Improved Antithrombotic Activity and Diminished Bleeding Side Effect of a PEGylated αIIbβ3 Antagonist, Disintegrin



Abstract
:1. Introduction
2. Results
2.1. Modification of Trimucrin Derivative RR with PEGylation
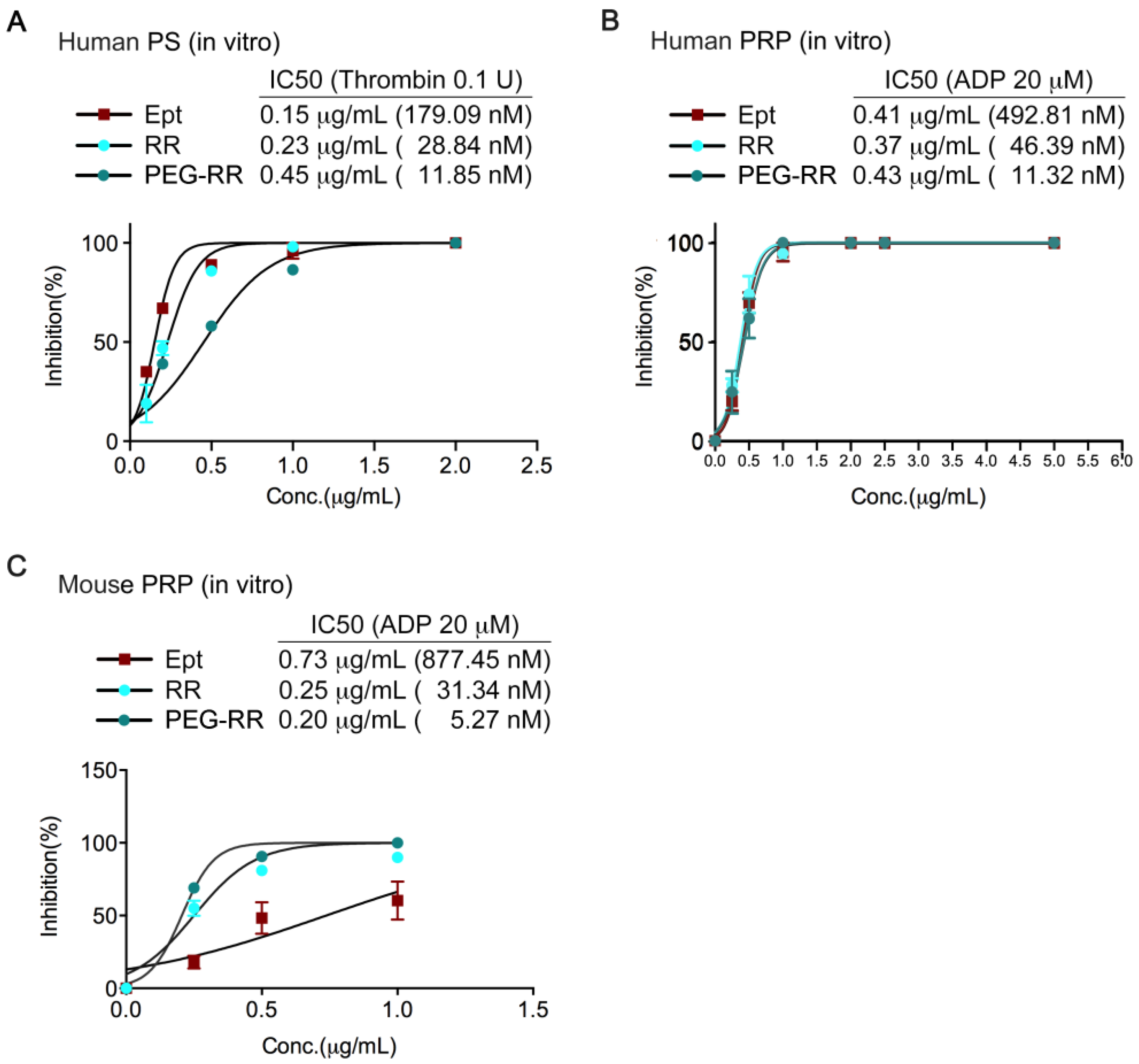
2.2. Inhibitory Efficacy of RR and PEG-RR on Platelet Aggregation
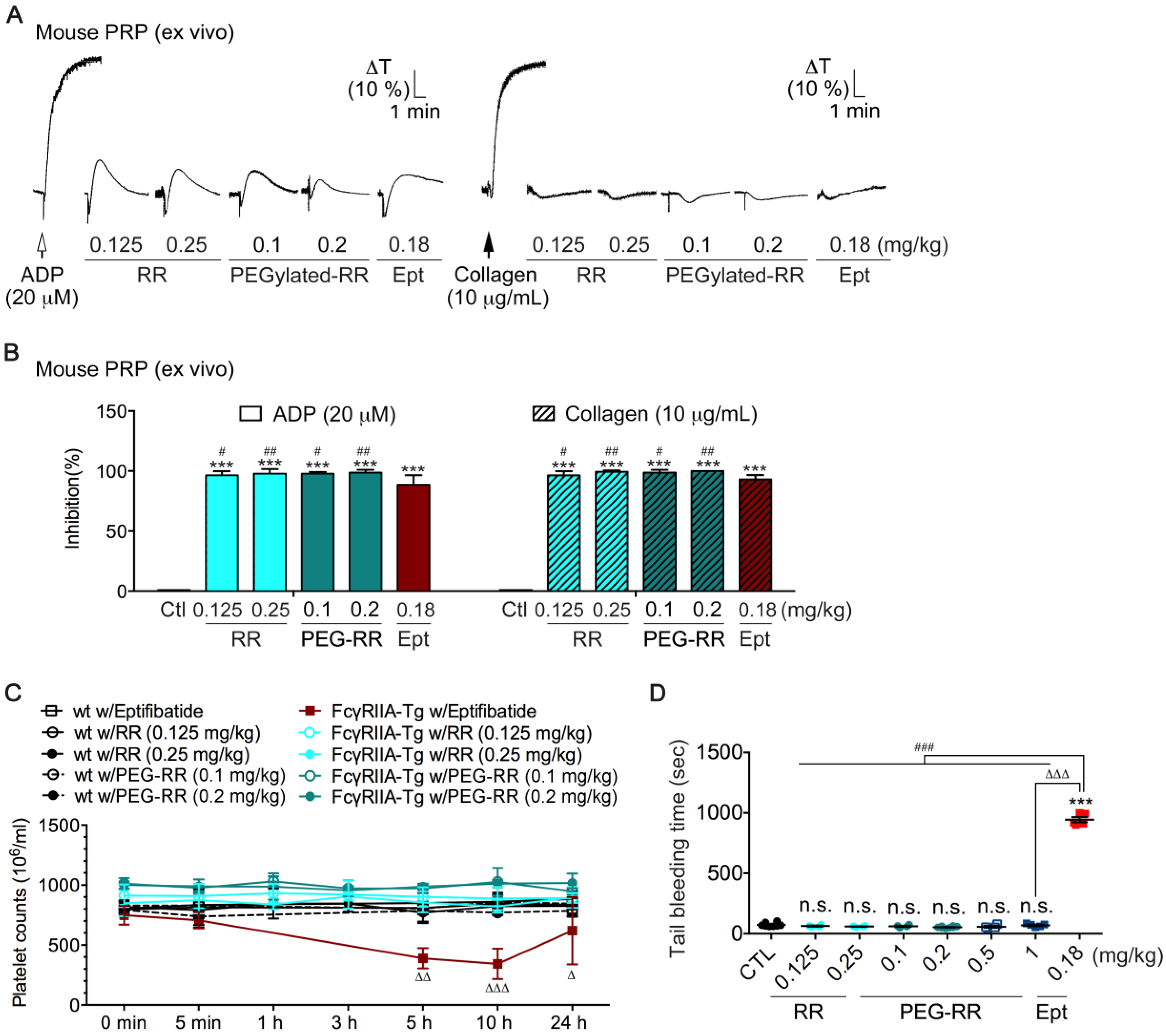
2.3. Comparison of the ex vivo Antiplatelet Activity between RR and PEG-RR
2.4. Effects of RR and PEG-RR on In Vivo Mouse Bleeding Tendency and Human Physiological Hemostatic Function in ROTEM Assays
2.5. Comparison of Anti-Thrombotic Activity between RR and PEG-RR in Illumination-Induced Thrombosis of Mouse Mesenteric Venules
2.6. In vivo Functional Half-Life (t1/2) of RR and PEG-RR in Inhibiting Platelet Aggregation
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Materials
5.2. Preparation of the PEG-RR
5.3. Mass Spectrometric Measurements
5.4. Protein Concentration
5.5. Analytical Methods
5.6. Preparation of Human PRP, PS, and Platelet Aggregation Assay
5.7. Definition of Safety Index
5.8. Rotational Thromboelastometry (ROTEM)
5.9. Ethics of Animal Experiments
5.10. In Vitro and ex vivo Mouse Platelet Aggregation
5.11. Drug Administration
5.12. Immune Clearance of Platelets in FcγRIIa-Transgenic Mice
5.13. Tail-Bleeding Time
5.14. Pharmacokinetic Studies
5.15. Irradiation-Induced Mesenteric Vessel Thrombosis Model
5.16. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ADP | Adenosine 5′-diphosphate |
| BSA | Bovine serum albumin |
| IV | Intravenous |
| PBS | Phosphate-buffered saline |
| PRP | Platelet-rich plasma |
| PS | Platelet suspension |
| RGD | Arg-Gly-Asp |
| SDS-PAGE | Sodium dodecyl sulfate-polyacrylamide gel electrophoresis |
References
- Bledzka, K.; Smyth, S.S.; Plow, E.F. Integrin alphaIIbbeta3: From discovery to efficacious therapeutic target. Circ. Res. 2013, 112, 1189–1200. [Google Scholar] [CrossRef] [Green Version]
- Xiao, T.; Takagi, J.; Coller, B.S.; Wang, J.H.; Springer, T.A. Structural basis for allostery in integrins and binding to fibrinogen-mimetic therapeutics. Nature 2004, 432, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Hynes, R.O. Integrins: Versatility, modulation, and signaling in cell adhesion. Cell 1992, 69, 11–25. [Google Scholar] [CrossRef]
- Huang, T.F.; Holt, J.C.; Lukasiewicz, H.; Niewiarowski, S. Trigramin. A low molecular weight peptide inhibiting fibrinogen interaction with platelet receptors expressed on glycoprotein IIb-IIIa complex. J. Biol. Chem. 1987, 262, 16157–16163. [Google Scholar]
- Selistre-de-Araujo, H.S.; Pontes, C.L.; Montenegro, C.F.; Martin, A.C. Snake venom disintegrins and cell migration. Toxins 2010, 2, 2606–2621. [Google Scholar] [CrossRef]
- Bassand, J.P. Current antithrombotic agents for acute coronary syndromes: Focus on bleeding risk. Int. J. Cardiol. 2013, 163, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Cook, J.J.; Huang, T.F.; Rucinski, B.; Strzyzewski, M.; Tuma, R.F.; Williams, J.A.; Niewiarowski, S. Inhibition of platelet hemostatic plug formation by trigramin, a novel RGD-peptide. Am. J. Physiol. 1989, 256, H1038–H1043. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.J.; Chen, Y.R.; Hsu, C.C.; Peng, H.C.; Huang, T.F. An alphaIIb beta3 antagonist prevents thrombosis without causing Fc receptor gamma-chain IIa-mediated thrombocytopenia. J. Thromb. Haemost. JTH 2017, 15, 2230–2244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hung, Y.C.; Kuo, Y.J.; Huang, S.S.; Huang, T.F. Trimucrin, an Arg-Gly-Asp containing disintegrin, attenuates myocardial ischemia-reperfusion injury in murine by inhibiting platelet function. Eur. J. Pharm. 2017, 813, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.M.; Chess, R.B. Effect of pegylation on pharmaceuticals. Nat. Rev. Drug Discov. 2003, 2, 214–221. [Google Scholar] [CrossRef]
- Abuchowski, A.; van Es, T.; Palczuk, N.C.; Davis, F.F. Alteration of immunological properties of bovine serum albumin by covalent attachment of polyethylene glycol. J. Biol. Chem. 1977, 252, 3578–3581. [Google Scholar] [PubMed]
- Roberts, M.J.; Bentley, M.D.; Harris, J.M. Chemistry for peptide and protein PEGylation. Adv. Drug Deliv. Rev. 2002, 54, 459–476. [Google Scholar] [CrossRef]
- Abuchowski, A.; McCoy, J.R.; Palczuk, N.C.; van Es, T.; Davis, F.F. Effect of covalent attachment of polyethylene glycol on immunogenicity and circulating life of bovine liver catalase. J. Biol. Chem. 1977, 252, 3582–3586. [Google Scholar] [PubMed]
- Jevsevar, S.; Kunstelj, M.; Porekar, V.G. PEGylation of therapeutic proteins. Biotechnol. J. 2010, 5, 113–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuo, Y.J.; Chung, C.H.; Pan, T.Y.; Chuang, W.J.; Huang, T.F. A Novel alphaIIbbeta3 Antagonist from Snake Venom Prevents Thrombosis without Causing Bleeding. Toxins 2019, 12, 11. [Google Scholar] [CrossRef] [Green Version]
- McKenzie, S.E.; Taylor, S.M.; Malladi, P.; Yuhan, H.; Cassel, D.L.; Chien, P.; Schwartz, E.; Schreiber, A.D.; Surrey, S.; Reilly, M.P. The role of the human Fc receptor Fc gamma RIIA in the immune clearance of platelets: A transgenic mouse model. J. Immunol. 1999, 162, 4311–4318. [Google Scholar] [PubMed]
- Worth, R.G.; Chien, C.D.; Chien, P.; Reilly, M.P.; McKenzie, S.E.; Schreiber, A.D. Platelet FcgammaRIIA binds and internalizes IgG-containing complexes. Exp. Hematol. 2006, 34, 1490–1495. [Google Scholar] [CrossRef]
- Ganter, M.T.; Hofer, C.K. Coagulation monitoring: Current techniques and clinical use of viscoelastic point-of-care coagulation devices. Anesth. Analg. 2008, 106, 1366–1375. [Google Scholar] [CrossRef] [Green Version]
- Lance, M.D. A general review of major global coagulation assays: Thrombelastography, thrombin generation test and clot waveform analysis. Thromb. J. 2015, 13, 1. [Google Scholar] [CrossRef] [Green Version]
- Greineder, C.F.; Howard, M.D.; Carnemolla, R.; Cines, D.B.; Muzykantov, V.R. Advanced drug delivery systems for antithrombotic agents. Blood 2013, 122, 1565–1575. [Google Scholar] [CrossRef]
- Hsu, C.C.; Chuang, W.J.; Chung, C.H.; Chang, C.H.; Peng, H.C.; Huang, T.F. Improved antithrombotic activity and diminished bleeding side effect of a PEGylated alphaIIbbeta3 antagonist, disintegrin. Thromb. Res. 2016, 143, 3–10. [Google Scholar] [CrossRef]
- Harris, J.M.; Martin, N.E.; Modi, M. Pegylation: A novel process for modifying pharmacokinetics. Clin. Pharm. 2001, 40, 539–551. [Google Scholar] [CrossRef]
- Swieringa, F.; Kuijpers, M.J.; Heemskerk, J.W.; van der Meijden, P.E. Targeting platelet receptor function in thrombus formation: The risk of bleeding. Blood Rev. 2014, 28, 9–21. [Google Scholar] [CrossRef]
- Kuo, Y.J.; Chung, C.H.; Huang, T.F. From Discovery of Snake Venom Disintegrins to A Safer Therapeutic Antithrombotic Agent. Toxins 2019, 11, 372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McFadyen, J.D.; Peter, K. Novel Antithrombotic Drugs on the Horizon: The Ultimate Promise to Prevent Clotting While Avoiding Bleeding. Circ. Res. 2017, 121, 1133–1135. [Google Scholar] [CrossRef] [PubMed]
- Ley, K.; Rivera-Nieves, J.; Sandborn, W.J.; Shattil, S. Integrin-based therapeutics: Biological basis, clinical use and new drugs. Nat. Rev. Drug Discov. 2016, 15, 173–183. [Google Scholar] [CrossRef] [Green Version]
- Shen, B.; Zhao, X.; O’Brien, K.A.; Stojanovic-Terpo, A.; Delaney, M.K.; Kim, K.; Cho, J.; Lam, S.C.; Du, X. A directional switch of integrin signalling and a new anti-thrombotic strategy. Nature 2013, 503, 131–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Negri, A.; Li, J.; Naini, S.; Coller, B.S.; Filizola, M. Structure-based virtual screening of small-molecule antagonists of platelet integrin alphaIIbbeta3 that do not prime the receptor to bind ligand. J. Comput. Aided Mol. Des. 2012, 26, 1005–1015. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Vootukuri, S.; Shang, Y.; Negri, A.; Jiang, J.K.; Nedelman, M.; Diacovo, T.G.; Filizola, M.; Thomas, C.J.; Coller, B.S. RUC-4: A novel alphaIIbbeta3 antagonist for prehospital therapy of myocardial infarction. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 2321–2329. [Google Scholar] [CrossRef] [Green Version]
- Van Agthoven, J.F.; Xiong, J.P.; Alonso, J.L.; Rui, X.; Adair, B.D.; Goodman, S.L.; Arnaout, M.A. Structural basis for pure antagonism of integrin alphaVbeta3 by a high-affinity form of fibronectin. Nat. Struct. Mol. Biol. 2014, 21, 383–388. [Google Scholar] [CrossRef] [Green Version]
- Akbarzadehlaleh, P.; Mirzaei, M.; Mashahdi-Keshtiban, M.; Shamsasenjan, K.; Heydari, H. PEGylated Human Serum Albumin: Review of PEGylation, Purification and Characterization Methods. Adv. Pharm. Bull. 2016, 6, 309–317. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.F.; Chang, C.H.; Ho, P.L.; Chung, C.H. FcgammaRII mediates platelet aggregation caused by disintegrins and GPIIb/IIIa monoclonal antibody, AP2. Exp. Hematol. 2008, 36, 1704–1713. [Google Scholar] [CrossRef]
- Chang, C.H.; Chung, C.H.; Kuo, H.L.; Hsu, C.C.; Huang, T.F. The highly specific platelet glycoprotein (GP) VI agonist trowaglerix impaired collagen-induced platelet aggregation ex vivo through matrix metalloproteinase-dependent GPVI shedding. J. Thromb. Haemost. JTH 2008, 6, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.C.; Wu, W.B.; Chang, Y.H.; Kuo, H.L.; Huang, T.F. Antithrombotic effect of a protein-type I class snake venom metalloproteinase, kistomin, is mediated by affecting glycoprotein Ib-von Willebrand factor interaction. Mol. Pharmacol. 2007, 72, 984–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, M.C.; Huang, T.F. In vivo effect of a thrombin-like enzyme on platelet plug formation induced in mesenteric microvessels of mice. Thromb. Res. 1994, 73, 31–38. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antithrombotic Agents | Trimucrin | RR | PEG-RR | Tirofiban | Eptifibatide | Control | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| IC50 (nM) | 58.51 | 36.9 | 8.6 | 85.1 | 625.0 | N/A | |||||
| Safety index | 23.4 | >3365.9 | >3071.3 | 4.0 | 3.6 | N/A | |||||
| Fold of IC50 | 4× | 20× | 4× | 20× | 4× | 20× | 4× | 20× | 4× | 20× | N/A |
| Clotting time (sec) | 200.0 (NS) | 224.3 (NS) | 204.1 (NS) | 221.5 (NS) | 201.3 (NS) | 196.8 (NS) | 263.3 (**) | 268.3 (***) | 246.0 (*) | 251.3 (*) | 202.1 |
| Clot formation time (sec) | 96.6 (NS) | 102.7 NS) | 91.5 (NS) | 101.9 (NS) | 86.7 (NS) | 93.1 (NS) | 235.7 (***) | 341.5 (***) | 248.3 (***) | 309.2 (***) | 87.2 |
| Maximum clot firmness (mm) | 59.8 (NS) | 55.7 (NS) | 61.3 (NS) | 57.4 (NS) | 61.3 (NS) | 60.5 (NS) | 50.7 (**) | 44.3 (***) | 52.3 (*) | 47.3 (***) | 60.9 |
| Tail-bleeding time (sec) | 95.0 (NS) | 190.2 (*) | 63.5 (NS) | 68.4 (NS) | 66.2 (NS) | 66.3 (NS) | 480.7 (***) | 836.2 (***) | 538.6 (***) | 799.5 (***) | 66.9 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuo, Y.-J.; Chang, Y.T.; Chung, C.-H.; Chuang, W.-J.; Huang, T.-F. Improved Antithrombotic Activity and Diminished Bleeding Side Effect of a PEGylated αIIbβ3 Antagonist, Disintegrin. Toxins 2020, 12, 426. https://doi.org/10.3390/toxins12070426
Kuo Y-J, Chang YT, Chung C-H, Chuang W-J, Huang T-F. Improved Antithrombotic Activity and Diminished Bleeding Side Effect of a PEGylated αIIbβ3 Antagonist, Disintegrin. Toxins. 2020; 12(7):426. https://doi.org/10.3390/toxins12070426
Chicago/Turabian StyleKuo, Yu-Ju, Yao Tsung Chang, Ching-Hu Chung, Woei-Jer Chuang, and Tur-Fu Huang. 2020. "Improved Antithrombotic Activity and Diminished Bleeding Side Effect of a PEGylated αIIbβ3 Antagonist, Disintegrin" Toxins 12, no. 7: 426. https://doi.org/10.3390/toxins12070426
APA StyleKuo, Y. -J., Chang, Y. T., Chung, C. -H., Chuang, W. -J., & Huang, T. -F. (2020). Improved Antithrombotic Activity and Diminished Bleeding Side Effect of a PEGylated αIIbβ3 Antagonist, Disintegrin. Toxins, 12(7), 426. https://doi.org/10.3390/toxins12070426