1. Introduction
Carukia barnesi was the first of many cubozoan jellyfish known to cause Irukandji syndrome [
1,
2]. While Huynh et al. [
3] showed that it was the principal causative animal in victims demonstrating Irukandji syndrome in Cairns, Australia, other regions of the tropical and subtropical world have identified other jellyfish species. Irukandji syndrome is typified by victims experiencing an innocuous sting while swimming in the ocean followed by delayed truncal pain, hypertension, tachycardia, and agitation among other symptoms and signs of catecholamine excess.
Although most victims settle over hours with supportive treatment, some victims deteriorate and develop severe Irukandji syndrome necessitating intensive care management of life-threatening cardiac failure and intracerebral haemorrhage [
3,
4,
5]. The toxic mechanisms involved remain largely speculative, including the general view that the venom is unlikely to have a direct acting cardiac toxin and instead acts to promote the release of endogenous catecholamines through which the symptoms and signs can be explained. This is indirectly supported by in vivo and in vitro research [
6,
7,
8,
9].
Morphological descriptions of
C. barnesi include characteristic clumps of nematocysts on the bell (called mammillations or warts) and on the tentacles (neckerchiefs) (
Figure 1).
Although both clumps are populated with the same nematocyst subtypes, they differ significantly in proportions. Morphological descriptions only recognised 2 subtypes of nematocysts, Type I (homotrichous microbasic rhopaloids), and Type II (homotrichous haplonemes) (
Table 1, [
10]), although a third subtype (microbasic mastigophores) was described in 2010 [
4] in
C. barnesi with larger than 25 mm inter-pedalial distance.
Underwood and Seymour [
10] suggested that the differences between bell and tentacle nematocyst concentrations may be ontogenetically related to changing prey targets from invertebrates to vertebrates during the jellyfish’s maturation and growth.
Clinically, Barnes [
2] noted that both the bell and tentacles produced local stings when applied to volunteers in his classic paper on causation. Notably, although all his victims developed Irukandji syndrome, both bell and tentacles were applied to each victim, and consequently he did not identify if either or both (bells and tentacles) are responsible for the development of Irukandji syndrome. There are no published papers that clarify this matter, and there have been only nominal attempts to identify differences where case studies provide dermatological descriptions of the sting site. As such, a “string of pearls” appearance, is attributed to the tentacular nematocyst distribution (neckerchiefs), while a single red wheal with localised piloerection (goose pimples) has been inferred to as an encounter with the bell [
11]. Furthermore, unsupported clinical opinions attribute significant envenomation to tentacular stings from presumptive exposure to more clumps of nematocysts and hence a greater venom load.
Apart from these convenient dermatographic associations, there are no scientific examinations of the difference between bell and tentacle stings, hence our attempt to examine their cardiac effects using pulse wave Doppler in the murine model described by Seymour et al. [
12]. In their study, reconstituted
C. barnesi venom (CBV, mixed tentacular and bell venoms) was parenterally administered to anaesthetised mice. They observed measurable increases in inotropy and chronotropy preceding cardiac compromise and death, which suggested a model consistent with toxin-induced stress cardiomyopathy.
We postulated that differences in effects between venom from the tentacles (CBVt) and bells (CBVb) may be quantitatively assessed through non-invasive pulse wave Doppler examination of the cardiac function.
3. Discussion
The controls demonstrated mild tachycardia and concurrent reduction in %SD. Although both ketamine and phenobarbitone are known direct myocardial depressants, ketamine offsets this effect by promoting endogenous catecholamine release (a useful characteristic in clinical medicine). We can only speculate if these were direct effects of the anaesthetic or if one was a compensatory response. Nevertheless, the combination maintained the %CO output at 100% throughout the observation period. Importantly, there were no changes in systolic (%IVCT) or diastolic (%IVRT) parameters.
Apart from the mode of envenomation, this study replicated the methodology used by Seymour et al. [
12]. As we reproduced cardiac parameter changes, we contend that we have provided evidence of validity to their parenterally administered methodology and, to a lesser extent, other cubozoan venom methodologies.
Many methods have been used in venom preparation for experiments with parenterally administered cubozoan venoms [
14]. These have been summarised by Yanagihara and Shohet and show marked variation in protein compositions, casting some doubt on the efficacy of the test venoms and outcomes [
14]. As mentioned in the previous paragraph, we used the same methodology as described by Bloom [
15] and Carrette [
16], and consequently this research has additionally provided validation for the venom preparation method described.
This study demonstrates statistically significant differences in measured cardiac effects between venoms from nematocysts on the bell (CBVb) compared to venoms from nematocysts on the tentacles (CBVt) of C. barnesi. Of importance, CBVb did not cause cardiac compromise or death in the Mbell group in mice at 60 min post-exposure, whereas CBVt resulted in death in all mice in the Mtent group with exposure to a single neckerchief (mean: 40 min; 95% CL: ± 11 min). Although our portrayal of endpoints appears equivalent between the Mtent and Mbell groups, the actual time difference is a substantial 20 min; for further emphasis, the Mbell group ran for 50% longer with no evidence of cardiovascular collapse and trended to normalisation of cardiovascular function. It is feasible that had the experiment run for an even longer period, the Mbell group may have registered some fatalities, but 60 min was an ethical and experimental time limit.
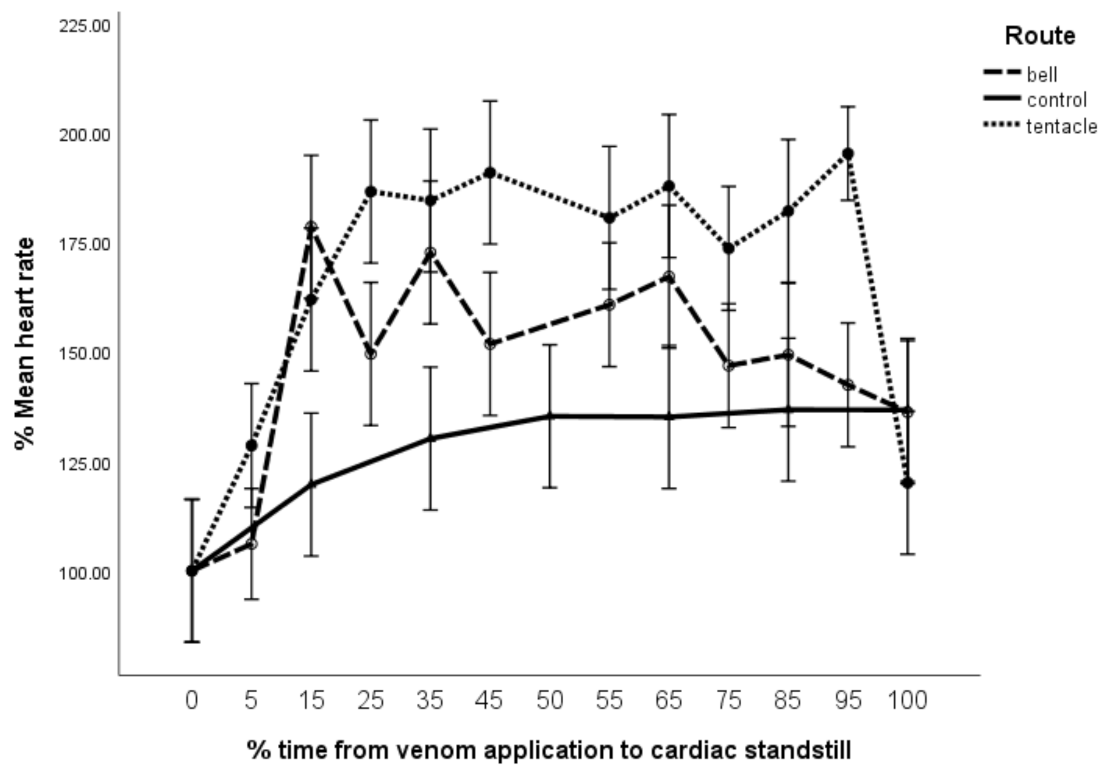
The M
bell group showed a rapid increase in %HR (
Figure 4) peaking at 175% at T
15 (~10 min) trending down to the control group at T
100. This group demonstrated an associated reduction in the %IVRT to 80%, which indicates an improved efficiency in ventricular relaxation. Significantly, there was no accompanying reduction in %IVCT, nominally suggesting no increase in ventricular contraction. As we did not assess aortic valvular flow, we were unable to provide other measures of ventricular contractility such as peak aortic velocity and aortic rise time as described by Seymour [
12], which would provide a more complete assessment of systolic function and inotropy. Despite their absence, our findings do not demonstrate any increase in inotropy, suggesting that the improved ventricular relaxation may just be an accompanying necessity of the observed moderate increase in %HR.
In contrast to the M
bell group, all M
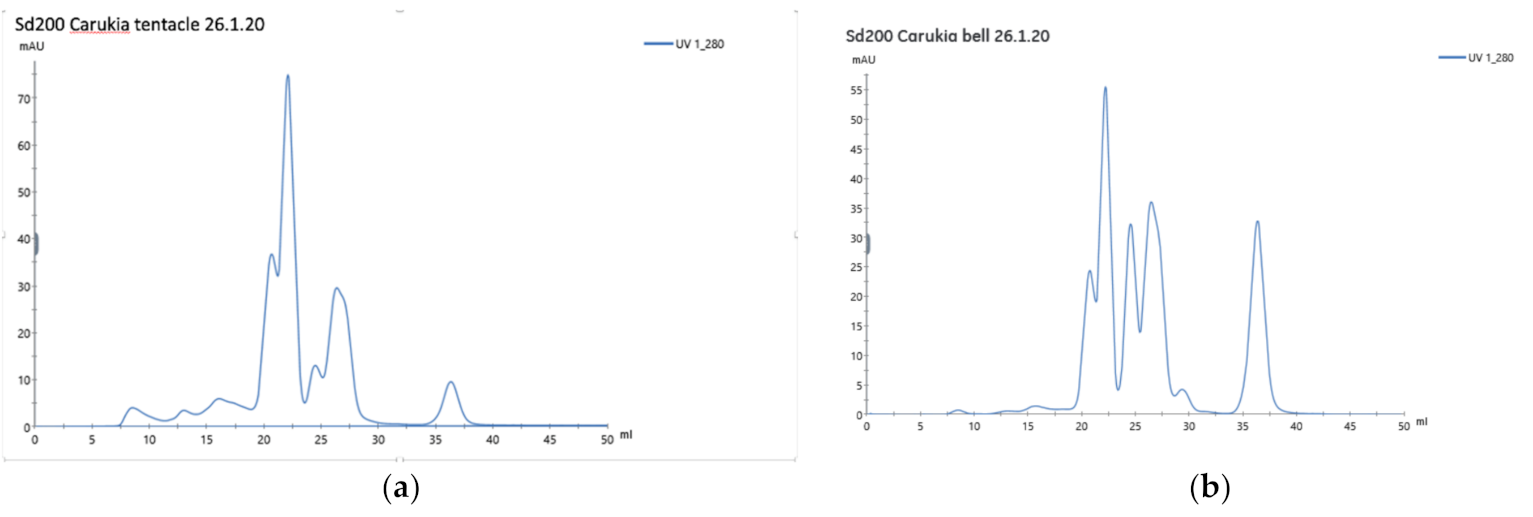
tent mice died before the 60 min endpoint. Clearly, the venoms differ significantly, with a lethal effect conferred by tentacular venom. Both the FPLC and SDS gel demonstrate different peaks and bands, respectively, between the two sources. Although there are other differences evident, our FPLC graphs (
Figure 2a,b) show greater prominence of the 7–20 min elution peaks in CBV
t when compared to the CBV
b. These peaks have gained attention in
C. fleckeri fractionated venom research, suspected to contain the lethal components responsible for box jellyfish fatalities [
17,
18]. We can only speculate that these peaks may be pertinent to our observed difference in lethality. Further research with fractionated CBV
t is intended to examine lethal fractions.
For most of the observed period, the %SD (effectively, the stroke volume) remained the same for both M
tent and M
bell groups and consequently the increase in %CO (
Figure 6) is attributable purely to the observed increased chronotropy.
For consistency across the figures,
Figure 4 shows M
tent’s T
100 based on the loss of %CO rather than the ECG waveform loss (as justified in
Section 5.3 of the Materials and Methods section). Had
Figure 4 been presented as %HR over 60 min observation instead, not only would M
tent have showed the same larger increase in %HR, but it would also have showed a stark reduction in %HR at T
60 (40 min) to below 50% followed by asystole till T
100, while both M
bell and control graphs would have remained unchanged.
Furthermore, it is significant that the loss in pulse wave Doppler (PWD) waveform (and hence %CO) preceded the reduction in heart rate by 4 min (
Figure 4,
Figure 5 and
Figure 6). Clearly, this represents a failure of contraction rather than the loss of electrical activity.
Additional support for this may be found in comparing parameter progression between
Figure 4,
Figure 5 and
Figure 6. It is notable that when M
tent demonstrates a reduction in %SD after T
75 (
Figure 5), there is an accompanying increase in %HR (
Figure 4), peaking at T
95. During this period, %CO is maintained (
Figure 6). This may be explained by a reduction in myocardial contractility (fall in %SD), which is compensated for by an increase in %HR between T
75 and T
95 (over 8 min). This compensation is overwhelmed at T
95, following which there is a precipitous fall in %CO at T
100.
While we nominally noted a widening of the QRS complex accompanying the PWD changes, the authors intend to formally assess ECG parameters (including QRS morphology) in an attempt to examine ion channelopathies that may implicate causation. Our QRS observations may be consistent with Yanagihara and Shohet’s findings of QRS prolongation in that they relate to K
+ leakage across cell membranes from venom-induced pores in their in vitro model [
14]. Ideally, this could have been tested in our experiment, had we measured serum K
+ changes. However, the need for repeat samples would have required a larger animal model that has a significantly larger blood volume than the mouse’s 2 mL to ensure that repeated sampling did not confound our PWD cardiac assessment.
Unlike the M
bell group, the M
tent group shows significant reduction in %IVCT as well as the same improved efficiency in ventricular relaxation (reduced %IVRT) (
Figure 7 and
Figure 8). This suggests that ventricular contraction was more time-efficient for the same ventricular stroke volume, equating to measurable evidence of increased inotropy in this group. Seymour et al. [
12] describe similar changes—increases in inotropy and chronotropy with reconstituted CBV, which were promoted as consistent with toxin-induced stress cardiomyopathy from sustained catecholaminergic activity. With this research, we ratify their findings and further refine the position that the measured cardiac decompensation from the
C. barnesi venom appears to be due to CBV
t alone.
This experiment achieved its aim to solely examine the difference between tentacular and bell envenomation. The following paragraphs speculate as to why these differences exist and how they are achieved. These can be separated into differences in the volume of venom delivered, differences in the potency of delivered venom, and differences in the effectiveness of venom delivery.
It appears that all immature cubozoans have warts, but only
Carybdea species retain these (albeit dwindling numbers) into maturity. Underwood and Seymour [
10] measured between 60 and 220 warts on
C. barnesi bells, the higher numbers associated with juvenile animals which feed solely on invertebrate crustaceans. Notably, these animals had stubby, developing tentacles and an absence of neckerchiefs. Our personal observation is that these immature animals manoeuvre into the path of their prey to effect their sting. As they mature and develop their (clear) tentacles and (visibly prominent) neckerchiefs (
Figure 1a), they appear to use these to catch larval fish by twitching their extended tentacles (note that
Figure 1a depicts a
C. barnesi with contracted tentacles, which, when extended, separate the individual neckerchiefs by up to 10 cm). It is promoted that the twitching of the separated neckerchiefs mimics the movement of their prey and this “angling” appears to entice larval fish into fatally attempting to feed on the neckerchiefs.
The inference from their findings is that the change in diet with maturity is achieved by ontogenetic changes to venom, which they ascribe to differences in populations of nematocyst subtypes (Type I and Type II) between warts and neckerchiefs (
Table 1). It is pertinent that our M
bell model involved the application of half the nematocysts of the bell’s surface and consequently half of the bell’s total nematocysts. This would also constitute a maximum dose in human envenomation. Despite this large exposure, there were no M
bell fatalities and only moderate elevations in HR. In contrast, the M
tent group was uniformly lethal after exposure to just a single neckerchief. It may be that a neckerchief contains vastly greater numbers of nematocysts than 30+ warts and can hence deliver a greater venom load. As yet, there are no studies into nematocyst density per clump.
Alternatively, the difference in lethality may be explained by a difference in venom composition within the same nematocysts that populate both groups (in different proportions). Further research is required to examine this.
Finally, it is possible that the difference in lethality between M
tent and M
bell groups could be explained purely by mechanical problems with venom delivery. For instance, it may be that tentacular nematocysts successfully penetrated and delivered venom into the dermis while bell nematocysts failed to do so. Cutaneous histological sections would have assisted in answering this possibility; however, this was not considered in the experiment’s design. In support of effective venom delivery from bell nematocysts, we can refer to the Barnes’ original paper where he described an observed cutaneous reaction to bell exposure which was separate from the tentacular reaction [
2].
Our Mbell methodology simulates a worst-case scenario with an adult human vs. bell encounter. That it does not result in fatality nor reduce the measured %CO in a mouse when the comparative dose/kg is at least 3000× greater than that with a human victim (24 g mouse vs. 70 kg human) is significant. Other than the remote possibility that this may be explained by mice having an innate resistance to CBVb when compared to humans, this would strongly suggest that the bell is not causative in the cardiac decompensation encountered in severe Irukandji syndrome.
Further extrapolation from a 20 g mouse to a 70 kg human suggests that the bell of C. barnesi may not cause any significant clinical catecholaminergic effects and may not be expected to produce any significant clinical systemic envenomation. This, of course, can only be ascertained through verifiable human exposure studies.
The bell’s apparent lack of significant effect may explain the wide variation in clinical severity described by Huynh et al. [
3], where some proven
C. barnesi stings were associated with no systemic symptoms of envenomation. It would also be consistent with other researchers’ ascription of bell exposure to mild stings [
11]. It also follows from this that encounters with juvenile
C. barnesi with poorly developed tentacles are more likely to induce minor stings.
Limitations
This study was an in vivo murine study, with only 9 mice examined and only 3 C. barnesi animals used. We have no way of knowing if there was equivalence in venom dosage between tentacle and bell groups when compared to the parenteral administration of reconstituted venoms, where protein concentrations are measured and dosage can be accounted for. We were careful to expose only a single neckerchief to each mouse in the tentacle group, which is likely to be a comparatively small exposure in a marine environment. In contrast, the application of up to half the bell would constitute a maximum exposure in a similar marine environment.
We were limited to one PWD probe per mouse and we elected to apply this to examine the mitral valve where we could obtain both diastolic and nominal systolic parameters of ventricular function. As such, we did not examine the aortic waveform, where a more complete measure of strength of ventricular contraction and systolic function can be obtained through additional parameters.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}