Zearalenone and Its Metabolites—General Overview, Occurrence, and Toxicity


Abstract
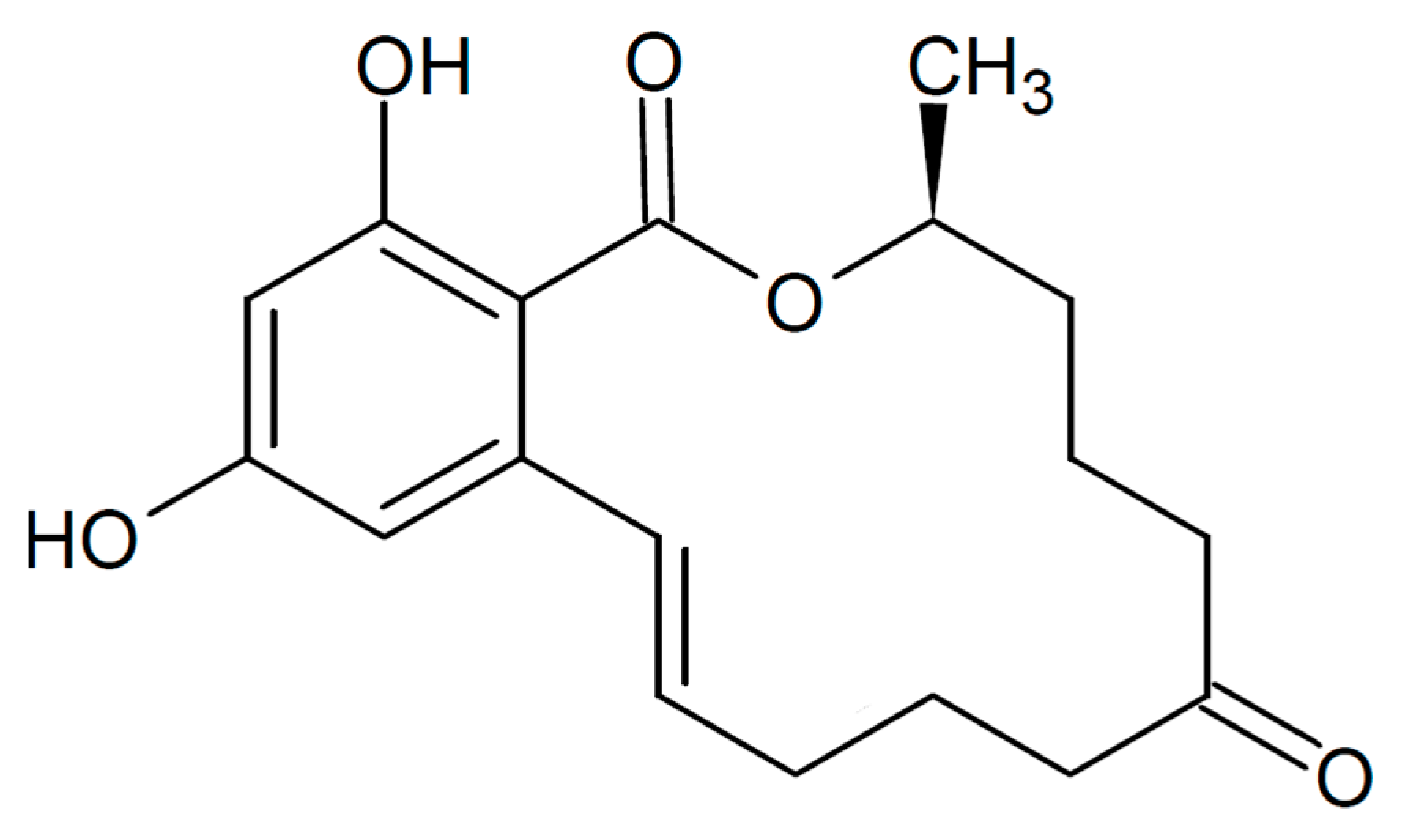
:1. Introduction
2. The Occurrence of ZEN in Food
3. The Occurrence of ZEN in Body Fluids
4. The Impact of ZEN on Organisms
5. Toxicokinetics of ZEN
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- SCOOP (European Commission, Directorate-General Health and Consumer Protection- Scientific Co-Operation on Questions relating to Food). SCOOP, Task 3.2.10. Collection of Occurrence Data of Fusarium Toxins in Food and Assessment of Dietary Intake by the Populstion of EU Member States. European Commission, Directorate-General Health and Consumer Protection, Reports on Tasks Forscientific Co-Operation; European Commission, Directorate-General Health and Consumer Protection: Brussel, Belgium, 2003; Available online: https://ec.europa.eu/food/sites/food/files/safety/docs/cs_contaminants_catalogue_fusarium_task3210.pdf (accessed on 2 May 2020).
- Gil-Serna, J.; Vázquez, C.; González-Jaén, M.T.; Patiño, B. Mycotoxins. Toxicology. In Encyclopedia of Food Microbiology; Batt, C.A., Tortorello, M.L., Eds.; Academic Press: Cambridge, MA, USA, 2014; pp. 887–892. [Google Scholar]
- Mostrom, M.S. Zearalenone. In Veterinary Toxicology; Basic and Clinical Principles, Gupta, R., Eds.; Academic Press: Cambridge, MA, USA, 2012; pp. 1266–1271. [Google Scholar]
- Mizutani, K.; Nagatomi, Y.; Mochizuki, N. Metabolism of Zearalenone in the Course of Beer Fermentation. Toxins 2011, 3, 134–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mally, A.; Solfrizzo, M.; Degen, G.H. Biomonitoring of the mycotoxin Zearalenone: Current state--of--the art and application to human exposure assessment. Arch. Toxicol. 2016, 90, 1281–1292. [Google Scholar] [CrossRef] [PubMed]
- Urry, W.H.; Wehrmeister, H.H.; Hodge, E.B.; Hidy, P.H. The structure of zearalenone. Tetrahedron Lett. 1966, 7, 3109–3114. [Google Scholar] [CrossRef]
- Stob, M.; Baldwin, R.S.; Tuite, J.; Andrews, F.N.; Gillette, K.G. Isolation of an anabolic, uterotrophic compound from corn infected with Gibberella zeae. Nature 1962, 196, 1318. [Google Scholar] [CrossRef]
- Tsakmakidis, I.A.; Lymberopoulos, A.G.; Alexopoulos, C.; Boscos, C.M.; Kyriakis, S.C. In vitro Effffect of Zearalenone and a-Zearalenol on Boar Sperm Characteristics and Acrosome Reaction. Reprod. Dom. Anim. 2006, 41, 394–401. [Google Scholar] [CrossRef]
- Zinedine, A.; Soriano, J.M.; Moltó, J.C.; Mañes, J. Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: An oestrogenic mycotoxin. Food Chem. Toxicol. 2007, 45, 1–18. [Google Scholar] [CrossRef]
- Gromadzka, K.; Waskiewicz, A.; Chełkowski, J.; Goliński, P. Zearalenone and its metabolites: Occurrence, detection, toxicity and guidelines. World Mycotoxin J. 2008, 1, 209–220. [Google Scholar] [CrossRef]
- Edite Bezerra da Rocha, M.; da Freire, F.C.O.; Erlan Feitosa Maia, F.; Guedes, M.I.F.; Rondina, D. Mycotoxins and their effects on human and animal health. Food Control 2014, 36, 159–165. [Google Scholar] [CrossRef]
- Martins, C.; Torres, D.; Lopes, C.; Correia, D.; Goios, A.; Assunção, R.; Alvito, P.; Vidal, A.; De Boevre, M.; De Saeger, S.; et al. Food Consumption Data as a Tool to Estimate Exposure to Mycoestrogens. Toxins 2020, 12, 118. [Google Scholar] [CrossRef] [Green Version]
- Rogowska, A.; Pomastowski, P.; Sagandykova, G.; Buszewski, B. Zearalenone and its metabolites: Effect on human health, metabolism and neutralisation methods. Toxicon 2019, 162, 46–56. [Google Scholar] [CrossRef]
- Döll, S.; Dänicke, S. The Fusarium toxins deoxynivalenol (DON) and zearalenone (ZON) in animal feeding. Prev. Veter. Med. 2011, 102, 132–145. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.; Kim, W.; Park, J.-H.; Kim, D.; Kim, C.-R.; Chung, S.; Lee, C. The Occurrence of Zearalenone in South Korean Feedstuffs between 2009 and 2016. Toxins 2017, 9, 223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hidy, P.H.; Baldwin, R.S.; Greasham, R.L.; Keith, C.L.; Mcmullen, J.R. Zearalenone and Some Derivatives: Production and Biological Activities. Adv. Appl. Microbiol. 1977, 22, 59–82. [Google Scholar] [PubMed]
- Ben Salah-Abbès, J.; Belgacem, H.; Ezzdini, K.; Abdel-Wahhab, M.A.; Abbès, S. Zearalenone nephrotoxicity: DNA fragmentation, apoptotic gene expression and oxidative stress protected by Lactobacillus plantarum MON03. Toxicon 2020, 175, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Kovalsky Paris, M.P.; Schweiger, W.; Hametner, C.; Stückler, R.; Muehlbauer, G.J.; Varga, E.; Krska, R.; Berthiller, F.; Adam, G. Zearalenone-16-O-glucoside: A New Masked Mycotoxin. J. Agric. Food Chem. 2014, 62, 1181–1189. [Google Scholar] [CrossRef]
- Ueberschär, K.-H.; Brezina, U.; Dänicke, S. Zearalenone (ZEN) and ZEN metabolites in feed, urine and bile of sows: Analysis, determination of the metabolic profile and evaluation of the binding forms. Appl. Agric. For. Res. 2016, 1, 21–28. [Google Scholar]
- Minervini, F.; Giannoccaro, A.; Fornelli, F.; Dell’Aquila, M.E.; Minoia, P.; Visconti, A. Influence of mycotoxin zearalenone and its derivatives (alpha and beta zearalenol) on apoptosis and proliferation of cultured granulosa cells from equine ovaries. Reprod. Biol. Endocrinol. 2006, 4, 62. [Google Scholar] [CrossRef] [Green Version]
- Mirocha, C.J.; Pathre, S.V.; Behrens, J.; Schauerhamer, B. Uterotropic activity of cis and trans isomers of zearalenone and zearalenol. Appl. Environ. Microbiol. 1978, 35, 986–987. [Google Scholar] [CrossRef] [Green Version]
- Rai, A.; Das, M.; Tripathi, A. Occurrence and toxicity of a fusarium mycotoxin, zearalenone. Crit. Rev. Food Sci. Nutr. 2019, 60, 2710–2729. [Google Scholar] [CrossRef]
- Songsermsakul, P.; Sontag, G.; Cichnamarkl, M.; Zentek, J.; Razzazifazeli, E. Determination of zearalenone and its metabolites in urine, plasma and faeces of horses by HPLC–APCI–MS. J. Chromatogr. B 2006, 843, 252–261. [Google Scholar] [CrossRef]
- Zhang, G.-L.; Feng, Y.-L.; Song, J.-L.; Zhou, X.-S. Zearalenone: A Mycotoxin With Different Toxic Effect in Domestic and Laboratory Animals’ Granulosa Cells. Front. Genet. 2018, 9, 667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Böswald, C.; Engelhardt, G.; Vogel, H.; Wallnöfer, P.R. Metabolism of the Fusarium mycotoxins zearalenone and deoxynivalenol by yeast strains of technological relevance. Nat. Toxins 1995, 3, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.; Liu, H.; Sun, J.; Wang, J.; Zhao, C.; Zhang, W.; Zhang, J.; Sun, C. Zearalenone Removal from Corn Oil by an Enzymatic Strategy. Toxins 2020, 12, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sprynskyy, M.; Gadzała-Kopciuch, R.; Nowak, K.; Buszewski, M. Removal zearalenone toxin from synthetics gastric and body flfluids using talc and diatomite: A batch kinetic study. Colloids Surf. B Biointerfaces 2012, 94, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Regulation of the European Commission (EC). No. 1881/2006 of December 19, 2006, as Amended d. Fixing Maximum Levels for Certain Contaminants in Foodstuffs (OJ. L. 364/5 of 20.12.2006, Annex “Maximum Levels for Certain Contaminants in fooDstuffs”. Off. J. Eur. Union. 2006, 364, 5–24. [Google Scholar]
- Commission Recommendation of 17 August 2006 on the Presence of Deoxynivalenol, Zearalenone, Ochratoxin A, T-2 and HT-2 and Fumonisins in Products Intended for Animal Nutrition (2006/576/EC as Amended) (OJ. L./229/7). Off. J. Eur. Union. 2006, 229, 7–9.
- Zinedine, A.; Ruiz, M.-J. Zearalenone. Mycotoxins Implic. Food Safety 2014, 52–66. [Google Scholar] [CrossRef]
- Fink-Gremmels, J.; Malekinejad, H. Clinical effects and biochemical mechanisms associated with exposure to the mycoestrogen zearalenone. Anim. Feed. Sci. Technol. 2007, 137, 326–341. [Google Scholar] [CrossRef]
- Prelusky, D.B.; Scott, P.M.; Trenholm, H.L.; Lawrence, G.A. Minimal transmission of zearalenone to milk of dairy cows. J. Environ. Sci. Heal. Part B 1990, 25, 87–103. [Google Scholar] [CrossRef]
- Coffey, R.; Cummins, E.; Ward, S. Exposure assessment of mycotoxins in dairy milk. Food Control 2009, 20, 239–249. [Google Scholar] [CrossRef]
- Domijan, A.-M.; Peraica, M.; Cvjetković, B.; Turčin, S.; Jurjević, Ž.; Ivić, D. Mould contamination and co-occurrence of mycotoxins in maize grain in Croatia. Acta Pharm. 2005, 55, 349–356. [Google Scholar] [PubMed]
- Scudamore, K.A.; Patel, S. Occurrence of Fusarium mycotoxins in maize imported into the UK, 2004–2007. Food Addit. Contam. Part A 2009, 26, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Manova, R.; MLadenova, R. Incidence of zearalenone and fumonisins in Bulgarian cereal production. Food Control 2009, 20, 362–365. [Google Scholar] [CrossRef]
- Zinedine, A.; Brera, C.; Elakhdari, S.; Catano, C.; Debegnach, F.; Angelini, S.; De Santis, B.; Faid, M.; BenlemLih, M.; Minardi, V.; et al. Natural occurrence of mycotoxins in cereals and spices commercialized in Morocco. Food Control 2006, 17, 868–874. [Google Scholar] [CrossRef]
- Schollenberger, M.; Müller, H.-M.; Rüfle, M.; Suchy, S.; Plank, S.; Drochner, W. Natural Occurrence of 16 Fusarium Toxins in Grains and Feedstuffs of Plant Origin from Germany. Mycopathologia 2006, 161, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Roigé, M.B.; Aranguren, S.M.; Riccio, M.B.; Pereyra, S.; Soraci, A.L.; Tapia, M.O. Mycobiota and mycotoxins in fermented feed, wheat grains and corn grains in Southeastern Buenos Aires Province, Argentina. Rev. Iberoam. Micol. 2009, 26, 233–237. [Google Scholar] [CrossRef]
- Cano-Sancho, G.; Marin, S.; Ramos, A.J.; Sanchis, V. Occurrence of zearalenone, an oestrogenic mycotoxin, in Catalonia (Spain) and exposure assessment. Food Chem. Toxicol. 2012, 50, 835–839. [Google Scholar] [CrossRef]
- Pleadin, J.; Zadravec, M.; Perši, N.; Vulić, A.; Jaki, V.; Mitak, M. Mould and mycotoxin contamination of pig feed in northwest Croatia. Mycotoxin Res. 2012, 28, 157–162. [Google Scholar] [CrossRef]
- Mwihia, E.W.; Lyche, J.L.; Mbuthia, P.G.; Ivanova, L.; Uhlig, S.; Gathumbi, J.K.; Maina, J.G.; Eshitera, E.E.; Eriksen, G.S. Co-Occurrence and Levels of Mycotoxins in Fish Feeds in Kenya. Toxins 2020, 12, 627. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, Q.; Yan, Z.; Tan, Y.; Zhu, R.; Yu, D.; Yang, H.; Wu, A. Occurrence and Quantitative Risk Assessment of Twelve Mycotoxins in Eggs and Chicken Tissues in China. Toxins 2018, 10, 477. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, S.Z.; Nisar, S.; Asi, M.R.; Jinap, S. Natural incidence of aflaToxins ochratoxin A and zearalenone in chicken meat and eggs. Food Control 2014, 43, 98–103. [Google Scholar] [CrossRef]
- Mahmoudi, R. Occurrence of Zearalenone in raw animal origin food produced in North-West of Iran. J. Food Qual. Hazards Control 2014, 1, 25–28. [Google Scholar]
- Meucci, V.; Soldani, G.; Razzuoli, E.; Saggese, G.; Massart, F. Mycoestrogen Pollution of Italian Infant Food. J. Pediatr. 2011, 159, 278–283.e1. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, S.Z.; Rabbani, T.; Asi, M.R.; Jinap, S. Assessment of aflaToxins ochratoxin A and zearalenone in breakfast cereals. Food Chem. 2014, 157, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Curtui, V.G.; Gareis, M.; Usleber, E.; Märtlbauer, E. Survey of Romanian slaughtered pigs for the occurrence of mycotoxins ochratoxins A and B, and zearalenone. Food Addit. Contam. 2001, 18, 730–738. [Google Scholar] [CrossRef] [PubMed]
- Stoev, S.D.; Dutton, M.F.; Njobeh, P.B.; Mosonik, J.S.; Steenkamp, P.A. Mycotoxic nephropathy in Bulgarian pigs and chickens: Complex aetiology and similarity to Balkan Endemic Nephropathy. Food Addit. Contam. Part A 2010, 27, 72–88. [Google Scholar] [CrossRef] [Green Version]
- Rubert, J.; León, N.; Sáez, C.; Martins, C.P.; Godula, M.; Yusà, V.; Mañes, J.; Soriano, J.M.; Soler, C. Evaluation of mycotoxins and their metabolites in human breast milk using liquid chromatography coupled to high resolution mass spectrometry. Anal. Chim. Acta 2014, 820, 39–46. [Google Scholar] [CrossRef]
- Massart, F.; Micillo, F.; Rivezzi, G.; Perrone, L.; Baggiani, A.; Miccoli, M.; Meucci, V. Zearalenone screening of human breast milk from the Naples area. Toxicol. Environ. Chem. 2015, 98, 128–136. [Google Scholar] [CrossRef]
- Valitutti, F.; De Santis, B.; Trovato, C.M.; Montuori, M.; Gatti, S.; Oliva, S.; Brera, C.; Catassi, C. Assessment of Mycotoxin Exposure in Breastfeeding Mothers with Celiac Disease. Nutrients 2018, 10, 336. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.C.; Zheng, N.; Zheng, B.Q.; Wen, F.; Cheng, J.B.; Han, R.W.; Xu, X.M.; Li, S.L.; Wang, J.Q. Simultaneous determination of aflatoxin M1, ochratoxin A, zearalenone and α-zearalenol in milk by UHPLC–MS/MS. Food Chem. 2014, 146, 242–249. [Google Scholar] [CrossRef]
- Pleadin, J.; Mihaljević, Ž.; Barbir, T.; Vulić, A.; Kmetič, I.; Zadravec, M.; Brumen, V.; Mitak, M. Natural incidence of zearalenone in Croatian pig feed, urine and meat in 2014. Food Addit. Contam. Part B 2015, 80, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Gambacorta, L.; Olsen, M.; Solfrizzo, M. Pig Urinary Concentration of Mycotoxins and Metabolites Reflects Regional Differences, Mycotoxin Intake and Feed Contaminations. Toxins 2019, 11, 378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Njumbe Ediage, E.; Diana Di Mavungu, J.; Song, S.; Sioen, I.; De Saeger, S. Multimycotoxin analysis in urines to assess infant exposure: A case study in Cameroon. Environ. Int. 2013, 57–58, 50–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ezekiel, C.N.; Warth, B.; Ogara, I.M.; Abia, W.A.; Ezekiel, V.C.; Atehnkeng, J.; Sulyok, M.; Turner, P.C.; Tayo, G.O.; Krska, R.; et al. Mycotoxin exposure in rural residents in northern Nigeria: A pilot study using multi-urinary biomarkers. Environ. Int. 2014, 66, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Solfrizzo, M.; Gambacorta, L.; Visconti, A. Assessment of Multi-Mycotoxin Exposure in Southern Italy by Urinary Multi-Biomarker Determination. Toxins 2014, 6, 523–538. [Google Scholar] [CrossRef]
- Wallin, S.; Gambacorta, L.; Kotova, N.; Warensjö Lemming, E.; Nälsén, C.; Solfrizzo, M.; Olsen, M. Biomonitoring of concurrent mycotoxin exposure among adults in Sweden through urinary multi-biomarker analysis. Food Chem. Toxicol. 2015, 83, 133–139. [Google Scholar] [CrossRef]
- Föllmann, W.; Ali, N.; Blaszkewicz, M.; Degen, G.H. Biomonitoring of Mycotoxins in Urine: Pilot Study in Mill Workers. J. Toxicol. Environ. Heal. Part A 2016, 79, 1015–1025. [Google Scholar] [CrossRef]
- Šarkanj, B.; Ezekiel, C.N.; Turner, P.C.; Abia, W.A.; Rychlik, M.; Krska, R.; Sulyok, M.; Warth, B. Ultra-sensitive, stable isotope assisted quantification of multiple urinary mycotoxin exposure biomarkers. Anal. Chim. Acta 2018, 1019, 84–92. [Google Scholar] [CrossRef]
- Ezekiel, C.N.; Abia, W.A.; Braun, D.; Šarkanj, B.; Ayeni, K.I.; Oyedele, O.A.; Michael-Chikezie, E.C.; Ezekiel, V.C.; Mark, B.; Ahuchaogu, C.P.; et al. Comprehensive mycotoxin exposure biomonitoring in breastfed and non-exclusively breastfed Nigerian children. MedRxiv 2020. [Google Scholar] [CrossRef]
- Mauro, T.; Hao, L.; Pop, L.C.; Buckley, B.; Schneider, S.H.; Bandera, E.V.; Shapses, S.A. Circulating zearalenone and its metabolites differ in women due to body mass index and food intake. Food Chem. Toxicol. 2018, 116, 227–232. [Google Scholar] [CrossRef]
- Pillay, D.; Chuturgoon, A.A.; Nevines, E.; Manickum, T.; Deppe, W.; Dutton, M.F. The Quantitative Analysis of Zearalenone and Its Derivatives in Plasma of Patients with Breast and Cervical Cancer. Clin. Chem. Lab. Med. 2002, 40, 40. [Google Scholar] [CrossRef] [PubMed]
- Fleck, S.C.; Churchwell, M.I.; Doerge, D.R.; Teeguarden, J.G. Urine and serum biomonitoring of exposure to environmental estrogens II: Soy isoflavones and zearalenone in pregnant women. Food Chem. Toxicol. 2016, 95, 19–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Santis, B.; Raggi, M.; Moretti, G.; Facchiano, F.; Mezzelani, A.; Villa, L.; Bonfanti, A.; Campioni, A.; Rossi, S.; Camposeo, S.; et al. Study on the Association among Mycotoxins and other Variables in Children with Autism. Toxins 2017, 9, 203. [Google Scholar] [CrossRef] [PubMed]
- Tassis, P.D.; Tsakmakidis, I.A.; Nagl, V.; Reisinger, N.; Tzika, E.; Gruber-Dorninger, C.; Michos, I.; Mittas, N.; Basioura, A.; Schatzmayr, D. Individual and Combined In Vitro Effects of Deoxynivalenol and Zearalenone on Boar Semen. Toxins 2020, 12, 495. [Google Scholar] [CrossRef]
- Buszewska-Forajta, M. MycoToxins invisible danger of feedstuff with toxic effect on animals. Toxicon 2020, 182, 34–53. [Google Scholar] [CrossRef]
- Gajęcka, M.; Gajęcki, M. Is mycotoxins can be used as inhibitors in milk? Innow. MLecz. 2014, 2, 22–29. [Google Scholar]
- Marin, D.E.; Pistol, G.C.; Bulgaru, C.V.; Taranu, I. Cytotoxic and inflammatory effects of individual and combined exposure of HepG2 cells to zearalenone and its metabolites. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2019, 392, 937–947. [Google Scholar] [CrossRef]
- Hueza, I.M.; Raspantini, P.C.F.; Raspantini, L.E.R.; Latorre, A.O.; Górniak, S.L. Zearalenone, an Estrogenic Mycotoxin, Is an Immunotoxic Compound. Toxins 2014, 6, 1080–1095. [Google Scholar] [CrossRef] [Green Version]
- Pietsch, C.; Kersten, S.; Valenta, H.; Dänicke, S.; Schulz, C.; Burkhardt-Holm, P.; Junge, R. Effects of Dietary Exposure to Zearalenone (ZEN) on Carp (Cyprinus carpio L.). Toxins 2015, 7, 3465–3480. [Google Scholar] [CrossRef] [Green Version]
- Maaroufi, K.; Chekir, L.; Creppy, E.E.; Ellouz, F.; Bacha, H. Zearalenone induces modifications of haematological and biochemical parameters in rats. Toxicon 1996, 34, 535–540. [Google Scholar] [CrossRef]
- Zwierzchowski, W.; Przybyłowicz, M.; Obremski, K.; Zielonka, L.; Skorska-Wyszyńska, E.; Gajecka, M.; Polak, M.; Jakimiuk, E.; Jana, B.; Rybarczyk, L.; et al. Level of zearalenone in blood serum and lesions in ovarian follicles of sexually immature gilts in the course of zearalenone micotoxicosis. Pol. J. Veter Sci. 2005, 8, 209–218. [Google Scholar]
- Jodlbauer, J.; Zöllner, P.; Lindner, W. Determination of zearalenone and its metabolites in urine and tissue samples of cow and pig by LC-MS/MS. Mycotoxin Res. 2000, 16 (Suppl. 2), 174–178. [Google Scholar] [CrossRef] [PubMed]
- Takagi, M.; Uno, S.; Kokushi, E.; Shiga, S.; Mukai, S.; Kuriyagawa, T.; Takagaki, K.; Hasunuma, H.; Matsumoto, D.; Okamoto, K.; et al. Measurement of urinary zearalenone concentrations for monitoring natural feed contamination in cattle herds: On-farm trials. J. Anim. Sci. 2011, 89, 287–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zain, M.E. Impact of mycotoxins on humans and animals. J. Saudi Chem. Soc. 2011, 15, 129–144. [Google Scholar] [CrossRef] [Green Version]
- Peraica, M.; Radić, B.; Lucić, A.; Pavlović, M. Toxic effects of mycotoxins in humans. Bull World Health Org. 1999, 77, 754–766. [Google Scholar] [PubMed]
- Binder, S.B.; Schwartz-Zimmermann, H.E.; Varga, E.; Bichl, G.; Michlmayr, H.; Adam, G.; Berthiller, F. Metabolism of Zearalenone and Its Major Modified Forms in Pigs. Toxins 2017, 9, 56. [Google Scholar] [CrossRef] [Green Version]
- Shier, W.T.; Shier, A.C.; Xie, W.; Mirocha, C.J. Structure-activity relationships for human estrogenic activity in zearalenone mycotoxins. Toxicon 2001, 39, 1435–1438. [Google Scholar] [CrossRef]
- Gliński, Z.; Kostro, K.; Gajęcki, M. Mikozy i Mikotoksykozy Zwierząt; Wyd. Uniwersytet Przyrodniczy w Lublinie: Lublin, Poland, 2011; p. 296. [Google Scholar]
- El-Sharkawy, S.H.; Selin, M.I.; Afifi, M.S.; Halaweish, F.T. Microbial Transformation of Zearalenone to a Zearalenone Sulfate. Appl. Environ. Microbiol. 1991, 57, 549–552. [Google Scholar] [CrossRef] [Green Version]
- Sáenz de Rodriguez, C.A.; Bongiovanni, A.M.; de Borrego, L.C. An epidemic of precocious development in Puerto Rican children. J. Pediatr. 1985, 107, 393–396. [Google Scholar] [CrossRef]
- Kwaśniewska, K.; Gadzała-Kopciuch, R.; Cendrowski, K. Analytical Procedure for the Determination of Zearalenone in Environmental and Biological Samples. Crit. Rev. Anal. Chem. 2015, 45, 119–130. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain (CONTAM). Risks for animal health related to the presence of zearalenone and its modified forms in feed. EFSA J. 2017, 15, 4851. [Google Scholar]
- Miksicek, R.J. Interaction of naturally occurring nonsteroidal estrogens with expressed recombinant human estrogen receptor. J. Steroid Biochem. Mol. Biol. 1994, 49, 153–160. [Google Scholar] [CrossRef]
- Kim, I. Zearalenone induces male germ cell apoptosis in rats. Toxicol. Lett. 2003, 138, 185–192. [Google Scholar] [CrossRef]
- International Programme On Chemical Safety (IPCS). Safety Evaluation of Certain Food Additives and Contaminants; World Health Organization: Geneva, Switzerland, 2000. [Google Scholar]
- Kuiper-Goodman, T. Uncertainties in the risk assessment of three mycotoxins: Aflatoxin, ochratoxin, and zearalenone. Can. J. Physiol. Pharmacol. 1990, 68, 1017–1024. [Google Scholar] [CrossRef]
- Obremski, K.; Gajęcki, M.; Zwierzchowski, W.; Bakuła, T.; Apoznański, J.; Wojciechowski, J. The level of zearalenone and α-zearalenol in the blood of gilts with clinical symptoms of toxicosis, fed diets with a low zearalenone content. J. Anim. Feed Sci. 2003, 12, 529–538. [Google Scholar] [CrossRef]
- EFSA. Scientific Opinion on the risks for public health related to the presence of zearalenone in food. EFSA J. 2011, 9, 2197. [Google Scholar] [CrossRef]
- D’Mello, J.P.F.; Placinta, C.M.; Macdonald, A.M.C. Fusarium mycotoxins: A review of global implications for animal health, welfare and productivity. Anim. Feed. Sci. Technol. 1999, 80, 183–205. [Google Scholar] [CrossRef] [Green Version]
- Rivera-Núñez, Z.; Barrett, E.S.; Szamreta, E.A.; Shapses, S.A.; Qin, B.; Lin, Y.; Zarbl, H.; Buckley, B.; Bandera, E.V. Urinary mycoestrogens and age and height at menarche in New Jersey girls. Environ. Health 2019, 18, 24. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Product | Highest Permissible Value [μg/kg] |
|---|---|
| Unprocessed cereals other than maize | 100 |
| Unprocessed maize | 350 |
| Cereals intended for direct human consumption, cereal flour, bran as end product marketed for direct human consumption and germ | 75 |
| Refined corn oil | 400 |
| Maize intended for direct human consumption, maize snacks, and maize-based breakfast cereals | 100 |
| Bread (including small bakery wares), cakes, biscuits, cereal snacks, and breakfast cereals, excluding maize snacks and maize based breakfast cereals | 50 |
| Processed cereal-based foods (excluding processed maize-based foods) and baby foods for infants and young children | 20 |
| Processed corn-based foods for infants and young children | 20 |
| Compound feed for piglets, gilts, puppies, kittens, dogs, and cats intended for reproduction | 0.1 |
| Compound feed for adult dogs and cats other than those intended for reproduction | 0.2 |
| Compound feed for sows and porkers | 0.25 |
| Compound feed for calves, dairy cattle, sheep (including lambs), and goats (including goatlings) | 0.5 |
| Country | Products | % of Positive Samples (Number of Samples) | Results | References |
|---|---|---|---|---|
| Croatia | Maize | 80% (12/15) | range 0.62–3.2 μg/kg | [34] |
| Argentina | Raw maize | 100% (26/26) | mean 15 μg/kg, maximum 42 μg/kg | [35] |
| Bulgaria | Maize | 21.1% (4/19) | mean 80.6 μg/kg, maximum 148 μg/kg | [36] |
| Morocco | Corn | 15% (3/20) | mean 14 μg/kg, maximum 17 μg/kg | [37] |
| Germany | Corn | 85% (35/41) | mean 48 μg/kg, maximum 860 μg/kg | [38] |
| Argentina | Corn grains | 36% (21/58) | maximum 1560 μg/kg | [39] |
| Spain | Corn snacks | 23.6% (17/72) | maximum 22.8 μg/kg | [40] |
| Germany | Wheat | 63% (26/41) | mean 15 μg/kg | [38] |
| Bulgaria | Wheat | 1.9% (1/54) | mean 10 μg/kg, maximum 10 μg/kg | [36] |
| Germany | Oats | 24% (4/17) | mean 21 μg/kg | [38] |
| Germany | Hay | 42% (12/28) | mean 24 μg/kg, maximum 115 μg/kg | [38] |
| Germany | Peas | 0% | - | [38] |
| South Korea | Beans | 100% (1/1) | maximum 15 µg/kg | [15] |
| South Korea | Grains | 77% (17/22) | maximum 277 µg/kg | [15] |
| Bulgaria | Barley | 11.1% (2/18) | mean 29 μg/kg, maximum 36.6 μg/kg | [36] |
| Germany | Soya meal | 69% (9/13) | mean 51 μg/kg, maximum 211 μg/kg | [38] |
| South Korea | Fibrous feed | 50% (4/8) | maximum 1315 µg/kg | [15] |
| South Korea | Food byproducts | 62% (8/13) | maximum 176 µg/kg | [15] |
| Croatia | Feed mixtures for fattening pigs | 93.3% (28/30) | range 8.93–866 μg/kg | [41] |
| Kenya | Fish feed | 40% (31/78) | range from < 38.0–757.9 ng/mL | [42] |
| China | Eggs | 44% (32/72) | range between 0.30–418 µg/kg | [43] |
| Pakistan | Eggs | 45% (18/40) | mean ± SD 2.23 ± 0.51 μg/kg | [44] |
| Pakistan | Chicken meat | 52% (60/115) | mean ± SD 2.01 ± 0.90 μg/kg | [44] |
| Iran | Buffalo meat | 41.42% (29/70) | range from 0.1–2.5 ng/mL | [45] |
| Iran | Buffalo liver | 68.57% (48/70) | range from 0.1–4.34 ng/mL | [45] |
| Italy | Cow’s milk-based infant formula | 9% (17/185) | maximum 0.76 μg/L | [46] |
| Pakistan | Bread (corn) | 43% (6/14) | mean ± SD 9.45 ± 2.76 μg/kg | [47] |
| Spain | Sliced bread | 43.6% (31/71) | maximum 20.9 μg/kg | [40] |
| Spain | Beer | 11.3% (8/71) | maximum 5.1 μg/kg | [40] |
| Spain | Pasta | 14.3% (10/70) | maximum 5.9 μg/kg | [40] |
| ZEN Metabolites | Country | Products | % of Positive Samples (Number of Samples) | Results | References |
|---|---|---|---|---|---|
| α-ZEL | Italy | Cow’s milk-based infant formula | 26% (49/185) | maximum 12.91 μg/L | [46] |
| β-ZEL | 28% (53/185) | maximum 73.24 μg/L | |||
| α-ZEL | China | Chicken heart | 40% (8/20) | mean 3.60 µg/kg | [43] |
| Chicken Gizzard | 40% (8/20) | mean 4.01 µg/kg | |||
| α-ZEL | Kenya | Fish feed | 24% (19/78) | range from < 22.2–288.4 ng/mL | [42] |
| β-ZEL | 33% (26/78) | range from < 16.0–79.8 ng/mL |
| Country | Body Fluid | % of Positive Samples (Number of Samples) | Results | References |
|---|---|---|---|---|
| Romania | Pig’s serum | 17.3% (9/52) | mean 0.8 ng/mL, maximum 0.96 ng/mL | [48] |
| Bulgaria | Pig’s serum | 50% (5/10) | mean ± SD 0.24 ± 0.12 μg/L | [49] |
| Bulgaria | Pig’s serum | 50% (5/10) | mean ± SD 0.33 ± 0.17μg/L | [49] |
| Iran | Buffaloes milk | 21.42% (15/70) | range between 0.1–3.55 ng/mL | [45] |
| Spain | Breast milk | 37% (13/35) | range between 2.1–14.3 ng/mL | [50] |
| Italy | Breast milk | 100% (47/47) | range between 0.26–1.78 μg/L | [51] |
| Italy | Breast milk (women with Celiac Disease) | 4% (12/275) | range between 2.0–17 ng/mL | [52] |
| Italy | Breast milk | 8% (15/178) | range between 2.0–22 ng/mL | [52] |
| China | Raw milk | 100% (30/30) | mean ± SD 14.9 ± 6.0 ng/kg | [53] |
| China | Liquid milk | 100% (12/12) | mean ± SD 20.5 ± 11.1 ng/kg | [53] |
| Croatia | Pig’s urine (male) | 100% (11/11) | mean ± SD 238 ± 30 µg/L, range between 104–350 µg/L | [54] |
| Croatia | Pig’s urine (female) | 100% (19/19) | mean ± SD 187 ± 27.1 µg/L, range between 22.7–390 µg/L | [54] |
| Sweden | Pig’s urine | 92% (179/195) | mean ± SD 2.44 ± 4.39 ng/mL | [55] |
| Cameroon | Human urine | 3.6% (8/220) | mean 0.97 ng/mL, range between 0.65–5.0 ng/mL | [56] |
| Nigeria | Human urine | 0.8% (1/120) | mean 0.3 µg/L | [57] |
| Italy | Human urine | 100% (52/52) | mean 0.057 ng/mL, maximum 0.120 ng/mL | [58] |
| Sweden | Human urine | 37% (92/252) | mean ± SD 0.09 ± 0.07 ng/mL | [59] |
| Germany | Male urine (control) | 100% (13/13) | mean ± SD 31 ± 23 ng/L, range between 7–90 ng/L | [60] |
| Germany | Male urine (Mill worker) | 100% (12/12) | mean ± SD 42 ± 26 ng/L, range between 4–100 ng/L | [60] |
| Germany | Female urine (Mill worker) | 100% (5/5) | mean ± SD 35 ± 28 ng/L, range between 6–78 ng/L | [60] |
| Nigeria | Human urine | 81.7% (98/120) | mean 0.75 ng/mL, range between 0.03–19.99 ng/mL | [61] |
| Nigeria | Breastfed infants urine | 57% (13/23) | mean 148 ng/L, range between 17–784 ng/L | [62] |
| Nigeria | Non-exclusively breastfed infants urine | 83% (35/42) | mean 140 ng/L, range between 13–678 ng/L | [62] |
| Parameter | Value |
|---|---|
| LOAEL | 200 µg/kg bw/day (15-day pig study) |
| LOAEL | 56 µg/kg bw/day (sheep) |
| LOAEL | 17.6 µg/kg bw/day (piglets) |
| LOAEL | 200 µg/kg bw/day (gilts) |
| LOAEL | 20 µg/kg bw/day (dogs) |
| NOEL | 40 µg/kg bw/day (pigs) |
| NOEL | 9200 µg (mice) |
| NOEL | 100 μg/kg bw (rats) |
| NOEL | 28 µg/kg bw/day (sheep) |
| NOEL | 10.4 µg/kg bw/day (piglets) |
| NOEL | 40 µg/kg bw/day (gilts) |
| LD50 | 2000 mg/kg (mice, rats, and guinea pigs) |
| TD50 | 20,000 µg (mice) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ropejko, K.; Twarużek, M. Zearalenone and Its Metabolites—General Overview, Occurrence, and Toxicity. Toxins 2021, 13, 35. https://doi.org/10.3390/toxins13010035
Ropejko K, Twarużek M. Zearalenone and Its Metabolites—General Overview, Occurrence, and Toxicity. Toxins. 2021; 13(1):35. https://doi.org/10.3390/toxins13010035
Chicago/Turabian StyleRopejko, Karolina, and Magdalena Twarużek. 2021. "Zearalenone and Its Metabolites—General Overview, Occurrence, and Toxicity" Toxins 13, no. 1: 35. https://doi.org/10.3390/toxins13010035
APA StyleRopejko, K., & Twarużek, M. (2021). Zearalenone and Its Metabolites—General Overview, Occurrence, and Toxicity. Toxins, 13(1), 35. https://doi.org/10.3390/toxins13010035