In Vitro Toxicological Screening of Stable and Senescing Cultures of Aphanizomenon, Planktothrix, and Raphidiopsis

 ,
, 
 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
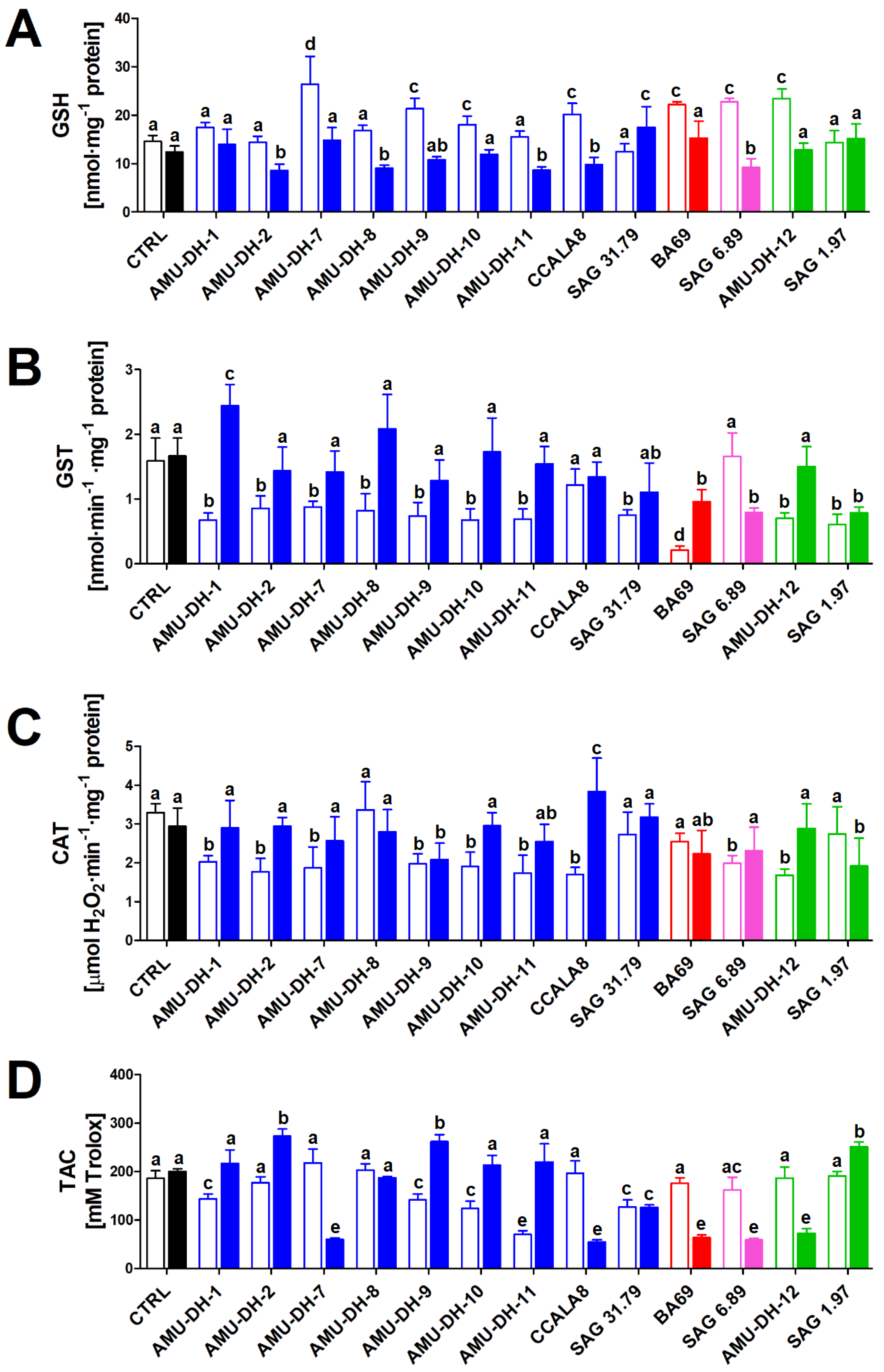
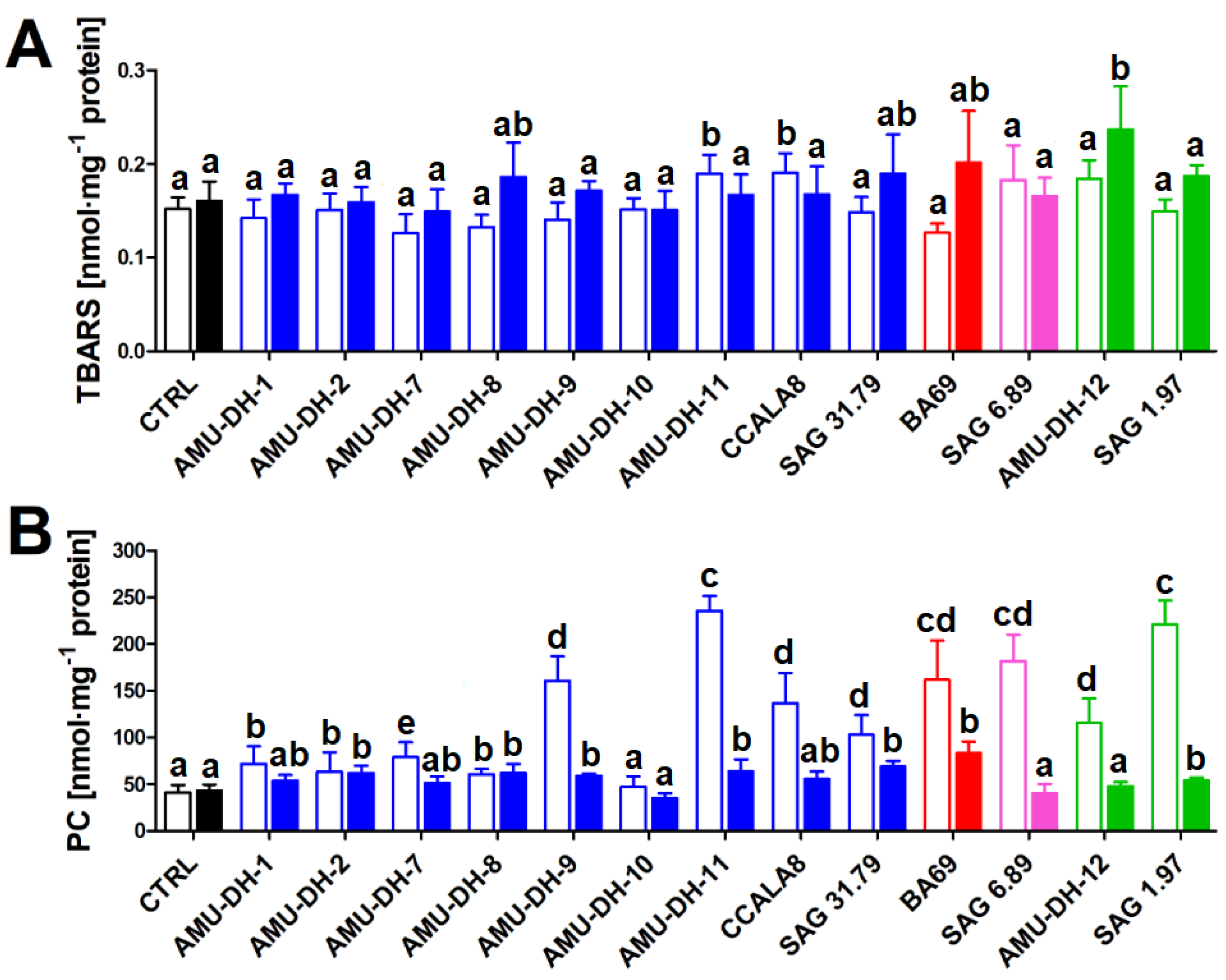
2.1. Oxidative Stress
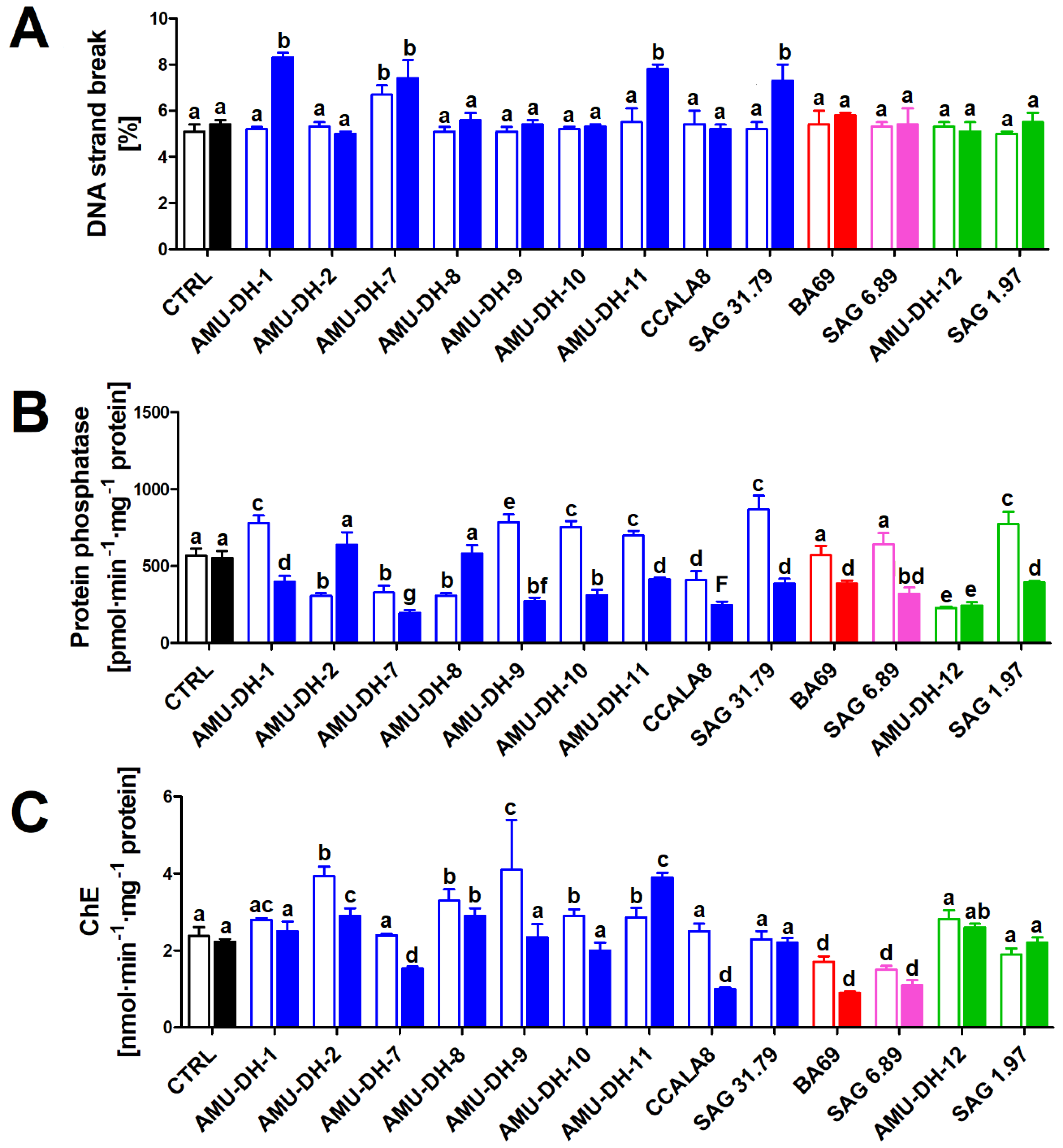
2.2. Genotoxicity
2.3. Serine/Threonine Protein Phosphatase Activity
2.4. Neurotoxicity
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Cyanobacterial Strains
5.2. High Performance Liquid Chromatography Analyses
5.3. Extract Preparation
5.4. Experimental Design
5.4.1. Oxidative Stress
5.4.2. DNA Fragmentation
5.4.3. Serine/Threonine Protein Phosphatase Activity
5.4.4. Cholinesterase Activity
5.5. Statistical Analyses
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Braun, A.; Pfeiffer, T. Cyanobacterial blooms as the cause of a Pleistocene large mammal assemblage. Paleobiology 2002, 28, 139–154. [Google Scholar] [CrossRef]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.H.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef]
- Henao, E.; Rzymski, P.; Waters, M.N. A Review on the Study of Cyanotoxins in Paleolimnological Research: Current Knowledge and Future Needs. Toxins Basel 2020, 12, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jöhnk, K.D.; Huisman, J.; Sharples, J.; Sommeijer, B.; Visser, P.M.; Stroom, J.M. Summer heatwaves promote blooms of harmful cyanobacteria. Glob. Chang. Biol. 2008, 14, 495–512. [Google Scholar] [CrossRef] [Green Version]
- Wojtal-Frankiewicz, A.; Kruk, A.; Frankiewicz, P.; Oleksińska, Z.; Izydorczyk, K. Long-term patterns in the population dynamics of Daphnia longispina, Leptodora kindtii and cyanobacteria in a shallow reservoir: A self-organising map (SOM) approach. PLoS ONE 2015, 10, e0144109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Neil, J.M.; Davis, T.W.; Burford, M.A.; Gobler, C.J. The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change. Harmful Algae 2012, 14, 313–334. [Google Scholar] [CrossRef]
- Lürling, M.; van Oosterhout, F.; Faassen, E. Eutrophication and warming boost cyanobacterial biomass and microcystins. Toxins Basel 2017, 9, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wells, M.L.; Karlson, B.; Wulff, A.; Kudela, R.; Trick, C.; Asnaghi, V.; Berdalet, E.; Cochlan, W.; Davidson, K.; De Rijcke, M.; et al. Future HAB science: Directions and challenges in a changing climate. Harmful Algae 2020, 91, 101632. [Google Scholar] [CrossRef]
- Toporowska, M.; Pawlik-Skowrońska, B.; Krupa, D.; Kornijów, R. Winter versus summer blooming of phytoplankton in a shallow lake: Effect of hypertrophic conditions. Pol. J. Ecol. 2010, 58, 3–12. [Google Scholar]
- Babanazarova, O.; Sidelev, S.; Schischeleva, S. The structure of winter phytoplankton in Lake Nero, Russia, a hypertrophic lake dominated by Planktothrix-like cyanobacteria. Aquat. Biosyst. 2013, 9, 18. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Qin, B.; Paerl, H.W.; Brookes, J.D.; Hall, N.S.; Shi, K.; Zhou, Y.; Guo, J.; Li, Z.; Xu, H.; et al. The persistence of cyanobacterial (Microcystis spp.) blooms throughout winter in Lake Taihu, China. Limnol. Oceanogr. 2016, 61, 711–722. [Google Scholar] [CrossRef] [Green Version]
- Wejnerowski, Ł.; Rzymski, P.; Kokociński, M.; Meriluoto, J. The structure and toxicity of winter cyanobacterial bloom in a eutrophic lake of the temperate zone. Ecotoxicology 2018, 27, 752–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coffer, M.M.; Schaeffer, B.A.; Darling, J.A.; Urquhart, E.A.; Salls, W.B. Quantifying national and regional cyanobacterial occurrence in US lakes using satellite remote sensing. Ecol. Indic. 2020, 111. [Google Scholar] [CrossRef]
- Von Elert, E.; Martin-Creuzburg, D.; Le Coz, J.R. Absence of sterols constrains carbon transfer between cyanobacteria and a freshwater herbivore (Daphnia galeata). Proc. Biol. Sci. 2003, 270, 1209–1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gliwicz, Z.M. Why do cladocerans fail to control algal blooms? Hydrobiologia 1990, 200–201, 83–97. [Google Scholar] [CrossRef]
- Wilk-Wozniak, E. An introduction to the “micronet” of cyanobacterial harmful algal blooms (CyanoHABs): Cyanobacteria, zooplankton and microorganisms: A review. Mar. Freshw. Res. 2019. [Google Scholar] [CrossRef]
- Hartmann, H.J.; Kunkel, D.D. Mechanisms of food selection in Daphnia. Hydrobiologia 1991, 225, 129–154. [Google Scholar] [CrossRef]
- Poniedziałek, B.; Rzymski, P.; Kokociński, M. Cylindrospermopsin: Water-linked potential threat to human health in Europe. Environ. Toxicol. Pharmacol. 2012, 34, 651–660. [Google Scholar] [CrossRef]
- Codd, G.A.; Meriluoto, J.; Metcalf, J.S. Introduction. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 1–8. [Google Scholar]
- Kaloudis, T.; Triantis, T.M.; Hiskia, A. Taste and odour compounds produced by cyanobacteria. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; John Wiley & Sons, Ltd.: Chicester, UK, 2017; pp. 196–201. [Google Scholar]
- Pavagadhi, S.; Balasubramanian, R. Toxicological evaluation of microcystins in aquatic fish species: Current knowledge and future directions. Aquat. Toxicol. 2013, 142–143, 1–16. [Google Scholar] [CrossRef]
- Baumann, H.I.; Jüttner, F. Inter-annual stability of oligopeptide patterns of Planktothrix rubescens blooms and mass mortality of Daphnia in Lake Hallwilersee. Limnologica 2008, 38, 350–359. [Google Scholar] [CrossRef] [Green Version]
- Wood, R. Acute animal and human poisonings from cyanotoxin exposure—A review of the literature. Environ. Int. 2016, 91, 276–282. [Google Scholar] [CrossRef]
- Poniedziałek, B.; Rzymski, P.; Kokociński, M.; Karczewski, J. Toxic potencies of metabolite(s) of non-cylindrospermopsin producing Cylindrospermopsis raciborskii isolated from temperate zone in human white cells. Chemosphere 2015, 120, 608–614. [Google Scholar] [CrossRef]
- Falfushynska, H.; Horyn, O.; Brzozowska, A.; Fedoruk, O.; Buyak, B.; Poznansky, D.; Poniedziałek, B.; Kokociński, M.; Rzymski, P. Is the presence of Central European strains of Raphidiopsis (Cylindrospermopsis) raciborskii a threat to a freshwater fish? An in vitro toxicological study in common carp cells. Aquat. Toxicol. 2019, 206, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Chislock, M.F.; Sarnelle, O.; Jernigan, L.M.; Wilson, A.E. Do high concentrations of microcystin prevent Daphnia control of phytoplankton? Water Res. 2013, 47, 1961–1970. [Google Scholar] [CrossRef] [PubMed]
- Chislock, M.F.; Sarnelle, O.; Olsen, B.K.; Doster, E.; Wilson, A.E. Large effects of consumer offense on ecosystem structure and function. Ecology 2013, 94, 2375–2380. [Google Scholar] [CrossRef]
- Mohamed, Z.A.; Bakr, A.A.; Ghramh, H.A. Grazing of the copepod Cyclops vicinus on toxic Microcystis aeruginosa: Potential for controlling cyanobacterial blooms and transfer of toxins. Oceanol. Hydrobiol. Stud. 2018, 47, 296–302. [Google Scholar] [CrossRef]
- Manganelli, M.; Stefanelli, M.; Vichi, S.; Andreani, P.; Nascetti, G.; Scialanca, F.; Scardala, S.; Testai, E.; Funari, E. Cyanobacteria biennal dynamic in a volcanic mesotrophic lake in central Italy: Strategies to prevent dangerous human exposures to cyanotoxins. Toxicon 2016, 115, 28–40. [Google Scholar] [CrossRef] [PubMed]
- Yunes, J.S.; De La Rocha, S.; Giroldo, D.; da Silveira, S.B.; Comin, R.; da Silva Bicho, M.; Melcher, S.S.; Sant’anna, C.L.; Vieira, A.A.H. Release of carbohydrates and proteins by a subtropical strain of Raphidiopsis brookii (cyanobacteria) able to produce saxitoxin at three nitrate concentrations. J. Phycol. 2009, 45, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Dias, E.; Pereira, P.; Franca, S. Production of paralytic shellfish toxins by Aphanizomenon sp LMECYA 31 (cyanobacteria). J. Phycol. 2002, 38, 705–712. [Google Scholar] [CrossRef]
- Carneiro, R.L.; Elisângela Venâncio, M.; Pacheco Furlanetto, A.B.; Feliciano de Oliveira e Azevedo, S.M. Effects of light intensity and light quality on growth and circadian rhythm of saxitoxins production in Cylindrospermopsis raciborskii (Cyanobacteria). J. Plankton Res. 2009, 31, 481–488. [Google Scholar] [CrossRef]
- Svirčev, Z.; Obradović, V.; Codd, G.A.; Marjanović, P.; Spoof, L.; Drobac, D.; Tokodi, N.; Petković, A.; Nenin, T.; Simeunović, J.; et al. Massive fish mortality and Cylindrospermopsis raciborskii bloom in Aleksandrovac Lake. Ecotoxicology 2016, 25, 1353–1363. [Google Scholar] [CrossRef] [PubMed]
- Rzymski, P.; Poniedziałek, B.; Kokociński, M.; Jurczak, T.; Lipski, D.; Wiktorowicz, K. Interspecific allelopathy in cyanobacteria: Cylindrospermopsin and Cylindrospermopsis raciborskii effect on the growth and metabolism of Microcystis aeruginosa. Harmful Algae 2014, 35, 1–8. [Google Scholar] [CrossRef]
- Dobronoki, D.; B-Béres, V.; Vasas, G.; Gonda, S.; Nagy, S.A.; Bácsi, I. Potential role of the cellular matrix of Aphanizomenon strains in the effects of cylindrospermopsin—An experimental study. J. Appl. Phycol. 2019, 31, 1805–1817. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.; Rzymski, P. Programmed cell death-like and accompanying release of microcystin in freshwater bloom-forming cyanobacterium microcystis: From identification to ecological relevance. Toxins Basel 2019, 11, 706. [Google Scholar] [CrossRef] [Green Version]
- Sivonen, K. Cyanobacterial toxins and toxin production. Phycologia 1996, 35, 12–24. [Google Scholar] [CrossRef]
- Merel, S.; Walker, D.; Chicana, R.; Snyder, S.; Baurès, E.; Thomas, O. State of knowledge and concerns on cyanobacterial blooms and cyanotoxins. Environ. Int. 2013, 59, 303–327. [Google Scholar] [CrossRef] [PubMed]
- Harland, F.M.J.; Wood, S.A.; Moltchanova, E.; Williamson, W.M.; Gaw, S. Phormidium autumnale growth and anatoxin-a production under iron and copper stress. Toxins Basel 2013, 5, 2504–2521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, Y.; Yu, Z.; Chen, H.; Yang, J.R.; Liu, M.; Liu, L.; Huang, B.; Yang, J. Cyanobacterial bloom significantly boosts hypolimnelic anammox bacterial abundance in a subtropical stratified reservoir. FEMS Microbiol. Ecol. 2017, 93, 118. [Google Scholar] [CrossRef] [Green Version]
- Abed, R.M.M. Interaction between cyanobacteria and aerobic heterotrophic bacteria in the degradation of hydrocarbons. Int. Biodeterior. Biodegrad. 2010, 64, 58–64. [Google Scholar] [CrossRef]
- Briand, E.; Humbert, J.-F.; Tambosco, K.; Bormans, M.; Gerwick, W.H. Role of bacteria in the production and degradation of Microcystis cyanopeptides. MicrobiologyOpen 2016, 5, 469–478. [Google Scholar] [CrossRef]
- Komárek, J.; Komárková, J. Taxonomic review of the cyanoprokaryotic genera Planktothrix and Planktothricoides. Czech Phycol. 2004, 4, 1–18. [Google Scholar]
- Pimentel, J.S.M.; Giani, A. Microcystin production and regulation under nutrient stress conditions in toxic Microcystis strains. Appl. Environ. Microbiol. 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robarts, R.D.; Waiser, M.J.; Arts, M.T.; Evans, M.S. Seasonal and diel changes of dissolved oxygen in a hypertrophic prairie lake. Lakes Reserv. Res. Manag. 2005, 10, 167–177. [Google Scholar] [CrossRef]
- Wejnerowski, L.; Cerbin, S.; Wojciechowicz, M.; Jurczak, T.; Glama, M.; Meriluoto, J.; Dziuba, M. Effects of Daphnia exudates and sodium octyl sulphates on filament morphology and cell wall thickness of Aphanizomenon gracile (Nostocales), Cylindrospermopsis raciborskii (Nostocales) and Planktothrix agardhii (Oscillatoriales). Eur. J. Phycol. 2018, 53. [Google Scholar] [CrossRef]
- Poniedziałek, B.; Rzymski, P.; Karczewski, J. The role of the enzymatic antioxidant system in cylindrospermopsin-induced toxicity in human lymphocytes. Toxicol. Vitr. 2015, 29, 926–932. [Google Scholar] [CrossRef]
- Buratti, F.M.; Manganelli, M.; Vichi, S.; Stefanelli, M.; Scardala, S.; Testai, E.; Funari, E. Cyanotoxins: Producing organisms, occurrence, toxicity, mechanism of action and human health toxicological risk evaluation. Arch. Toxicol. 2017, 91, 1049–1130. [Google Scholar] [CrossRef]
- Massey, I.Y.; Yang, F.; Ding, Z.; Yang, S.; Guo, J.; Tezi, C.; Al-Osman, M.; Kamegni, R.B.; Zeng, W. Exposure routes and health effects of microcystins on animals and humans: A mini-review. Toxicon 2018, 151, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Henao, E.; Murphy, P.J.; Falfushynska, H.; Horyn, O.; Evans, D.M.; Klimaszyk, P.; Rzymski, P. Polymethoxy-1-alkenes screening of Chlorella and Spirulina food supplements coupled with in vivo toxicity studies. Toxins Basel 2020, 12, 111. [Google Scholar] [CrossRef] [Green Version]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014. [Google Scholar] [CrossRef]
- Fedorova, M.; Bollineni, R.C.; Hoffmann, R. Protein carbonylation as a major hallmark of oxidative damage: Update of analytical strategies. Mass Spectrom. Rev. 2014, 33, 79–97. [Google Scholar] [CrossRef]
- Burford, M.A.; Willis, A.; Chuang, A.; Man, X.; Orr, P.T. Recent insights into physiological responses to nutrients by the cylindrospermopsin producing cyanobacterium, Cylindrospermopsis raciborskii. J. Oceanol. Limnol. 2018, 36, 1032–1039. [Google Scholar] [CrossRef] [Green Version]
- Xiao, M.; Willis, A.; Burford, M.A. Differences in cyanobacterial strain responses to light and temperature reflect species plasticity. Harmful Algae 2017, 62, 84–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willis, A.; Chuang, A.W.; Woodhouse, J.N.; Neilan, B.A.; Burford, M.A. Intraspecific variation in growth, morphology and toxin quotas for the cyanobacterium, Cylindrospermopsis raciborskii. Toxicon 2016, 119, 307–310. [Google Scholar] [CrossRef] [PubMed]
- Johansson, E.; Legrand, C.; Björnerås, C.; Godhe, A.; Mazur-Marzec, H.; Säll, T.; Rengefors, K. High diversity of microcystin chemotypes within a summer bloom of the cyanobacterium Microcystis botrys. Toxins Basel 2019, 11, 698. [Google Scholar] [CrossRef] [Green Version]
- Takenaka, S.; Watanabe, M.F. Microcystin LR degradation by Pseudomonas aeruginosa alkaline protease. Chemosphere 1997, 34, 749–757. [Google Scholar] [CrossRef]
- Kato, H.; Imanishi, S.Y.; Tsuji, K.; Harada, K. Microbial degradation of cyanobacterial cyclic peptides. Water Res. 2007, 41, 1754–1762. [Google Scholar] [CrossRef]
- Gumbo, R.; Ross, G.; Cloete, E. Biological control of Microcystis dominated harmful algal blooms. Afr. J. Biotechnol. 2008, 7. [Google Scholar] [CrossRef]
- Mazur-Marzec, H.; Toruńska, A.; Błońska, M.J.; Moskot, M.; Pliński, M.; Jakóbkiewicz-Banecka, J.; Wegrzyn, G. Biodegradation of nodularin and effects of the toxin on bacterial isolates from the Gulf of Gdańsk. Water Res. 2009, 43, 2801–2810. [Google Scholar] [CrossRef]
- Kormas, K.A.; Lymperopoulou, D.S. Cyanobacterial toxin degrading bacteria: Who are they? Biomed. Res. Int. 2013, 2013, 12. [Google Scholar] [CrossRef] [Green Version]
- Wejnerowski, L.; Cerbin, S.; Dziuba, M.K. Thicker filaments of Aphanizomenon gracile are more harmful to Daphnia than thinner Cylindrospermopsis raciborskii. Zool. Stud. 2015, 54. [Google Scholar] [CrossRef] [Green Version]
- Ballot, A.; Fastner, J.; Wiedner, C. Paralytic shellfish poisoning toxin-producing cyanobacterium Aphanizomenon gracile in northeast Germany. Appl. Environ. Microbiol. 2010, 76, 1173–1180. [Google Scholar] [CrossRef] [Green Version]
- Carneiro, R.L.; Pacheco, A.B.F.; De Oliveira, E.; Azevedo, S.M.F. Growth and saxitoxin production by Cylindrospermopsis raciborskii (cyanobacteria) correlate with water hardness. Mar. Drugs 2013, 11, 2949–2963. [Google Scholar] [CrossRef] [Green Version]
- Casero, M.C.; Ballot, A.; Agha, R.; Quesada, A.; Cirés, S. Characterization of saxitoxin production and release and phylogeny of sxt genes in paralytic shellfish poisoning toxin-producing Aphanizomenon gracile. Harmful Algae 2014, 37, 28–37. [Google Scholar] [CrossRef]
- Rzymski, P.; Poniedziałek, B.; Mankiewicz-Boczek, J.; Faassen, E.J.; Jurczak, T.; Gągała-Borowska, I.; Ballot, A.; Lürling, M.; Kokociński, M. Polyphasic toxicological screening of Cylindrospermopsis raciborskii and Aphanizomenon gracile isolated in Poland. Algal Res. 2017, 24, 72–80. [Google Scholar] [CrossRef]
- Soreq, H.; Seidman, S. Acetylcholinesterase—New roles for an old actor. Nat. Rev. Neurosci. 2001, 2, 294–302. [Google Scholar] [CrossRef]
- Harland, F.; Wood, S.A.; Broady, P.; Williamson, W.; Gaw, S. Changes in saxitoxin-production through growth phases in the metaphytic cyanobacterium Scytonema cf. crispum. Toxicon 2015, 103, 74–79. [Google Scholar] [CrossRef]
- Da Silva, C.A.; Oba, E.T.; Ramsdorf, W.A.; Magalhães, V.F.; Cestari, M.M.; Oliveira Ribeiro, C.A.; Silva de Assis, H.C. First report about saxitoxins in freshwater fish Hoplias malabaricus through trophic exposure. Toxicon 2011, 57, 141–147. [Google Scholar] [CrossRef]
- Viaggiu, E.; Melchiorre, S.; Volpi, F.; Di Corcia, A.; Mancini, R.; Garibaldi, L.; Crichigno, G.; Bruno, M. Anatoxin-a toxin in the cyanobacterium Planktothrix rubescens from a fishing pond in northern Italy. Environ. Toxicol. 2004, 19, 191–197. [Google Scholar] [CrossRef]
- Hodoki, Y.; Ohbayashi, K.; Kobayashi, Y.; Takasu, H.; Okuda, N.; Nakano, S. Anatoxin-a-producing Raphidiopsis mediterranea Skuja var. grandis Hill is one ecotype of non-heterocytous Cuspidothrix issatschenkoi (Usačev) Rajaniemi et al. in Japanese lakes. Harmful Algae 2013, 21–22, 44–53. [Google Scholar] [CrossRef]
- John, N.; Baker, L.; Ansell, B.R.E.; Newham, S.; Crosbie, N.D.; Jex, A.R. First report of anatoxin-a producing cyanobacteria in Australia illustrates need to regularly up-date monitoring strategies in a shifting global distribution. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Vehovszky, A.; Kovács, A.W.; Szabó, H.; Gyori, J.; Farkas, A. Neurotoxic effects evoked by cyanobacterial extracts suggest multiple receptors involved in electrophysiological responses of molluscan (CNS, heart) models. Acta Biol. Hung. 2012, 63, 160–170. [Google Scholar] [CrossRef]
- Vehovszky, Á.; Kovács, A.W.; Farkas, A.; Győri, J.; Szabó, H.; Vasas, G. Pharmacological studies confirm neurotoxic metabolite(s) produced by the bloom-forming Cylindrospermopsis raciborskii in Hungary. Environ. Toxicol. 2015, 30, 501–512. [Google Scholar] [CrossRef] [Green Version]
- Zapomělová, E.; Řeháková, K.; Znachor, P.; Komárková, J. Morphological diversity of coiled planktonic types of the genus Anabaena (cyanobacteria) in natural populations—Taxonomic consequences. Cryptogam. Algol. 2007, 28, 353–371. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota: Bd. 2/Part 2: Oscillatoriales. In Süßwasserflora von Mitteleuropa, Bd. 19/2: Cyanoprokaryota; Büdel, B., Krienitz, L., Gärtner, G., Schagerl, L., Eds.; Springer Spektrum: Berlin/Heildberg, Germany, 2007; p. 759. ISBN 978-3-8274-1914-9. [Google Scholar]
- Komárek, J. Cyanoprokaryota: 3. Teil/3rd Part: Heterocytous genera. In Süßwasserflora von Mitteleuropa, Bd. 19/3: Cyanoprokaryota; Büdel, B., Gärtner, G., Krienitz, L., Schagerl, L., Eds.; Springer Spektrum: Berlin/Heildeberg, Germany, 2013; ISBN 978-3-8274-0932-4. [Google Scholar]
- Guillard, R.R.L.; Lorenzen, C.J. Yellow-green algae with chlorophyllide c. J. Phycol. 1972, 8, 10–14. [Google Scholar] [CrossRef]
- Kokociński, M.; Mankiewicz-Boczek, J.; Jurczak, T.; Spoof, L.; Meriluoto, J.; Rejmonczyk, E.; Hautala, H.; Vehniäinen, M.; Pawełczyk, J.; Soininen, J. Aphanizomenon gracile (Nostocales), a cylindrospermopsin-producing cyanobacterium in Polish lakes. Environ. Sci. Pollut. Res. Int. 2013, 20, 5243–5264. [Google Scholar] [CrossRef] [Green Version]
- Hautala, H.; Lamminmäki, U.; Spoof, L.; Nybom, S.; Meriluoto, J.; Vehniäinen, M. Quantitative PCR detection and improved sample preparation of microcystin-producing Anabaena, Microcystis and Planktothrix. Ecotoxicol. Environ. Saf. 2013, 87, 49–56. [Google Scholar] [CrossRef]
- Wetzel, R.G.; Likens, G.E. Limnological Analyses; Springer: New York, NY, USA, 2000. [Google Scholar]
- Evans, D.M.; Hughes, J.; Jones, L.F.; Murphy, P.J.; Falfushynska, H.; Horyn, O.; Sokolova, I.M.; Christensen, J.; Coles, S.J.; Rzymski, P. Elucidating cylindrospermopsin toxicity via synthetic analogues: An in vitro approach. Chemosphere 2019, 234, 139–147. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Anderson, M.E. Determination of glutathione and glutathione disulfide in biological samples. Methods Enzymol. 1985, 113, 548–555. [Google Scholar] [CrossRef]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S transferases. The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar]
- Aebi, H.; Wyss, S.R.; Scherz, B.; Skvaril, F. Heterogeneity of erythrocyte catalase II: Isolation and characterization of normal and variant erythrocyte catalase and their subunits. Eur. J. Biochem. 1974, 48, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Poniedziałek, B.; Rzymski, P.; Pięt, M.; Gąsecka, M.; Stroińska, A.; Niedzielski, P.; Mleczek, M.; Rzymski, P.; Wilczak, M. Relation between polyphenols, malondialdehyde, antioxidant capacity, lactate dehydrogenase and toxic elements in human colostrum milk. Chemosphere 2018, 191, 548–554. [Google Scholar] [CrossRef] [PubMed]
- Reznick, A.Z.; Packer, L. Oxidative damage to proteins: Spectrophotometric method for carbonyl assay. Methods Enzymol. 1994, 233, 357–363. [Google Scholar] [CrossRef] [PubMed]
- McAvoy, T.; Nairn, A.C. Serine/threonine protein phosphatase assays. Curr. Protoc. Mol. Biol. 2010. [Google Scholar] [CrossRef] [Green Version]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species, Strain | CYN | MC-RR | MC-YR | MC-LR | Source of Information | Origin | Optical Density (750 nm) in Cultures (Chlorophyll Concentration in µg L−1) | ||
|---|---|---|---|---|---|---|---|---|---|
| Day 0 | Day 25 | Day 70 | |||||||
| A. gracile | |||||||||
| AMU-DH-1 | – | – | – | – | this study | Lake Buszewskie (52°32′42″ N, 16°22′47″ E) | 0.008 ± 0.001 | 0.819 ± 0.095 (2376) | 0.774 ± 0.136 (3729) |
| AMU-DH-2 | – | – | – | – | [46] | Lake Lubosińskie (52°31′40″N, 16°22′56″E) | 0.006 ± 0.001 | 0.712 ± 0.078 (3814) | 0.682 ± 0.232 (4599) |
| AMU-DH-7 | – | n.e. | n.e. | n.e. | this study | Lake Pniewskie (52°30′41″N, 16°14′27″) | 0.057 ± 0.006 | 0.654 ± 0.12 (2477) | 0.689 ± 0.294 (5731) |
| AMU-DH-8 | – | n.e. | n.e. | n.e. | this study | Lake Pniewskie (52°30′41″N, 16°14′27″) | 0.008 ± 0.002 | 0.619 ± 0.057 (1733) | 0.755 ± 0.129 (2161) |
| AMU-DH-9 | – | n.e. | n.e. | n.e. | this study | Lake Pniewskie (52°30′41″N, 16°14′27″) | 0.06 ± 0.024 | 0.604 ± 0.066 (2329) | 0.607 ± 0.107 (4164) |
| AMU-DH-10 | – | n.e. | n.e. | n.e. | this study | Lake Pniewskie (52°30′41″N, 16°14′27″) | 0.008 ± 0.001 | 0.491 ± 0.042 (5100) | 0.594 ± 0.243 (4362) |
| AMU-DH-11 | – | n.e. | n.e. | n.e. | this study | Lake Pniewskie (52°30′41″N, 16°14′27″) | 0.007 ± 0.005 | 0.448 ± 0.122 (2508) | 0.653 ± 0.161 (2385) |
| CCALA 8 | – | – | – | – | [46] | Lake Lough Neagh (54°37’06’’N, 6°23’43’’E) | 0.008 ± 0.002 | 0.666 ± 0.121 (1415) | 0.762 ± 0.131 (3272) |
| SAG 31.79 | – | – | – | – | [46] | Lake Plußee (54°11’00.7’’N, 10°26’45.9’’E) | 0.008 ± 0.001 | 0.726 ± 0.147 (2719) | 0.679 ± 0.189 (3794) |
| Aphanizomenon sp. | |||||||||
| BA 69 | n.e. | n.e. | n.e. | n.e. | Puck Bay, Baltic Sea | 0.008 ± 0.001 | 0.553 ± 0.272 (2545) | 0.809 ± 0.248 (3505) | |
| P. agardhii | |||||||||
| SAG 6.89 | – | + | + | + | [46] | Lake Plußee (54°11’00.7’’N, 10°26’45.9’’E) | 0.007 ± 0.002 | 0.46 ± 0.067 (1416) | 0.61 ± 0.146 (2625) |
| R. raciborskii | |||||||||
| AMU-DH-12 | – | n.e. | n.e. | n.e. | this study | Lake Pniewskie (52°30′41″N, 16°14′27″) | 0.008 ± 0.001 | 0.586 ± 0.09 (2851) | 0.582 ± 0.199 (2186) |
| SAG 1.97 | – | – | – | – | [46] | Lake Balaton (46°48’51.0’’N, 17°45’52.8’’E) | 0.013 ± 0.003 | 0.459 ± 0.064 (2705) | 0.819 ± 0.171 (3151) |
| Studied Biomarker | Stage of Culture Development | Strain | Stage of Culture Development × Strain |
|---|---|---|---|
| GSH | F1,140 = 250.3; p < 0.001 | F13,140 = 13.1; p < 0.001 | F13,140 = 10.0; p < 0.001 |
| GST | F1,140 = 148.3; p < 0.001 | F13,140 = 8.7; p < 0.001 | F13,140 = 10.2; p < 0.001 |
| CAT | F1,140 = 36.6; p < 0.001 | F13,140 = 3.6; p < 0.001 | F13,140 = 5.5; p < 0.001 |
| TAC | F1,140 = 0.3; p = 0.601 | F13,140 = 11.6; p < 0.001 | F13,140 = 20.6; p < 0.001 |
| TBARS | F1,140 = 35.3; p < 0.001 | F13,140 = 3.7; p < 0.001 | F13,140 = 2.6; p < 0.01 |
| PC | F1,140 = 385.3; p < 0.001 | F13,140 = 33.8; p < 0.001 | F13,140 = 25.1; p < 0.001 |
| DSBs | F1,140 = 24.6; p < 0.001 | F13,140 = 6.8; p < 0.001 | F13,140 = 3.9; p < 0.001 |
| PSP | F1,140 = 124.9; p < 0.001 | F13,140 = 14.7; p < 0.001 | F13,140 = 12.4; p < 0.001 |
| ChE | F1,140 = 38.6; p < 0.001 | F13,140 = 26.6; p < 0.001 | F13,140 = 6.8; p < 0.001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wejnerowski, Ł.; Falfushynska, H.; Horyn, O.; Osypenko, I.; Kokociński, M.; Meriluoto, J.; Jurczak, T.; Poniedziałek, B.; Pniewski, F.; Rzymski, P. In Vitro Toxicological Screening of Stable and Senescing Cultures of Aphanizomenon, Planktothrix, and Raphidiopsis. Toxins 2020, 12, 400. https://doi.org/10.3390/toxins12060400
Wejnerowski Ł, Falfushynska H, Horyn O, Osypenko I, Kokociński M, Meriluoto J, Jurczak T, Poniedziałek B, Pniewski F, Rzymski P. In Vitro Toxicological Screening of Stable and Senescing Cultures of Aphanizomenon, Planktothrix, and Raphidiopsis. Toxins. 2020; 12(6):400. https://doi.org/10.3390/toxins12060400
Chicago/Turabian StyleWejnerowski, Łukasz, Halina Falfushynska, Oksana Horyn, Inna Osypenko, Mikołaj Kokociński, Jussi Meriluoto, Tomasz Jurczak, Barbara Poniedziałek, Filip Pniewski, and Piotr Rzymski. 2020. "In Vitro Toxicological Screening of Stable and Senescing Cultures of Aphanizomenon, Planktothrix, and Raphidiopsis" Toxins 12, no. 6: 400. https://doi.org/10.3390/toxins12060400
APA StyleWejnerowski, Ł., Falfushynska, H., Horyn, O., Osypenko, I., Kokociński, M., Meriluoto, J., Jurczak, T., Poniedziałek, B., Pniewski, F., & Rzymski, P. (2020). In Vitro Toxicological Screening of Stable and Senescing Cultures of Aphanizomenon, Planktothrix, and Raphidiopsis. Toxins, 12(6), 400. https://doi.org/10.3390/toxins12060400