Targeting the Early Endosome-to-Golgi Transport of Shiga Toxins as a Therapeutic Strategy

Abstract
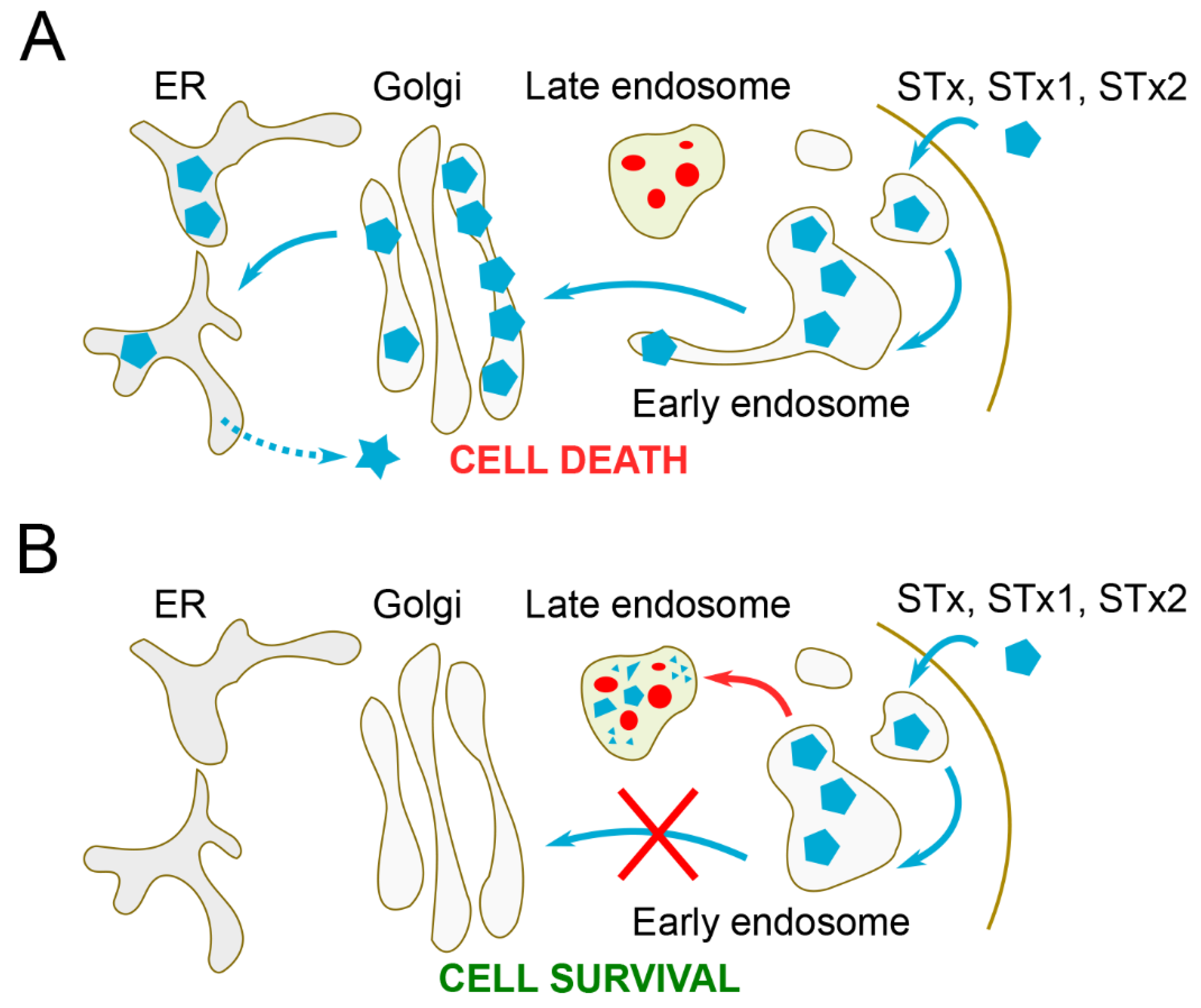
:1. Introduction
2. Early Endosome-to-Golgi Transport of STx and STx1
2.1. GPP130—The Endosomal Receptor for STxB/STx1B
2.2. Cholesterol and Membrane Microdomains
2.3. Clathrin and Retromer Coats and Accessory Proteins
2.4. GTPase Dynamin
2.5. Cytoskeletal Factors
2.6. Tethering, Docking, and Fusion with the Golgi
2.7. Other Regulators
3. Early Endosome-to-Golgi Transport of STx2
3.1. Similarities and Differences in the Transport of STx2B and STxB/STx1B
3.2. Late Endosome–Lysosome Fusion Modulates Early Endosome-to-Golgi Trafficking of STx2B—Insights Obtained from a Genome-Wide siRNA Screen
4. Small-Molecule Inhibitors Targeting Early Endosome-to-Golgi Transport of STx, STx1, and STx2
4.1. Compound 134
4.2. Retro-2 Substances
4.3. Manganese
4.4. Tamoxifen and Derivatives
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nataro, J.P.; Kaper, J.B. Diarrheagenic Escherichia coli. Clin. Microbiol. Rev. 1998, 11, 142–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahsay, A.G.; Muthupandian, S. A review on Sero diversity and antimicrobial resistance patterns of Shigella species in Africa, Asia and South America, 2001–2014. BMC Res. Notes 2016, 9, 422. [Google Scholar] [CrossRef] [Green Version]
- Mead, P.S.; Slutsker, L.; Dietz, V.; McCaig, L.F.; Bresee, J.S.; Shapiro, C.; Griffin, P.M.; Tauxe, R.V. Food-related illness and death in the United States. Emerg. Infect. Dis. 1999, 5, 607–625. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, A.D.; Holmes, R.K. Shiga and Shiga-like toxins. Microbiol. Rev. 1987, 51, 206–220. [Google Scholar] [CrossRef]
- Obrig, T.G.; Karpman, D. Shiga toxin pathogenesis: Kidney complications and renal failure. Curr. Top. Microbiol. Immunol. 2012, 357, 105–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsushiro, A.; Sato, K.; Miyamoto, H.; Yamamura, T.; Honda, T. Induction of Prophages of EnterohemorrhagicEscherichia coli O157:H7 with Norfloxacin. J. Bacteriol. 1999, 181, 2257–2260. [Google Scholar] [CrossRef] [Green Version]
- McGannon, C.M.; Fuller, C.A.; Weiss, A.A. Different Classes of Antibiotics Differentially Influence Shiga Toxin Production. Antimicrob. Agents Chemother. 2010, 54, 3790–3798. [Google Scholar] [CrossRef] [Green Version]
- Beddoe, T.; Paton, A.W.; Le Nours, J.; Rossjohn, J.; Paton, J.C. Structure, biological functions and applications of the AB5 toxins. Trends Biochem. Sci. 2010, 35, 411–418. [Google Scholar] [CrossRef] [Green Version]
- Mukhopadhyay, S.; Linstedt, A.D. Retrograde trafficking of AB5 toxins: Mechanisms to therapeutics. J. Mol. Med. 2013, 91, 1131–1141. [Google Scholar] [CrossRef] [Green Version]
- Mukhopadhyay, S.; Redler, B.; Linstedt, A. Shiga toxin–binding site for host cell receptor GPP130 reveals unexpected divergence in toxin-trafficking mechanisms. Mol. Boil. Cell 2013, 24, 2311–2318. [Google Scholar] [CrossRef]
- Strockbine, N.A.; Jackson, M.P.; Sung, L.M.; Holmes, R.K.; O’Brien, A.D. Cloning and sequencing of the genes for Shiga toxin from Shigella dysenteriae type 1. J. Bacteriol. 1988, 170, 1116–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tesh, V.L.; Burris, J.A.; Owens, J.W.; Gordon, V.M.; Wadolkowski, E.A.; O’Brien, A.D.; Samuel, J.E. Comparison of the relative toxicities of Shiga-like toxins type I and type II for mice. Infect. Immun. 1993, 61, 3392–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boerlin, P.; McEwen, S.A.; Boerlin-Petzold, F.; Wilson, J.B.; Johnson, R.P.; Gyles, C.L. Associations between Virulence Factors of Shiga Toxin-ProducingEscherichia coli and Disease in Humans. J. Clin. Microbiol. 1999, 37, 497–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selyunin, A.S.; Mukhopadhyay, S. A Conserved Structural Motif Mediates Retrograde Trafficking of Shiga Toxin Types 1 and 2. Traffic 2015, 16, 1270–1287. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Mukhopadhyay, S. Functional analyses of the UDP-galactose transporter SLC35A2 using the binding of bacterial Shiga toxins as a novel activity assay. Glycobiology 2019, 29, 490–503. [Google Scholar] [CrossRef]
- Lingwood, C. Role of verotoxin receptors in pathogenesis. Trends Microbiol. 1996, 4, 147–153. [Google Scholar] [CrossRef]
- Saenz, J.B.; Doggett, T.A.; Haslam, D.B. Identification and Characterization of Small Molecules That Inhibit Intracellular Toxin Transport. Infect. Immun. 2007, 75, 4552–4561. [Google Scholar] [CrossRef] [Green Version]
- Stechmann, B.; Bai, S.-K.; Gobbo, E.; Lopez, R.; Merer, G.; Pinchard, S.; Panigai, L.; Tenza, D.; Raposo, G.; Beaumelle, B.; et al. Inhibition of Retrograde Transport Protects Mice from Lethal Ricin Challenge. Cell 2010, 141, 231–242. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.; Linstedt, A. Manganese Blocks Intracellular Trafficking of Shiga Toxin and Protects Against Shiga Toxicosis. Science 2012, 335, 332–335. [Google Scholar] [CrossRef] [Green Version]
- Selyunin, A.S.; Hutchens, S.; McHardy, S.F.; Mukhopadhyay, S. Tamoxifen blocks retrograde trafficking of Shiga toxin 1 and 2 and protects against lethal toxicosis. Life Sci. Alliance 2019, 2, e201900439. [Google Scholar] [CrossRef]
- Sandvig, K.; Van Deurs, B. Membrane Traffic Exploited by Protein Toxins. Annu. Rev. Cell Dev. Boil. 2002, 18, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Johannes, L.; Popoff, V. Tracing the Retrograde Route in Protein Trafficking. Cell 2008, 135, 1175–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallard, F.; Antony, C.; Tenza, D.; Salamero, J.; Goud, B.; Johannes, L. Direct Pathway from Early/Recycling Endosomes to the Golgi Apparatus Revealed through the Study of Shiga Toxin B-fragment Transport. J. Cell Boil. 1998, 143, 973–990. [Google Scholar] [CrossRef]
- Linstedt, A.D.; Mehta, A.; Suhan, J.; Reggio, H.; Hauri, H.P. Sequence and overexpression of GPP130/GIMPc: Evidence for saturable pH-sensitive targeting of a type II early Golgi membrane protein. Mol. Boil. Cell 1997, 8, 1073–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bachert, C.; Lee, T.H.; Linstedt, A. Lumenal Endosomal and Golgi-Retrieval Determinants Involved in pH-sensitive Targeting of an Early Golgi Protein. Mol. Boil. Cell 2001, 12, 3152–3160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Natarajan, R.; Linstedt, A. A Cyclingcis-Golgi Protein Mediates Endosome-to-Golgi Traffic. Mol. Boil. Cell 2004, 15, 4798–4806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forrester, A.; Rathjen, S.J.; Garcia-Castillo, M.D.; Bachert, C.; Couhert, A.; Tepshi, L.; Pichard, S.; Martinez, J.; Munier, M.; Sierocki, R.; et al. Functional dissection of the retrograde Shiga toxin trafficking inhibitor Retro-2. Nat. Methods 2020, 16, 327–336. [Google Scholar] [CrossRef]
- Falguières, T.; Mallard, F.; Baron, C.; Hanau, D.; Lingwood, C.; Goud, B.; Salamero, J.; Johannes, L. Targeting of Shiga Toxin B-Subunit to Retrograde Transport Route in Association with Detergent-resistant Membranes. Mol. Boil. Cell 2001, 12, 2453–2468. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.C.; Sillence, D.J.; Falguières, T.; Jarvis, R.M.; Johannes, L.; Lord, J.M.; Platt, F.M.; Roberts, L.M. The Association of Shiga-like Toxin with Detergent-resistant Membranes Is Modulated by Glucosylceramide and Is an Essential Requirement in the Endoplasmic Reticulum for a Cytotoxic Effect. Mol. Boil. Cell 2006, 17, 1375–1387. [Google Scholar] [CrossRef] [Green Version]
- Saint-Pol, A.; Yélamos, B.; Amessou, M.; Mills, I.G.; Dugast, M.; Tenza, D.; Schu, P.; Antony, C.; McMahon, H.T.; Lamaze, C.; et al. Clathrin Adaptor epsinR Is Required for Retrograde Sorting on Early Endosomal Membranes. Dev. Cell 2004, 6, 525–538. [Google Scholar] [CrossRef] [Green Version]
- Lauvrak, S.U.; Torgersen, M.L.; Sandvig, K. Efficient endosome-to-Golgi transport of Shiga toxin is dependent on dynamin and clathrin. J. Cell Sci. 2004, 117, 2321–2331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhury, R.; Diao, A.; Zhang, F.; Eisenberg, E.; Saint-Pol, A.; Williams, C.; Konstantakopoulos, A.; Lucocq, J.; Johannes, L.; Rabouille, C.; et al. Lowe Syndrome Protein OCRL1 Interacts with Clathrin and Regulates Protein Trafficking between Endosomes and the Trans-Golgi Network. Mol. Boil. Cell 2005, 16, 3467–3479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Utskarpen, A.; Slagsvold, H.H.; Dyve, A.B.; Skånland, S.S.; Sandvig, K. SNX1 and SNX2 mediate retrograde transport of Shiga toxin. Biochem. Biophys. Res. Commun. 2007, 358, 566–570. [Google Scholar] [CrossRef] [PubMed]
- Bujny, M.V.; Popoff, V.; Johannes, L.; Cullen, P.J. The retromer component sorting nexin-1 is required for efficient retrograde transport of Shiga toxin from early endosome to the trans Golgi network. J. Cell Sci. 2007, 120, 2010–2021. [Google Scholar] [CrossRef] [Green Version]
- Popoff, V.; Mardones, G.; Tenza, D.; Rojas, R.; Lamaze, C.; Bonifacino, J.S.; Raposo, G.; Johannes, L. The retromer complex and clathrin define an early endosomal retrograde exit site. J. Cell Sci. 2007, 120, 2022–2031. [Google Scholar] [CrossRef] [Green Version]
- Naslavsky, N.; McKenzie, J.; Altan-Bonnet, N.; Sheff, D.; Caplan, S. EHD3 regulates early-endosome-to-Golgi transport and preserves Golgi morphology. J. Cell Sci. 2009, 122, 389–400. [Google Scholar] [CrossRef] [Green Version]
- Popoff, V.; Mardones, G.A.; Bai, S.-K.; Chambon, V.; Tenza, D.; Burgos, P.V.; Shi, A.; Benaroch, P.; Urbe, S.; Lamaze, C.; et al. Analysis of Articulation Between Clathrin and Retromer in Retrograde Sorting on Early Endosomes. Traffic 2009, 10, 1868–1880. [Google Scholar] [CrossRef]
- Hehnly, H.; Sheff, D.; Stamnes, M. Shiga Toxin Facilitates Its Retrograde Transport by Modifying Microtubule Dynamics. Mol. Boil. Cell 2006, 17, 4379–4389. [Google Scholar] [CrossRef] [Green Version]
- Hehnly, H.; Longhini, K.M.; Chen, J.-L.; Stamnes, M. Retrograde Shiga Toxin Trafficking Is Regulated by ARHGAP21 and Cdc42. Mol. Boil. Cell 2009, 20, 4303–4312. [Google Scholar] [CrossRef] [Green Version]
- Wilcke, M.; Johannes, L.; Galli, T.; Mayau, V.; Goud, B.; Salamero, J. Rab11 Regulates the Compartmentalization of Early Endosomes Required for Efficient Transport from Early Endosomes to the Trans-Golgi Network. J. Cell Boil. 2000, 151, 1207–1220. [Google Scholar] [CrossRef] [Green Version]
- Mallard, F.; Tang, B.L.; Galli, T.; Tenza, D.; Saint-Pol, A.; Yue, X.; Antony, C.; Hong, W.; Goud, B.; Johannes, L. Early/recycling endosomes-to-TGN transport involves two SNARE complexes and a Rab6 isoform. J. Cell Boil. 2002, 156, 653–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Nery, E.; Miserey-Lenkei, S.; Falguières, T.; Nizak, C.; Johannes, L.; Perez, F.; Goud, B. Rab6A and Rab6A′ GTPases Play Non-overlapping Roles in Membrane Trafficking. Traffic 2006, 7, 394–407. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Tai, G.; Hong, W. Autoantigen Golgin-97, an Effector of Arl1 GTPase, Participates in Traffic from the Endosome to the Trans-Golgi Network. Mol. Boil. Cell 2004, 15, 4426–4443. [Google Scholar] [CrossRef] [Green Version]
- Yoshino, A.; Setty, S.R.G.; Poynton, C.; Whiteman, E.L.; Saint-Pol, A.; Burd, C.; Johannes, L.; Holzbaur, E.L.; Koval, M.; McCaffery, J.M.; et al. tGolgin-1 (p230, golgin-245) modulates Shiga-toxin transport to the Golgi and Golgi motility towards the microtubule-organizing centre. J. Cell Sci. 2005, 118, 2279–2293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derby, M.C.; Lieu, Z.Z.; Brown, D.; Stow, J.L.; Goud, B.; Gleeson, P.A. The trans-Golgi Network Golgin, GCC185, is Required for Endosome-to-Golgi Transport and Maintenance of Golgi Structure. Traffic 2007, 8, 758–773. [Google Scholar] [CrossRef]
- Yamane, J.; Kubo, A.; Nakayama, K.; Yuba-Kubo, A.; Katsuno, T.; Tsukita, S.; Tsukita, S. Functional involvement of TMF/ARA160 in Rab6-dependent retrograde membrane traffic. Exp. Cell Res. 2007, 313, 3472–3485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Victoria, F.J.; Mardones, G.A.; Bonifacino, J.S. Requirement of the Human GARP Complex for Mannose 6-phosphate-receptor-dependent Sorting of Cathepsin D to Lysosomes. Mol. Boil. Cell 2008, 19, 2350–2362. [Google Scholar] [CrossRef] [Green Version]
- Tai, G.; Lu, L.; Wang, T.L.; Tang, B.L.; Goud, B.; Johannes, L.; Hong, W. Participation of the syntaxin 5/Ykt6/GS28/GS15 SNARE complex in transport from the early/recycling endosome to the trans-Golgi network. Mol. Biol. Cell 2004, 15, 4011–4022. [Google Scholar] [CrossRef] [Green Version]
- Wälchli, S.; Skånland, S.S.; Gregers, T.F.; Lauvrak, S.U.; Torgersen, M.L.; Ying, M.; Kuroda, S.; Maturana, A.; Sandvig, K. The Mitogen-activated Protein Kinase p38 Links Shiga Toxin-dependent Signaling and Trafficking. Mol. Boil. Cell 2008, 19, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Torgersen, M.L.; Walchli, S.; Grimmer, S.; Skanland, S.S.; Sandvig, K. Protein kinase Cdelta is activated by Shiga toxin and regulates its transport. J. Biol. Chem. 2007, 282, 16317–16328. [Google Scholar] [CrossRef] [Green Version]
- Lingelem, A.B.D.; Bergan, J.; Sandvig, K. Inhibitors of Intravesicular Acidification Protect Against Shiga Toxin in a pH-Independent Manner. Traffic 2011, 13, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Suchy, S.F.; Olivos-Glander, I.M.; Nussbaum, R.L. Lowe Syndrome, a deficiency of a phosphatidyl-inositol 4,5-bisphosphate 5-phosphatase in the Golgi apparatus. Hum. Mol. Genet. 1995, 4, 2245–2250. [Google Scholar] [CrossRef] [PubMed]
- Bonifacino, J.S.; Hurley, J.H. Retromer. Curr. Opin. Cell Biol. 2008, 20, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, S.R. Rab GTPases: Master regulators that establish the secretory and endocytic pathways. Mol. Boil. Cell 2017, 28, 712–715. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, E.; Haas, A.K.; Spooner, R.A.; Yoshimura, S.-I.; Lord, J.M.; Barr, F.A. Specific Rab GTPase-activating proteins define the Shiga toxin and epidermal growth factor uptake pathways. J. Cell Boil. 2007, 177, 1133–1143. [Google Scholar] [CrossRef]
- Munro, S. The Golgin Coiled-Coil Proteins of the Golgi Apparatus. Cold Spring Harb. Perspect. Boil. 2011, 3, a005256. [Google Scholar] [CrossRef] [Green Version]
- Yoon, T.-Y.; Munson, M. SNARE complex assembly and disassembly. Curr. Boil. 2018, 28, R397–R401. [Google Scholar] [CrossRef] [Green Version]
- Selyunin, A.S.; Iles, L.R.; Bartholomeusz, G.; Mukhopadhyay, S. Genome-wide siRNA screen identifies UNC50 as a regulator of Shiga toxin 2 trafficking. J. Cell Boil. 2017, 216, 3249–3262. [Google Scholar] [CrossRef] [Green Version]
- Poluzzi, C.; Casulli, J.; Goyal, A.; Mercer, T.J.; Neill, T.; Iozzo, R.V. Endorepellin Evokes Autophagy in Endothelial Cells. J. Boil. Chem. 2014, 289, 16114–16128. [Google Scholar] [CrossRef] [Green Version]
- Akizu, N.; Cantagrel, V.; Zaki, M.; Al-Gazali, L.; Wang, X.; Rosti, R.O.; Dikoglu, E.; Gelot, A.B.; Rosti, B.; Vaux, K.; et al. Biallelic mutations in SNX14 cause a syndromic form of cerebellar atrophy and lysosome-autophagosome dysfunction. Nat. Genet. 2015, 47, 528–534. [Google Scholar] [CrossRef]
- García-Rúa, V.; Feijóo-Bandín, S.; Rodríguez-Penas, D.; Mosquera-Leal, A.; Abu-Assi, E.; Beiras, A.; Seoane, L.M.; Lear, P.; Parrington, J.; Portolés, M.; et al. Endolysosomal two-pore channels regulate autophagy in cardiomyocytes. J. Physiol. 2016, 594, 3061–3077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, P.; Nishimura, T.; Sakamaki, Y.; Itakura, E.; Hatta, T.; Natsume, T.; Mizushima, N. The HOPS complex mediates autophagosome–lysosome fusion through interaction with syntaxin 17. Mol. Boil. Cell 2014, 25, 1327–1337. [Google Scholar] [CrossRef] [PubMed]
- Luzio, J.P.; Pryor, P.R.; Bright, N.A. Lysosomes: Fusion and function. Nat. Rev. Mol. Cell Boil. 2007, 8, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Pols, M.; Brink, C.T.; Gosavi, P.; Oorschot, V.; Klumperman, J. The HOPS Proteins hVps41 and hVps39 Are Required for Homotypic and Heterotypic Late Endosome Fusion. Traffic 2012, 14, 219–232. [Google Scholar] [CrossRef]
- Wartosch, L.; Günesdogan, U.; Graham, S.C.; Luzio, J.P. Recruitment of VPS33A to HOPS by VPS16 Is Required for Lysosome Fusion with Endosomes and Autophagosomes. Traffic 2015, 16, 727–742. [Google Scholar] [CrossRef] [Green Version]
- Rink, J.; Ghigo, E.; Kalaidzidis, Y.; Zerial, M. Rab Conversion as a Mechanism of Progression from Early to Late Endosomes. Cell 2005, 122, 735–749. [Google Scholar] [CrossRef] [Green Version]
- Komatsu, M.; Waguri, S.; Ueno, T.; Iwata, J.; Murata, S.; Tanida, I.; Ezaki, J.; Mizushima, N.; Ohsumi, Y.; Uchiyama, Y.; et al. Impairment of starvation-induced and constitutive autophagy in Atg7-deficient mice. J. Cell Boil. 2005, 169, 425–434. [Google Scholar] [CrossRef]
- Saenz, J.B.; Sun, W.J.; Chang, J.W.; Li, J.; Bursulaya, B.; Gray, N.S.; Haslam, D.B. Golgicide A reveals essential roles for GBF1 in Golgi assembly and function. Nat. Methods 2009, 5, 157–165. [Google Scholar] [CrossRef] [Green Version]
- Morgens, D.W.; Chan, C.; Kane, A.J.; Weir, N.R.; Li, A.; Dubreuil, M.M.; Tsui, C.K.; Hess, G.T.; Lavertu, A.; Han, K.; et al. Retro-2 protects cells from ricin toxicity by inhibiting ASNA1-mediated ER targeting and insertion of tail-anchored proteins. eLife 2019, 8, 48434. [Google Scholar] [CrossRef]
- Noel, R.; Gupta, N.; Pons, V.; Goudet, A.; Garcia-Castillo, M.D.; Michau, A.; Martinez, J.; Buisson, D.-A.; Johannes, L.; Gillet, D.; et al. N-Methyldihydroquinazolinone Derivatives of Retro-2 with Enhanced Efficacy against Shiga Toxin. J. Med. Chem. 2013, 56, 3404–3413. [Google Scholar] [CrossRef]
- Gupta, N.; Pons, V.; Noël, R.; Buisson, D.-A.; Michau, A.; Johannes, L.; Gillet, D.; Barbier, J.; Cintrat, J.-C. (S)-N-Methyldihydroquinazolinones are the Active Enantiomers of Retro-2 Derived Compounds against Toxins. ACS Med. Chem. Lett. 2013, 5, 94–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Secher, T.; Shima, A.; Hinsinger, K.; Cintrat, J.-C.; Johannes, L.; Barbier, J.; Gillet, D.; Oswald, E. Retrograde Trafficking Inhibitor of Shiga Toxins Reduces Morbidity and Mortality of Mice Infected with Enterohemorrhagic Escherichia coli. Antimicrob. Agents Chemother. 2015, 59, 5010–5013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukhopadhyay, S.; Bachert, C.; Smith, N.R.; Linstedt, A. Manganese-induced Trafficking and Turnover of the cis-Golgi Glycoprotein GPP130. Mol. Boil. Cell 2010, 21, 1282–1292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukhopadhyay, S.; Linstedt, A. Identification of a gain-of-function mutation in a Golgi P-type ATPase that enhances Mn2+ efflux and protects against toxicity. Proc. Natl. Acad. Sci. USA 2010, 108, 858–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuda, M.; Braun-Sommargren, M.; Crooks, D.; Smith, D.R. Golgi phosphoprotein 4 (GPP130) is a sensitive and selective cellular target of manganese exposure. Synapsis 2013, 67, 205–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tewari, R.; Jarvela, T.; Linstedt, A. Manganese induces oligomerization to promote down-regulation of the intracellular trafficking receptor used by Shiga toxin. Mol. Boil. Cell 2014, 25, 3049–3058. [Google Scholar] [CrossRef]
- Tewari, R.; Bachert, C.; Linstedt, A. Induced oligomerization targets Golgi proteins for degradation in lysosomes. Mol. Boil. Cell 2015, 26, 4427–4437. [Google Scholar] [CrossRef]
- Taylor, C.A.; Tuschl, K.; Nicolai, M.M.; Bornhorst, J.; Gubert, P.; Varão, A.M.; Aschner, M.; Smith, D.R.; Mukhopadhyay, S. Maintaining Translational Relevance in Animal Models of Manganese Neurotoxicity. J. Nutr. 2020, 066. [Google Scholar] [CrossRef]
- Van Weert, A.W.; Dunn, K.W.; Gueze, H.J.; Maxfield, F.R.; Stoorvogel, W. Transport from late endosomes to lysosomes, but not sorting of integral membrane proteins in endosomes, depends on the vacuolar proton pump. J. Cell Boil. 1995, 130, 821–834. [Google Scholar] [CrossRef] [Green Version]
- Huotari, J.; Helenius, A. Endosome maturation. EMBO J. 2011, 30, 3481–3500. [Google Scholar] [CrossRef]
- Morello, K.C.; Wurz, G.T.; DeGregorio, M.W. Pharmacokinetics of Selective Estrogen Receptor Modulators. Clin. Pharmacokinet. 2003, 42, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Schindler, M.; Simon, S.M. A Mechanism for Tamoxifen-mediated Inhibition of Acidification. J. Boil. Chem. 1999, 274, 18364–18373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altan, N.; Chen, Y.; Schindler, M.; Simon, S.M. Tamoxifen inhibits acidification in cells independent of the estrogen receptor. Proc. Natl. Acad. Sci. USA 1999, 96, 4432–4437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Touitou, I.; Mathieu, M.; Rochefort, H. Stable transfection of the estrogen receptor cDNA into Hela cells induces estrogen responsiveness of endogenous cathepsin D gene but not of cell growth. Biochem. Biophys. Res. Commun. 1990, 169, 109–115. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Function | Reference |
|---|---|---|
| GPP130 | Endosomal sorting receptor | [19] |
| Cholesterol | Microdomain organization | [28] |
| Clathrin | Coat | [30,31] |
| EpsinR | Clathrin adaptor | [30] |
| OCRL1 | Phosphatidylinositol 4,5-bisphosphate 5-phosphatase | [32] |
| Retromer (SNX1, SNX2, Vps26/29/35) | Coat | [33,34,35] |
| EHD3 | Retromer interactor | [36] |
| Rabenosyn5 | EHD3 interactor | [36] |
| RME-8 | Recruit Hsc-70 to SNX1 | [37] |
| Hsc-70 | Clathrin uncoating ATPase | [37] |
| Hrs | Clathrin interactor | [37] |
| Dynamin | Membrane scission | [31] |
| Microtubule | Cytoskeleton | [38] |
| Dynein | Motor | [38] |
| Actin | Cytoskeleton | [39] |
| Cdc42 | Rho GTPase | [39] |
| ARHGAP21 | Cdc42 activating protein | [39] |
| Rab11 | Small GTPase | [40,41] |
| Rab6a’ | Small GTPase | [41,42] |
| Arl1 | Small GTPase, Golgin anchor | [43] |
| Golgin 97 | Tether, Arl1 effector | [43] |
| Golgin 245 | Tether, Arl1 effector | [44] |
| GCC185 | Tether, Arl1 effector | [45] |
| TMF | Tether, Rab6 effector | [46] |
| GARP | Tether | [47] |
| Syntaxin 5, Ykt6, GS15, GS28 | SNARE complex | [48] |
| Syntaxin 6, syntaxin 16, Vti1a, Vamp3/4 | SNARE complex | [41] |
| p38 | Kinase | [49] |
| PKCδ | Kinase | [50] |
| V-ATPase | Proton pump | [51] |
| Factor | Function | Reference |
|---|---|---|
| Dynamin | Membrane scission | [14] |
| EpsinR | Clathrin adaptor | [14] |
| Vps26 | Retromer component | [14] |
| Syntaxin 5 | SNARE complex component | [14] |
| UNC50 | GBF1 interactor, recruits GBF1 to the Golgi | [58] |
| Rab2a | Small GTPase | [20] |
| STAM | ESCRT (endosomal sorting complexes required for transport)-0 complex component | [20] |
| FUT1 | Fucosylation enzyme | [20] |
| Vps39 | HOPS (homotypic fusion and vacuole protein sorting) complex component | [20] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, D.; Selyunin, A.; Mukhopadhyay, S. Targeting the Early Endosome-to-Golgi Transport of Shiga Toxins as a Therapeutic Strategy. Toxins 2020, 12, 342. https://doi.org/10.3390/toxins12050342
Li D, Selyunin A, Mukhopadhyay S. Targeting the Early Endosome-to-Golgi Transport of Shiga Toxins as a Therapeutic Strategy. Toxins. 2020; 12(5):342. https://doi.org/10.3390/toxins12050342
Chicago/Turabian StyleLi, Danyang, Andrey Selyunin, and Somshuvra Mukhopadhyay. 2020. "Targeting the Early Endosome-to-Golgi Transport of Shiga Toxins as a Therapeutic Strategy" Toxins 12, no. 5: 342. https://doi.org/10.3390/toxins12050342
APA StyleLi, D., Selyunin, A., & Mukhopadhyay, S. (2020). Targeting the Early Endosome-to-Golgi Transport of Shiga Toxins as a Therapeutic Strategy. Toxins, 12(5), 342. https://doi.org/10.3390/toxins12050342