Cylindrospermopsin-Microcystin-LR Combinations May Induce Genotoxic and Histopathological Damage in Rats

 , ,
, ,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Micronucleus Assay
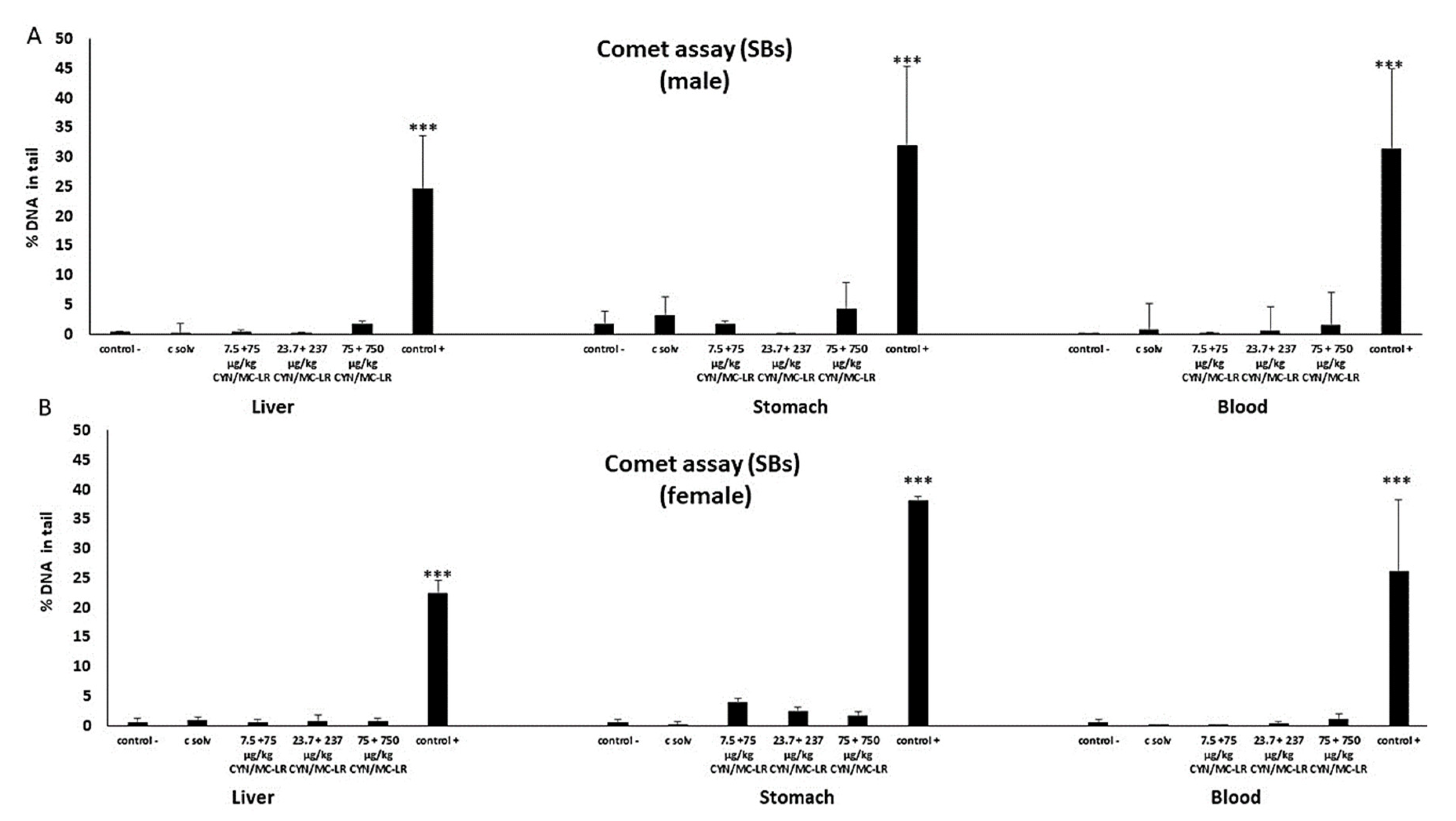
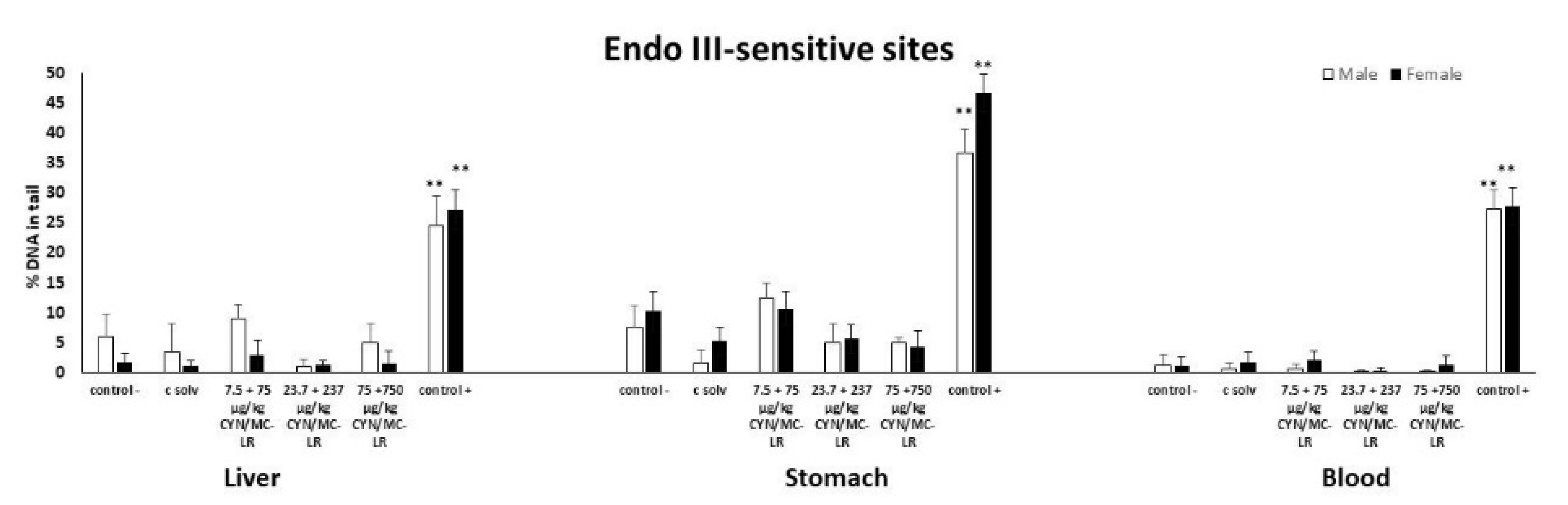
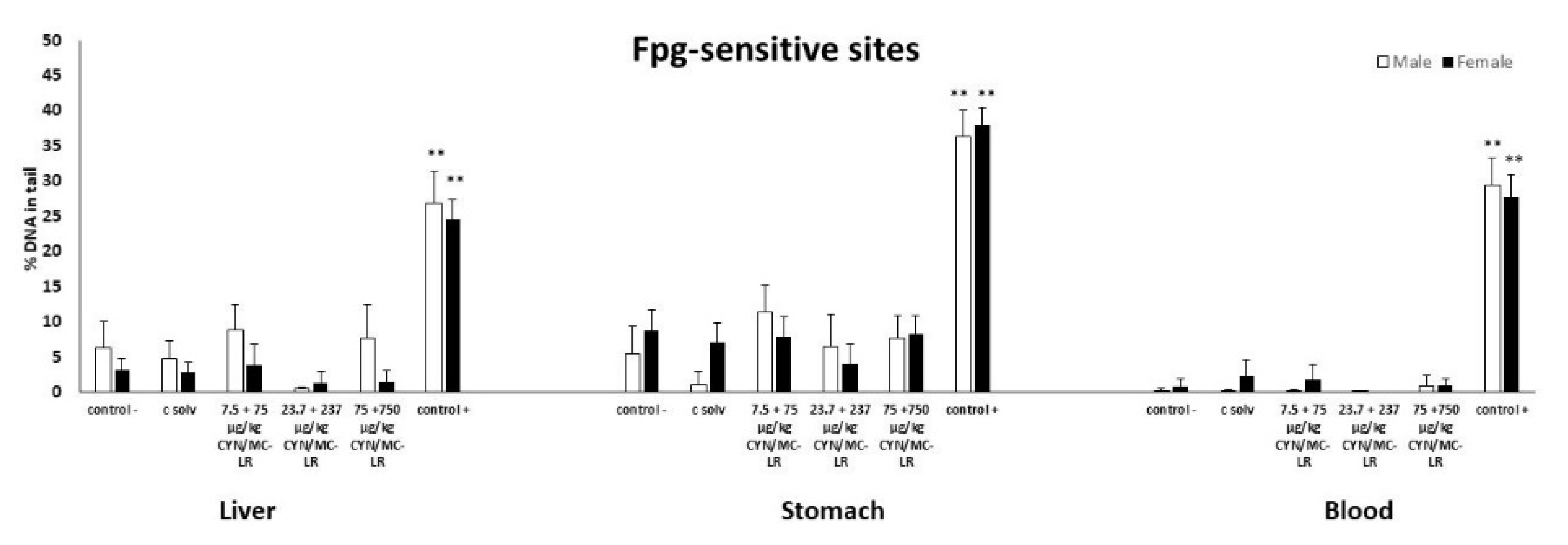
2.2. Standard and Enzyme-Modified Comet Assay
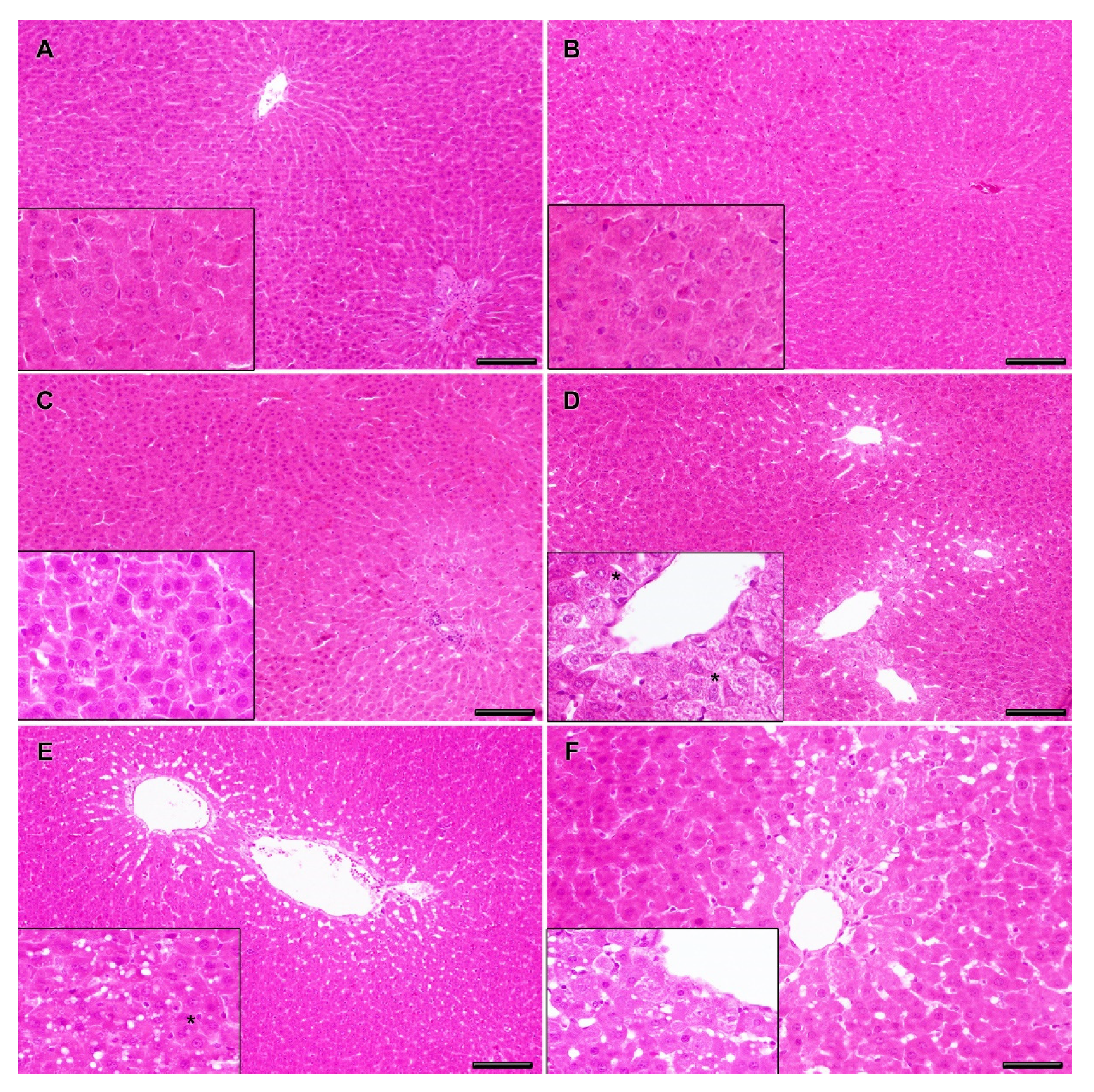
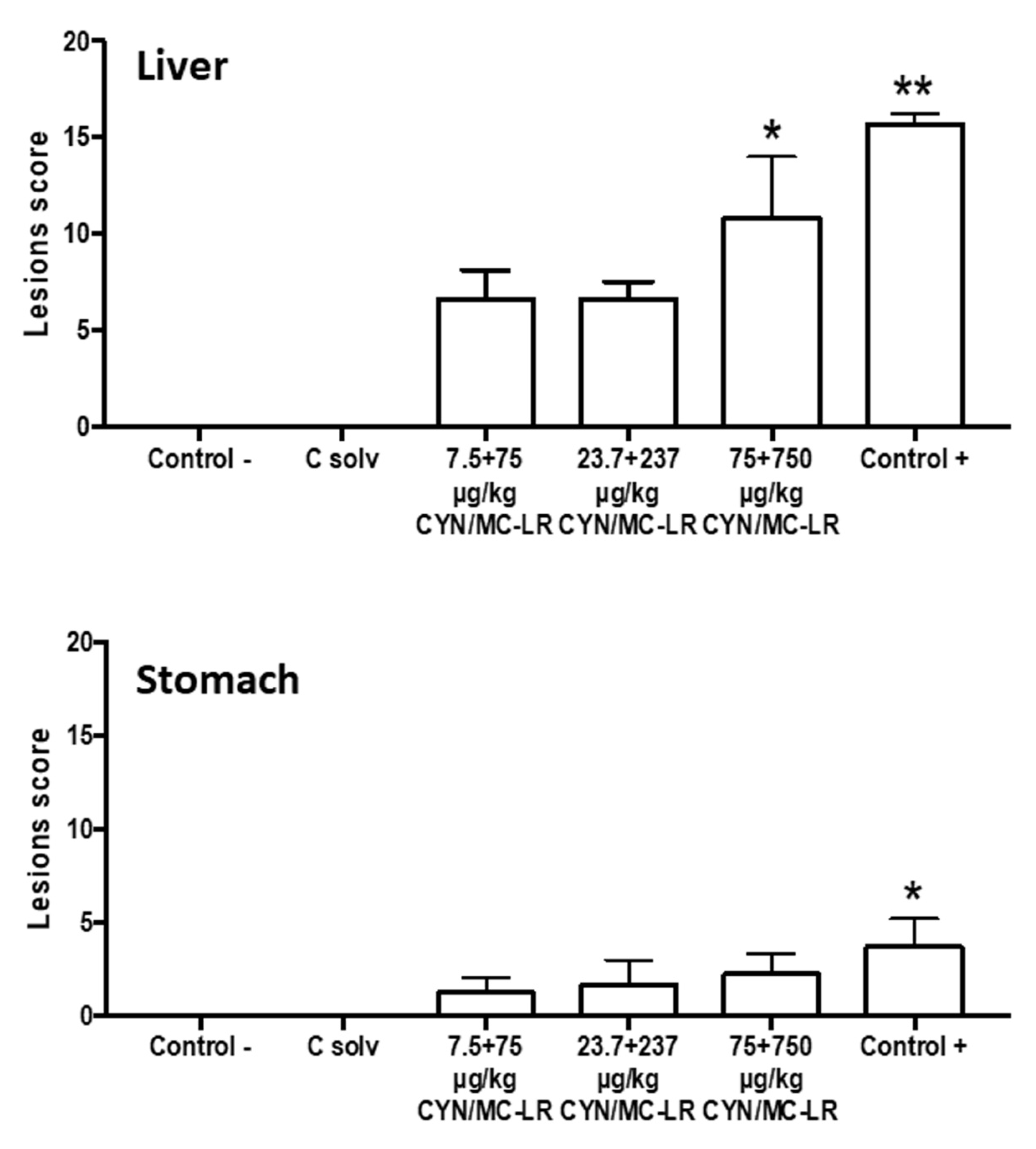
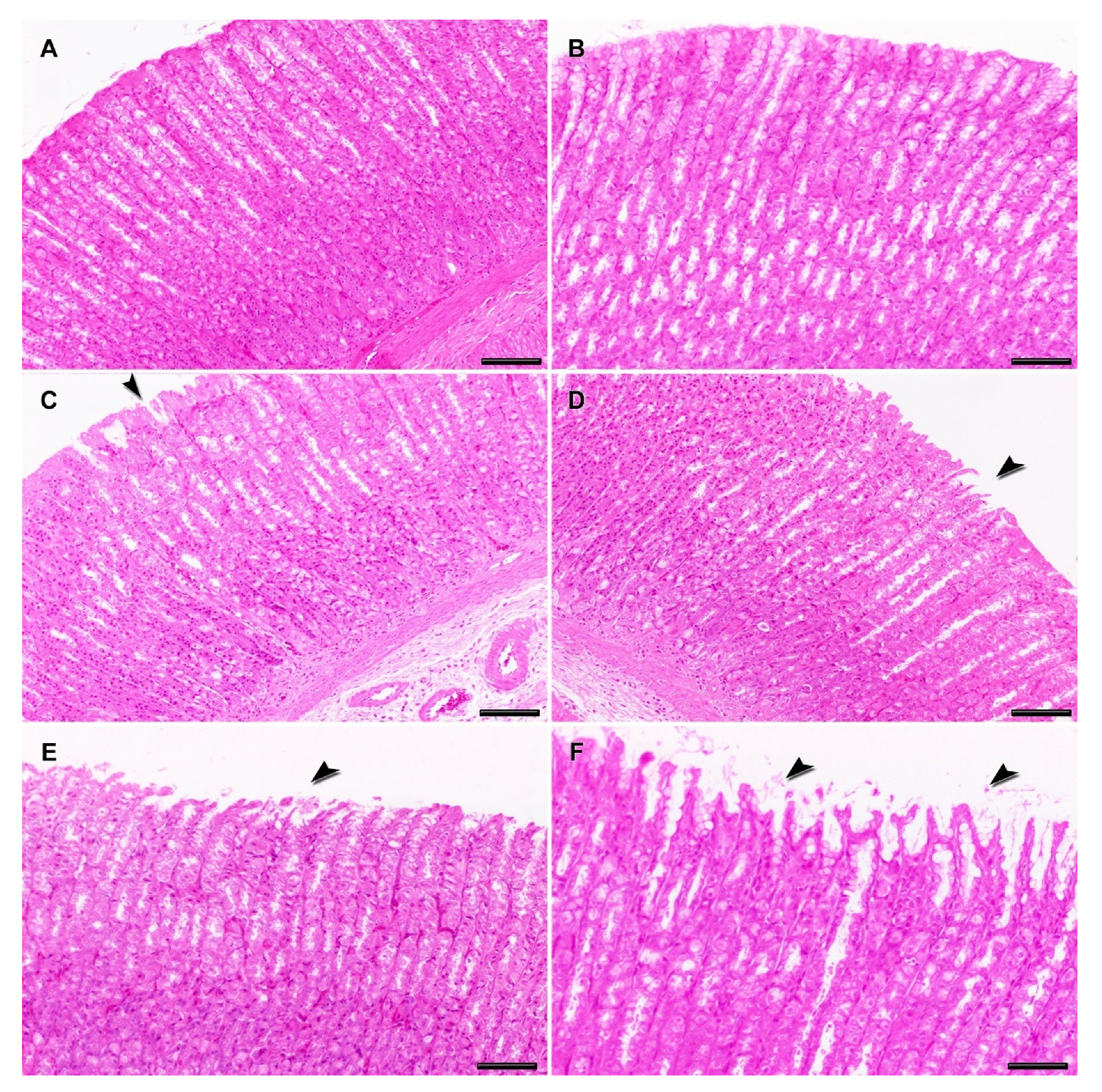
2.3. Clinical and Histopathological Analysis
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals and Reagents
5.2. Animal Housing and Feeding Conditions
5.3. Experimental Design and Treatment
5.4. Sample Collection
5.5. Micronucleus Assay
5.6. Standard and Enzyme-Modified Comet Assay
5.7. Histopathological Analysis
5.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Glibert, P.M. Harmful algae at the complex nexus of eutrophication and climate change. Harmful Algae 2020, 91, 101583. [Google Scholar] [CrossRef] [PubMed]
- Buratti, F.M.; Manganelli, M.; Vichi, S.; Stefanelli, M.; Scardala, S.; Testai, E.; Funari, E. Cyanotoxins: Producing organisms, occurrence, toxicity, mechanism of action and human health toxicological risk evaluation. Arch. Toxicol. 2017, 91, 1049–1130. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, W.W. Cyanobacteria secondary metabolites – the cyanotoxins. J. Appl. Bacteriol. 1992, 72, 445–459. [Google Scholar] [CrossRef]
- Environmental Protection Agency. Health Effects Support Document for the Cyanobacterial Toxin Cylindrospermopsin. 2015. Available online: https://www.epa.gov/sites/production/files/2017-06/documents/cylindrospermopsin-support-report-2015.pdf (accessed on 4 May 2020).
- Gutiérrez-Praena, D.; Jos, Á.; Pichardo, S.; Moreno, I.M.; Cameán, A.M. Presence and bioaccumulation of microcystins and cylindrospermopsin in food and the effectiveness of some cooking techniques at decreasing their concentrations: A review. Food Chem. Toxicol. 2013, 53, 139–152. [Google Scholar] [CrossRef]
- Gupta, N.; Pant, S.C.; Vijayaraghavan, R.; Lakshmana Rao, P.V. Comparative toxicity evaluation of cyanobacterial cyclic peptide toxin microcystin variants (LR, RR, YR) in mice. Toxicology 2003, 188, 285–296. [Google Scholar] [CrossRef]
- Spoof, L.; Catherine, A. Appendix 3: Tables of microcystins and nodularins. Handb. Cyanobact. Monit. Cyanotoxin Anal. 2017, 526–537. [Google Scholar] [CrossRef]
- Diez-Quijada, L.; Prieto, A.I.; Guzmán-Guillen, R.; Jos, Á.; Cameán, A.M. Occurrence and toxicity of microcystin congeners other than MC-LR and MC-RR: A review. Food Chem. Toxicol. 2019, 125, 106–132. [Google Scholar] [CrossRef]
- Puerto, M.; Pichardo, S.; Jos, Á.; Cameán, A.M. Comparison of the toxicity induced by microcystin-RR and microcystin-YR in differentiated and undifferentiated Caco-2 cells. Toxicon 2009, 54, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Fischer, W.J.; Altheimer, S.; Cattori, V.; Meier, P.J.; Dietrich, D.R.; Hagenbuch, B. Organic anion transporting polypeptides expressed in liver and brain mediate uptake of microcystin. Toxicol. Appl. Pharmacol. 2005, 203, 257–263. [Google Scholar] [CrossRef] [Green Version]
- MacKintosh, C.; Beattie, K.A.; Klumpp, S.; Cohen, P.; Codd, G.A. Cyanobacterial microcystin-LR is a potent and specific inhibitor of protein phosphatases 1 and 2A from both mammals and higher plants. FEBS Lett. 1990, 264, 187–192. [Google Scholar] [CrossRef] [Green Version]
- Falconer, I.R.; Yeung, D.S.K. Cytoskeletal changes in hepatocytes induced by Microcystis toxins and their relation to hyperphosphorylation of cell proteins. Chem. Biol. Interact. 1992, 81, 181–196. [Google Scholar] [CrossRef]
- Chorus, I.; Bartram, J. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; Chorus, I., Bertram, J., Eds.; E & FN Spon: London, UK, 1999; pp. 1–400. ISBN 0-419-23930-8. [Google Scholar]
- Guzman, R.E.; Solter, P.F. Hepatic oxidative stress following prolonged subletal Microcystin-LR exposure. Toxicol. Pathol. 1999, 27, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Ding, W.-X.; Shen, H.-M.; Ong, C.-N. Critical role of reactive oxygen species formation in microcystin-induced cytoskeleton disruption in primary cultured hepatocytes. J. Toxicol. Environ. Health A 2001, 64, 507–519. [Google Scholar] [CrossRef] [PubMed]
- Žegura, B.; Lah, T.T.; Filipič, M. The role of reactive oxygen species in microcystin-LR induced DNA damage. Toxicology 2004, 200, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Puerto, M.; Pichardo, S.; Jos, Á.; Prieto, A.I.; Sevilla, E.; Frías, J.E.; Cameán, A.M. Differential oxidative stress response to pure Microcystin-LR and Microcystin-containing and non-containing cyanobacterial crude extracts on Caco-2 cells. Toxicon 2010, 55, 514–522. [Google Scholar] [CrossRef]
- Nishiwaki-Matsushima, R.; Ohta, T.; Nishiwaki, S.; Suganuma, M.; Kohyama, K.; Ishikawa, T.; Carmichael, W.W.; Fujiki, H. Liver tumor promotion by the cyanobacterial cyclic peptide toxin microcystin-LR. J. Cancer Res. Clin. Oncol. 1992, 118, 420–424. [Google Scholar] [CrossRef]
- Abramsson-Zetterberg, L.; Sundh, U.B.; Mattsson, R. Cyanobacterial extracts and microcystin-LR are inactive in the micronucleus assay in vivo and in vitro. Mutat. Res. 2010, 699, 5–10. [Google Scholar] [CrossRef]
- Žegura, B. An overview of the mechanisms of microcystin-LR genotoxicity and potential carcinogenicity. Mini Rev. Med. Chem. 2016, 16, 1042–1062. [Google Scholar] [CrossRef]
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. Ingested Nitrate and Nitrite, and Cyanobacterial Peptide Toxins. 2010; World Health Organization, International Agency for Research on Cancer. Available online: https://www.ncbi.nlm.nih.gov/books/NBK326544/pdf/Bookshelf_NBK326544.pdf (accessed on 6 April 2020).
- Žegura, B.; Štraser, A.; Filipič, M. Genotoxicity and potential carcinogenicity of cyanobacterial toxins – a review. Mutat. Res.-Rev. Mutat. Res. 2011, 727, 16–41. [Google Scholar] [CrossRef]
- Rao, P.V.L.; Bhattacharya, R. The cyanobacterial toxin Microcystin-LR induced DNA damage in mouse liver in vivo. Toxicology 1996, 114, 29–36. [Google Scholar] [CrossRef]
- Rao, P.V.L.; Gupta, N.; Jayaraj, R.; Bhaskar, A.S.B.; Jatav, P.C. Age-dependent effects on biochemical variables and toxicity induced by cyclic peptide toxin microcystin-LR in mice. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2005, 140, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Gaudin, J.; Huet, S.; Jarry, G.; Fessard, V. in vivo DNA damage induced by the cyanotoxin microcystin-LR: Comparison of intra-peritoneal and oral administrations by use of the comet assay. Mutat. Res. 2008, 652, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Dias, E.; Louro, H.; Pinto, M.; Santos, T.; Antunes, S.; Pereira, P.; Silva, M.J. Genotoxicity of microcystin-LR in in vitro and in vivo experimental models. BioMed. Res. Int. 2014, 2014, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Zhang, X.; Xie, P.; Liang, H.; Zhang, X. The suppression of hematopoiesis function in Balb/c mice induced by prolonged exposure of microcystin-LR. Toxicol. Lett. 2013, 219, 194–201. [Google Scholar] [CrossRef]
- Gaudin, J.; Le Hegarat, L.; Nesslany, F.; Marzin, D.; Fessard, V. In vivo genotoxic potential of microcystin-LR: A cyanobacterial toxin, investigated both by the unscheduled DNA synthesis (UDS) and the comet assays after intravenous administration. Environ. Toxicol. 2009, 24, 200–209. [Google Scholar] [CrossRef]
- Zhan, L.; Honma, M.; Wang, L.; Hayashi, M.; Wu, D.-S.; Zhang, L.-S.; Rajaguru, P.; Suzuki, T. Microcystin-LR is not Mutagenic in vivo in the λ/lacZ Transgenic Mouse (MutaTMMouse). Gene. Environ. 2006, 28, 68–73. [Google Scholar] [CrossRef] [Green Version]
- Ohtani, I.; Moore, R.E.; Runnegar, M.T.C. Cylindrospermopsin: A potent hepatotoxin from the blue-green alga Cylindrospermopsis raciborskii. J. Am. Chem. Soc. 1992, 114, 7941–7942. [Google Scholar] [CrossRef]
- Kinnear, S. Cylindrospermopsin: A decade of progress on bioaccumulation research. Mar. Drugs 2010, 8, 542–564. [Google Scholar] [CrossRef] [Green Version]
- Manning, S.R.; Nobles, D.R. Impact of global warming on water toxicity: Cyanotoxins. Curr. Opin. Food Sci. 2017, 18, 14–20. [Google Scholar] [CrossRef]
- Pichardo, S.; Cameán, A.M.; Jos, Á. In vitro toxicological assessment of Cylindrospermopsin: A review. Toxins 2017, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Terao, K.; Ohmori, S.; Igarashi, K.; Ohtani, I.; Watanabe, M.F.; Harada, K.I.; Ito, E.; Watanabe, M. Electron microscopic studies on experimental poisoning in mice induced by cylindrospermopsin isolated from blue-green alga Umezakia natans. Toxicon 1994, 32, 833–843. [Google Scholar] [CrossRef]
- Runnegar, M.T.; Kong, S.-M.; Zhong, Y.-Z.; Lu, S.C. Inhibition of reduced glutathione synthesis by cyanobacterial alkaloid cylindrospermopsin in cultured rat hepatocytes. Biochem. Pharmacol. 1995, 49, 219–225. [Google Scholar] [CrossRef]
- Froscio, S.M.; Humpage, A.R.; Burcham, P.C.; Falconer, I.R. Cylindrospermopsin-induced protein synthesis inhibition and its dissociation from acute toxicity in mouse hepatocytes. Environ. Toxicol. Int. J. 2003, 18, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Puerto, M.; Jos, Á.; Pichardo, S.; Moyano, R.; Blanco, A.; Cameán, A.M. Acute exposure to pure Cylindrospermopsin results in oxidative stress and pathological alterations in Tilapia (Oreochromis niloticus). Environ. Toxicol. 2014, 29, 371–385. [Google Scholar] [CrossRef] [PubMed]
- Humpage, A.R.; Fontaine, F.; Froscio, S.; Burcham, P.; Falconer, I.R. Cylindrospermopsin genotoxicity and cytotoxicity: Role of cytochrome P-450 and oxidative stress. J. Toxicol. Environ. Health A 2005, 68, 739–753. [Google Scholar] [CrossRef]
- Bazin, E.; Mourot, A.; Humpage, A.R.; Fessard, V. Genototoxicity of a freshwater cyanotoxin, cylindrospermopsin, in two human cell lines: Caco-2 and HepaRG. Environ. Mol. Mutat. 2010, 51, 251–259. [Google Scholar] [CrossRef]
- Shen, X.; Lam, P.K.S.; Shaw, G.R.; Wickramasinghe, W. Genotoxity investigation of a cyanobacterial toxin, cylindrospermopsin. Toxicon 2002, 40, 1499–1501. [Google Scholar] [CrossRef]
- Bazin, E.; Huet, S.; Jarry, G.; Le Hégarat, L.; Munday, J.S.; Humpage, A.R.; Fessard, V. Cytotoxic and genotoxic effects of cylindrospermopsin in mice treated by gavage or intraperitoneal injection. Environ. Toxicol. 2012, 27, 277–284. [Google Scholar] [CrossRef]
- Dordevic, N.B.; Matíc, S.L.J.; Simić, S.B.; Stanić, S.M.; Mihailivić, V.B.; Stancović, N.M.; Stancović, V.D.; Cirić, A.R. Impact of the toxicity of Cylindrospermopsis raciboskii (Woloszynska) Seenayya & Subba Raju on laboratory rats in vivo. Environ. Sci. Pollut. Res. 2017, 24, 14259–14272. [Google Scholar] [CrossRef]
- Diez-Quijada, L.; Llana-Ruiz-Cabello, M.; Cătunescu, M.G.; Puerto, M.; Moyano, R.; Jos, Á.; Cameán, A.M. In vivo genotoxicity evaluation of cylindrospermopsin in rats using a combined micronucleus and comet assay. Food. Chem. Toxicol. 2019, 132, 1–9. [Google Scholar] [CrossRef]
- Bittencourt-Oliveira, M.; Carmo, D.; Piccin-Santos, V.; Moura, A.N.; Aragão-Tavares, N.K.; Cordeiro-Araújo, M.K. Cyanobacteria, microcystins and cylindrospermopsin in public drinking supply reservoirs of Brazil. An. Acad. Bras. Cienc. 2014, 86, 297–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jančula, D.; Straková, L.; Sadílek, J.; Maršálek, B.; Babica, P. Survey of cyanobacterial toxins in Czech water reservoirs—The first observation of neurotoxic saxitoxins. Environ. Sci. Pollut. Res. Int. 2014, 21, 8006–8015. [Google Scholar] [CrossRef] [PubMed]
- León, C.; Peñuela, G.A. Detected cyanotoxins by UHPLC MS/MS technique in tropical reservoirs of northeastern Colombia. Toxicon 2019, 167, 38–48. [Google Scholar] [CrossRef]
- Testai, E.; Buratti, F.M.; Funari, E.; Manganelli, M.; Vichi, S.; Arnich, N.; Biré, R.; Fessard, V.; Sialehaamoa, A. Review and analysis of occurrence, exposure and toxicity of cyanobacteria toxins in food. EFSA Support. Publ. 2016, 13, 1–309. [Google Scholar] [CrossRef]
- Hercog, K.; Maisanaba, S.; Filipič, M.; Jos, Á.; Cameán, A.M.; Žegura, B. Genotoxic potential of the binary mixture of cyanotoxins microcystin-LR and cylindrospermopsin. Chemosphere 2017, 189, 319–329. [Google Scholar] [CrossRef]
- Diez-Quijada, L.; Prieto, A.I.; Jos, Á.; Cameán, A.M. in vitro mutagenic and genotoxic assessment of a mixture of the cyanotoxins Microcystin-LR and Cylindrospermopsin. Toxins 2019, 11, 318. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-Praena, D.; Guzmán-Guillén, R.; Pichardo, S.; Moreno, F.J.; Vasconcelos, V.; Jos, Á.; Cameán, A.M. Cytotoxic and morphological effects of microcystin-LR, cylindrospermopsin, and their combinations on the human hepatic cell line HepG2. Environ. Toxicol. 2018, 34, 240–251. [Google Scholar] [CrossRef] [PubMed]
- Hinojosa, M.G.; Prieto, A.I.; Gutiérrez-Praena, D.; Moreno, F.J.; Cameán, A.M.; Jos, Á. Neurotoxic assessment of Microcystin-LR, cylindrospermopsin and their combination on the human neuroblastoma SH-SY5Y cell line. Chemosphere 2019, 224, 751–764. [Google Scholar] [CrossRef] [PubMed]
- EFSA Scientific Committee. Scientific opinion on genotoxicity testing strategies applicable to food and feed safety assessment. EFSA J. 2011, 9, 2379. [Google Scholar] [CrossRef]
- OECD Guidelines for the Testing of Chemicals: Mammalian Erythrocyte Micronucleus Test. 2016. Guideline 474. pp. 1–21. Available online: https://www.oecd.org/env/test-no-474-mammalian-erythrocyte-micronucleus-test-9789264264762-en.htm (accessed on 6 April 2020).
- OECD Guideline for the Testing of Chemicals: In vivo Mammalian Alkaline Comet Assay. 2016. Guideline 489. pp. 1–27. Available online: https://www.oecd.org/env/test-no-489-in-vivo-mammalian-alkaline-comet-assay-9789264264885-en.htm (accessed on 6 April 2020).
- Zervou, S.K.; Christophoridis, C.; Kaloudis, T.; Triantis, T.M.; Hiskia, A. New SPE-LC-MS/MS method for simultaneous determination of multi-class cyanobacterial and algal toxins. J. Hazard. Mater. 2017, 323, 56–66. [Google Scholar] [CrossRef]
- Pinheiro, C.; Azevedo, J.; Campos, A.; Vasconcelos, V.; Loureiro, S. The interactive effects of microcystin-LR and cylindrospermopsin on the growth rate of the freshwater algae Chlorella vulgaris. Ecotoxicology 2016, 25, 745–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puerto, M.; Prieto, A.I.; Maisanaba, S.; Gutiérrez-Praena, D.; Mellado-García, P.; Jos, Á.; Cameán, A.M. Mutagenic and genotoxic potential of pure Cylindrospermopsin by a battery of in vitro tests. Food Chem. Toxicol. 2018, 121, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Bowen, D.E.; Whitwell, J.H.; Lillford, L.; Henderson, D.; Kidd, D.; McGarry, S.; Pearce, G.; Beevers, C.; Kirkland, D.J. Evaluation of a multi-endpoint assay in rats, combining the bone-marrow micronucleus test, the comet assay and the flow-cytometric peripheral blood micronucleus test. Mutat. Res. Genet. Toxicol. Environ. Mutagen 2011, 722, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Kirkland, D.; Levy, D.D.; LeBaron, M.J.; Aardema, M.J.; Beevers, C.; Bhalli, J.; Douglas, G.R.; Escobar, P.A.; Farabaugh, C.S.; Guerard, M.; et al. A comparison of transgenic rodent mutation and in vivo comet assay responses for 91 chemicals. Mutat. Res. Genet.Toxicol. Environ. Mutagen. 2019, 839, 21–35. [Google Scholar] [CrossRef]
- Azqueta, A.; Shaposhnikov, S.; Collins, A.R. DNA oxidation: Investigating its key role in environmental mutagenesis with the comet assay. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2009, 674, 101–108. [Google Scholar] [CrossRef]
- Llana-Ruiz-Cabello, M.; Maisanaba, S.; Puerto, M.; Prieto, A.I.; Pichardo, S.; Moyano, R.; González-Pérez, J.A.; Cameán, A.M. Genotoxicity evaluation of carvacrol in rats using a combined micronucleus and comet assay. Food Chem. Toxicol. 2016, 98, 240–250. [Google Scholar] [CrossRef]
- Hooser, S.B.; Beasley, V.R.; Lovell, R.A.; Carmichael, W.W.; Haschek, W.M. Toxicity of Microcystin LR, a Cyclic Heptapeptide Hepatotoxin from Microcystis aeruginosa, to Rats and Mice. Vet. Pathol. 1989, 26, 246–252. [Google Scholar] [CrossRef] [Green Version]
- Chernoff, N.; Hill, D.J.; Chorus, I.; Diggs, D.L.; Huang, H.; King, D.; Lang, J.R.; Le, T.T.; Schmid, J.E.; Travlos, G.S.; et al. Cylindrospermopsin toxicity in mice following a 90-d oral exposure. J. Toxicol. Environ. Health A 2018, 81, 549–566. [Google Scholar] [CrossRef]
- Diez-Quijada, L.; Puerto, M.; Gutiérrez-Praena, D.; Llana-Ruiz-Cabello, M.; Jos, Á.; Cameán, A.M. Microcystin-RR: Occurrence, content in water and food and toxicological studies. A review. Environ. Res. 2019, 168, 467–489. [Google Scholar] [CrossRef]
- Humpage, A.R.; Fenech, M.; Thomas, P.; Falconer, I.R. Micronucleus induction and chromosome loss in transformed human white cells indicate clastogenic and aneugenic action of the cyanobacterial toxin, cylindrospermopsin. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2000, 472, 155–161. [Google Scholar] [CrossRef]
- Sieroslawska, A.; Rymuszka, A. Cylindrospermopsin induces oxidative stress and genotoxic effects in the fish CLC cell line. J. Appl. Toxicol. 2015, 35, 426–433. [Google Scholar] [CrossRef]
- Štraser, A.; Žegura, B.; Filipič, M. Genotoxic effects of the cyanobacterial hepatotoxin cylindrospermopsin in the HepG2 cell line. Arch. Toxicol. 2011, 85, 1617–1626. [Google Scholar] [CrossRef]
- Zhan, L.; Sakamoto, H.; Sakuraba, M.; Wu, D.-S.; Zhang, L.-S.; Suzuki, T.; Hayashi, M.; Honma, M. Genotoxicity of microcystin-LR in human lymphoblastoid TK6 cells. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2004, 557, 1–6. [Google Scholar] [CrossRef]
- Štraser, A.; Filipič, M.; Gorenc, I.; Žegura, B. The influence of cylindrospermopsin on oxidative DNA damage and apoptosis induction in HepG2 cells. Chemosphere 2013, 92, 24–30. [Google Scholar] [CrossRef]
- Žegura, B.; Sedmak, B.; Filipič, M. Microcystin-LR induces oxidative DNA damage in human hepatoma cell line HepG2. Toxicon 2003, 41, 41–48. [Google Scholar] [CrossRef]
- Žegura, B.; Gajski, G.; Štraser, A.; Garaj-Vrhovac, V.; Filipič, M. Microcystin-LR induced DNA damage in human peripheral blood lymphocytes. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2011, 726, 116–122. [Google Scholar] [CrossRef]
- Amé, M.V.; Baroni, M.V.; Galanti, L.N.; Bocco, J.L.; Wunderlin, D.A. Effects of microcystin-LR on the expression of P-glycoprotein in Jenynsia multidentata. Chemosphere 2009, 74, 1179–1186. [Google Scholar] [CrossRef]
- Qiu, T.; Xie, P.; Liu, Y.; Li, G.; Xiong, Q.; Hao, L.; Li, H. The profound effects of microcystin on cardiac antioxidant enzymes, mitochondrial function and cardiac toxicity in rat. Toxicology 2009, 257, 86–94. [Google Scholar] [CrossRef]
- Arman, T.; Lynch, K.D.; Montonye, M.L.; Goedken, M.; Clarke, J.D. Sub-Chronic Microcystin-LR Liver Toxicity in Preexisting Diet-Induced Nonalcoholic Steatohepatitis in Rats. Toxins 2019, 11, 398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, T.; Makita, Y.; Nagata, S.; Tsutsumi, T.; Yoshida, F.; Sekijima, M.; Tamura, S.I.; Ueno, Y. Acute oral toxicity of microcystin-LR, a cyanobacterial hepatotoxin, in mice. Nat Toxins 1997, 5, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Sedan, D.; Andrinolo, D.; Telese, L.; Giannuzzi, L.; de Alaniz, M.J.; Marra, C.A. Alteration and recovery of the antioxidant system induced by sub-chronic exposure to microcystin-LR in mice: Its relation to liver lipid composition. Toxicon 2010, 55, 333–342. [Google Scholar] [CrossRef]
- Zhang, X.X.; Zhang, Z.; Fu, Z.; Wang, T.; Qin, W.; Xu, L.; Cheng, S.; Yang, L. Stimulation effect of microcystin-LR on matrix metalloproteinase-2/-9 expression in mouse liver. Toxicol. Lett. 2010, 199, 377–382. [Google Scholar] [CrossRef]
- Sun, X.; Mi, L.; Liu, J.; Song, L.; Chung, F.L.; Gan, N. Sulforaphane prevents microcystin-LR-induced oxidative damage and apoptosis in BALB/c mice. Toxicol. Appl. Pharmacol. 2011, 255, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Weng, D.; Lu, Y.; Wei, Y.; Liu, Y.; Shen, P. The role of ROS in microcystin-LR-induced hepatocyte apoptosis and liver injury in mice. Toxicology 2007, 232, 15–23. [Google Scholar] [CrossRef]
- Han, Z.X.; Yang, L.; Zhang, L.; Xu, C.; Shu, W.Q. The antagonistic action of epigallocatechin-3-gallate on microcystin LR-induced oxidative damage on hepatocytes of mice and the expression of cytochrome P450 2E1. Zhonghua Yu Fang Yi Xue Za Zhi 2010, 44, 24–29. [Google Scholar] [CrossRef]
- Sibaldo de Almeida, C.; Costa de Arruda, A.C.; Caldas de Queiroz, E.; Matias de Lima Costa, H.T.; Fernandes Barbosa, P.; Araújo Moura Lemos, T.M.; Nunes Oliveira, C.; Pinto, E.; Schwarz, A.; Kujbida, P. Oral exposure to cylindrospermopsin in pregnant rats: Reproduction and foetal toxicity studies. Toxicon 2013, 74, 127–129. [Google Scholar] [CrossRef]
- Humpage, A.R.; Falconer, I.R. Oral toxicity of the cyanobacterial toxin cylindrospermopsin in male Swiss albino mice: Determination of no observed adverse effect level for deriving a drinking water guideline value. Environ. Toxicol. 2003, 18, 94–103. [Google Scholar] [CrossRef]
- Chernoff, N.; Rogers, E.H.; Zehr, R.D.; Gage, M.I.; Malarkey, D.E.; Bradfield, C.A.; Liu, Y.; Schmid, J.E.; Jaskot, R.H.; Richards, J.H.; et al. Toxicity and recovery in the pregnant mouse after gestational exposure to the cyanobacterial toxin, cylindrospermopsin. J. Appl. Toxicol. 2010, 31, 242–254. [Google Scholar] [CrossRef] [Green Version]
- Seawright, A.A.; Nolan, C.C.; Shaw, G.R.; Chiswell, R.K.; Norris, R.L.; Moore, M.R.; Smith, M.J. The oral toxicity for mice of the tropical cyanobacterium Cylindrospermopsis raciborskii (Wolonszynska). Environ. Toxicol. 1999, 14, 135–142. [Google Scholar] [CrossRef]
- Pouria, S.; de Andrade, A.; Barbosa, J.; Cavalcanti, R.L.; Barreto, V.T.; Ward, C.J.; Preiser, W.; Poon, G.K.; Neild, G.H.; Codd, G.A. Fatal microcystin intoxication in haemodialysis unit in Caruaru, Brazil. Lancet 1998, 352, 21–26. [Google Scholar] [CrossRef]
- Frangež, R.; Kosec, M.; Sedmak, B.; Beravs, K.; Demsar, F.; Juntes, P.; Pogačnik, M.; Šuput, D. Subchronic liver injuries caused by microcystins. Pflügers Arch. 2000, 440, 103–104. [Google Scholar] [CrossRef]
- Ito, E.; Kondo, F.; Harada, K.I. Hepatic necrosis in aged mice by oral administration of microcystin-LR. Toxicon 1997, 35, 231–239. [Google Scholar] [CrossRef]
- Iwabuchi, T.; Iijima, K.; Ara, N.; Koike, T.; Shinkai, H.; Ichikawa, T.; Kamata, Y.; Ishihara, K.; Shimosegawa, T. Increased gastric mucus secretion alleviates non-steroidal anti-inflammatory drug-induced abdominal pain. Tohoku J. Exp. Med. 2013, 231, 29–36. [Google Scholar] [CrossRef] [Green Version]
- De La Cruz, A.A.; Hiskia, A.; Kaloudis, T.; Chernoff, N.; Hill, D.; Antoniou, M.G.; He, X.; Loftin, K.; O’Shea, K.; Zhao, C.; et al. A review on cylindrospermopsin: The global occurrence, detection, toxicity and degradation of a potent cyanotoxin. Environ. Sci. Process. Impacts 2013, 15, 1979–2003. [Google Scholar] [CrossRef]
- International Conference of Harmonisation (ICH). Guidance on Genotoxicity Testing and Data Interpretation for Pharmaceuticals Intended for Human Use S2 (R1). 2012. Available online: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/s2r1-genotoxicity-testing-and-data-interpretation-pharmaceuticals-intended-human-use (accessed on 6 April 2020).
- Easterbrook, J.; Lu, C.; Sakai, Y.; Li, A.P. Effects of organic solvents on the activities of cytochrome P450 isoforms, UDP-dependent glucuronyl transferase, and phenol sulfotransferase in human hepatocytes. Drug Metab. Dispos. 2001, 29, 141–144. [Google Scholar]
- Corcuera, L.A.; Vettorazzi, A.; Arbillaga, L.; Pérez, N.; Gil, A.G.; Azqueta, A.; González-Peñas, E.; García-Jalón, J.A.; López de Cerain, A. Genotoxicity of Aflatoxin B1 and Ochratoxin A after simultaneous application of the in vivo micronucleus and comet assay. Food Chem. Toxicol. 2015, 76, 116–124. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Sex | n | Doses | PCE/Total | % MN-PCE´s | PCE/NCE |
|---|---|---|---|---|---|---|
| Negative Control (water) | ♂ | 5 | 0.49 ± 0.02 | 0.45 ± 0.64 | 0.95 ± 0.08 | |
| ♀ | 5 | 0.49 ± 0.03 | 0.66 ± 0.32 | 0.96 ± 0.14 | ||
| Solvent Control (0.5% MeOH) | ♂ | 5 | 0.50 ± 0.02 | 0.83 ± 0.51 | 0.98 ± 0.08 | |
| ♀ | 5 | 0.50 ± 0.03 | 0.58 ± 0.24 | 1.00 ± 0.11 | ||
| Positive Control (EMS *) | ♂ | 3 | 200 mg/kg b.w. | 0.34 ± 0.02 ** | 1.93 ± 0.15 ** | 0.51 ± 0.05 ** |
| ♀ | 3 | 0.36 ± 0.03 ** | 2.47 ± 0.9 ** | 0.57 ± 0.08 ** | ||
| CYN/MC-LR | ♂ | 5 | 7.5 + 75 µg/kg b.w. | 0.51 ± 0.02 | 1.75 ± 0.42 ** | 1.03 ± 0.07 |
| ♀ | 5 | 0.50 ± 0.03 | 1.98 ± 0.21 ** | 1.05 ± 0.13 | ||
| ♂ | 5 | 23.7 + 237 µg/kg b.w. | 0.46 ± 0.04 | 1.83 ± 0.41 ** | 0.86 ± 0.14 | |
| ♀ | 5 | 0.47 ± 0.03 | 2.09 ± 0.15 ** | 0.88 ± 0.09 | ||
| ♂ | 5 | 75 + 750 µg/kg b.w. | 0.37 ± 0.03 ** | 1.88 ± 0.51 ** | 0.60 ± 0.08 ** | |
| ♀ | 5 | 0.34 ± 0.06 ** | 2.22 ± 0.27 ** | 0.51 ± 0.12 ** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Díez-Quijada, L.; Medrano-Padial, C.; Llana-Ruiz-Cabello, M.; Cătunescu, G.M.; Moyano, R.; Risalde, M.A.; Cameán, A.M.; Jos, Á. Cylindrospermopsin-Microcystin-LR Combinations May Induce Genotoxic and Histopathological Damage in Rats. Toxins 2020, 12, 348. https://doi.org/10.3390/toxins12060348
Díez-Quijada L, Medrano-Padial C, Llana-Ruiz-Cabello M, Cătunescu GM, Moyano R, Risalde MA, Cameán AM, Jos Á. Cylindrospermopsin-Microcystin-LR Combinations May Induce Genotoxic and Histopathological Damage in Rats. Toxins. 2020; 12(6):348. https://doi.org/10.3390/toxins12060348
Chicago/Turabian StyleDíez-Quijada, Leticia, Concepción Medrano-Padial, María Llana-Ruiz-Cabello, Giorgiana M. Cătunescu, Rosario Moyano, Maria A. Risalde, Ana M. Cameán, and Ángeles Jos. 2020. "Cylindrospermopsin-Microcystin-LR Combinations May Induce Genotoxic and Histopathological Damage in Rats" Toxins 12, no. 6: 348. https://doi.org/10.3390/toxins12060348
APA StyleDíez-Quijada, L., Medrano-Padial, C., Llana-Ruiz-Cabello, M., Cătunescu, G. M., Moyano, R., Risalde, M. A., Cameán, A. M., & Jos, Á. (2020). Cylindrospermopsin-Microcystin-LR Combinations May Induce Genotoxic and Histopathological Damage in Rats. Toxins, 12(6), 348. https://doi.org/10.3390/toxins12060348