5 | 44 | 410 ± 78 | 4.6 ± 0.9 |
| 5 | 486 ± 52 | 3.8 ± 0.4 | |||
| 3 | 2.5 | 270 ± 51 | 7.0 ± 1.2 | ||
| 5 | 381 ± 44 | 4.9 ± 0.6 | |||
| August 2018 | 1 | 2.5 | 44 | 14,611 ± 4817 | 0.14 ± 0.04 |
| 5 | 4937 ± 966 | 0.38 ± 0.08 | |||
| 6 | 2.5 | 44 | 4589 ± 149 | 0.40 ± 0.01 | |
| 71 | 3661 ± 1391 | 0.54 ± 0.21 | |||
| 5 | 44 | 2290 ± 410 | 0.82 ± 0.14 | ||
| 71 | 1095 ± 95 | 1.7 ± 0.2 |
| Field Experiment | Deployment Time (Days) | Resin Load (g) | Surface of Exposure (cm2) | Toxin Content 1 (ng Toxin SPATT−1) | |
|---|---|---|---|---|---|
| OA | DTX1 | ||||
| November 2016 | 1 | 2.5 | 44 | 9.0 ± 1.6 | ND 2 |
| 5 | 21.3 ± 4.3 | ND | |||
| 3 | 2.5 | 13.0 ± 1.3 | ND | ||
| 5 | 37.0 ± 13.0 | 4.2 ± 1.6 | |||
| August 2018 | 1 | 2.5 | 44 | 3.3 ± 0.1 | ND |
| 5 | 6.2 ± 0.8 | ND | |||
| 6 | 2.5 | 44 | 18.6 ± 0.2 | 2.1 ± 0.5 | |
| 71 | 25.4 ± 1.3 | 2.0 ± 0.1 | |||
| 5 | 44 | 25.4 ± 6.4 | 3.1 ± 0.8 | ||
| 71 3 | 47.0 | 7.0 | |||
| Compound | Pseudo Molecular Ion | MRM Transitions (m/z) | DP (V) | CE (eV) | LOD (ng mL−1) | LOQ (ng mL−1) |
|---|---|---|---|---|---|---|
| P-CTX1B | [M + NH4]+ | 1128.6/1093.6 | 105 | 20 | ||
| 1128.6/1075.6 1 | 105 | 30 | ||||
| 1128.6/95.1 | 105 | 90 | ||||
| P-CTX3C & P-CTX3B | [M + NH4]+ | 1040.5/1005.6 | 105 | 30 | ||
| [M + H]+ | 1023.6/1005.6 1 | 105 | 20 | 2 | 6 | |
| 1023.6/125.1 | 105 | 50 | ||||
| P-CTX4A & P-CTX4B | [M + NH4]+ | 1078.6/1043.6 | 105 | 30 | ||
| [M + H]+ | 1061.6/1043.6 1 | 105 | 20 | |||
| 1061.6/125.1 | 105 | 50 | ||||
| 2,3-diOH-P-CTX3C | [M + NH4]+ | 1074.6/1039.6 | 105 | 30 | ||
| [M + H]+ | 1057.6/1039.6 1 | 105 | 20 | |||
| 1057.6/125.1 | 105 | 50 | ||||
| 51-OH-P-CTX3C | [M + NH4]+ | 1056.6/1021.6 1 | 105 | 30 | ||
| [M + H]+ | 1039.6/1021.6 | 105 | 20 | |||
| 1039.6/1003.6 | 105 | 20 | ||||
| M-seco-P-CTX3C | [M + H]+ | 1041.6/1023.6 1 | 105 | 30 | ||
| 1041.6/1005.6 | 105 | 20 | ||||
| 1041.6/125.1 | 105 | 50 | ||||
| P-CTX2 & P-CTX3 | [M + NH4]+ | 1112.6/1077.6 | 105 | 20 | ||
| 1112.6/1059.6 1 | 105 | 30 | ||||
| 112.6/95.1 | 105 | 90 | ||||
| 2-OH-P-CTX3C & 3-OH-P-CTX3C | [M + NH4]+ | 1058.6/1023.6 1 | 105 | 30 | ||
| 1058.6/1005.6 | 105 | 20 | ||||
| 1058.6/125.1 | 105 | 50 | ||||
| OA | [M − H]− | 803.4/255.1 1 | −170 | −62 | 1 | 3 |
| 803.4/113.1 | −170 | −92 | ||||
| DTX2 | [M − H]− | 803.4/255.1 1 | −170 | −62 | 1 | 3 |
| 803.4/113.1 | −170 | −92 | ||||
| DTX1 | [M − H]− | 817.5/255.1 1 | −170 | −68 | 1 | 3 |
| 817.5/113.1 | −170 | −92 | ||||
| YTX | [M − H]− | 1141.4/1061.6 1 | −120 | −48 | 1 | 3 |
| 1141.4/855.6 | −120 | −98 | ||||
| Homo-YTX | [M − H]− | 1155.5/1075.6 1 | −120 | −48 | ||
| 1155.5/869.4 | −120 | −98 | ||||
| 45-OH YTX | [M − H]− | 1157.5/1077.5 1 | −120 | −48 | ||
| 1157.5/855.5 | −120 | −98 | ||||
| 45-OH homo YTX | [M − H]− | 1171.5/1091.5 1 | −120 | −48 | ||
| 1171.5/869.4 | −120 | −98 | ||||
| COOH YTX | [M − H]− | 1173.5/1093.5 1 | −120 | −48 | ||
| 1173.5/855.5 | −120 | −98 | ||||
| Homo COOH YTX | [M − H]− | 1187.5/1107.5 1 | −120 | −48 | ||
| 1187.5/869.4 | −120 | −98 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roué, M.; Smith, K.F.; Sibat, M.; Viallon, J.; Henry, K.; Ung, A.; Biessy, L.; Hess, P.; Darius, H.T.; Chinain, M. Assessment of Ciguatera and Other Phycotoxin-Related Risks in Anaho Bay (Nuku Hiva Island, French Polynesia): Molecular, Toxicological, and Chemical Analyses of Passive Samplers. Toxins 2020, 12, 321. https://doi.org/10.3390/toxins12050321
Roué M, Smith KF, Sibat M, Viallon J, Henry K, Ung A, Biessy L, Hess P, Darius HT, Chinain M. Assessment of Ciguatera and Other Phycotoxin-Related Risks in Anaho Bay (Nuku Hiva Island, French Polynesia): Molecular, Toxicological, and Chemical Analyses of Passive Samplers. Toxins. 2020; 12(5):321. https://doi.org/10.3390/toxins12050321
Chicago/Turabian StyleRoué, Mélanie, Kirsty F. Smith, Manoella Sibat, Jérôme Viallon, Kévin Henry, André Ung, Laura Biessy, Philipp Hess, Hélène Taiana Darius, and Mireille Chinain. 2020. "Assessment of Ciguatera and Other Phycotoxin-Related Risks in Anaho Bay (Nuku Hiva Island, French Polynesia): Molecular, Toxicological, and Chemical Analyses of Passive Samplers" Toxins 12, no. 5: 321. https://doi.org/10.3390/toxins12050321
APA StyleRoué, M., Smith, K. F., Sibat, M., Viallon, J., Henry, K., Ung, A., Biessy, L., Hess, P., Darius, H. T., & Chinain, M. (2020). Assessment of Ciguatera and Other Phycotoxin-Related Risks in Anaho Bay (Nuku Hiva Island, French Polynesia): Molecular, Toxicological, and Chemical Analyses of Passive Samplers. Toxins, 12(5), 321. https://doi.org/10.3390/toxins12050321