Formulation and Safety Tests of a Wickerhamomyces anomalus–Based Product: Potential Use of Killer Toxins of a Mosquito Symbiotic Yeast to Limit Malaria Transmission

 ,
,  ,
,  and
and 
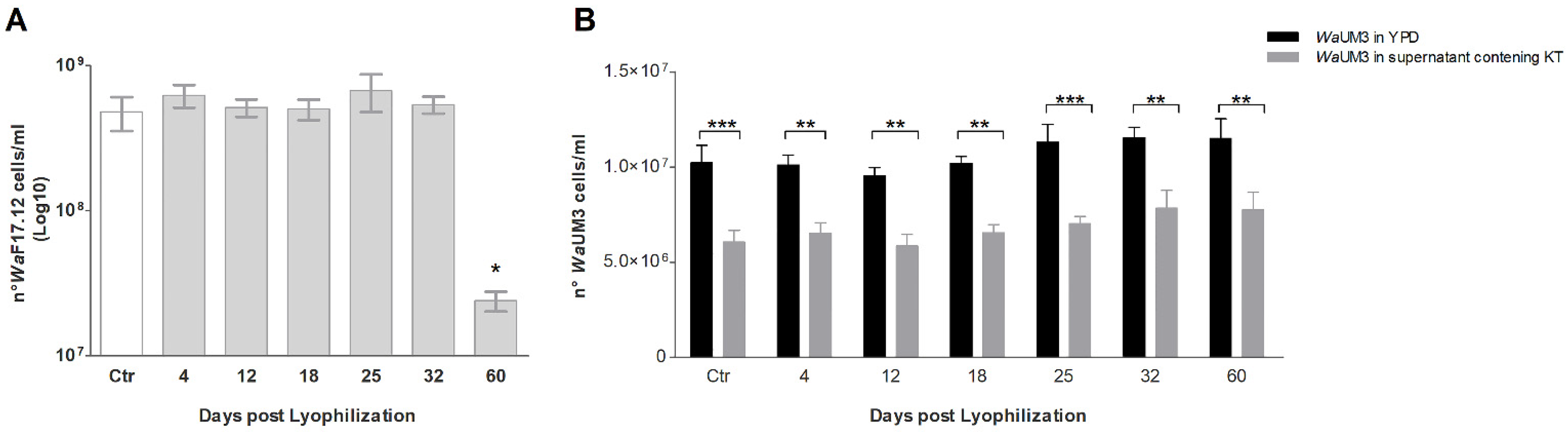{kind=link}
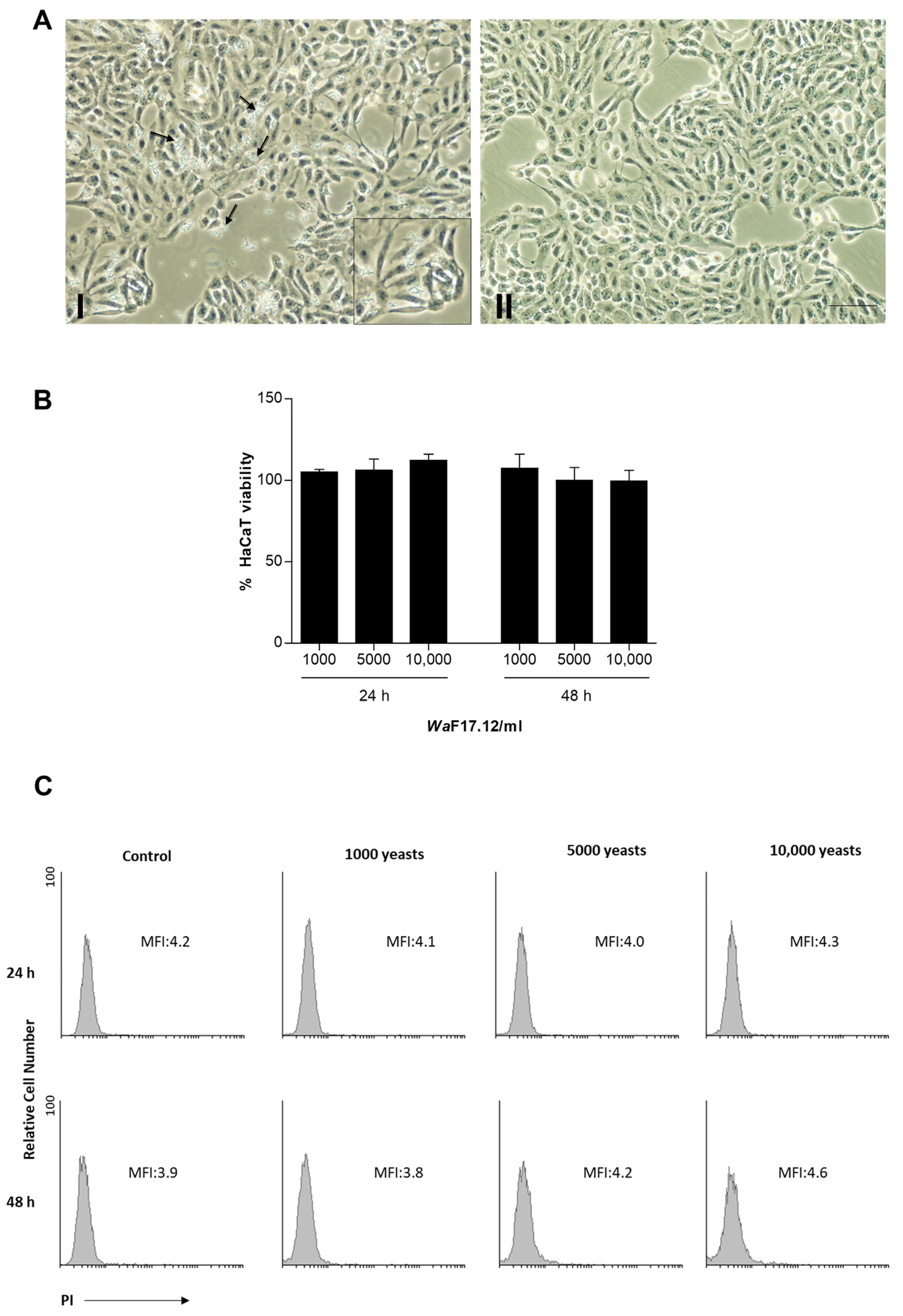{kind=link}
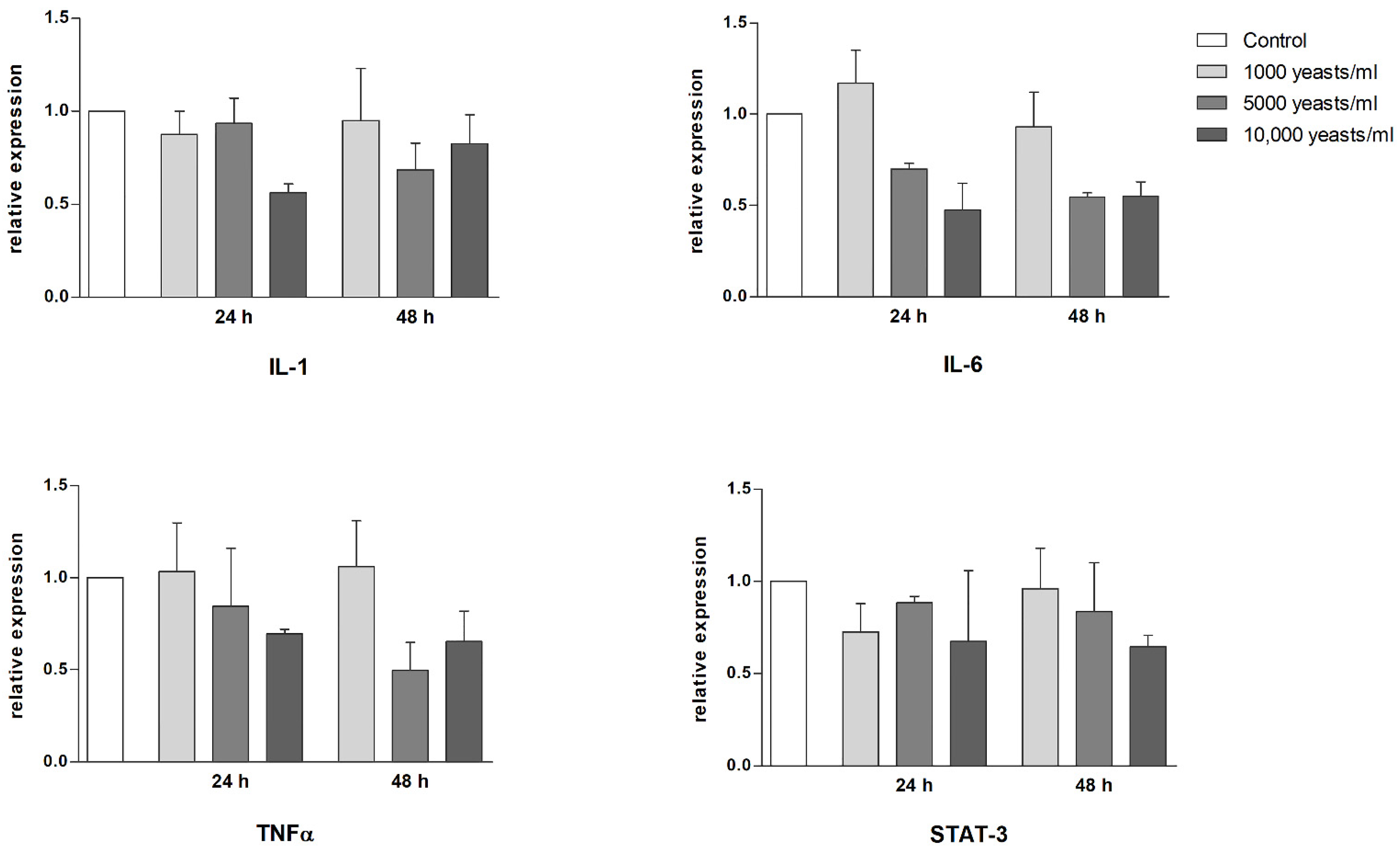{kind=link}
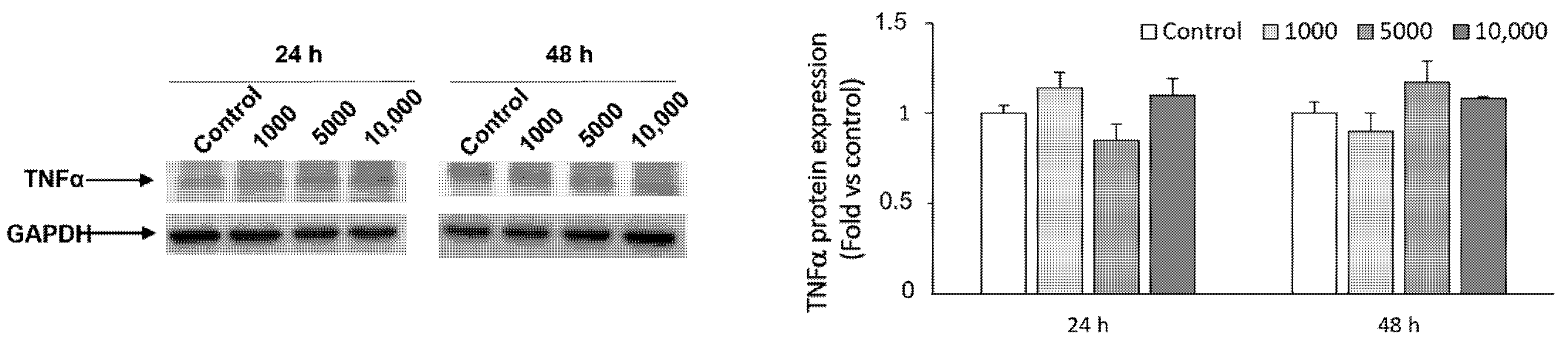{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Growth Reactivation and Killer Toxin Activity Maintenance of WaF17.12 after Lyophilization
2.2. Effects of WaF17.12 on Viability of Human Keratinocytes (HaCaT)
2.3. Effect of WaF17.12 on the Proinflammatory Response
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Yeast
5.2. HaCaT Cells Cultures
5.3. Freeze-Drying of WaF17.12
5.4. Growth Rate Assessment and Killer Toxin Activity Assay Post-Lyophilization
5.5. Treatment of HaCaT Cells with WaF17.12
5.6. Cell Viability Assays in HaCaT
5.7. Gene Expression Analysis of Proinflammatory Mediators
5.8. Western Blot Analysis of TNFα
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Walker, G.M. Pichia anomala: Cell physiology and biotechnology relative to other yeasts. Antonie Van Leeuwenhoek 2011, 99, 25–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Salvador, P.; Escamez, F.; Girones, R.; Herman, L.; Koutsoumanis, K.; Lindqvist, R.; et al. Update of the list of QPS-recommended biological agents intentionally added to food or feed as notified to EFSA 4: Suitability of taxonomic units notified to EFSA until March 2016. EFSA J. 2017, 15, 4663. [Google Scholar] [CrossRef]
- Cappelli, A.; Favia, G.; Ricci, I. Wickerhamomyces anomalus in Mosquitoes: A Promising Yeast-Based Tool for the “Symbiotic Control” of Mosquito-Borne Diseases. Front. Microbiol. 2021, 11, 621605. [Google Scholar] [CrossRef] [PubMed]
- Martin, E.; Bongiorno, G.; Giovati, L.; Montagna, M.; Crotti, E.; Damiani, C.; Gradoni, L.; Polonelli, L.; Ricci, I.; Favia, G.; et al. Isolation of a Wickerhamomyces anomalus yeast strain from the sandfly Phlebotomus perniciosus, displaying the killer phenotype. Med. Vet. Entomol. 2016, 30, 101–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, N.N. Insecticide Resistance in Mosquitoes: Impact, Mechanisms, and Research Directions. Annu. Rev. Entomol. 2015, 60, 537–559. [Google Scholar] [CrossRef] [PubMed]
- Benelli, G.; Jeffries, C.L.; Walker, T. Biological Control of Mosquito Vectors: Past, Present, and Future. Insects 2016, 7, 52. [Google Scholar] [CrossRef] [PubMed]
- Ricci, I.; Damiani, C.; Scuppa, P.; Mosca, M.; Crotti, E.; Rossi, P.; Rizzi, A.; Capone, A.; Gonella, E.; Ballarini, P.; et al. The yeast Wickerhamomyces anomalus (Pichia anomala) inhabits the midgut and reproductive system of the Asian malaria vector Anopheles stephensi. Environ. Microbiol. 2011, 13, 911–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cappelli, A.; Valzano, M.; Cecarini, V.; Bozic, J.; Rossi, P.; Mensah, P.; Amantini, C.; Favia, G.; Ricci, I. Killer yeasts exert anti-plasmodial activities against the malaria parasite Plasmodium berghei in the vector mosquito Anopheles stephensi and in mice. Parasites Vectors 2019, 12, 329. [Google Scholar] [CrossRef]
- Stefanini, I. Yeast-insect associations: It takes guts. Yeast 2018, 35, 315–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madden, A.A.; Epps, M.J.; Fukami, T.; Irwin, R.E.; Sheppard, J.; Sorger, D.M.; Dunn, R.R. The ecology of insect-yeast relationships and its relevance to human industry. Proc. Biol. Sci. 2018, 285, 20172733. [Google Scholar] [CrossRef] [Green Version]
- Passoth, V.; Olstorpe, M.; Schnurer, J. Past, present and future research directions with Pichia anomala. Antonie Van Leeuwenhoek 2011, 99, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.R.; Zhao, L.N.; Li, Z.B.; Li, C.; Li, B.; Gu, X.Y.; Zhang, X.Y.; Zhang, H.Y. Screening and identification of an antagonistic yeast controlling postharvest blue mold decay of pears and the possible mechanisms involved. Biol. Control 2019, 133, 26–33. [Google Scholar] [CrossRef]
- Junges, D.S.B.; Delabeneta, M.F.; Rosseto, L.R.B.; Nascimento, B.L.; Paris, A.P.; Persel, C.; Loth, E.A.; Simao, R.C.G.; Menolli, R.A.; Paula, C.R.; et al. Antibiotic Activity of Wickerhamomyces anomalus Mycocins on Multidrug-Resistant Acinetobacter baumannii. Microb. Ecol. 2020, 80, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Jarque, S.; Bittner, M.; Hilscherova, K. Freeze-drying as suitable method to achieve ready-to-use yeast biosensors for androgenic and estrogenic compounds. Chemosphere 2016, 148, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Stefanello, R.F.; Nabeshima, E.H.; Iamanaka, B.T.; Ludwig, A.; Fries, L.L.M.; Bernardi, A.O.; Copetti, M.V. Survival and stability of Lactobacillus fermentum and Wickerhamomyces anomalus strains upon lyophilisation with different cryoprotectant agents. Food Res. Int. 2019, 115, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Cappelli, A.; Ulissi, U.; Valzano, M.; Damiani, C.; Epis, S.; Gabrielli, M.G.; Conti, S.; Polonelli, L.; Bandi, C.; Favia, G.; et al. A Wickerhamomyces anomalus killer strain in the malaria vector Anopheles stephensi. PLoS ONE 2014, 9, e95988. [Google Scholar] [CrossRef]
- Rossi, P.; Cappelli, A.; Marinelli, O.; Valzano, M.; Pavoni, L.; Bonacucina, G.; Petrelli, R.; Pompei, P.; Mazzara, E.; Ricci, I.; et al. Mosquitocidal and Anti-Inflammatory Properties of The Essential Oils Obtained from Monoecious, Male, and Female Inflorescences of Hemp. Molecules 2020, 25, 3451. [Google Scholar] [CrossRef]
- Colombo, I.; Sangiovanni, E.; Maggio, R.; Mattozzi, C.; Zava, S.; Corbett, Y.; Fumagalli, M.; Carlino, C.; Corsetto, P.A.; Scaccabarozzi, D.; et al. HaCaT Cells as a Reliable In Vitro Differentiation Model to Dissect the Inflammatory/Repair Response of Human Keratinocytes. Mediat. Inflamm. 2017, 2017, 7435621. [Google Scholar] [CrossRef] [PubMed]
- Sangiovanni, E.; Fumagalli, M.; Pacchetti, B.; Piazza, S.; Magnavacca, A.; Khalilpour, S.; Melzi, G.; Martinelli, G.; Dell’Agli, M. Cannabis sativa L. extract and cannabidiol inhibit in vitro mediators of skin inflammation and wound injury. Phytother. Res. 2019, 33, 2083–2093. [Google Scholar] [CrossRef]
- Sano, S.; Chan, K.S.; Carbajal, S.; Clifford, J.; Peavey, M.; Kiguchi, K.; Itami, S.; Nickoloff, B.J.; DiGiovanni, J. Stat3 links activated keratinocytes and immunocytes required for development of psoriasis in a novel transgenic mouse model. Nat. Med. 2005, 11, 43–49. [Google Scholar] [CrossRef]
- Mannazzu, I.; Domizio, P.; Carboni, G.; Zara, S.; Zara, G.; Comitini, F.; Budroni, M.; Ciani, M. Yeast killer toxins: From ecological significance to application. Crit. Rev. Biotechnol. 2019, 39, 603–617. [Google Scholar] [CrossRef]
- Parafati, L.; Vitale, A.; Restuccia, C.; Cirvilleri, G. The effect of locust bean gum (LBG)-based edible coatings carrying biocontrol yeasts against Penicillium digitatum and Penicillium italicum causal agents of postharvest decay of mandarin fruit. Food Microbiol. 2016, 58, 87–94. [Google Scholar] [CrossRef]
- Oro, L.; Feliziani, E.; Ciani, M.; Romanazzi, G.; Comitini, F. Volatile organic compounds from Wickerhamomyces anomalus, Metschnikowia pulcherrima and Saccharomyces cerevisiae inhibit growth of decay causing fungi and control postharvest diseases of strawberries. Int. J. Food Microbiol. 2018, 265, 18–22. [Google Scholar] [CrossRef]
- Ricci, I.; Mosca, M.; Valzano, M.; Damiani, C.; Scuppa, P.; Rossi, P.; Crotti, E.; Cappelli, A.; Ulissi, U.; Capone, A.; et al. Different mosquito species host Wickerhamomyces anomalus (Pichia anomala): Perspectives on vector-borne diseases symbiotic control. Antonie Van Leeuwenhoek 2011, 99, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Steyn, A.; Roets, F.; Botha, A. Yeasts Associated with Culex pipiens and Culex theileri Mosquito Larvae and the Effect of Selected Yeast Strains on the Ontogeny of Culex pipiens. Microb. Ecol. 2016, 71, 747–760. [Google Scholar] [CrossRef]
- Valzano, M.; Cecarini, V.; Cappelli, A.; Capone, A.; Bozic, J.; Cuccioloni, M.; Epis, S.; Petrelli, D.; Angeletti, M.; Eleuteri, A.M.; et al. A yeast strain associated to Anopheles mosquitoes produces a toxin able to kill malaria parasites. Malar. J. 2016, 15, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melin, P.; Schnurer, J.; Hakansson, S. Formulation and stabilisation of the biocontrol yeast Pichia anomala. Antonie Van Leeuwenhoek 2011, 99, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Dutra, V.R.; Silva, L.F.; Oliveira, A.N.M.; Beirigo, E.F.; Arthur, V.M.; da Silva, R.B.; Ferreira, T.B.; Andrade-Silva, L.; Silva, M.V.; Fonseca, F.M.; et al. Fatal Case of Fungemia by Wickerhamomyces anomalus in a Pediatric Patient Diagnosed in a Teaching Hospital from Brazil. J. Fungi 2020, 6, 147. [Google Scholar] [CrossRef] [PubMed]
- Epis, S.; Capone, A.; Martin, E.; Paolucci, M.; Bazzocchi, C.; Valzano, M.; Bozic, J.; Novati, S.; Favia, G.; Ricci, I. A rapid qPCR method to investigate the circulation of the yeast Wickerhamomyces anomalus in humans. New Microbiol. 2015, 38, 577–581. [Google Scholar] [PubMed]
- Cecarini, V.; Cuccioloni, M.; Bonfili, L.; Ricciutelli, M.; Valzano, M.; Cappelli, A.; Amantini, C.; Favia, G.; Eleuteri, A.M.; Angeletti, M.; et al. Identification of a Killer Toxin from Wickerhamomyces anomalus with beta-Glucanase Activity. Toxins 2019, 11, 568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polonelli, L.; Seguy, N.; Conti, S.; Gerloni, M.; Bertolotti, D.; Cantelli, C.; Magliani, W.; Cailliez, J.C. Monoclonal yeast killer toxin-like candidacidal anti-idiotypic antibodies. Clin. Diagn. Lab. Immunol. 1997, 4, 142–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melin, P.; Hakansson, S.; Schnurer, J. Optimisation and comparison of liquid and dry formulations of the biocontrol yeast Pichia anomala J121. Appl. Microbiol. Biotechnol. 2007, 73, 1008–1016. [Google Scholar] [CrossRef] [PubMed]
- Nahr, F.K.; Mokarram, R.R.; Hejazi, M.A.; Ghanbarzadeh, B.; Khiyabani, M.S.; Benis, K.Z. Optimization of the nanocellulose based cryoprotective medium to enhance the viability of freeze dried Lactobacillus plantarum using response surface methodology. Lwt-Food Sci. Technol. 2015, 64, 326–332. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cappelli, A.; Amantini, C.; Maggi, F.; Favia, G.; Ricci, I. Formulation and Safety Tests of a Wickerhamomyces anomalus–Based Product: Potential Use of Killer Toxins of a Mosquito Symbiotic Yeast to Limit Malaria Transmission. Toxins 2021, 13, 676. https://doi.org/10.3390/toxins13100676
Cappelli A, Amantini C, Maggi F, Favia G, Ricci I. Formulation and Safety Tests of a Wickerhamomyces anomalus–Based Product: Potential Use of Killer Toxins of a Mosquito Symbiotic Yeast to Limit Malaria Transmission. Toxins. 2021; 13(10):676. https://doi.org/10.3390/toxins13100676
Chicago/Turabian StyleCappelli, Alessia, Consuelo Amantini, Federica Maggi, Guido Favia, and Irene Ricci. 2021. "Formulation and Safety Tests of a Wickerhamomyces anomalus–Based Product: Potential Use of Killer Toxins of a Mosquito Symbiotic Yeast to Limit Malaria Transmission" Toxins 13, no. 10: 676. https://doi.org/10.3390/toxins13100676
APA StyleCappelli, A., Amantini, C., Maggi, F., Favia, G., & Ricci, I. (2021). Formulation and Safety Tests of a Wickerhamomyces anomalus–Based Product: Potential Use of Killer Toxins of a Mosquito Symbiotic Yeast to Limit Malaria Transmission. Toxins, 13(10), 676. https://doi.org/10.3390/toxins13100676