Risk Assessment and the Effects of Refuge Availability on the Defensive Behaviors of the Southern Unstriped Scorpion (Vaejovis carolinianus)



Abstract
:1. Introduction
2. Results
2.1. General Use of Defensive Behaviors
2.2. Results of Mixed Model Analysis
2.2.1. Cycle
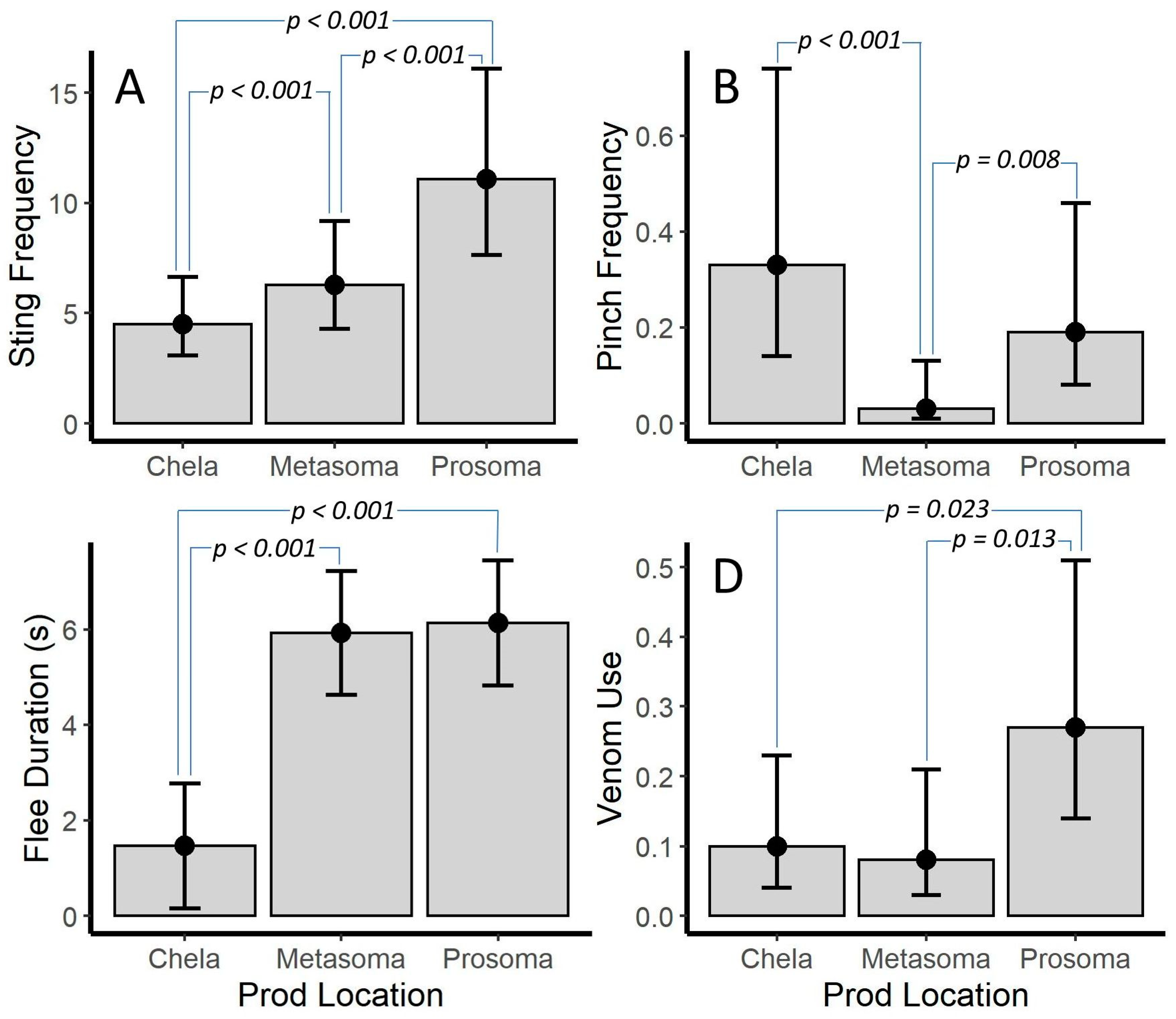
2.2.2. Prod Location
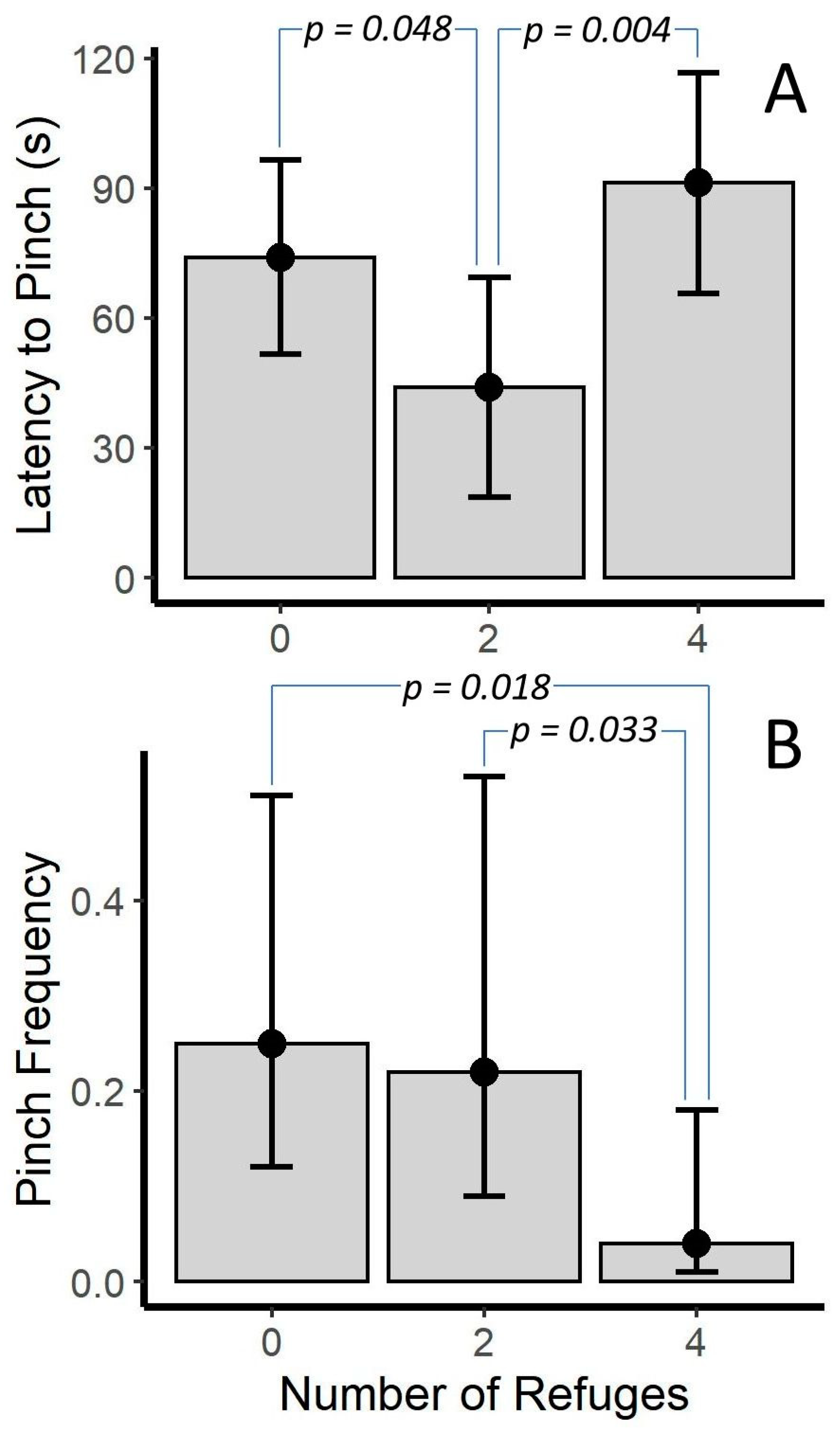
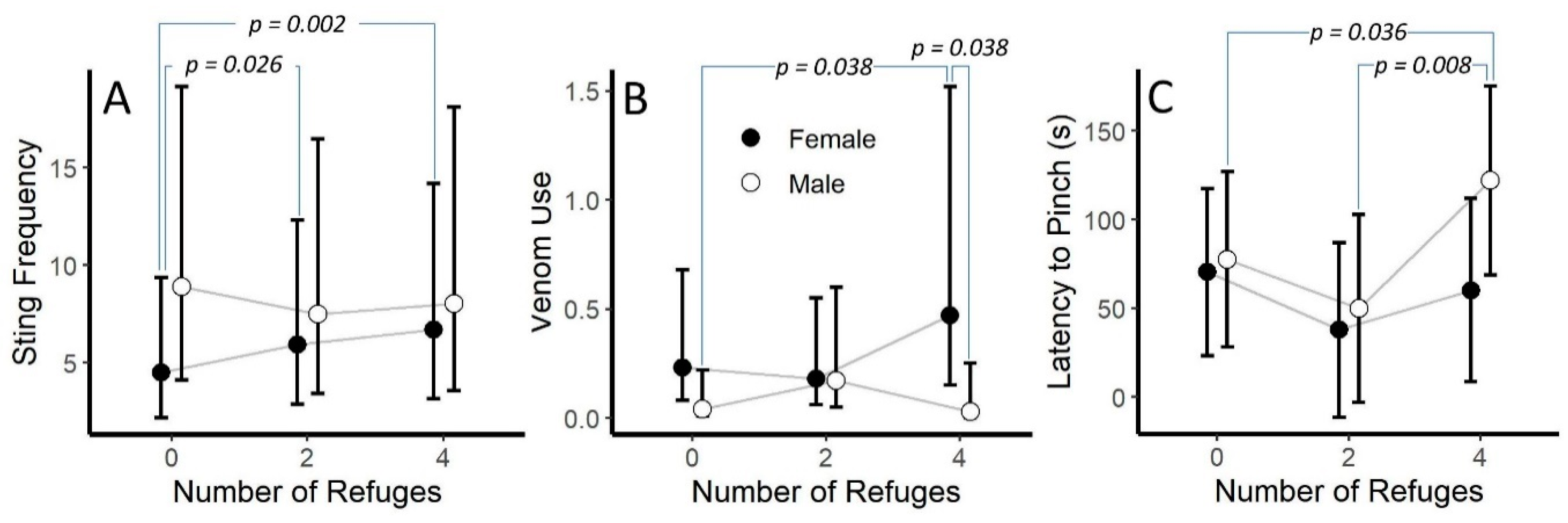
2.2.3. Number of Refuges and Sex
2.3. Covariate Controls
3. Discussion
3.1. Risk Assessment Based on Prod Location
3.2. Venom Use and Associated Behaviors in the Context of Other Defensive Behaviors
3.3. Changes in Defensive Behavior across Cycle
3.4. Presence and Number of Refuges
4. Conclusions
5. Materials and Methods
5.1. Collection
5.2. Care
5.3. Testing Arenas for V. carolinianus Risk Assesment Trails: (A) no, (B) Two, and (C) Four Refuge Condition
5.4. Experimental Randomization/Standardization
5.5. Exploration of Arenas
5.6. Simulated Predatory Encounter
5.7. Data Abstraction
5.7.1. Recordings of Overnight Movements
5.7.2. Recordings of Defensive Behaviors
5.8. Statistical Methods
5.8.1. General Analysis and Order of Defensive Behaviors
5.8.2. Behavioral Analysis Using Linear and Generalized Liner Mixed Models
5.8.3. General Statistical Conventions and Statistical Software
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McFarland, D.J. Decision making in animals. Nature 1977, 269, 15–21. [Google Scholar] [CrossRef]
- Dukas, R.; Ratcliffe, J.M. Cognitive Ecology II; University of Chicago Press: Chicago, IL, USA, 2009; ISBN 9780226169378. [Google Scholar]
- Janetos, A.C.; Cole, B.J. Imperfectly optimal animals. Behav. Ecol. Sociobiol. 1981, 9, 203–209. [Google Scholar] [CrossRef]
- Owen, M.A.; Swaisgood, R.R.; Blumstein, D.T. Contextual influences on animal decision-making: Significance for behavior-based wildlife conservation and management. Integr. Zool. 2017, 12, 32–48. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, D.C.; Griebel, G.; Pobbe, R.; Blanchard, R.J. Risk assessment as an evolved threat detection and analysis process. Neurosci. Biobehav. Rev. 2011, 35, 991–998. [Google Scholar] [CrossRef]
- Hayes, W.K. The Snake Venom-Metering Controversy: Levels of Analysis, Assumptions, and Evidence. In The Biology of Rattlesnakes; Loma Linda University Press: Loma Linda, CA, USA, 2008; pp. 1–30. [Google Scholar]
- Wigger, E.; Kuhn-Nentwig, L.; Nentwig, W. The venom optimisation hypothesis: A spider injects large venom quantities only into difficult prey types. Toxicon 2002, 40, 749–752. [Google Scholar] [CrossRef]
- Morgenstern, D.; King, G.F. The venom optimization hypothesis revisited. Toxicon 2013, 63, 120–128. [Google Scholar] [CrossRef]
- Cooper, A.M.; Nelsen, D.R.; Hayes, W.K. The Strategic Use of Venom by Spiders. In Evolution of Venomous Animals and Their Toxins; Gopalakrishnakone, P., Malhotra, A., Eds.; Springer: Dordrecht, The Netherlands, 2015; pp. 1–18. ISBN 978-94-007-6727-0. [Google Scholar]
- Evans, E.R.J.; Northfield, T.D.; Daly, N.L.; Wilson, D.T. Venom costs and optimization in scorpions. Front. Ecol. Evol. 2019, 7, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Hayes, W.K. Effects of hunger on striking, prey-handling, and venom expenditure of Prairie Rattlesnakes (Crotalus v. viridis). Herpetologica 1993, 49, 305–310. [Google Scholar]
- Hostettler, S.; Nentwig, W. Olfactory information saves venom during prey-capture of the hunting spider Cupiennius salei (Araneae: Ctenidae). Funct. Ecol. 2006, 20, 369–375. [Google Scholar] [CrossRef]
- Wullschleger, B.; Nentwig, W. Influence of venom availability on a spider’s prey-choice behaviour. Funct. Ecol. 2002, 16, 802–807. [Google Scholar] [CrossRef]
- Boevé, J.-L.J. Injection of venom into an insect prey by the free hunting spider Cupiennius salei (Araneae, Ctenidae). J. Zool. 1994, 234, 165–175. [Google Scholar] [CrossRef]
- Hayes, W.K.; Lavin-Murcio, P.; Kardong, K.V. Northern Pacific rattlesnakes (Crotalus viridis oreganus) meter venom when feeding on prey of different sizes. Copeia 1995, 2, 337–343. [Google Scholar] [CrossRef]
- Hayes, W.K.; Herbert, S.S.; Rehling, G.C.; Gennaro, J.F. Factors that influence venom expenditure in viperids and other snake species during predatory and defensive contexts. In Biology of the Vipers; Greene, H.W., Ed.; Eagle Mountain Publishing: Eagle Mountain, UT, USA, 1996; pp. 207–233. [Google Scholar]
- Malli, H.; Imboden, H.; Kuhn-Nentwig, L. Quantifying the venom dose of the spider Cupiennius salei using monoclonal antibodies. Toxicon 1998, 36, 1959–1969. [Google Scholar] [CrossRef]
- Malli, H.; Kuhn-Nentwig, L.; Imboden, H.; Nentwig, W.; Berne, C.; Neurobiology, D. Effects of size, motility and paralysation time of prey on the quantity of venom injected by the hunting spider Cupiennius salei. J. Exp. Biol. 1999, 202, 2083–2089. [Google Scholar] [PubMed]
- Rodriguez-Robles, J.A.; Leal, M. Effects of Prey Type on the Feeding Behavior of Alsophis portoricensis (Serpentes: Colubridae). J. Herpetol. 1993, 27, 163. [Google Scholar] [CrossRef]
- Boevé, J.-L.L.; Kuhn-Nentwig, L.; Keller, S.; Nentwig, W. Quantity and quality of venom released by a spider (Cupiennius salei, Ctenidae). Toxicon 1995, 33, 1347–1357. [Google Scholar] [CrossRef]
- Hayes, W.K. Factors associated with the mass of venom expended by prairie rattlesnakes (Crotalus v. viridis) feeding on mice. Toxicon 1992, 30, 449–460. [Google Scholar] [CrossRef]
- Carlin, N.F.; Gladstein, D.S. The “Bouncer” Defense of Odontomachus Ruginodis and Other Odontomachine Ants (Hymenoptera: Formicidae). Psyche 1989, 96, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Fink, L.L.S. Venom spitting by the green lynx spider, Peucetia viridans (Araneae, Oxyopidae). J. Arachnol. 1984, 12, 372–373. [Google Scholar]
- Haight, K.L. Defensiveness of the fire ant, Solenopsis invicta, is increased during colony rafting. Insectes Soc. 2006, 53, 32–36. [Google Scholar] [CrossRef]
- Nisani, Z.; Hayes, W.K. Defensive stinging by Parabuthus transvaalicus scorpions: Risk assessment and venom metering. Anim. Behav. 2011, 81, 627–633. [Google Scholar] [CrossRef]
- Nelsen, D.R.; Kelln, W.; Hayes, W.K. Poke but don’t pinch: Risk assessment and venom metering in the western black widow spider, Latrodectus hesperus. Anim. Behav. 2014, 89, 107–114. [Google Scholar] [CrossRef]
- Cushing, B.S.; Matherne, A. Stinger utilization and predation in the scorpion Paruroctonus boreus. Great Basin Nat. 1980, 40, 193–195. [Google Scholar]
- Casper, G. Prey capture and stinging behavior in the emperor scorpion, Pandinus imperator (Koch) (Scorpiones, Scorpionidae). J. Arachnol. 1985, 13, 277–283. [Google Scholar]
- Rein, J. Sting use in two species of Parabuthus Scorpions (Buthidae). J. Arachnol. 1993, 21, 60–63. [Google Scholar] [CrossRef]
- Edmunds, M.C.; Sibly, R.M. Optimal sting use in the feeding behavior of the scorpion Hadrurus spadix. J. Arachnol. 2010, 38, 123–125. [Google Scholar] [CrossRef] [Green Version]
- Simone, Y.; Garcia, L.F.; Lacava, M.; van der Meijden, A.; Viera, C. Predatory versatility in females of the scorpion Bothriurus bonariensis (Scorpiones: Bothriuridae): Overcoming prey with different defensive mechanisms. J. Insect Behav. 2018, 31, 402–415. [Google Scholar] [CrossRef]
- Carlson, B.E.; McGinley, S.; Rowe, M.P. Meek males and fighting females: Sexually-dimorphic antipredator behavior and locomotor performance is explained by morphology in bark scorpions (Centruroides vittatus). PLoS ONE 2014, 9, e97648. [Google Scholar] [CrossRef] [Green Version]
- Miller, D.W.; Jones, A.D.; Goldston, J.S.; Rowe, M.P.; Rowe, A.H. Sex differences in defensive behavior and venom of the striped bark scorpion Centruroides vittatus (Scorpiones: Buthidae). Integr. Comp. Biol. 2016, 56, 1022–1031. [Google Scholar] [CrossRef] [Green Version]
- Lira, A.F.A.; Santos, A.B.; Silva, N.A.; Martins, R.D. Threat level influences the use of venom in a scorpion species, Tityus stigmurus (Scorpiones, Buthidae). Acta Ethol. 2017, 20, 291–295. [Google Scholar] [CrossRef]
- Silva, M.A.; Silva, N.A.; Lira, A.F.A.; Martins, R.D. Role of venom quantity in the feeding behavior of Jaguajir rochae (Scorpiones: Buthidae). Acta Ethol. 2019, 99–104. [Google Scholar] [CrossRef]
- Nisani, Z.; Hayes, W.K. Venom-spraying behavior of the scorpion Parabuthus transvaalicus (Arachnida: Buthidae). Behav. Process. 2015, 115, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Nisani, Z.; Curiel, R. Antipredator responses of Hadrurus arizonensis (Scorpiones: Caraboctonidae) to chemosensory cue from a mammalian predator. J. Arachnol. 2019, 47, 389. [Google Scholar] [CrossRef]
- Rasko, M.; Coelho, P.; Simone, Y.; van der Meijden, A. How to attack a scorpion: Venom metering during a repeated attack. Anim. Behav. 2018, 145, 125–129. [Google Scholar] [CrossRef]
- Glaudas, X.; Farrell, T.M.; May, P.G.; Glaudas, X.; Farrell, T.M.; May, P.G. Defensive behavior of free-ranging pygmy rattlesnakes (Sistrurus miliarius). Copeia 2005, 2005, 196–200. [Google Scholar] [CrossRef]
- Glaudas, X.; Winne, C.T.; Fedewa, L.A. Ontogeny of anti-predator behavioral habituation in cottonmouths (Agkistrodon piscivorus). Ethology 2006, 112, 608–615. [Google Scholar] [CrossRef]
- Glaudas, X.; Winne, C.T. Do warning displays predict striking behavior in a viperid snake, the cottonmouth (Agkistrodon piscivorus)? Can. J. Zool. 2007, 85, 574–578. [Google Scholar] [CrossRef]
- Llewelyn, J.; Webb, J.K.; Shine, R. Flexible defense: Context-dependent antipredator responses of two species of australian elapid snakes. Herpetologica 2010, 66, 1–11. [Google Scholar] [CrossRef]
- Carlson, B.E.; Rowe, M.P. Temperature and desiccation effects on the antipredator behavior of Centruroides vittatus (Scorpiones: Buthidae). J. Arachnol. 2009, 37, 321–330. [Google Scholar] [CrossRef]
- Van der Meijden, A.; Lobo Coelho, P.; Sousa, P.; Herrel, A. Choose your weapon: Defensive behavior is associated with morphology and performance in scorpions. PLoS ONE 2013, 8, e78955. [Google Scholar] [CrossRef] [Green Version]
- Lira, A.F.A.; Almeida, F.M.F.; Albuquerque, C.M.R. Reaction under the risk of predation: Effects of age and sexual plasticity on defensive behavior in scorpion Tityus pusillus (Scorpiones: Buthidae). J. Ethol. 2020, 38, 13–19. [Google Scholar] [CrossRef]
- Domenici, P. Context-dependent variability in the components of fish escape response: Integrating locomotor performance and behavior. J. Exp. Zool. Part A Ecol. Genet. Physiol. 2010, 313, 59–79. [Google Scholar] [CrossRef] [PubMed]
- Krochmal, A.R.; Bakken, G.S. Thermoregulation is the pits: Use of thermal radiation for retreat site selection by rattlesnakes. J. Exp. Biol. 2003, 206, 2539–2545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collett, T.S.; Graham, P. Animal navigation: Path integration, visual landmarks and cognitive maps. Curr. Biol. 2004, 14, 475–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baddeley, B.; Graham, P.; Husbands, P.; Philippides, A. A model of ant route navigation driven by scene familiarity. PLoS Comput. Biol. 2012, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaffin, D.D.; Brayfield, B.P. Exploring the chemo-textural familiarity hypothesis for scorpion navigation. J. Arachnol. 2017, 45, 265–270. [Google Scholar] [CrossRef]
- Mattoni, C.I.; García-Hernández, S.; Botero-Trujillo, R.; Ochoa, J.A.; Ojanguren-Affilastro, A.A.; Pinto-da-Rocha, R.; Prendini, L. Scorpion sheds “tail” to escape: Consequences and implications of autotomy in scorpions (Buthidae: Ananteris). PLoS ONE 2015, 10, e0116639. [Google Scholar] [CrossRef] [Green Version]
- Stewart, D.M.; Martin, A.W. Blood pressure in the tarantula, Dugesiella hentzi. J. Comp. Physiol. 1974, 88, 141–172. [Google Scholar] [CrossRef]
- Foelix, R. Biology of Spiders; Oxford University Press: Oxford, UK, 2011; ISBN 9780199734825. [Google Scholar]
- Polis, G.A.G. The Biology of Scorpions; Polis, G.A., Ed.; Stanford University Press: Stanford, CA, USA, 1990. [Google Scholar]
- Stockmann, R. Introduction to Scorpion Biology and Ecology. In Scorpion Venoms: Scorpion Venoms; Gopalakrishnakone, P., Ferroni Schwartz, E., Possani, L.D., de la Vega, R.C., Eds.; Springer: Dordrecht, The Netherlands, 2015; pp. 1–29. ISBN 978-94-007-6647-1. [Google Scholar]
- Elston, D.M. What’s eating you? Pain eastern stripeless scorpion (Vaejovis carolinianus). Cutis Cutan. Med. Pract. 2006, 78, 393–394. [Google Scholar]
- Kang, A.M.; Brooks, D.E. Nationwide scorpion exposures reported to us poison control centers from 2005 to 2015. J. Med. Toxicol. 2017, 13, 158–165. [Google Scholar] [CrossRef]
- McCue, M.D. Cost of producing venom in three North American pitviper species. Copeia 2006, 818–825. [Google Scholar] [CrossRef]
- Nisani, Z.; Dunbar, S.G.; Hayes, W.K. Cost of venom regeneration in Parabuthus transvaalicus (Arachnida: Buthidae). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2007, 147, 509–513. [Google Scholar] [CrossRef] [PubMed]
- Nisani, Z.; Boskovic, D.S.; Dunbar, S.G.; Kelln, W.; Hayes, W.K. Investigating the chemical profile of regenerated scorpion (Parabuthus transvaalicus) venom in relation to metabolic cost and toxicity. Toxicon 2012, 60, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Pintor, A.F.V.; Krockenberger, A.K.; Seymour, J.E. Costs of venom production in the common death adder (Acanthophis antarcticus). Toxicon 2010, 56, 1035–1042. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.T.; Ortega, J.; Beaupre, S.J. Metabolic cost of venom replenishment by prairie rattlesnakes (Crotalus viridis viridis). Toxicon 2014, 86, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Sih, A.; Bell, A.M.; Johnson, J.C.; Ziemba, R.E. Behavioral syndromes: An integrative overview. Q. Rev. Biol. 2004, 79, 241–277. [Google Scholar] [CrossRef] [PubMed]
- Herbert, S.S.; Hayes, W.K. Denim clothing reduces venom expenditure by rattlesnakes striking defensively at model human limbs. Ann. Emerg. Med. 2009, 54, 830–836. [Google Scholar] [CrossRef] [PubMed]
- Inceoglu, B.; Lango, J.; Jing, J.; Chen, L.; Doymaz, F.; Pessah, I.N.; Hammock, B.D. One scorpion, two venoms: Prevenom of Parabuthus transvaalicus acts as an alternative type of venom with distinct mechanism of action. Proc. Natl. Acad. Sci. USA 2003, 100, 922–927. [Google Scholar] [CrossRef] [Green Version]
- Rowe, A.H.; Rowe, M.P. Risk assessment by grasshopper mice (Onychomys spp.) feeding on neurotoxic prey (Centruroides spp.). Anim. Behav. 2006, 71, 725–734. [Google Scholar] [CrossRef]
- Brown, C.A.; O’Connell, D.J. Plant climbing behavior in the scorpion Centruroides vittatus. Am. Midl. Nat. 2000, 144, 406–418. [Google Scholar] [CrossRef]
- Baggett, B.A. Preferred Soil Moisture Content, and Aggregation Tendency in Female Vaejovis carolinianus Populations in Tennessee’s Upper Cumberland Region; Tennessee Technological University: Cookeville, TN, USA, 2015. [Google Scholar]
- Vinnedge, J.E.; Gaffin, D.D. Determination of in-lab site fidelity and movement patterns of Paruroctonus utahensis (Scorpiones: Vaejovidae). J. Arachnol. 2015, 43, 54–58. [Google Scholar] [CrossRef]
- Rasband, W.S. ImageJ (Version 1.53a); U.S. National Institutes of Health: Bethesda, MD, USA, 2018. Available online: https://imagej.nih.gov/ij/ (accessed on 21 July 2020).
- The Jamovi Project (Version 1.2). 2020. Available online: https://www.jamovi.org (accessed on 29 June 2020).
- Gallucci, M. GAMLj: General Analyses for Linear Models. [Jamovi Module]. 2019. Available online: https://gamlj.github.io/ (accessed on 29 June 2020).
- Russell Length. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. R Package Version 1.4.8. 2020. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 3 August 2020).
- Nakagawa, S.; Schielzeth, H. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Nakagawa, S. A farewell to Bonferroni: The problems of low statistical power and publication bias. Behav. Ecol. 2004, 15, 1044–1045. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing, Version 3.6.1; 2019. Available online: https://cran.r-project.org/ (accessed on 1 July 2020).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Terminology | Definitions |
|---|---|
| Sting | Metasoma movement directed at the stimulus that included the extension of the last metasoma segment so that the telson struck the target, or if the target had moved, where the target had been. |
| Venom Use | A droplet of venom observed at the tip of the telson. If the droplet grew larger during a successive sting, then this was counted as a new instance of venom use. |
| Pinch | Open and closure of a chela or chelae around the stimulus. |
| Flee * | Movement in response to a prod from the stimulus. Fleeing events were measured from the first sign of limb movement until the scorpion became and remained motionless for several frames. |
| Cycle | Unit of experimental stimulus used to elicit defensive behavior. Each cycle consisted of 10 brief prods to the chelae, 10 prods to the metasoma, and then 10 prods to the prosoma. Each cycle lasted 30 s and always followed the same order: chelae, metasoma, then prosoma. |
| Mean movement | The average speed of the scorpion as it moved around the arena during overnight acclimation. |
| Sting Frequency 1 | Pinch Frequency 1 | Venom Use 1 | Flee Duration 2 | |||||
|---|---|---|---|---|---|---|---|---|
| χ2(df) | p | χ2(df) | p | χ2(df) | p | F(df) | p | |
| Cycle | 31.56(2) | <0.001 | 1.58(2) | 0.455 | 2.43(2) | 0.296 | 112.94(2) | <0.001 |
| R2 Marginal | 0.03 | 0.01 | 0.02 | 0.07 | ||||
| R2 Conditional | 0.86 | 0.36 | 0.34 | 0.48 | ||||
| Sting Frequency 1 | Pinch Frequency 1 | Venom Use 1 | Flee Duration 2 | Latency to Sting 2 | Latency to Pinch 2 | Latency to Flee 2 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| χ2(df) | p | χ2(df) | p | χ2(df) | p | F(df) | p | F(df) | p | F(df) | p | F(df) | p | |
| Prod Location | 167.27(2) | <0.001 | 14.74(2) | <0.001 | 9.03(2) | 0.011 | 25.36(2) | <0.001 | - | - | - | - | - | - |
| # of Refuges | 2.13(2) | 0.344 | 5.63(2) | 0.06 | 1.56(2) | 0.459 | 0.55(2) | 0.578 | 1.00(2) | 0.38 | 7.04(2) | 0.007 | 0.38(2) | 0.687 |
| Sex | 0.29(1) | 0.588 | 0.01(1) | 0.909 | 2.88(1) | 0.09 | 0.29(1) | 0.596 | 0.25(1) | 0.622 | 0.44(1) | 0.516 | 2.10(1) | 0.115 |
| Refuges × Sex | 11.66(2) | 0.003 | 1.88(2) | 0.39 | 6.07(2) | 0.048 | 1.54(2) | 0.218 | 0.52(2) | 0.602 | 3.04(2) | 0.073 | 0.49(2) | 0.617 |
| Movement | 17.64(1) | <0.001 | 1.09(1) | 0.297 | 6.84(1) | 0.009 | 0.07(1) | 0.797 | 5.41(1) | 0.026 | 0.47(1) | 0.499 | 0.50(1) | 0.486 |
| Length | 0.63(1) | 0.426 | 0.10(1) | 0.749 | 0.01(1) | 0.927 | 0.09(1) | 0.764 | 1.00(1) | 0.333 | 0.02(1) | 0.881 | 2.04(1) | 0.161 |
| Time Observed | 0.67(1) | 0.414 | 2.49(1) | 0.114 | 0.41(1) | 0.521 | 0.59(1) | 0.218 | - | - | - | - | - | - |
| R2 Marginal | 0.36 | 0.5 | 0.46 | 0.27 | 0.16 | 0.25 | 0.07 | |||||||
| R2 Conditional | 0.84 | 0.59 | 0.47 | 0.39 | 0.44 | 0.68 | 0.08 | |||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nelsen, D.R.; David, E.M.; Harty, C.N.; Hector, J.B.; Corbit, A.G. Risk Assessment and the Effects of Refuge Availability on the Defensive Behaviors of the Southern Unstriped Scorpion (Vaejovis carolinianus). Toxins 2020, 12, 534. https://doi.org/10.3390/toxins12090534
Nelsen DR, David EM, Harty CN, Hector JB, Corbit AG. Risk Assessment and the Effects of Refuge Availability on the Defensive Behaviors of the Southern Unstriped Scorpion (Vaejovis carolinianus). Toxins. 2020; 12(9):534. https://doi.org/10.3390/toxins12090534
Chicago/Turabian StyleNelsen, David R., Emily M. David, Chad N. Harty, Joseph B. Hector, and Aaron G. Corbit. 2020. "Risk Assessment and the Effects of Refuge Availability on the Defensive Behaviors of the Southern Unstriped Scorpion (Vaejovis carolinianus)" Toxins 12, no. 9: 534. https://doi.org/10.3390/toxins12090534
APA StyleNelsen, D. R., David, E. M., Harty, C. N., Hector, J. B., & Corbit, A. G. (2020). Risk Assessment and the Effects of Refuge Availability on the Defensive Behaviors of the Southern Unstriped Scorpion (Vaejovis carolinianus). Toxins, 12(9), 534. https://doi.org/10.3390/toxins12090534