The Southeast Alaska Tribal Ocean Research (SEATOR) Partnership: Addressing Data Gaps in Harmful Algal Bloom Monitoring and Shellfish Safety in Southeast Alaska


 , and
, and
Abstract
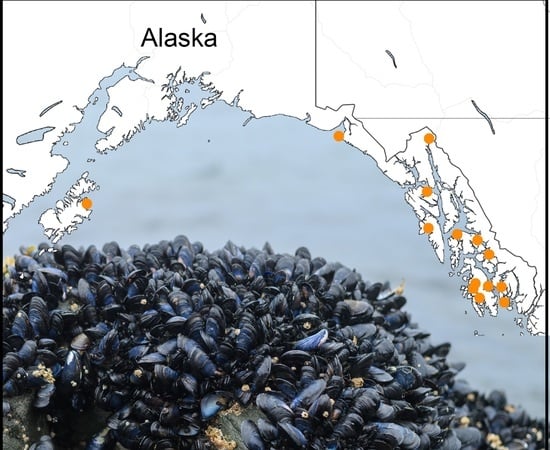
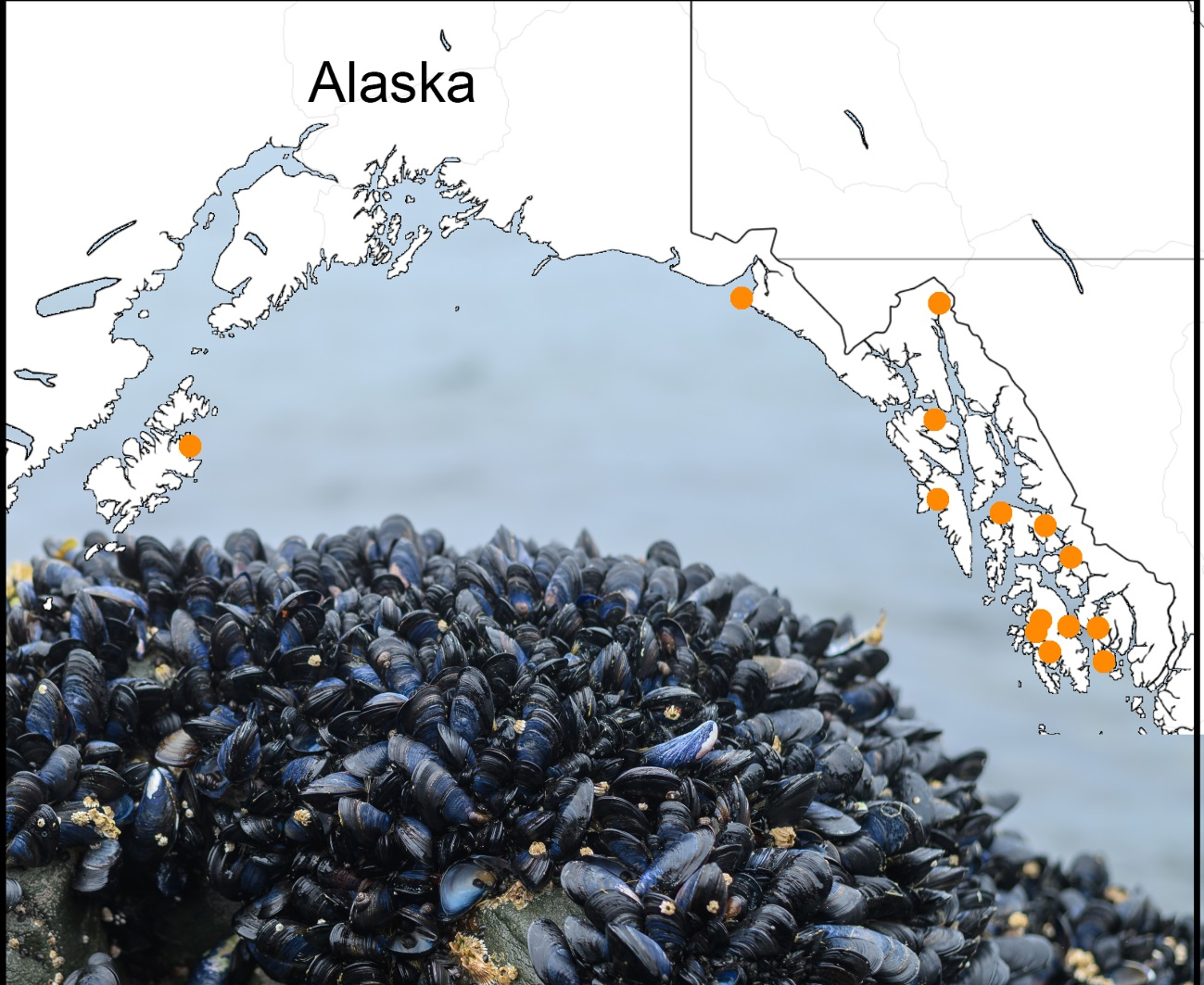
:
1. Introduction
2. Results and Discussion
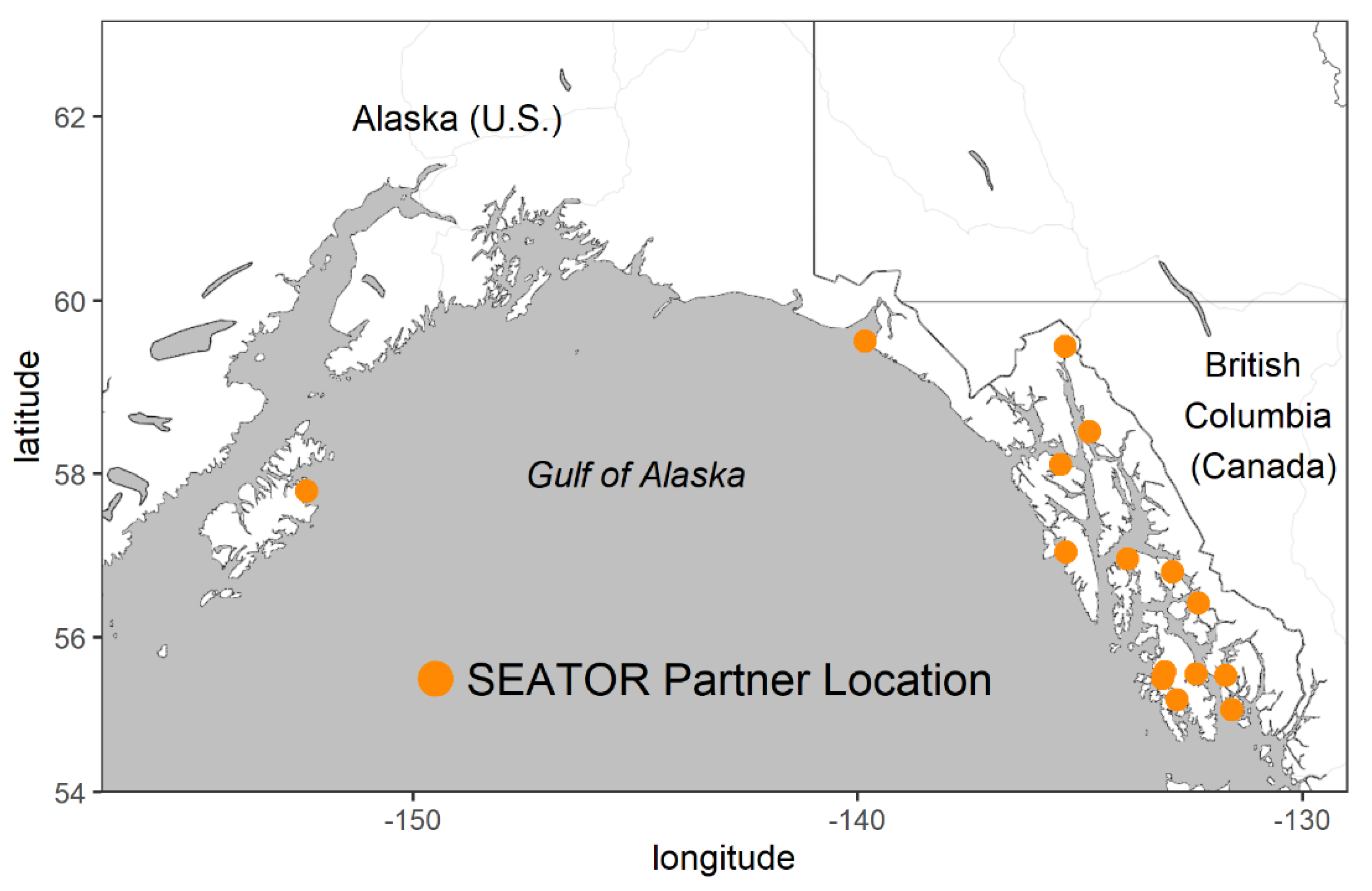
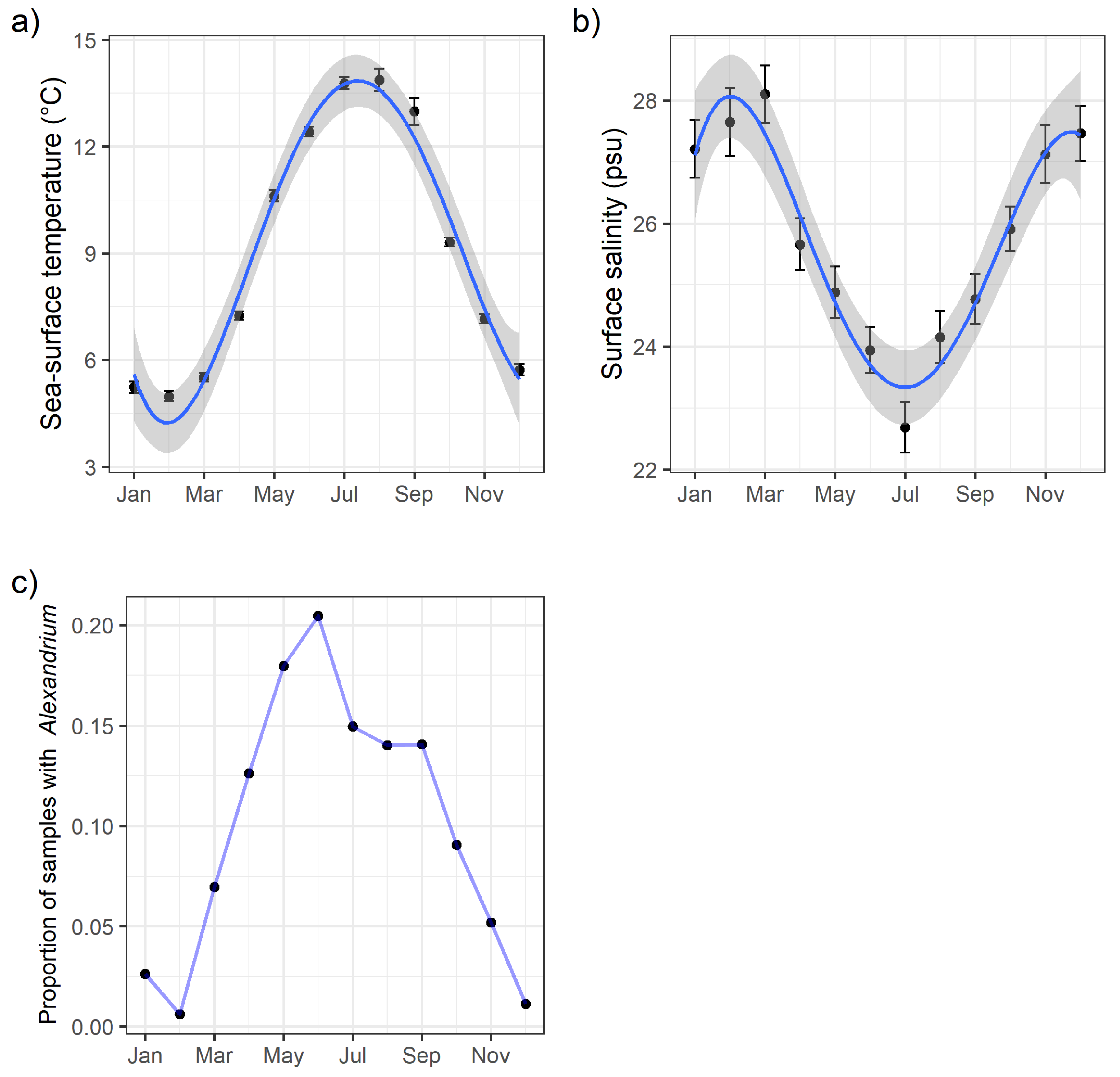
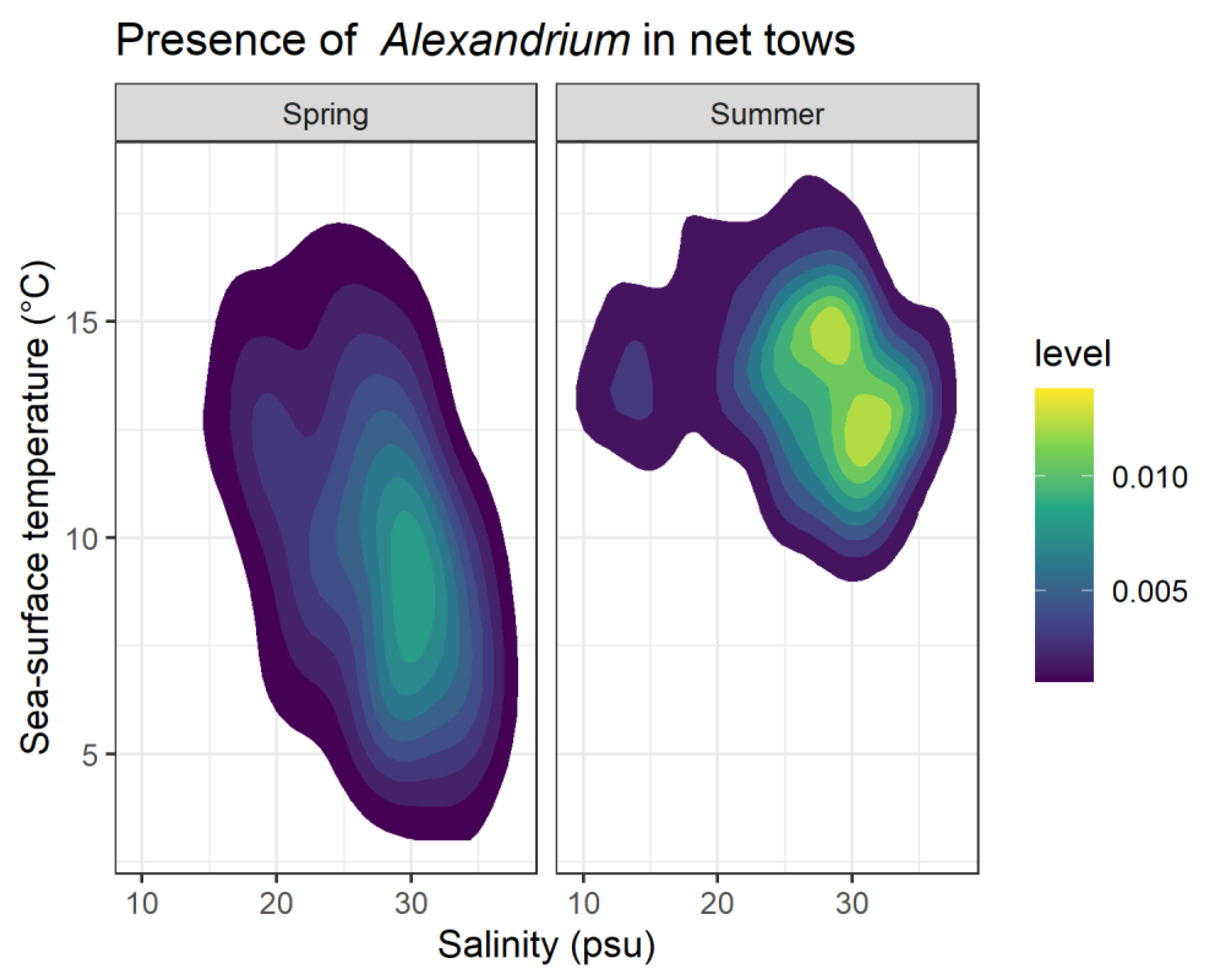
2.1. HAB and Oceanographic Observations
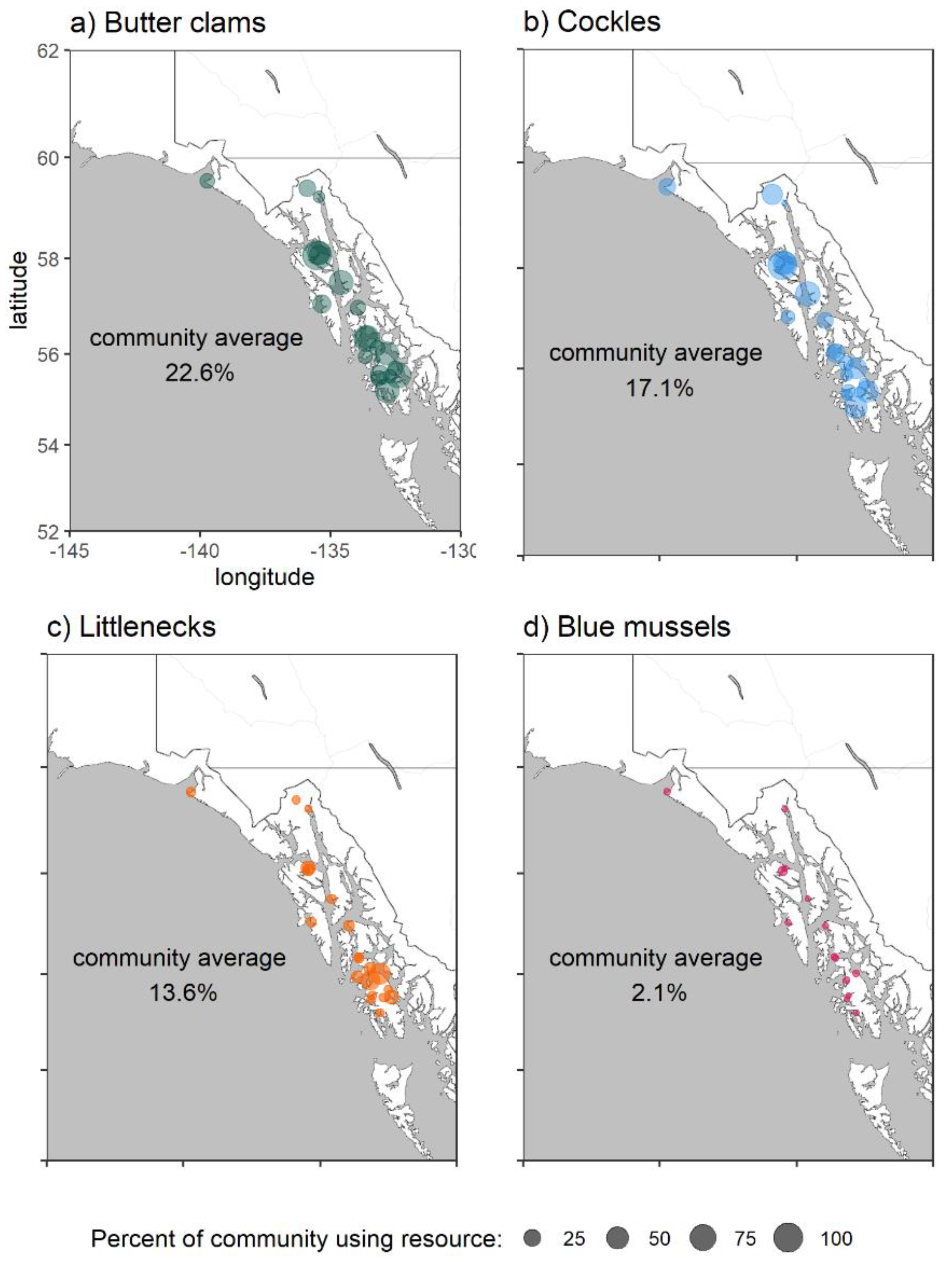
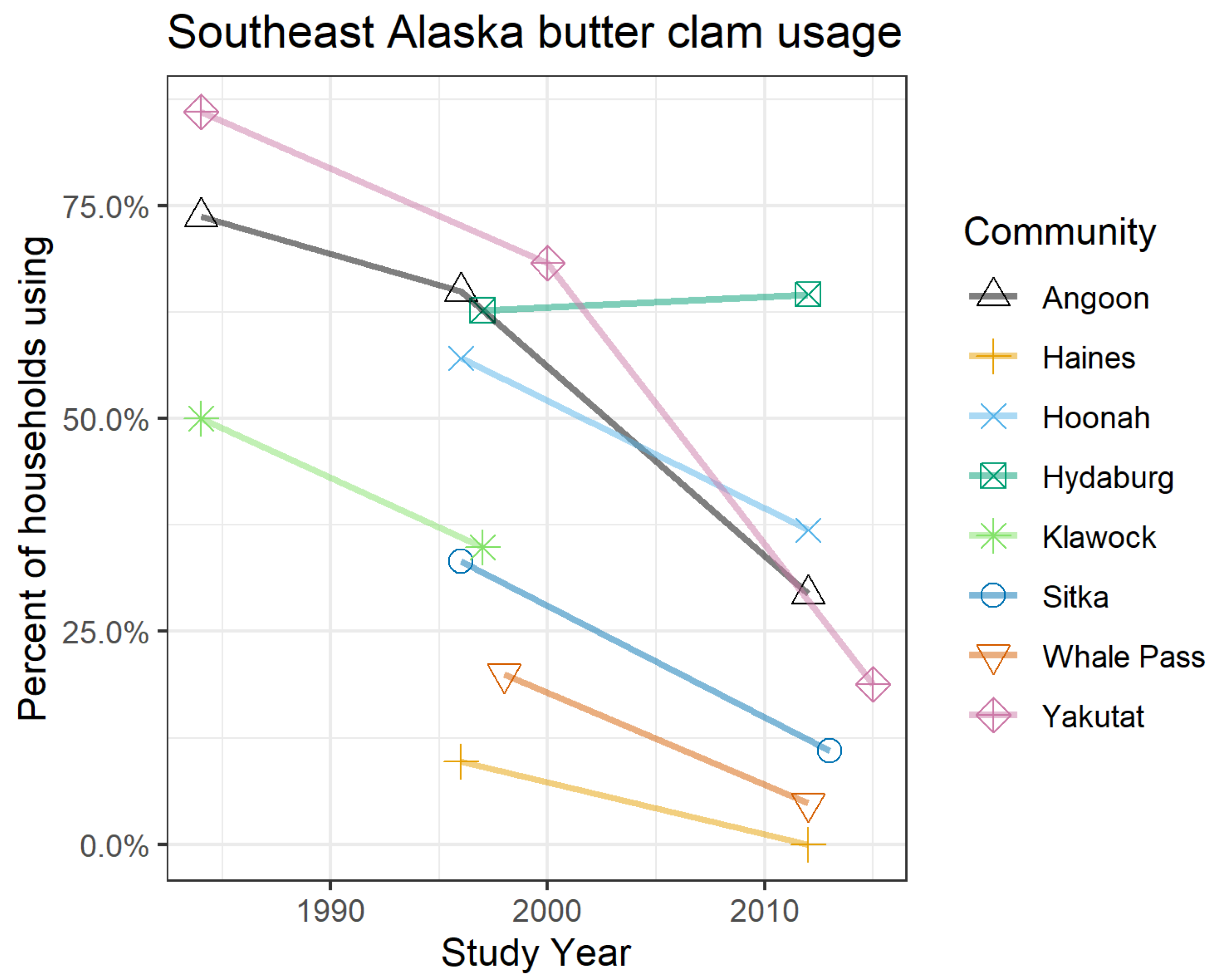
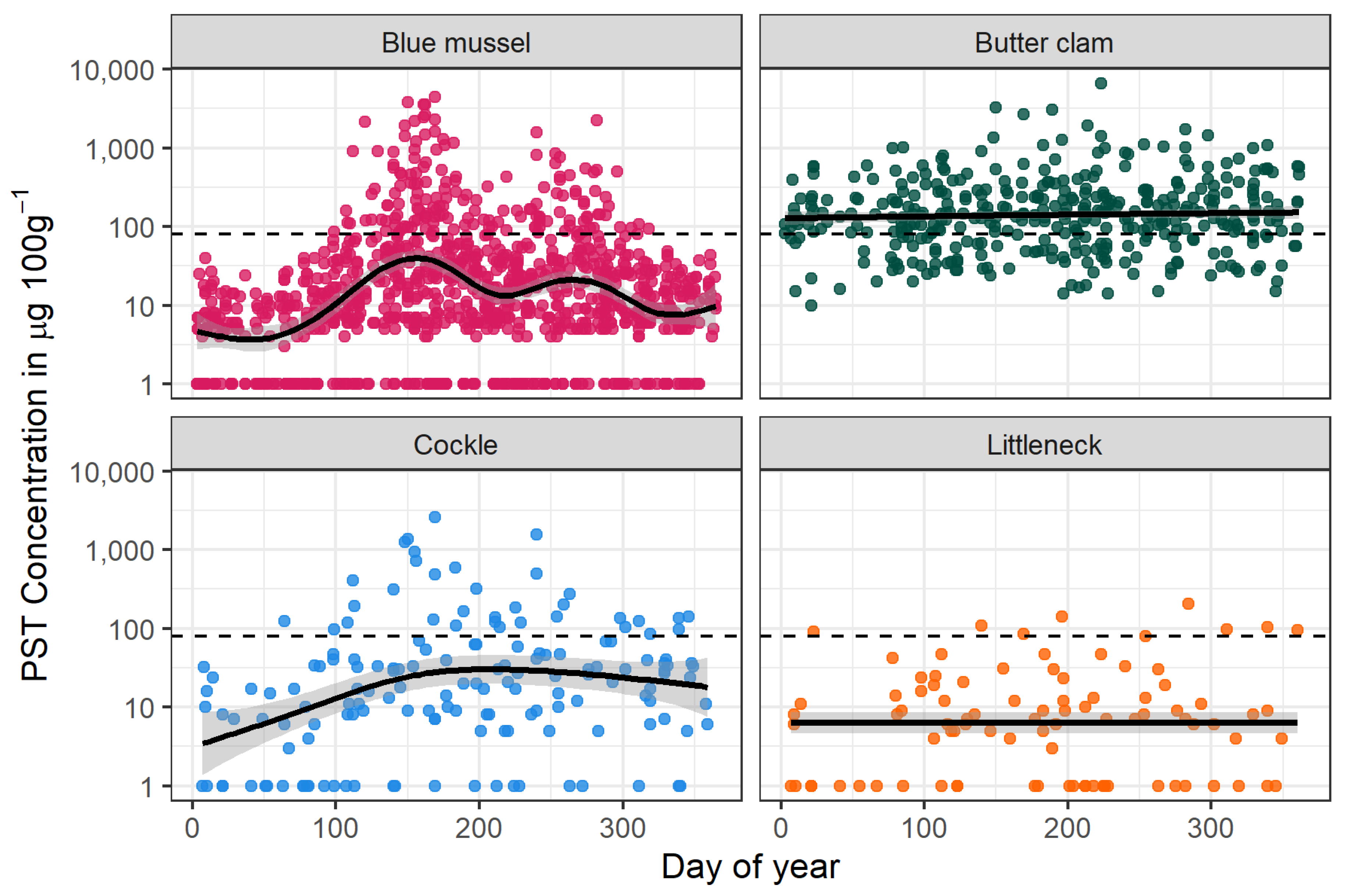
2.2. PSTs in Southeast Alaska Shellfish
2.3. Value of These Data as a Platform for Modeling Efforts
3. Conclusions
4. Materials and Methods
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Van Dolah, F.M. Marine algal toxins: Origins, health effects, and their increased occurrence. Environ. Health Perspect. 2000, 108, 133–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, F.X.; Tatters, A.O.; Hutchins, D.A. A Global change and the future of harmful algal blooms in the ocean. Mar. Ecol. Prog. Ser. 2012, 470, 207–233. [Google Scholar] [CrossRef] [Green Version]
- Grattan, L.M.; Holobaugh, S.; Morris, J.G. Harmful algal blooms and public health. Harmful Algae 2016, 57, 2–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, K.J.; Carey, B.; O’halloran, J.; van Pelt, F.N.A.M.; Škrabáková, Z. Shellfish toxicity: Human health implications of marine algal toxins. Epidemiol. Infect. 2010, 138, 927–940. [Google Scholar] [CrossRef] [Green Version]
- Gobler, C.J.; Doherty, O.M.; Hattenrath-Lehmann, T.K.; Griffith, A.W.; Kang, Y.; Litaker, R.W. Ocean warming since 1982 has expanded the niche of toxic algal blooms in the North Atlantic and North Pacific oceans. Proc. Natl. Acad. Sci. USA 2017, 114. [Google Scholar] [CrossRef] [Green Version]
- Wells, M.L.; Karlson, B.; Wulff, A.; Kudela, R.; Trick, C.; Asnaghi, V.; Berdalet, E.; Cochlan, W.; Davidson, K.; De Rijcke, M.; et al. Future HAB science: Directions and challenges in a changing climate. Harmful Algae 2020, 91, 101632. [Google Scholar] [CrossRef]
- Anderson, D.M.; Alpermann, T.J.; Cembella, A.D.; Collos, Y.; Masseret, E.; Montresor, M. The globally distributed genus Alexandrium: Multifaceted roles in marine ecosystems and impacts on human health. Harmful Algae 2012, 14, 10–35. [Google Scholar] [CrossRef] [Green Version]
- Trainer, V.L.; Sullivan, K.; Eberhart, B.-T.L.; Shuler, A.; Hignutt, E.; Kiser, J.; Eckert, G.L.; Shumway, S.E.; Morton, S.L. Enhancing Shellfish Safety in Alaska through Monitoring of Harmful Algae and Their Toxins. J. Shellfish Res. 2014, 33, 531–539. [Google Scholar] [CrossRef]
- Hallegraeff, G.M. A review of harmful algal blooms and their apparent global increase. Phycologia 1993, 32, 79–99. [Google Scholar] [CrossRef] [Green Version]
- Gessner, B.D.; Middaugh, J.P. Paralytic shellfish poisoning in Alaska: A 20-year retrospective analysis. Am. J. Epidemiol. 1995, 141, 766–770. [Google Scholar] [CrossRef]
- Castrodale, L. Paralytic Shellfish Poisoning—Alaska, 1993–2014; State of Alaska Epidemiology Bulletin; Department of Health and Social Services: Anchorage, AK, USA, 2015.
- Trainer, V.L.; Hardy, F.J. Integrative Monitoring of Marine and Freshwater Harmful Algae in Washington State for Public Health Protection. Toxins (Basel) 2015, 7, 1206–1234. [Google Scholar] [CrossRef] [PubMed]
- Newton, R.; Moss, M. The Subsistence Lifeway of the Tlingit People: Excerpts of Oral Interviews; U.S. Dept. of Agriculture, Forest Service, Alaska Region: Juneau, AK, USA, 1984.
- Sill, L.A.; Koster, D. The Harvest and Use of Wild Resources in Haines, Hoonah, Angoon, Whale Pass, and Hydaburg, Alaska, 2012; Alaska Department of Fish and Game Division of Subsistence: Juneau, AK, USA, 2017; Technical Paper No. 399.
- Alaska Department of Fish and Game Community Subsistence Information System (CSIS). Available online: https://www.adfg.alaska.gov/sb/CSIS/ (accessed on 13 January 2020).
- Moss, M.L. Shellfish, Gender, and Status on the Northwest Coast: Reconciling Archeological, Ethnographic, and Ethnohistorical Records of the Tlingit. Am. Anthropol. 1993, 95, 631–652. [Google Scholar] [CrossRef]
- Finnis, S.; Krstic, N.; McIntyre, L.; Nelson, T.A.; Henderson, S.B. Spatiotemporal patterns of paralytic shellfish toxins and their relationships with environmental variables in British Columbia, Canada from 2002 to 2012. Environ. Res. 2017, 156, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Valbi, E.; Ricci, F.; Capellacci, S.; Casabianca, S.; Scardi, M.; Penna, A. A model predicting the PSP toxic dinoflagellate Alexandrium minutum occurrence in the coastal waters of the NW Adriatic Sea. Sci. Rep. 2019, 9, 4166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, S.K.; Mantua, N.J.; Hickey, B.M.; Trainer, V.L. Recent trends in paralytic shellfish toxins in Puget Sound, relationships to climate, and capacity for prediction of toxic events. Harmful Algae 2009, 8, 463–477. [Google Scholar] [CrossRef]
- Peng, M.; Zhang, A.; Yang, Z. Implementation of the Gulf of Maine Operational Forecast System (GOMOFS) and the Semi-Operational Nowcast/forecast Skill Assessment; NOS CO-OPS; Center for Operational Oceanographic Products and Services: Spring, MD, USA, 2018.
- Tobin, E.D.; Wallace, C.L.; Crumpton, C.; Johnson, G.; Eckert, G.L. Environmental drivers of paralytic shellfish toxin producing Alexandrium catenella blooms in a fjord system of northern Southeast Alaska. Harmful Algae 2019, 88, 101659. [Google Scholar] [CrossRef]
- Weingartner, T.; Eisner, L.; Eckert, G.L.; Danielson, S.; Bellwood, D. Southeast Alaska: Oceanographic Habitats and Linkages. J. Biogeogr. 2009, 36, 387–400. [Google Scholar] [CrossRef]
- Bill, B.D.; Moore, S.K.; Hay, L.R.; Anderson, D.M.; Trainer, V.L. Effects of temperature and salinity on the growth of Alexandrium (Dinophyceae) isolates from the Salish Sea. J. Phycol. 2016, 52, 230–238. [Google Scholar] [CrossRef] [Green Version]
- Bruce, H.E.; McLain, D.R.; Wing, B.L. Annual Physical and Chemical Oceanographic Cycles of Auke Bay, Southeastern Alaska; NMFS SSRF; NOAA: Silver Spring, MD, USA, 1977.
- Reverdin, G.; Morisset, S.; Boutin, J.; Martin, N. Rain-induced variability of near sea-surface T and S from drifter data. J. Geophys. Res. Ocean. 2012, 117. [Google Scholar] [CrossRef] [Green Version]
- McCabe, R.M.; Hickey, B.M.; Kudela, R.M.; Lefebvre, K.A.; Adams, N.G.; Bill, B.D.; Gulland, F.M.; Thomson, R.E.; Cochlan, W.P.; Trainer, V.L. An unprecedented coastwide toxic algal bloom linked to anomalous ocean conditions. Geophys. Res. Lett. 2016, 43, 10–366. [Google Scholar] [CrossRef]
- Bricelj, V.M.; Shumway, S.E. Paralytic Shellfish Toxins in Bivalve Molluscs: Occurrence, Transfer Kinetics, and Biotransformation. Rev. Fish. Sci. 1998, 6, 315–383. [Google Scholar] [CrossRef]
- Beyer, J.; Green, N.W.; Brooks, S.; Allan, I.J.; Ruus, A.; Gomes, T.; Bråte, I.L.N.; Schøyen, M. Blue mussels (Mytilus edulis spp.) as sentinel organisms in coastal pollution monitoring: A review. Mar. Environ. Res. 2017, 130, 338–365. [Google Scholar] [CrossRef] [PubMed]
- Kvitek, R.G.; Beitler, M.K. Relative insensitivity of butter clam neurons to saxitoxin: A pre-adaptation for sequestering paralytic shellfish poisoning toxins as a chemical defense. Mar. Ecol. Prog. Ser. 1991, 69, 47–54. [Google Scholar] [CrossRef]
- Brown, C.W.; Green, D.; Hickey, B.M.; Jacobs, J.M.; Lanerolle, L.W.J.; Moore, S.; Schwab, D.J.; Trainer, V.L.; Trtanj, J.; Turner, E.; et al. Towards operational forecasts of algal blooms and pathogens. In Environmental Tracking for Public Health Surveillance; Morain, S.A., Budge, A.M., Eds.; CRC Press: Boca Raton, FL, USA, 2012; Volume 11, pp. 345–368. [Google Scholar]
- Alaska Ocean Observing System. Available online: https://aoos.org/ (accessed on 17 March 2020).
- Campbell, J.L.; Rustad, L.E.; Porter, J.H.; Taylor, J.R.; Dereszynski, E.W.; Shanley, J.B.; Gries, C.; Henshaw, D.L.; Martin, M.E.; Sheldon, W.M. Quantity is nothing without quality: Automated QA/QC for streaming environmental sensor data. BioScience 2013, 63, 574–585. [Google Scholar] [CrossRef] [Green Version]
- Van Dolah, F.M.; Fire, S.E.; Leighfield, T.A.; Mikulski, C.M.; Doucette, G.J. Determination of paralytic shellfish toxins in shellfish by receptor binding assay: Collaborative study. J. AOAC Int. 2012, 95, 795–812. [Google Scholar] [CrossRef]
- King, T.; Claassen, L.; Borchert, J.; Trainer, V. SoundToxins: A Puget Sound Harmful Algae Monitoring Partnership. Salish Sea Ecosystem Conference (2018: Seattle, Wash). Available online: cedar.wwu.edu/ssec/2018ssec/allsessions/269/ (accessed on 19 June 2020).
- Washington Sea Grant. SoundToxins Manual; Washington Sea Grant: Seattle, WA, USA, 2016; p. 36. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Year | |||
|---|---|---|---|---|
| 2016 | 2017 | 2018 | 2019 | |
| Blue mussel | 916 | 3791 | 2243 | 4412 |
| (M. trossulus) | ||||
| Butter clam | 723 | 6624 | 1712 | 3081 |
| (S. gigantea) | ||||
| California mussel | NT | NT | 24 | 29 |
| (Mytilus californianus) | ||||
| Cockle | 202 | 1367 | 1565 | 2603 |
| (C. nuttallii) | ||||
| Eastern softshell | 15 | 6 | 8 | 21 |
| (Mya arenaria) | ||||
| Horse clam | 13 | BDL | 42 | 37 |
| (Tresus capax) | ||||
| Littleneck clam | 24 | 47 | 206 | 142 |
| (Leukoma staminea) | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harley, J.R.; Lanphier, K.; Kennedy, E.G.; Leighfield, T.A.; Bidlack, A.; Gribble, M.O.; Whitehead, C. The Southeast Alaska Tribal Ocean Research (SEATOR) Partnership: Addressing Data Gaps in Harmful Algal Bloom Monitoring and Shellfish Safety in Southeast Alaska. Toxins 2020, 12, 407. https://doi.org/10.3390/toxins12060407
Harley JR, Lanphier K, Kennedy EG, Leighfield TA, Bidlack A, Gribble MO, Whitehead C. The Southeast Alaska Tribal Ocean Research (SEATOR) Partnership: Addressing Data Gaps in Harmful Algal Bloom Monitoring and Shellfish Safety in Southeast Alaska. Toxins. 2020; 12(6):407. https://doi.org/10.3390/toxins12060407
Chicago/Turabian StyleHarley, John R., Kari Lanphier, Esther G. Kennedy, Tod A. Leighfield, Allison Bidlack, Matthew O. Gribble, and Christopher Whitehead. 2020. "The Southeast Alaska Tribal Ocean Research (SEATOR) Partnership: Addressing Data Gaps in Harmful Algal Bloom Monitoring and Shellfish Safety in Southeast Alaska" Toxins 12, no. 6: 407. https://doi.org/10.3390/toxins12060407
APA StyleHarley, J. R., Lanphier, K., Kennedy, E. G., Leighfield, T. A., Bidlack, A., Gribble, M. O., & Whitehead, C. (2020). The Southeast Alaska Tribal Ocean Research (SEATOR) Partnership: Addressing Data Gaps in Harmful Algal Bloom Monitoring and Shellfish Safety in Southeast Alaska. Toxins, 12(6), 407. https://doi.org/10.3390/toxins12060407