Metabolic Traits of Bovine Shiga Toxin-Producing Escherichia coli (STEC) Strains with Different Colonization Properties

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Acid Resistance
2.2. Biofilm, Curli, and Cellulose Production
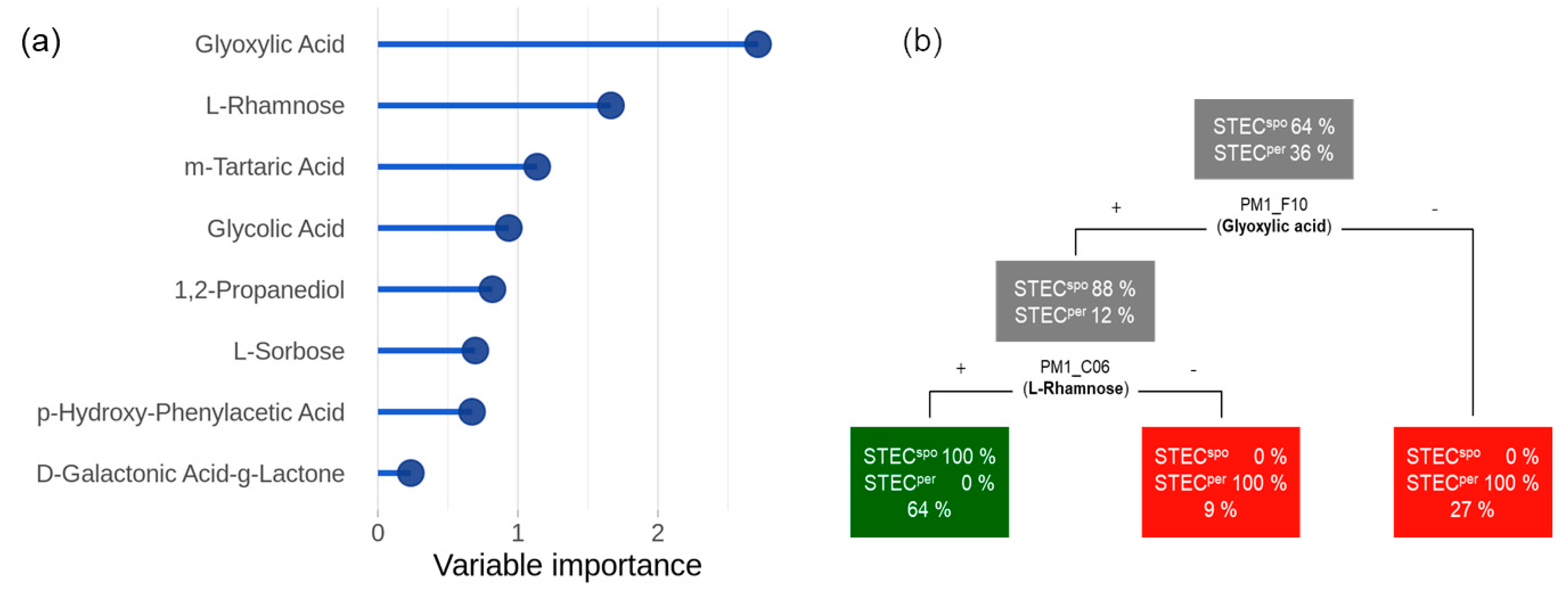
2.3. Metabolic Activity of the Strains and Classification According to Their Colonization Type
2.4. Genetic Basis for Key Metabolic Reactions
3. Discussion
4. Material and Methods
4.1. Strain Collection
4.2. Illumina Sequencing
4.3. Genome Analysis
4.4. Acid Resistance Assay
4.5. Biofilm Crystal Violet Plate Assay
4.6. Curli Detection by Congo Red Agar
4.7. Metabolic Phenotype Microarray (Omnilog®)
4.8. M9 Minimal Medium Growth Kinetics with Differing C-Sources
4.9. Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Scheutz, F.; Teel, L.D.; Beutin, L.; Pierard, D.; Buvens, G.; Karch, H.; Mellmann, A.; Caprioli, A.; Tozzoli, R.; Morabito, S.; et al. Multicenter evaluation of a sequence-based protocol for subtyping Shiga toxins and standardizing Stx nomenclature. J. Clin. Microbiol. 2012, 50, 2951–2963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, X.; Fu, S.; Zhang, J.; Fan, R.; Xu, Y.; Sun, H.; He, X.; Xu, J.; Xiong, Y. Identification and pathogenomic analysis of an Escherichia coli strain producing a novel Shiga toxin 2 subtype. Sci. Rep. 2018, 8, 6756. [Google Scholar] [CrossRef] [PubMed]
- Brooks, J.T.; Sowers, E.G.; Wells, J.G.; Greene, K.D.; Griffin, P.M.; Hoekstra, R.M.; Strockbine, N.A. Non-O157 Shiga toxin-producing Escherichia coli infections in the United States, 1983–2002. J. Infect. Dis. 2005, 192, 1422–1429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashid, R.A.; Tabata, T.A.; Oatley, M.J.; Besser, T.E.; Tarr, P.I.; Moseley, S.L. Expression of putative virulence factors of Escherichia coli O157:H7 differs in bovine and human infections. Infect. Immun. 2006, 74, 4142–4148. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, J.M.; Wagner, P.L.; Acheson, D.W.; Waldor, M.K. Comparison of Shiga toxin production by hemolytic-uremic syndrome-associated and bovine-associated Shiga toxin-producing Escherichia coli isolates. Appl. Environ. Microbiol. 2003, 69, 1059–1066. [Google Scholar] [CrossRef] [Green Version]
- Lejeune, J.T.; Abedon, S.T.; Takemura, K.; Christie, N.P.; Sreevatsan, S. Human Escherichia coli O157:H7 genetic marker in isolates of bovine origin. Emerg. Infect. Dis. 2004, 10, 1482–1485. [Google Scholar] [CrossRef]
- Lupolova, N.; Dallman, T.J.; Matthews, L.; Bono, J.L.; Gally, D.L. Support vector machine applied to predict the zoonotic potential of E. coli O157 cattle isolates. Proc. Natl. Acad. Sci. USA 2016, 113, 11312–11317. [Google Scholar] [CrossRef] [Green Version]
- Bielaszewska, M.; Mellmann, A.; Zhang, W.; Kock, R.; Fruth, A.; Bauwens, A.; Peters, G.; Karch, H. Characterisation of the Escherichia coli strain associated with an outbreak of haemolytic uraemic syndrome in Germany, 2011: A microbiological study. Lancet Infect. Dis. 2011, 11, 671–676. [Google Scholar] [CrossRef] [Green Version]
- Rasko, D.A.; Webster, D.R.; Sahl, J.W.; Bashir, A.; Boisen, N.; Scheutz, F.; Paxinos, E.E.; Sebra, R.; Chin, C.S.; Iliopoulos, D.; et al. Origins of the E. coli strain causing an outbreak of hemolytic-uremic syndrome in Germany. N. Engl. J. Med. 2011, 365, 709–717. [Google Scholar] [CrossRef] [Green Version]
- Stalb, S.; Barth, S.A.; Sobotta, K.; Liebler-Tenorio, E.; Geue, L.; Menge, C. Pro-inflammatory capacity of Escherichia coli O104:H4 outbreak strain during colonization of intestinal epithelial cells from human and cattle. Int. J. Med. Microbiol. 2018, 308, 899–911. [Google Scholar] [CrossRef]
- Hamm, K.; Barth, S.A.; Stalb, S.; Geue, L.; Liebler-Tenorio, E.; Teifke, J.P.; Lange, E.; Tauscher, K.; Kotterba, G.; Bielaszewska, M.; et al. Experimental infection of calves with Escherichia coli O104:H4 outbreak strain. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabal, A.; Geue, L.; Gomez-Barrero, S.; Barth, S.; Barcena, C.; Hamm, K.; Porrero, M.C.; Valverde, A.; Canton, R.; Menge, C.; et al. Detection of virulence-associated genes characteristic of intestinal Escherichia coli pathotypes, including the enterohemorrhagic/enteroaggregative O104:H4 in bovines from Germany and Spain. Microbiol. Immunol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Arimizu, Y.; Kirino, Y.; Sato, M.P.; Uno, K.; Sato, T.; Gotoh, Y.; Auvray, F.; Brugere, H.; Oswald, E.; Mainil, J.G.; et al. Large-scale genome analysis of bovine commensal Escherichia coli reveals that bovine-adapted E. coli lineages are serving as evolutionary sources of the emergence of human intestinal pathogenic strains. Genome Res. 2019, 29, 1495–1505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, M.A.; Menge, C.; Casey, T.A.; Laegreid, W.; Bosworth, B.T.; Dean-Nystrom, E.A. Bovine immune response to shiga-toxigenic Escherichia coli O157:H7. Clin. Vaccine Immunol. 2006, 13, 1322–1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menge, C.; Stamm, I.; Blessenohl, M.; Wieler, L.H.; Baljer, G. Verotoxin 1 from Escherichia coli affects Gb3/CD77+ bovine lymphocytes independent of interleukin-2, tumor necrosis factor-alpha, and interferon-alpha. Exp. Biol. Med. 2003, 228, 377–386. [Google Scholar] [CrossRef]
- Menge, C.; Loos, D.; Bridger, P.S.; Barth, S.; Werling, D.; Baljer, G. Bovine macrophages sense Escherichia coli Shiga toxin 1. Innate Immun. 2015, 21, 655–664. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, N.; Barth, S.A.; Frahm, J.; Meyer, U.; Danicke, S.; Geue, L.; Menge, C. Decreased STEC shedding by cattle following passive and active vaccination based on recombinant Escherichia coli Shiga toxoids. Vet. Res. 2018, 49. [Google Scholar] [CrossRef] [Green Version]
- Chase-Topping, M.E.; McKendrick, I.J.; Pearce, M.C.; MacDonald, P.; Matthews, L.; Halliday, J.; Allison, L.; Fenlon, D.; Low, J.C.; Gunn, G.; et al. Risk factors for the presence of high-level shedders of Escherichia coli O157 on Scottish farms. J. Clin. Microbiol. 2007, 45, 1594–1603. [Google Scholar] [CrossRef] [Green Version]
- Matthews, L.; Reeve, R.; Gally, D.L.; Low, J.C.; Woolhouse, M.E.; McAteer, S.P.; Locking, M.E.; Chase-Topping, M.E.; Haydon, D.T.; Allison, L.J.; et al. Predicting the public health benefit of vaccinating cattle against Escherichia coli O157. Proc. Natl. Acad. Sci. USA 2013, 110, 16265–16270. [Google Scholar] [CrossRef] [Green Version]
- Munns, K.D.; Selinger, L.B.; Stanford, K.; Guan, L.; Callaway, T.R.; McAllister, T.A. Perspectives on super-shedding of Escherichia coli O157:H7 by cattle. Foodborne Pathog. Dis. 2015, 12, 89–103. [Google Scholar] [CrossRef]
- Fitzgerald, S.F.; Beckett, A.E.; Palarea-Albaladejo, J.; McAteer, S.; Shaaban, S.; Morgan, J.; Ahmad, N.I.; Young, R.; Mabbott, N.A.; Morrison, L.; et al. Shiga toxin sub-type 2a increases the efficiency of Escherichia coli O157 transmission between animals and restricts epithelial regeneration in bovine enteroids. PLoS Pathog. 2019, 15, e1008003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geue, L.; Segura-Alvarez, M.; Conraths, F.J.; Kuczius, T.; Bockemühl, J.; Karch, H.; Gallien, P. A long-term study on the prevalence of Shiga toxin-producing Escherichia coli (STEC) on four German cattle farms. Epidemiol. Infect. 2002, 129, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Geue, L.; Schares, S.; Mintel, B.; Conraths, F.J.; Muller, E.; Ehricht, R. Rapid microarray-based genotyping of enterohemorrhagic Escherichia coli serotype O156:H25/H-/Hnt isolates from cattle and clonal relationship analysis. Appl. Environ. Microbiol. 2010, 76, 5510–5519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geue, L.; Selhorst, T.; Schnick, C.; Mintel, B.; Conraths, F.J. Analysis of the clonal relationship of shiga toxin-producing Escherichia coli serogroup O165:H25 isolated from cattle. Appl. Environ. Microbiol. 2006, 72, 2254–2259. [Google Scholar] [CrossRef] [Green Version]
- Geue, L.; Klare, S.; Schnick, C.; Mintel, B.; Meyer, K.; Conraths, F.J. Analysis of the clonal relationship of serotype O26:H11 enterohemorrhagic Escherichia coli isolates from cattle. Appl. Environ. Microbiol. 2009, 75, 6947–6953. [Google Scholar] [CrossRef] [Green Version]
- Barth, S.A.; Menge, C.; Eichhorn, I.; Semmler, T.; Wieler, L.H.; Pickard, D.; Belka, A.; Berens, C.; Geue, L. The accessory genome of Shiga toxin-producing Escherichia coli defines a persistent colonization type in cattle. Appl. Environ. Microbiol. 2016, 82, 5455–5464. [Google Scholar] [CrossRef] [Green Version]
- Pellicer, M.T.; Badia, J.; Aguilar, J.; Baldoma, L. glc locus of Escherichia coli: Characterization of genes encoding the subunits of glycolate oxidase and the glc regulator protein. J. Bacteriol. 1996, 178, 2051–2059. [Google Scholar] [CrossRef] [Green Version]
- Tate, C.G.; Muiry, J.A.; Henderson, P.J. Mapping, cloning, expression, and sequencing of the rhaT gene, which encodes a novel L-rhamnose-H+ transport protein in Salmonella typhimurium and Escherichia coli. J. Biol. Chem. 1992, 267, 6923–6932. [Google Scholar]
- Via, P.; Badia, J.; Baldoma, L.; Obradors, N.; Aguilar, J. Transcriptional regulation of the Escherichia coli rhaT gene. Microbiology 1996, 142, 1833–1840. [Google Scholar] [CrossRef] [Green Version]
- Rodionova, I.A.; Li, X.; Thiel, V.; Stolyar, S.; Stanton, K.; Fredrickson, J.K.; Bryant, D.A.; Osterman, A.L.; Best, A.A.; Rodionov, D.A. Comparative genomics and functional analysis of rhamnose catabolic pathways and regulons in bacteria. Front. Microbiol. 2013, 4, 407. [Google Scholar] [CrossRef] [Green Version]
- Moralejo, P.; Egan, S.M.; Hidalgo, E.; Aguilar, J. Sequencing and characterization of a gene cluster encoding the enzymes for L-rhamnose metabolism in Escherichia coli. J. Bacteriol. 1993, 175, 5585–5594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, K.S.; Kim, C.; Kim, I.; Yoo, S.; Choi, B.S.; Park, C. NMR application probes a novel and ubiquitous family of enzymes that alter monosaccharide configuration. J. Biol. Chem. 2004, 279, 25544–25548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaw, D.J.; Jenkins, C.; Pearce, M.C.; Cheasty, T.; Gunn, G.J.; Dougan, G.; Smith, H.R.; Woolhouse, M.E.; Frankel, G. Shedding patterns of verocytotoxin-producing Escherichia coli strains in a cohort of calves and their dams on a Scottish beef farm. Appl. Environ. Microbiol. 2004, 70, 7456–7465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callaway, T.R.; Carr, M.A.; Edrington, T.S.; Anderson, R.C.; Nisbet, D.J. Diet, Escherichia coli O157:H7, and cattle: A review after 10 years. Curr. Issues Mol. Biol. 2009, 11, 67–79. [Google Scholar]
- Fröhlich, J.; Baljer, G.; Menge, C. Maternally and naturally acquired antibodies to Shiga toxins in a cohort of calves shedding Shiga-toxigenic Escherichia coli. Appl. Environ. Microbiol. 2009, 75, 3695–3704. [Google Scholar] [CrossRef] [Green Version]
- Wirth, T.; Falush, D.; Lan, R.; Colles, F.; Mensa, P.; Wieler, L.H.; Karch, H.; Reeves, P.R.; Maiden, M.C.; Ochman, H.; et al. Sex and virulence in Escherichia coli: An evolutionary perspective. Mol. Microbiol. 2006, 60, 1136–1151. [Google Scholar] [CrossRef] [Green Version]
- Tuttle, J.; Gomez, T.; Doyle, M.P.; Wells, J.G.; Zhao, T.; Tauxe, R.V.; Griffin, P.M. Lessons from a large outbreak of Escherichia coli O157:H7 infections: Insights into the infectious dose and method of widespread contamination of hamburger patties. Epidemiol. Infect. 1999, 122, 185–192. [Google Scholar] [CrossRef]
- Van Winden, S.C.L.; Muller, K.E.; Kuiper, R.; Noordhuizen, J.P.T.M. Studies on the pH value of abomasal contents in dairy cows during the first 3 weeks after calving. J. Vet. Med. A 2002, 49, 157–160. [Google Scholar] [CrossRef]
- Sarashina, T.; Ichijo, S.; Takahashi, J.; Osame, S. Origin of abomasum gas in the cows with displaced abomasum. Jpn. J. Vet. Sci. 1990, 52, 371–378. [Google Scholar] [CrossRef]
- Bhaskar, K.R.; Garik, P.; Turner, B.S.; Bradley, J.D.; Bansil, R.; Stanley, H.E.; Lamont, J.T. Viscous fingering of HCl through gastric mucin. Nature 1992, 360, 458–461. [Google Scholar] [CrossRef]
- Aquino, P.; Honda, B.; Jaini, S.; Lyubetskaya, A.; Hosur, K.; Chiu, J.G.; Ekladious, I.; Hu, D.; Jin, L.; Sayeg, M.K.; et al. Coordinated regulation of acid resistance in Escherichia coli. BMC Syst. Biol. 2017, 11, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, S.; Meng, J.; Bhagwat, A.A. Availability of glutamate and arginine during acid challenge determines cell density-dependent survival phenotype of Escherichia coli strains. Appl. Environ. Microbiol. 2001, 67, 4914–4918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Audia, J.P.; Webb, C.C.; Foster, J.W. Breaking through the acid barrier: An orchestrated response to proton stress by enteric bacteria. Int. J. Med. Microbiol. 2001, 291, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Parker, C.T.; Kyle, J.L.; Huynh, S.; Carter, M.Q.; Brandl, M.T.; Mandrell, R.E. Distinct transcriptional profiles and phenotypes exhibited by Escherichia coli O157:H7 isolates related to the 2006 spinach-associated outbreak. Appl. Environ. Microbiol. 2012, 78, 455–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zogaj, X.; Nimtz, M.; Rohde, M.; Bokranz, W.; Römling, U. The multicellular morphotypes of Salmonella typhimurium and Escherichia coli produce cellulose as the second component of the extracellular matrix. Mol. Microbiol. 2001, 39, 1452–1463. [Google Scholar] [CrossRef] [PubMed]
- Römling, U.; Bian, Z.; Hammar, M.; Sierralta, W.D.; Normark, S. Curli fibers are highly conserved between Salmonella typhimurium and Escherichia coli with respect to operon structure and regulation. J. Bacteriol. 1998, 180, 722–731. [Google Scholar] [CrossRef] [Green Version]
- Uhlich, G.A.; Chen, C.Y.; Cottrell, B.J.; Nguyen, L.H. Growth media and temperature effects on biofilm formation by serotype O157:H7 and non-O157 Shiga toxin-producing Escherichia coli. FEMS Microbiol. Lett. 2014, 354, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Brown, P.K.; Dozois, C.M.; Nickerson, C.A.; Zuppardo, A.; Terlonge, J.; Curtiss, R. MlrA, a novel regulator of curli (AgF) and extracellular matrix synthesis by Escherichia coli and Salmonella enterica serovar typhimurium. Mol. Microbiol. 2001, 41, 349–363. [Google Scholar] [CrossRef]
- Mika, F.; Hengge, R. Small RNAs in the control of RpoS, CsgD, and biofilm architecture of Escherichia coli. RNA Biol. 2014, 11, 494–507. [Google Scholar] [CrossRef] [Green Version]
- Meric, G.; Kemsley, E.K.; Falush, D.; Saggers, E.J.; Lucchini, S. Phylogenetic distribution of traits associated with plant colonization in Escherichia coli. Environ. Microbiol. 2013, 15, 487–501. [Google Scholar] [CrossRef]
- Shaikh, N.; Tarr, P.I. Escherichia coli O157:H7 Shiga toxin-encoding bacteriophages: Integrations, excisions, truncations, and evolutionary implications. J. Bacteriol. 2003, 185, 3596–3605. [Google Scholar] [CrossRef] [Green Version]
- Uhlich, G.A.; Chen, C.Y.; Cottrell, B.J.; Hofmann, C.S.; Dudley, E.G.; Strobaugh, T.P.; Nguyen, L.H. Phage insertion in mlrA and variations in rpoS limit curli expression and biofilm formation in Escherichia coli serotype O157: H7. Microbiology 2013, 159, 1586–1596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geue, L.; Menge, C.; Berens, C.; Barth, S.A. Complete annotated genome sequences of two Shiga toxin-producing Escherichia coli strains and one atypical enteropathogenic E. coli strain, isolated from naturally colonized cattle of german origin. Genome Announc. 2017, 5. [Google Scholar] [CrossRef] [Green Version]
- Tate, C.G.; Henderson, P.J. Membrane topology of the L-rhamnose-H+ transport protein (RhaT) from enterobacteria. J. Biol. Chem. 1993, 268, 26850–26857. [Google Scholar]
- Alqasim, A.; Emes, R.; Clark, G.; Newcombe, J.; La Ragione, R.; McNally, A. Phenotypic microarrays suggest Escherichia coli ST131 is not a metabolically distinct lineage of extra-intestinal pathogenic E. coli. PLoS ONE 2014, 9, e88374. [Google Scholar] [CrossRef] [Green Version]
- Davis, M.A.; Besser, T.E.; Orfe, L.H.; Baker, K.N.K.; Lanier, A.S.; Broschat, S.L.; New, D.; Call, D.R. Genotypic-phenotypic discrepancies between antibiotic resistance characteristics of Escherichia coli Isolates from calves in management settings with high and low antibiotic use. Appl. Environ. Microbiol. 2011, 77, 3293–3299. [Google Scholar] [CrossRef] [Green Version]
- Ruppe, E.; Cherkaoui, A.; Charretier, Y.; Girard, M.; Schicklin, S.; Lazarevic, V.; Schrenzel, J. From genotype to antibiotic susceptibility phenotype in the order enterobacterales: A clinical perspective. Clin. Microbiol. Infect. 2019. [Google Scholar] [CrossRef] [PubMed]
- Berger, P.; Kouzel, I.U.; Berger, M.; Haarmann, N.; Dobrindt, U.; Koudelka, G.B.; Mellmann, A. Carriage of Shiga toxin phage profoundly affects Escherichia coli gene expression and carbon source utilization. BMC Genome 2019, 20, 504. [Google Scholar] [CrossRef] [PubMed]
- Saile, N.; Voigt, A.; Kessler, S.; Stressler, T.; Klumpp, J.; Fischer, L.; Schmidt, H. Escherichia coli O157:H7 strain EDL933 harbors multiple functional prophage-associated genes necessary for the utilization of 5-N-acetyl-9-O-acetyl neuraminic acid as a growth substrate. Appl. Environ. Microbiol. 2016, 82, 5940–5950. [Google Scholar] [CrossRef] [Green Version]
- Langridge, G.C.; Fookes, M.; Connor, T.R.; Feltwell, T.; Feasey, N.; Parsons, B.N.; Seth-Smith, H.M.B.; Barquist, L.; Stedman, A.; Humphrey, T.; et al. Patterns of genome evolution that have accompanied host adaptation in Salmonella. Proc. Natl. Acad. Sci. USA 2015, 112, 863–868. [Google Scholar] [CrossRef] [Green Version]
- Leopold, S.R.; Magrini, V.; Holt, N.J.; Shaikh, N.; Mardis, E.R.; Cagno, J.; Ogura, Y.; Iguchi, A.; Hayashi, T.; Mellmann, A.; et al. A precise reconstruction of the emergence and constrained radiations of Escherichia coli O157 portrayed by backbone concatenomic analysis. Proc. Natl. Acad. Sci. USA 2009, 106, 8713–8718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barth, S.A.; Menge, C.; Eichhorn, I.; Semmler, T.; Pickard, D.; Geue, L. Evaluation of applicability of DNA microarray-based characterization of bovine Shiga toxin-producing Escherichia coli isolates using whole genome sequence analysis. J. Vet. Diagn. Investig. 2017, 29, 721–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clermont, O.; Bonacorsi, S.; Bingen, E. Rapid and simple determination of the Escherichia coli phylogenetic group. Appl. Environ. Microbiol. 2000, 66, 4555–4558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, R.K.; Jain, M. NGS QC Toolkit: A toolkit for quality control of next generation sequencing data. PLoS ONE 2012, 7, e30619. [Google Scholar] [CrossRef]
- Coldewey, S.M.; Hartmann, M.; Schmidt, D.S.; Engelking, U.; Ukena, S.N.; Gunzer, F. Impact of the rpoS genotype for acid resistance patterns of pathogenic and probiotic Escherichia coli. BMC Microbiol. 2007, 7, 21. [Google Scholar] [CrossRef] [Green Version]
- O’Toole, G.A. Microtiter dish biofilm formation assay. J. Vis. Exp. JoVE 2011. [Google Scholar] [CrossRef]
- Richter, A.M.; Povolotsky, T.L.; Wieler, L.H.; Hengge, R. Cyclic-di-GMP signalling and biofilm-related properties of the Shiga toxin-producing 2011 German outbreak Escherichia coli O104:H4. EMBO Mol. Med. 2014, 6, 1622–1637. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing (R.app GUI 1.69); R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Vaas, L.A.I.; Sikorski, J.; Hofner, B.; Fiebig, A.; Buddruhs, N.; Klenk, H.P.; Goker, M. opm: An R package for analysing OmniLog® phenotype microarray data. Bioinformatics 2013, 29, 1823–1824. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Geno-Serotype | log10 cfu/mL (Mean of n = 3) | log10 Reduction Compared to Control pH 7.8 | RpoS Protein | ||||
|---|---|---|---|---|---|---|---|---|
| Inoculum | pH 1.5 | pH 2.5 | pH 7.8 | pH 1.5 | pH 2.5 | |||
| Persistent Colonization Type (STECper): | ||||||||
| 13E0591 | O26:H11 | 7.33 | 0.52 | 7.02 | 8.23 | 7.71 | 1.21 | full-length |
| 13E0663 | O26:H11 | 7.31 | 1.00 | 6.89 | 7.83 | 6.83 | 0.95 | full-length |
| 13E0674 | O26:H11 | 7.42 | 3.91 | 6.96 | 7.93 | 4.02 | 0.97 | full-length |
| 13E0634 | O26:H11 | 7.36 | 3.43 | 6.88 | 7.95 | 4.52 | 1.07 | full-length |
| 13E0753 | O156:H25 | 7.46 | n.d. | 5.10 | 8.29 | 8.29 | 3.19 | full-length |
| 13E0780 | O156:H25 | 7.58 | n.d. | 3.35 | 8.24 | 8.24 | 4.89 | truncated (26 AS) due to an SNP at pos. 79G→T |
| 13E0725 | O182:H25 | 7.43 | n.d. | 3.17 | 8.27 | 8.27 | 5.10 | truncated (289 AS) due to a frame shift at pos. 847 (deletion of 4 nucleotides) |
| 13E0711 | O165:H25 | 7.26 | 0.82 | 5.51 | 7.77 | 6.94 | 2.25 | full-length |
| 13E0718 | O165:H25 | 7.47 | n.d. | 5.76 | 7.90 | 7.90 | 2.14 | truncated (177 AS) due to an SNP at pos. 559C→G |
| 13E0734 | O165:H25 | 6.99 | 2.90 | 4.51 | 7.40 | 4.50 | 2.90 | deleted |
| 13E0812 | O172:H25 | 7.30 | n.d. | 1.82 | 8.03 | 8.03 | 6.20 | truncated (189 AS) due to an insertion of the phage-related gene mom |
| Sporadic Colonization Type (STECspo): | ||||||||
| 13E0609 | O157:H7 | 7.32 | n.d. | 2.62 | 7.70 | 7.70 | 5.08 | deleted |
| 13E0613 | O157:H7 | 6.99 | 1.82 | 5.14 | 7.52 | 5.70 | 2.39 | truncated (70 AS) due to a 22 nucleotide insertion at pos. 170 |
| 13E0762 | O157:H7 | 7.16 | n.d. | 4.43 | 7.38 | 7.38 | 2.95 | full-length |
| 13E0793 | O157:H7 | 6.99 | n.d. | 3.97 | 7.78 | 7.78 | 3.80 | full-length |
| 13E0611 | O84:H2 | 7.51 | n.d. | 5.74 | 7.91 | 7.91 | 2.17 | full-length |
| 13E0822 | O103:H2 | 7.45 | 2.43 | 5.66 | 8.37 | 5.94 | 2.70 | full-length |
| 13E0704 | O185:H28 | 7.48 | 4.53 | 7.16 | 8.37 | 3.84 | 1.20 | full-length |
| 13E0895 | O8:HNT | 7.48 | 1.30 | 5.49 | 8.32 | 7.01 | 2.82 | deleted |
| 13E0899 | Ont:H25 | 7.39 | n.d. | 2.52 | 8.39 | 8.39 | 5.87 | 331 AS, due to a duplication of triplet GTA at pos. 307 |
| 13E0659 | O35:H2 | 7.51 | 5.00 | 7.14 | 8.20 | 3.20 | 1.06 | full-length |
| 13E0867 | O171:H2 | 7.36 | 0.52 | 5.35 | 8.22 | 7.69 | 2.87 | full-length |
| 13E0869 | O6:H49 | 7.44 | 2.32 | 5.60 | 8.05 | 5.74 | 2.45 | full-length |
| 13E0892 | O6:H49 | 7.23 | 1.60 | 6.17 | 7.98 | 6.37 | 1.81 | full-length |
| 13E0767 | O156:H8 | 7.57 | n.d. | 6.75 | 8.30 | 8.30 | 1.55 | full-length |
| Unknown Colonization Type: | ||||||||
| 12E0115 | O84:H2 | 7.41 | 6.26 | 7.37 | 7.88 | 1.62 | 0.52 | full-length |
| 12E0117 | O8:H21 | 7.52 | 2.23 | 7.22 | 7.82 | 5.59 | 0.60 | full-length |
| 12E0119 | O174:H21 | 7.13 | n.d. | 6.97 | 8.27 | 8.27 | 1.30 | deleted |
| Control Strains: | ||||||||
| EcN | O6:K5:H1 | 7.40 | n.d. | 3.70 | 8.32 | 8.32 | 4.63 | not known |
| C600 | K-12 | 6.75 | 1.94 | 3.76 | 8.27 | 6.33 | 4.50 | not known |
| Strain | Geno-Serotype | MLST 1 | Phylo-Group 2 | Selected VAGs 3 | SRA 4 Accession No. | |||
|---|---|---|---|---|---|---|---|---|
| ST Complex | ST | stx Subtype | Other Toxins | Adhesins | ||||
| Strains Representing the Persistent Colonization Type (STECper): | ||||||||
| 13E0591 | O26:H11 | ST29 Cplx | 21 | B1 | none | astA, ehxA, toxB | β-eae, efa-1, iha | SRR9972680 |
| 13E0663 | O26:H11 | ST29 Cplx | 21 | B1 | 1a | astA, ehxA | β-eae, efa-1, iha | SRR9972674 |
| 13E0674 | O26:H11 | ST29 Cplx | 21 | B1 | 1a, 2a | astA, ehxA, toxB | β-eae, efa-1, iha | SRR9972669 |
| 13E0634 | O26:H11 | ST29 Cplx | 1705 | B1 | 1a, 2a | astA, ehxA, toxB | β-eae, efa-1, iha | SRR9972676 |
| 13E0753 | O156:H25 | none | 688 | B1 | 1a | astA, ehxA | ζ-eae | SRR9972663 |
| 13E0780 | O156:H25 | none | 300 | B1 | 1a | astA, ehxA | ζ-eae | SRR9972672 |
| 13E0725 | O182:H25 | none | 300 | B1 | 1a | astA, ehxA | ζ-eae | SRR9972665 |
| 13E0711 | O165:H25 | none | 119 | A | none | none | ε-eae | SRR9972667 |
| 13E0718 | O165:H25 | none | 119 | A | 2a | astA | ε-eae, efa-1 | SRR9972668 |
| 13E0734 | O165:H25 | none | 119 | A | 2a | astA | ε-eae, efa-1 | SRR9972666 |
| 13E0812 | O172:H25 | none | 660 | A | 2a | astA | ε-eae, efa-1 | SRR9972661 |
| Strains Representing the Sporadic Colonization Type (STECspo): | ||||||||
| 13E0609 | O157:H7 | ST11 Cplx | 11 | D | 2c | astA, cdt, ehxA, toxB | γ-eae, iha | SRR9972677 |
| 13E0613 | O157:H7 | ST11 Cplx | 11 | D | 1a, 2c | astA, cdt, ehxA, toxB | γ-eae | SRR9972675 |
| 13E0762 | O157:H7 | ST11 Cplx | 11 | D | none | astA, cdt, ehxA, toxB | γ-eae, iha | SRR9972664 |
| 13E0793 | O157:H7 | ST11 Cplx | 11 | D | none | astA, cdt, ehxA, toxB | γ-eae, iha | SRR9972662 |
| 13E0611 | O84:H2 | none | 306 | B1 | 1a | astA, cdt, ehxA | ζ-eae | SRR9972678 |
| 13E0822 | O103:H2 | ST20 Cplx | 17 | B1 | 1a | ehxA | ε-eae, efa-1 | SRR9972660 |
| 13E0704 | O185:H28 | none | 658 | D | 2a | ehxA | LEE-neg., iha, saa | SRR9972670 |
| 13E0895 | O8:HNT | ST155 Cplx | 155 | D | none | none | LEE-neg. | SRR9972656 |
| 13E0899 | Ont:H25 | ST155 Cplx | 58 | B1 | 2a | astA, ehxA, subA | LEE-neg., iha, saa | SRR9972655 |
| 13E0659 | O35:H2 | none | 5266 | B1 | none | none | β-eae | SRR9972673 |
| 13E0867 | O171:H2 | none | 332 | B1 | 2d | none | LEE-neg., iha | SRR9972659 |
| 13E0869 | O6:H49 | none | 1079 | B1 | none | none | LEE-neg., iha | SRR9972658 |
| 13E0892 | O6:H49 | none | 1079 | B1 | 1a | none | LEE-neg., iha | SRR9972657 |
| 13E0767 | O156:H8 | none | 327 | B1 | none | none | θ-eae, iha | SRR9972671 |
| Strains with Unknown Colonization Type: | ||||||||
| 12E0115 | O84:H2 | none | 306 | B1 | 1a | astA, ehxA | ζ-eae | SRR9972681 |
| 12E0117 | O8:H21 | none | 1794 | B1 | 2d | astA | LEE-neg. | SRR9972682 |
| 12E0119 | O174:H21 | none | 677 | B1 | 2c | none | LEE-neg. | SRR9972679 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barth, S.A.; Weber, M.; Schaufler, K.; Berens, C.; Geue, L.; Menge, C. Metabolic Traits of Bovine Shiga Toxin-Producing Escherichia coli (STEC) Strains with Different Colonization Properties. Toxins 2020, 12, 414. https://doi.org/10.3390/toxins12060414
Barth SA, Weber M, Schaufler K, Berens C, Geue L, Menge C. Metabolic Traits of Bovine Shiga Toxin-Producing Escherichia coli (STEC) Strains with Different Colonization Properties. Toxins. 2020; 12(6):414. https://doi.org/10.3390/toxins12060414
Chicago/Turabian StyleBarth, Stefanie A., Michael Weber, Katharina Schaufler, Christian Berens, Lutz Geue, and Christian Menge. 2020. "Metabolic Traits of Bovine Shiga Toxin-Producing Escherichia coli (STEC) Strains with Different Colonization Properties" Toxins 12, no. 6: 414. https://doi.org/10.3390/toxins12060414
APA StyleBarth, S. A., Weber, M., Schaufler, K., Berens, C., Geue, L., & Menge, C. (2020). Metabolic Traits of Bovine Shiga Toxin-Producing Escherichia coli (STEC) Strains with Different Colonization Properties. Toxins, 12(6), 414. https://doi.org/10.3390/toxins12060414