A Short Review of the Venoms and Toxins of Spider Wasps (Hymenoptera: Pompilidae)


Abstract
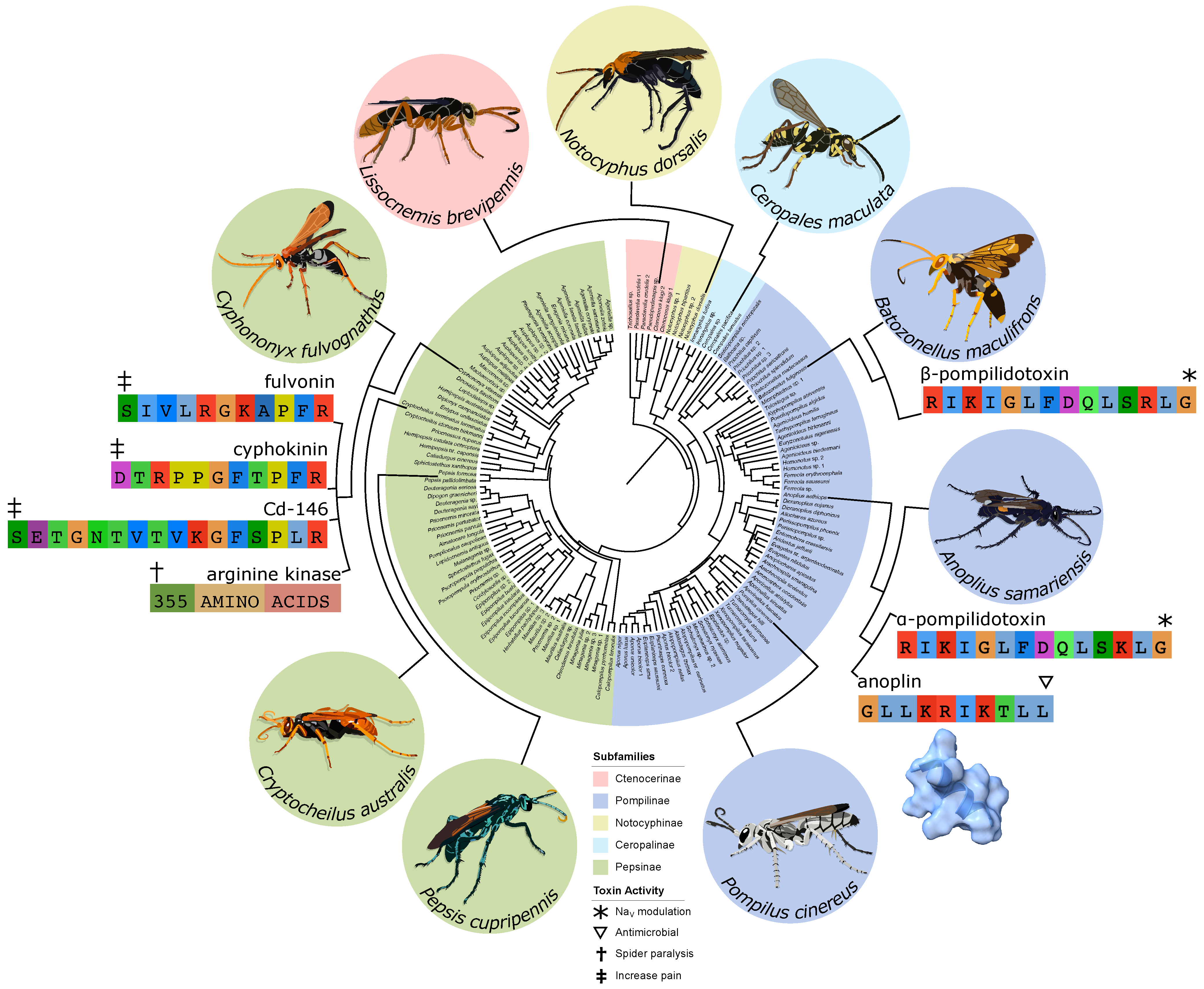
:1. Introduction
2. Natural History Observations

{kind=link}
| Wasp | Spider | Behavioral Notes | Paralysis | Further Notes | Ref. |
|---|---|---|---|---|---|
| Paracyphononyx africanus | Lycosidae | Spiders live freely with egg attached | 15 min | [90] | |
| Fabriogenia sp. | Lycosidae | Wasp amputated spider’s legs | <1 day | palps responded to stimuli | per. obs. |
| Anoplius nigerrimus | Lycosidae | Spiders restrained in small cells | 1–2 h | [94] | |
| Anoplius apiculatus autumnalis | Arctosa littoralis | Spiders restrained in small cells | 1–2 h | [95] | |
| Anoplius apiculatus autumnalis | Lycosidae | Spiders restrained in small cells | 1–2 h | [96] | |
| Anoplius apiculatus pretiosus | Lycosidae | Spiders restrained in small cells | 1–2 h | [96] | |
| Anoplius semirufus | Lycosidae | Spiders restrained in small cells | 1–2 h | [96] | |
| Anoplius semirufus | various | Spiders restrained in small cells | 1–4 h | [97] | |
| Anoplius marginalis | Lycosidae | Spiders restrained in small cells | 2 h | [98] | |
| Anoplius tenebrosus | unspecified | Spiders restrained in small cells | 4 h | [97] | |
| Anoplius semirufus | Trochosa avara | Spiders restrained in small cells | 6 h | [99] | |
| Anoplius apiculatus pretiosus | Arctosa littoralis | Spiders restrained in small cells | 7 h | [100] | |
| Pompilus scelestus | Geolycosa rafaelana | >1 d | nesting takes up to a day | [98] | |
| Pompilus quinquenotatus | Larinioides cornutus | 30 d | [89] | ||
| Sericopompilus apicalis | unspecified | >46 d | spiders died | [96] | |
| Pompilus bigutattus | Metepeira labyrinthea | 62 d | [89] | ||
| Pepsis marginata | Cyrtopholis portoricae | 2.5 mo | [101] | ||
| Cryptocheilus affinis | unspecified | >4 mo | spiders died | [102] | |
| Pepsis cupripennis | unclear if Acanthoscurria suina, Eupalaestrus weijenberghi, or both | 8 mo | [91] |
3. Lab Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| PMTX | pompilidotoxin |
| NaV | voltage-gated sodium channel |
| CSIRO | Commonwealth Science & Industry Research Organisation |
| CERC | CSIRO Early Research Career |
References
- Nelsen, D.R.; Nisani, Z.; Cooper, A.M.; Fox, G.A.; Gren, E.C.K.; Corbit, A.G.; Hayes, W.K. Poisons, toxungens, and venoms: Redefining and classifying toxic biological secretions and the organisms that employ them. Biol. Rev. 2014, 89, 450–465. [Google Scholar] [CrossRef]
- Jackson, T.N.; Fry, B.G. A tricky trait: Applying the fruits of the “function debate” in the philosophy of biology to the “venom debate” in the science of toxinology. Toxins 2016, 8, 263. [Google Scholar] [CrossRef] [Green Version]
- Beard, R.L. Insect toxins and venoms. Annu. Rev. Entomol. 1963, 8, 1–18. [Google Scholar] [CrossRef]
- Schmidt, J.O. Biochemistry of insect venoms. Annu. Rev. Entomol. 1982, 27, 339–368. [Google Scholar] [CrossRef]
- Fry, B.G.; Roelants, K.; Champagne, D.E.; Scheib, H.; Tyndall, J.D.; King, G.F.; Nevalainen, T.J.; Norman, J.A.; Lewis, R.J.; Norton, R.S.; et al. The toxicogenomic multiverse: Convergent recruitment of proteins into animal venoms. Annu. Rev. Genom. Hum. Genet. 2009, 10, 483–511. [Google Scholar] [CrossRef] [Green Version]
- Arbuckle, K.; Harris, R.J. Radiating pain: Venom has contributed to the diversification of the largest radiations of vertebrate and invertebrate animals. BMC Ecol. Evol. 2021, 21, 150. [Google Scholar] [CrossRef]
- Gaston, K.J. The magnitude of global insect species richness. Conserv. Biol. 1991, 5, 283–296. [Google Scholar] [CrossRef]
- Stork, N.E. How many species of insects and other terrestrial arthropods are there on Earth? Annu. Rev. Entomol. 2018, 63, 31–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forbes, A.A.; Bagley, R.K.; Beer, M.A.; Hippee, A.C.; Widmayer, H.A. Quantifying the unquantifiable: Why Hymenoptera, not Coleoptera, is the most speciose animal order. BMC Ecol. 2018, 18, 21. [Google Scholar] [CrossRef] [Green Version]
- Whitfield, J.B. Phylogeny and evolution of host-parasitoid interactions in Hymenoptera. Annu. Rev. Entomol. 1998, 43, 129–151. [Google Scholar] [CrossRef]
- Heraty, J. Parasitoid biodiversity and insect pest management. In Insect Biodiversity; Blackwell Publishing: Hoboken, NJ, USA, 2009; pp. 445–462. [Google Scholar]
- Peters, R.S.; Krogmann, L.; Mayer, C.; Donath, A.; Gunkel, S.; Meusemann, K.; Kozlov, A.; Podsiadlowski, L.; Petersen, M.; Lanfear, R.; et al. Evolutionary history of the Hymenoptera. Curr. Biol. 2017, 27, 1013–1018. [Google Scholar] [CrossRef] [Green Version]
- Fry, B.G.; Koludarov, I.; Jackson, T.N.; Holford, M.; Terrat, Y.; Casewell, N.R.; Undheim, E.A.; Vetter, I.; Ali, S.A.; Low, D.H. Seeing the woods for the trees: Understanding venom evolution as a guide for biodiscovery. In Venoms to Drugs: Venom as a Source for the Development of Human Therapeutics; King, G., Ed.; Royal Society of Chemistry: Cambridge, UK, 2015; pp. 1–36. [Google Scholar]
- Walker, A.A.; Robinson, S.D.; Yeates, D.K.; Jin, J.; Baumann, K.; Dobson, J.; Fry, B.G.; King, G.F. Entomo-venomics: The evolution, biology and biochemistry of insect venoms. Toxicon 2018, 154, 15–27. [Google Scholar] [CrossRef] [Green Version]
- Greener, M. The next generation of venom-based drugs. Prescriber 2020, 31, 28–32. [Google Scholar] [CrossRef]
- Poirié, M.; Carton, Y.; Dubuffet, A. Virulence strategies in parasitoid Hymenoptera as an example of adaptive diversity. Comptes Rendus Biol. 2009, 332, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Cruz, Y.P. A sterile defender morph in a polyembryonic hymenopterous parasite. Nature 1981, 294, 446–447. [Google Scholar] [CrossRef]
- Cruz, Y.P. The defender role of the precocious larvae of Copidosomopsis tanytmemus Caltagirone (Encyrtidae, Hymenoptera). J. Exp. Zool. 1986, 237, 309–318. [Google Scholar] [CrossRef]
- Fleming, J.G.; Summers, M.D. Polydnavirus DNA is integrated in the DNA of its parasitoid wasp host. Proc. Natl. Acad. Sci. USA 1991, 88, 9770–9774. [Google Scholar] [CrossRef] [Green Version]
- Webb, B.A. Polydnavirus Biology, Genome Structure, and Evolution. In The Insect Viruses; The Viruses; Miller, L.K., Ball, L.A., Eds.; Springer: Boston, MA, USA, 1998; pp. 105–139. [Google Scholar] [CrossRef]
- Whitfield, J.B. Estimating the age of the polydnavirus/braconid wasp symbiosis. Proc. Natl. Acad. Sci. USA 2002, 99, 7508–7513. [Google Scholar] [CrossRef] [Green Version]
- Whitfield, J.B.; Asgari, S. Virus or not? Phylogenetics of polydnaviruses and their wasp carriers. J. Insect Physiol. 2003, 49, 397–405. [Google Scholar] [CrossRef]
- Gauthier, J.; Drezen, J.M.; Herniou, E.A. The recurrent domestication of viruses: Major evolutionary transitions in parasitic wasps. Parasitology 2018, 145, 713–723. [Google Scholar] [CrossRef]
- Fleming, J.A.G. Polydnaviruses: Mutualists and pathogens. Annu. Rev. Entomol. 1992, 37, 401–425. [Google Scholar] [CrossRef]
- Lavine, M.D.; Beckage, N.E. Polydnaviruses: Potent mediators of host insect immune dysfunction. Parasitol. Today 1995, 11, 368–378. [Google Scholar] [CrossRef]
- Strand, M.R.; Burke, G.R. Polydnavirus-wasp associations: Evolution, genome organization, and function. Curr. Opin. Virol. 2013, 3, 587–594. [Google Scholar] [CrossRef]
- Brodeur, J.; Vet, L.E.M. Usurpation of host behaviour by a parasitic wasp. Anim. Behav. 1994, 48, 187–192. [Google Scholar] [CrossRef] [Green Version]
- Kester, K.M.; Jackson, D.M. When good bugs go bad: Intraguild predation by Jalysus wickhami on the parasitoid, Cotesia congregata. Entomol. Exp. Appl. 1996, 81, 271–276. [Google Scholar] [CrossRef]
- Grosman, A.H.; Janssen, A.; Brito, E.F.d.; Cordeiro, E.G.; Colares, F.; Fonseca, J.O.; Lima, E.R.; Pallini, A.; Sabelis, M.W. Parasitoid increases survival of its pupae by inducing hosts to fight predators. PLoS ONE 2008, 3, e2276. [Google Scholar] [CrossRef] [Green Version]
- Harvey, J.A.; Tanaka, T.; Kruidhof, M.; Vet, L.E.M.; Gols, R. The ‘usurpation hypothesis’ revisited: Dying caterpillar repels attack from a hyperparasitoid wasp. Anim. Behav. 2011, 81, 1281–1287. [Google Scholar] [CrossRef]
- Mohan, P.; Sinu, P.A. Parasitoid wasp usurps its host to guard its pupa against hyperparasitoids and induces rapid behavioral changes in the parasitized host. PLoS ONE 2017, 12, e0178108. [Google Scholar] [CrossRef] [Green Version]
- Maure, F.; Brodeur, J.; Ponlet, N.; Doyon, J.; Firlej, A.; Elguero, E.; Thomas, F. The cost of a bodyguard. Biol. Lett. 2011, 7, 843–846. [Google Scholar] [CrossRef] [PubMed]
- Maure, F.; Brodeur, J.; Droit, A.; Doyon, J.; Thomas, F. Bodyguard manipulation in a multipredator context: Different processes, same effect. Behav. Process. 2013, 99, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Dheilly, N.M.; Maure, F.; Ravallec, M.; Galinier, R.; Doyon, J.; Duval, D.; Leger, L.; Volkoff, A.N.; Missé, D.; Nidelet, S.; et al. Who is the puppet master? Replication of a parasitic wasp-associated virus correlates with host behaviour manipulation. Proc. R. Soc. B Biol. Sci. 2015, 282, 20142773. [Google Scholar] [CrossRef] [Green Version]
- Gonzaga, M.O.; Kloss, T.G.; Sobczak, J.F. Host Behavioural Manipulation of Spiders by Ichneumonid Wasps. In Behaviour and Ecology of Spiders: Contributions from the Neotropical Region; Viera, C., Gonzaga, M.O., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 417–437. [Google Scholar] [CrossRef]
- Weinersmith, K.L. What’s gotten into you? A review of recent research on parasitoid manipulation of host behavior. Curr. Opin. Insect Sci. 2019, 33, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Weinersmith, K.L.; Liu, S.M.; Forbes, A.A.; Egan, S.P. Tales from the crypt: A parasitoid manipulates the behaviour of its parasite host. Proc. R. Soc. B Biol. Sci. 2017, 284, 20162365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Marle, J.; Piek, T. Morphology of the Venom Apparatus. In Venoms of the Hymenoptera: Biochemical, Pharmacological and Behavioural Aspects; Academic Press: London, UK, 1986; pp. 17–44. [Google Scholar] [CrossRef]
- Steiner, A.L. Stinging behaviour of solitary wasps. In Venoms of the Hymenoptera: Biochemical, Pharmacological and Behavioural Aspects; Academic Press: London, UK, 1986; pp. 63–160. [Google Scholar] [CrossRef]
- Williams, F.X. Ampulex compressa (Fabr.), a cockraoch-hunting wasp introduced from New Caledonia into Hawaii. Proc. Hawaii. Entomol. Soc. 1942, 11, 221–233. [Google Scholar]
- Fouad, K.; Rathmayer, W.; Libersat, F. Neuromodulation of the escape behavior of the cockroach Periplaneta americana by the venom of the parasitic wasp Ampulex compressa. J. Comp. Physiol. A 1996, 178, 91–100. [Google Scholar] [CrossRef]
- Catania, K. Getting the most out of your zombie: Abdominal sensors and neural manipulations help jewel wasps find the roach’s weak spot. Brain Behav. Evol. 2020, 95, 181–202. [Google Scholar] [CrossRef] [PubMed]
- Piek, T.; Spanjer, W. Chemistry and pharmacology of solitary wasp venoms. In Venoms of the Hymenoptera: Biochemical, Pharmacological and Behavioural Aspects; Academic Press: London, UK, 1986; pp. 161–307. [Google Scholar] [CrossRef]
- Poirié, M.; Colinet, D.; Gatti, J.L. Insights into function and evolution of parasitoid wasp venoms. Curr. Opin. Insect Sci. 2014, 6, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Bland, R.G.; Jaques, H.E. How to Know the Insects; Waveland Press: Long Grove, IL, USA, 2010. [Google Scholar]
- Banks, B.E.; Shipolini, R.A. Chemistry and pharmacology of honey-bee venom. In Venoms of the Hymenoptera: Biochemical, Pharmacological and Behavioural Aspects; Academic Press: London, UK, 1986; pp. 329–416. [Google Scholar]
- Dotimas, E.M.; Hider, R.C. Honeybee venom. Bee World 1987, 68, 51–70. [Google Scholar] [CrossRef]
- De Lima, P.R.; Brochetto-Braga, M.R. Hymenoptera venom review focusing on Apis mellifera. J. Venom. Anim. Toxins Incl. Trop. Dis. 2003, 9, 149–162. [Google Scholar]
- Azam, M.N.K.; Ahmed, M.N.; Biswas, S.; Ara, N.; Rahman, M.M.; Hirashima, A.; Hasan, M.N. A review on bioactivities of honey bee venom. Annu. Res. Rev. Biol. 2018, 30, 1–13. [Google Scholar] [CrossRef]
- Moreau, S.J.M.; Asgari, S. Venom proteins from parasitoid wasps and their biological functions. Toxins 2015, 7, 2385–2412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mrinalini, W.J.; Werren, J.H. Parasitoid wasps and their venoms. In Evolution of Venomous Animals and Their Toxins; Toxinology; Gopalakrishnakone, P., Malhotra, A., Eds.; Springer: Dordrecht, The Netherlands, 2017; pp. 187–212. [Google Scholar] [CrossRef]
- Williams, F.X. Studies in tropical wasps–their hosts and associates (with descriptions of new species). Bull. Exp. Stn. Hawaii. Sugar Plant. Assoc. 1928, 19, 1882–1967. [Google Scholar]
- Wasbauer, M.S.; Powell, J.A. Host records for some North American spider wasps, with notes on prey selection (Hymenoptera: Pompilidae). J. Kans. Entomol. Soc. 1962, 35, 393–401. [Google Scholar]
- Evans, H.E. Studies on neotropical Pompilidae (Hymenoptera) V. Austrochares Banks. Psyche J. Entomol. 1969, 76, 18–28. [Google Scholar] [CrossRef]
- Kurczewski, F.E.; Spofford, M.G. Observations on the behaviors of some Scoliidae and Pompilidae (Hymenoptera) in Florida. Fla. Entomol. 1986, 69, 636. [Google Scholar] [CrossRef]
- Fitton, M.G.; Shaw, M.R.; Austin, A.D. The Hymenoptera associated with spiders in Europe. Zool. J. Linn. Soc. 1987, 90, 65–93. [Google Scholar] [CrossRef] [Green Version]
- Harris, A.C. Pompilidae (Insecta: Hymenoptera); Number 12 in Fauna of New Zealand; Science Information Pub. Centre, DSIR: Wellington, New Zealand, 1987. [Google Scholar]
- Day, M.C. Spider Wasps: Hymenoptera: Pompilidae. In Handbooks for the Identification of British Insects; British Museum (Natural History): London, UK, 1988; Volume 6. [Google Scholar]
- Elliott, M.G. Annotated catalogue of the Pompilidae (Hymenoptera) of Australia. Zootaxa 2007, 1428, 1–83. [Google Scholar] [CrossRef]
- Loktionov, V.M.; Lelej, A.S. An annotated catalogue of the spider wasps (Hymenoptera: Pompilidae) of Russia. Zootaxa 2017, 4280, 1–95. [Google Scholar] [CrossRef] [Green Version]
- Williams, F.X. Life History Studies of Pepsis and Hemipepsis Wasps California (Hymenoptera, Pompilidae). Ann. Entomol. Soc. Am. 1956, 49, 447–466. [Google Scholar] [CrossRef] [Green Version]
- Evans, H.E. The ecology and nesting behaviour of the Pompilidae (Hymenoptera) of the northeastern United States. Misc. Publ. Entomol. Soc. Am. 1962, 3, 63–119. [Google Scholar]
- Cazier, M.A.; Mortenson, M.A. Bionomical observations on tarantula-hawks and their prey (Hymenoptera: Pompilidae: Pepsis). Ann. Entomol. Soc. Am. 1964, 57, 533–541. [Google Scholar] [CrossRef]
- Shimizu, A. Nesting behavior of the semi-aquatic spider wasp, Anoplius eous, which transports its prey on the surface film of water (Hymenoptera, Pompilidae). J. Ethol. 1992, 10, 85–102. [Google Scholar] [CrossRef]
- Punzo, F. The biology of the spider wasp Pepsis thisbe (Hymenoptera: Pompilidae) from Trans Pecos, Texas. I. Adult morphometrics, larval development and the ontogeny of larval feeding patterns. Psyche J. Entomol. 1994, 101, 229–241. [Google Scholar] [CrossRef] [Green Version]
- Punzo, F. The biology of the spider wasp, Pepsis thisbe (Hymenoptera: Pompilidae) from Trans Pecos, Texas. II. Temporal patterns of activity and hunting behavior with special reference to the effects of experience. Psyche J. Entomol. 1994, 101, 243–256. [Google Scholar] [CrossRef] [Green Version]
- Punzo, F. Studies on the natural history, ecology, and behavior of Pepsis cerberus and P. mexicana (Hymenoptera: Pompilidae) from Big Bend National Park, Texas. Entomol. Am. 2005, 113, 84–95. [Google Scholar] [CrossRef]
- Kurczewski, F.E.; Edwards, G.B. Hosts, nesting behavior, and ecology of some North American spider wasps (Hymenoptera: Pompilidae). Southeast. Nat. 2012, 11, 1–71. [Google Scholar] [CrossRef] [Green Version]
- Waichert, C.; Rodriguez, J.; Von Dohlen, C.D.; Pitts, J.P. Spider wasps (Hymenoptera: Pompilidae) of the Dominican Republic. Zootaxa 2012, 3353, 1–47. [Google Scholar] [CrossRef]
- Kephart, S.R. The Floral Ecology and Reproductive Isolation of Three Sympatric Species of Asclepias. Ph.D. Thesis, Indiana University, Bloomington, IN, USA, 1979. [Google Scholar]
- Forster, P.I. Diurnal insects associated with the flowers of Gomphocarpus physocarpus E. Mey. (Asclepiadaceae), an introduced weed in Australia. Biotropica 1994, 26, 214–217. [Google Scholar] [CrossRef]
- Punzo, F. Plants whose flowers are utilized by adults of Pepsis grossa Fabricius (Hymenoptera: Pompilidae) as a source of nectar. J. Hymenopt. Res. 2006, 15, 171–176. [Google Scholar]
- Steiner, K.E.; Whitehead, V.B.; Johnson, S.D. Floral and pollinator divergence in two sexually deceptive South African orchids. Am. J. Bot. 1994, 81, 185–194. [Google Scholar] [CrossRef]
- Johnson, S.D. Specialized pollination by spider-hunting wasps in the African orchid Disa sankeyi. Plant Syst. Evol. 2005, 251, 153–160. [Google Scholar] [CrossRef]
- Shuttleworth, A.; Johnson, S.D. Specialized pollination by large spider-hunting wasps and self-incompatibility in the African milkweed Pachycarpus asperifolius. Int. J. Plant Sci. 2006, 167, 1177–1186. [Google Scholar] [CrossRef]
- Shuttleworth, A.; Johnson, S.D. A key role for floral scent in a wasp-pollination system in Eucomis (Hyacinthaceae). Ann. Bot. 2009, 103, 715–725. [Google Scholar] [CrossRef] [Green Version]
- Shuttleworth, A.; Johnson, S.D. The importance of scent and nectar filters in a specialized wasp-pollination system. Funct. Ecol. 2009, 23, 931–940. [Google Scholar] [CrossRef]
- Shuttleworth, A.; Johnson, S.D. The Hemipepsis wasp-pollination system in South Africa: A comparative analysis of trait convergence in a highly specialized plant guild. Bot. J. Linn. Soc. 2012, 168, 278–299. [Google Scholar] [CrossRef]
- Pupulin, F.; Bogarín, D. The genus Brassia in Costa Rica: A survey of four species and a new species. Orchids 2005, 74, 202–207. [Google Scholar]
- Wiemer, A.P.; Sérsic, A.N.; Marino, S.; Simões, A.O.; Cocucci, A.A. Functional morphology and wasp pollination of two South American asclepiads (Asclepiadoideae–Apocynaceae). Ann. Bot. 2012, 109, 77–93. [Google Scholar] [CrossRef] [Green Version]
- Phillips, R.D.; Bohman, B.; Peakall, R. Pollination by nectar-foraging pompilid wasps: A new specialized pollination strategy for the Australian flora. Plant Biol. 2021. [Google Scholar] [CrossRef]
- Schmidt, J.O. Venom and the good life in tarantula hawks (Hymenoptera: Pompilidae): How to eat, not be eaten, and live long. J. Kans. Entomol. Soc. 2004, 77, 402–413. [Google Scholar] [CrossRef]
- Schmidt, J.O. The Sting of the Wild; Johns Hopkins University Press: Baltimore, MD, USA, 2016. [Google Scholar]
- Lincecum, G. The tarantula killers of Texas. Am. Nat. 1867, 1, 137–141. [Google Scholar] [CrossRef]
- Iwata, K. Comparative studies on the habits of solitary wasps. Tenthredo 1942, 4, 1–146. [Google Scholar]
- Petrunkevitch, A. The spider and the wasp. Sci. Am. 1952, 187, 20–23. [Google Scholar] [CrossRef]
- Waichert, C.; Rodriguez, J.; Wasbauer, M.S.; von Dohlen, C.D.; Pitts, J.P. Molecular phylogeny and systematics of spider wasps (Hymenoptera: Pompilidae): Redefining subfamily boundaries and the origin of the family. Zool. J. Linn. Soc. 2015, 175, 271–287. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, J.; Pitts, J.P.; Florez, J.A.; Bond, J.E.; von Dohlen, C.D. Molecular phylogeny of Pompilinae (Hymenoptera: Pompilidae): Evidence for rapid diversification and host shifts in spider wasps. Mol. Phylogenetics Evol. 2016, 94, 55–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peckham, G.W.; Peckham, E.G. On the Instincts and Habits of the Solitary Wasps; Wisconsin Geological and Natural History Survey: Madison, WI, USA, 1898; Volume 1. [Google Scholar]
- Grout, D. Behaviour of a parasitic pompilid wasp (Hymenoptera). J. Entomol. Soc. S. Afr. 1982, 45, 217–220. [Google Scholar]
- Costa, F.G.; Pérez-Miles, F.; Mignone, A. Pompilid wasp interactions with burrowing tarantulas: Pepsis cupripennis versus Eupalaestrus weijenberghi and Acanthoscurria suina (Araneae, Theraphosidae). Stud. Neotrop. Fauna Environ. 2004, 39, 37–43. [Google Scholar] [CrossRef]
- Yamamoto, T.; Arimoto, H.; Kinumi, T.; Oba, Y.; Uemura, D. Identification of proteins from venom of the paralytic spider wasp, Cyphononyx dorsalis. Insect Biochem. Mol. Biol. 2007, 37, 278–286. [Google Scholar] [CrossRef]
- Jensen, T.; Walker, A.A.; Nguyen, S.H.; Jin, A.H.; Deuis, J.R.; Vetter, I.; King, G.F.; Schmidt, J.O.; Robinson, S.D. Venom chemistry underlying the painful stings of velvet ants (Hymenoptera: Mutillidae). Cell. Mol. Life Sci. 2021. [Google Scholar] [CrossRef] [PubMed]
- Richards, O.W.; Hamm, A.H. The biology of British Pompilidae (Hymenoptera). Trans. Soc. Br. Entomol. 1939, 6, 51–114. [Google Scholar]
- Evans, H.E.; Lin, C.S.; Yoshimoto, C.M. A biological study of Anoplius apiculatus autumnalis (Banks) Its Parasite, Evagetes mohave (Banks) (Hymenoptera, Pompilidae). J. N. Y. Entomol. Soc. 1953, 61, 61–78. [Google Scholar]
- Evans, H.E. Comparative ethology and the systematics of spider wasps. Syst. Zool. 1953, 2, 155–172. [Google Scholar] [CrossRef]
- Kurczewski, F.E.; Kurczewski, E.J. Host records for some North American Pompilidae (Hymenoptera). Third Supplement. Tribe Pompilini. J. Kans. Entomol. Soc. 1973, 46, 65–81. [Google Scholar]
- Gwynne, D.T. Nesting biology of the spider wasps (Hymenoptera: Pompilidae) which prey on burrowing wolf spiders (Araneae: Lycosidae, Geolycosa). J. Nat. Hist. 1979, 13, 681–692. [Google Scholar] [CrossRef]
- Kurczewski, F.E.; Kurczewski, E.J. Host records for some North American Pompilidae (Hymenoptera). First supplement. J. Kans. Entomol. Soc. 1968, 41, 367–382. [Google Scholar]
- Kurczewski, F.E.; Kurczewski, E.J. Host records for some North American Pompilidae (Hymenoptera) with a discussion of factors in prey selection. J. Kans. Entomol. Soc. 1968, 41, 1–33. [Google Scholar]
- Petrunkevitch, A. Tarantula versus tarantula-hawk: A study in instinct. J. Exp. Zool. 1926, 45, 367–397. [Google Scholar] [CrossRef]
- Minkiewicz, R. Les Pompilides à nid fixe et ceux à nid momentané: (étude d’éthologie comparée). Pol. Pismo Entomol. 1934, 13, 43–60. [Google Scholar]
- Piek, T. A toxinological argument in favour of the close relationship of the Vespidae and the Scoliidae (Hymenoptera). Entomol. Ber. 1987, 47, 96–98. [Google Scholar]
- Piek, T.; Schmidt, J.O.; de Jong, J.M.; Mantel, P. Kinins in ant venoms—A comparison with venoms of related hymenoptera. Comp. Biochem. Physiol. Part C Comp. Pharmacol. 1989, 92, 117–124. [Google Scholar] [CrossRef]
- Piek, T. Neurotoxins from venoms of the hymenoptera—Twenty-five years of research in Amsterdam. Comp. Biochem. Physiol. Part C Comp. Pharmacol. 1990, 96, 223–233. [Google Scholar] [CrossRef]
- Schoeters, E.; Billen, J.; Schmidt, J.O. Venom gland morphology in Pepsis pallidolimbata pallidolimbata and biological use and activity of Pepsis venom. Can. J. Zool. 1997, 75, 1014–1019. [Google Scholar] [CrossRef]
- Konno, K.; Miwa, A.; Takayama, H.; Hisada, M.; Itagaki, Y.; Naoki, H.; Yasuhara, T.; Kawai, N. α-Pompilidotoxin (α-PMTX), a novel neurotoxin from the venom of a solitary wasp, facilitates transmission in the crustacean neuromuscular synapse. Neurosci. Lett. 1997, 238, 99–102. [Google Scholar] [CrossRef]
- Konno, K.; Hisada, M.; Itagaki, Y.; Naoki, H.; Kawai, N.; Miwa, A.; Yasuhara, T.; Takayama, H. Isolation and structure of pompilidotoxins, novel peptide neurotoxins in solitary wasp venoms. Biochem. Biophys. Res. Commun. 1998, 250, 612–616. [Google Scholar] [CrossRef]
- Konno, K.; Kazuma, K.; Nihei, K.I. Peptide toxins in solitary wasp venoms. Toxins 2016, 8, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahara, Y.; Gotoh, M.; Konno, K.; Miwa, A.; Tsubokawa, H.; Robinson, H.P.C.; Kawai, N. A new class of neurotoxin from wasp venom slows inactivation of sodium current: Slowing of Na channel inactivation by a novel wasp toxin. Eur. J. Neurosci. 2000, 12, 1961–1970. [Google Scholar] [CrossRef]
- Harsch, A.; Konno, K.; Takayama, H.; Kawai, N.; Robinson, H. Effects of α-pompilidotoxin on synchronized firing in networks of rat cortical neurons. Neurosci. Lett. 1998, 252, 49–52. [Google Scholar] [CrossRef]
- Yokota, H.; Tsubokawa, H.; Miyawaki, T.; Konno, K.; Nakayama, H.; Masuzawa, T.; Kawai, N. Modulation of synaptic transmission in hippocampal CA1 neurons by a novel neurotoxin (β-pompilidotoxin) derived from wasp venom. Neurosci. Res. 2001, 41, 365–371. [Google Scholar] [CrossRef]
- Kinoshita, E.; Maejima, H.; Yamaoka, K.; Konno, K.; Kawai, N.; Shimizu, E.; Yokote, S.; Nakayama, H.; Seyama, I. Novel wasp toxin discriminates between neuronal and cardiac sodium channels. Mol. Pharmacol. 2001, 59, 1457–1463. [Google Scholar] [CrossRef] [Green Version]
- Schiavon, E.; Stevens, M.; Zaharenko, A.J.; Konno, K.; Tytgat, J.; Wanke, E. Voltage-gated sodium channel isoform-specific effects of pompilidotoxins: Pompilidotoxins and voltage-gated NaV isoforms. FEBS J. 2010, 277, 918–930. [Google Scholar] [CrossRef] [PubMed]
- Konno, K.; Hisada, M.; Naoki, H.; Itagaki, Y.; Yasuhara, T.; Nakata, Y.; Miwa, A.; Kawai, N. Molecular determinants of binding of a wasp toxin (PMTXs) and its analogs in the Na+ channels proteins. Neurosci. Lett. 2000, 285, 29–32. [Google Scholar] [CrossRef]
- Kawai, N.; Konno, K. Molecular determinants of two neurotoxins that regulate sodium current inactivation in rat hippocampal neurons. Neurosci. Lett. 2004, 361, 44–46. [Google Scholar] [CrossRef]
- Grieco, T.M.; Raman, I.M. Production of resurgent current in NaV1.6-null Purkinje neurons by slowing sodium channel inactivation with β-pompilidotoxin. J. Neurosci. 2004, 24, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Nakayama, H.; Yokote, S.; Setoguchi, R.; Shimizu, E.; Kawahara, K.; Kuniyasu, A.; Shirasaki, T.; Takahama, K.; Kawai, N.; Yamaoka, K.; et al. A synthetic approach to develop peptide inhibitors selective for brain-type sodium channels on the basis of pompilidotoxin structure. HeteroCycles 2009, 79, 925. [Google Scholar] [CrossRef]
- Yu, F.H.; Catterall, W.A. Overview of the voltage-gated sodium channel family. Genome Biol. 2003, 4, 207. [Google Scholar] [CrossRef] [Green Version]
- Fux, J.E.; Mehta, A.; Moffat, J.; Spafford, J.D. Eukaryotic voltage-gated sodium channels: On their origins, asymmetries, losses, diversification and adaptations. Front. Physiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, F.C.; Lewis, R.J. Sodium channels and pain: From toxins to therapies. Br. J. Pharmacol. 2018, 175, 2138–2157. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Ma, H.; Zhang, F.; Zhang, C.; Zou, X.; Cao, Z. Selective voltage-gated sodium channel peptide toxins from animal venom: Pharmacological probes and analgesic drug development. ACS Chem. Neurosci. 2018, 9, 187–197. [Google Scholar] [CrossRef]
- Konno, K.; Hisada, M.; Fontana, R.; Lorenzi, C.C.B.; Naoki, H.; Itagaki, Y.; Miwa, A.; Kawai, N.; Nakata, Y.; Yasuhara, T.; et al. Anoplin, a novel antimicrobial peptide from the venom of the solitary wasp Anoplius samariensis. Biochim. Biophys. Acta (BBA) Protein Struct. Mol. Enzymol. 2001, 1550, 70–80. [Google Scholar] [CrossRef]
- Dos Santos Cabrera, M.P.; Arcisio-Miranda, M.; Broggio Costa, S.T.; Konno, K.; Ruggiero, J.R.; Procopio, J.; Ruggiero Neto, J.a. Study of the mechanism of action of anoplin, a helical antimicrobial decapeptide with ion channel-like activity, and the role of the amidated C-terminus. J. Pept. Sci. 2008, 14, 661–669. [Google Scholar] [CrossRef]
- Ifrah, D.; Doisy, X.; Ryge, T.S.; Hansen, P.R. Structure-activity relationship study of anoplin. J. Pept. Sci. 2005, 11, 113–121. [Google Scholar] [CrossRef]
- Won, A.; Pripotnev, S.; Ruscito, A.; Ianoul, A. Effect of point mutations on the secondary structure and membrane interaction of antimicrobial peptide anoplin. J. Phys. Chem. B 2011, 115, 2371–2379. [Google Scholar] [CrossRef] [PubMed]
- Munk, J.K.; Uggerhøj, L.E.; Poulsen, T.J.; Frimodt-Møller, N.; Wimmer, R.; Nyberg, N.T.; Hansen, P.R. Synthetic analogs of anoplin show improved antimicrobial activities. J. Pept. Sci. 2013, 19, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Munk, J.K.; Ritz, C.; Uggerhøj, L.E.; Poulsen, T.J.; Wimmer, R.; Frimodt-Møller, N.; Hansen, P.R. Statistical Design and Analysis of Anoplin Sequence Chemical Space. In Biopolymers; Wiley-Blackwell: Hoboken, NJ, USA, 2013; Volume 100, p. 256. [Google Scholar]
- Slootweg, J.C.; van Schaik, T.B.; van Ufford, H.L.C.Q.; Breukink, E.; Liskamp, R.M.; Rijkers, D.T. Improving the biological activity of the antimicrobial peptide anoplin by membrane anchoring through a lipophilic amino acid derivative. Bioorganic Med. Chem. Lett. 2013, 23, 3749–3752. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, J.; Zheng, X.; Yang, X.; Ma, P.; Cai, Y.; Zhang, B.; Chen, Y. Design of novel analogues of short antimicrobial peptide anoplin with improved antimicrobial activity. J. Pept. Sci. 2014, 20, 945–951. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhu, N.; Zhong, C.; Zhu, Y.; Gou, S.; Chang, L.; Bao, H.; Liu, H.; Zhang, Y.; Ni, J. Effect of N-methylated and fatty acid conjugation on analogs of antimicrobial peptide Anoplin. Eur. J. Pharm. Sci. 2020, 152, 105453. [Google Scholar] [CrossRef]
- Wu, Y.; Huang, R.; Jin, J.M.; Zhang, L.J.; Zhang, H.; Chen, H.Z.; Chen, L.L.; Luan, X. Advances in the study of structural modification and biological activities of anoplin. Front. Chem. 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Jindřichová, B.; Burketová, L.; Novotná, Z. Novel properties of antimicrobial peptide anoplin. Biochem. Biophys. Res. Commun. 2014, 444, 520–524. [Google Scholar] [CrossRef]
- Sher, D.; Fishman, Y.; Zhang, M.; Lebendiker, M.; Gaathon, A.; Mancheño, J.M.; Zlotkin, E. Hydralysins, a new category of β-pore-forming toxins in Cnidaria. J. Biol. Chem. 2005, 280, 22847–22855. [Google Scholar] [CrossRef] [Green Version]
- Dreon, M.S.; Frassa, M.V.; Ceolín, M.; Ituarte, S.; Qiu, J.W.; Sun, J.; Fernández, P.E.; Heras, H. Novel animal defenses against predation: A snail egg neurotoxin combining lectin and pore-forming chains that resembles plant defense and bacteria attack toxins. PLoS ONE 2013, 8, e63782. [Google Scholar] [CrossRef] [Green Version]
- Ellisdon, A.M.; Reboul, C.F.; Panjikar, S.; Huynh, K.; Oellig, C.A.; Winter, K.L.; Dunstone, M.A.; Hodgson, W.C.; Seymour, J.; Dearden, P.K.; et al. Stonefish toxin defines an ancient branch of the perforin-like superfamily. Proc. Natl. Acad. Sci. USA 2015, 112, 15360–15365. [Google Scholar] [CrossRef] [Green Version]
- Blake, K.J.; Baral, P.; Voisin, T.; Lubkin, A.; Pinho-Ribeiro, F.A.; Adams, K.L.; Roberson, D.P.; Ma, Y.C.; Otto, M.; Woolf, C.J.; et al. Staphylococcus aureus produces pain through pore-forming toxins and neuronal TRPV1 that is silenced by QX-314. Nat. Commun. 2018, 9, 37. [Google Scholar] [CrossRef] [PubMed]
- Picolo, G.; Hisada, M.; Moura, A.B.; Machado, M.F.M.; Sciani, J.M.; Conceição, I.M.; Melo, R.L.; Oliveira, V.; Lima-Landman, M.T.R.; Cury, Y.; et al. Bradykinin-related peptides in the venom of the solitary wasp Cyphononyx fulvognathus. Biochem. Pharmacol. 2010, 79, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Kazandjian, T.D.; Petras, D.; Robinson, S.D.; Thiel, J.V.; Greene, H.W.; Arbuckle, K.; Barlow, A.; Carter, D.A.; Wouters, R.M.; Whiteley, G.; et al. Convergent evolution of pain-inducing defensive venom components in spitting cobras. Science 2021, 371, 386–390. [Google Scholar] [CrossRef]
- Piek, T.; Hue, B.; Mantel, P.; Nakajima, T.; Pelhate, M.; Yasuhara, T. Threonine6-bradykinin in the venom of the wasp Colpa interrupta (F.) presynaptically blocks nicotinic synaptic transmission in the insect CNS. Comp. Biochem. Physiol. Part C Comp. Pharmacol. 1990, 96, 157–162. [Google Scholar] [CrossRef]
- Hisada, M.; Satake, H.; Masuda, K.; Aoyama, M.; Murata, K.; Shinada, T.; Iwashita, T.; Ohfune, Y.; Nakajima, T. Molecular components and toxicity of the venom of the solitary wasp, Anoplius samariensis. Biochem. Biophys. Res. Commun. 2005, 330, 1048–1054. [Google Scholar] [CrossRef]
- Hisada, M.; Konno, K.; Itagaki, Y.; Naoki, H.; Nakajima, T. Sequencing wasp venom peptides by endopeptidase digestion and nested collision-induced dissociation/post-source decay methods. Rapid Commun. Mass Spectrom. 2002, 16, 1040–1048. [Google Scholar] [CrossRef]
- Huicab-Uribe, M.; Verdel-Aranda, K.; Martínez-Hernández, A.; Zamudio, F.; Jiménez-Vargas, J.; Lara-Reyna, J. Molecular composition of the paralyzing venom of three solitary wasps (Hymenoptera: Pompilidae) collected in southeast Mexico. Toxicon 2019, 168, 98–102. [Google Scholar] [CrossRef]
- Nolasco, M.; Biondi, I.; Pimenta, D.C.; Branco, A. Extraction and preliminary chemical characterization of the venom of the spider wasp Pepsis decorata (Hymenoptera: Pompilidae). Toxicon 2018, 150, 74–76. [Google Scholar] [CrossRef]
- Walker, A.A.; Robinson, S.D.; Hamilton, B.F.; Undheim, E.A.B.; King, G.F. Deadly proteomes: A practical guide to proteotranscriptomics of animal venoms. PROTEOMICS 2020, 20, 1900324. [Google Scholar] [CrossRef]
- Calvete, J.J.; Lomonte, B.; Saviola, A.J.; Bonilla, F.; Sasa, M.; Williams, D.J.; Undheim, E.A.B.; Sunagar, K.; Jackson, T.N.W. Mutual enlightenment: A toolbox of concepts and methods for integrating evolutionary and clinical toxinology via snake venomics and the contextual stance. Toxicon X 2021, 9-10, 100070. [Google Scholar] [CrossRef]
- Hisada, M.; Konno, K.; Itagaki, Y.; Naoki, H.; Nakajima, T. Advantages of using nested collision induced dissociation/post-source decay with matrix-assisted laser desorption/ionization time-of-flight mass spectrometry: Sequencing of novel peptides from wasp venom. Rapid Commun. Mass Spectrom. 2000, 14, 1828–1834. [Google Scholar] [CrossRef]
- Hisada, M. Sequencing novel peptides from solitary wasps by nested CID/PSD. J. Mass Spectrom. Soc. Jpn. 2003, 51, 310–316. [Google Scholar] [CrossRef]
- Daly, N.L.; Wilson, D. Structural diversity of arthropod venom toxins. Toxicon 2018, 152, 46–56. [Google Scholar] [CrossRef]
- Aird, S.D. Ophidian envenomation strategies and the role of purines. Toxicon 2002, 40, 335–393. [Google Scholar] [CrossRef]
- Villar-Briones, A.; Aird, S.D. Organic and peptidyl constituents of snake venoms: The picture is vastly more complex than we imagined. Toxins 2018, 10, 392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, E.R.J.; McIntyre, L.; Northfield, T.D.; Daly, N.L.; Wilson, D.T. Small molecules in the venom of the scorpion Hormurus waigiensis. Biomedicines 2020, 8, 259. [Google Scholar] [CrossRef]
- Rodriguez, J.; Waichert, C.; von Dohlen, C.D.; Pitts, J.P. The geological record and phylogeny of spider wasps (Hymenoptera: Pompilidae): A revision of fossil species and their phylogenetic placement. PLoS ONE 2017, 12, e0185379. [Google Scholar] [CrossRef] [Green Version]
- Gilchrist, J.; Olivera, B.M.; Bosmans, F. Animal toxins influence voltage-gated sodium channel function. Handb. Exp. Pharmacol. 2014, 221, 203–229. [Google Scholar] [CrossRef] [Green Version]
- Moran, Y.; Gordon, D.; Gurevitz, M. Sea anemone toxins affecting voltage-gated sodium channels—Molecular and evolutionary features. Toxicon 2009, 54, 1089–1101. [Google Scholar] [CrossRef] [Green Version]
- Jouiaei, M.; Sunagar, K.; Federman Gross, A.; Scheib, H.; Alewood, P.F.; Moran, Y.; Fry, B.G. Evolution of an ancient venom: Recognition of a novel family of Cnidarian toxins and the common evolutionary origin of sodium and potassium neurotoxins in sea anemone. Mol. Biol. Evol. 2015, 32, 1598–1610. [Google Scholar] [CrossRef] [Green Version]
- Jouiaei, M.; Yanagihara, A.A.; Madio, B.; Nevalainen, T.J.; Alewood, P.F.; Fry, B.G. Ancient venom systems: A review on Cnidaria toxins. Toxins 2015, 7, 2251–2271. [Google Scholar] [CrossRef] [Green Version]
- Eitan, M.; Fowler, E.; Herrmann, R.; Duval, A.; Pelhate, M.; Zlotkin, E. A scorpion venom neurotoxin paralytic to insects that affects sodium current inactivation: Purification, primary structure, and mode of action. Biochemistry 1990, 29, 5941–5947. [Google Scholar] [CrossRef]
- Karbat, I.; Frolow, F.; Froy, O.; Gilles, N.; Cohen, L.; Turkov, M.; Gordon, D.; Gurevitz, M. Molecular basis of the high insecticidal potency of scorpion α-toxins. J. Biol. Chem. 2004, 279, 31679–31686. [Google Scholar] [CrossRef] [Green Version]
- Bosmans, F.; Tytgat, J. Voltage-gated sodium channel modulation by scorpion α-toxins. Toxicon 2007, 49, 142–158. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, J.I.; Chapman, B.E.; Mackay, J.P.; Howden, M.E.; King, G.F. The structure of versutoxin (δ-atracotoxin-Hv1) provides insights into the binding of site 3 neurotoxins to the voltage-gated sodium channel. Structure 1997, 5, 1525–1535. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Diao, J.; Li, J.; Tang, J.; Lin, Y.; Hu, W.; Zhang, Y.; Xiao, Y.; Liang, S. JZTX-IV, a unique acidic sodium channel toxin isolated from the spider Chilobrachys jingzhao. Toxicon 2008, 52, 871–880. [Google Scholar] [CrossRef]
- Shon, K.J.; Grilley, M.M.; Marsh, M.; Yoshikami, D.; Hall, A.R.; Kurz, B.; Gray, W.R.; Imperial, J.S.; Hillyard, D.R.; Olivera, B.M. Purification, characterization, synthesis, and cloning of the lockjaw peptide from Conus purpurascens venom. Biochemistry 1995, 34, 4913–4918. [Google Scholar] [CrossRef] [PubMed]
- Green, B.R.; Olivera, B.M. Venom peptides from cone snails: Pharmacological probes for voltage-gated sodium channels. In Current Topics in Membranes; Na Channels from Phyla to, Function; French, R.J., Noskov, S.Y., Eds.; Academic Press: Cambridge, MA, USA, 2016; Volume 78, pp. 65–86. [Google Scholar] [CrossRef]
- Morales Duque, H.; Campos Dias, S.; Franco, O.L. Structural and functional analyses of cone snail toxins. Mar. Drugs 2019, 17, 370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.C.; Deuis, J.R.; Dashevsky, D.; Dobson, J.; Jackson, T.N.W.; Brust, A.; Xie, B.; Koludarov, I.; Debono, J.; Hendrikx, I.; et al. The snake with the scorpion’s sting: Novel three-finger toxin sodium channel activators from the venom of the long-glanded blue coral snake (Calliophis bivirgatus). Toxins 2016, 8, 303. [Google Scholar] [CrossRef] [PubMed]
- Dutertre, S.; Jin, A.H.; Vetter, I.; Hamilton, B.; Sunagar, K.; Lavergne, V.; Dutertre, V.; Fry, B.G.; Antunes, A.; Venter, D.J.; et al. Evolution of separate predation- and defence-evoked venoms in carnivorous cone snails. Nat. Commun. 2014, 5, 3521. [Google Scholar] [CrossRef] [Green Version]
- Walker, A.A.; Mayhew, M.L.; Jin, J.; Herzig, V.; Undheim, E.A.B.; Sombke, A.; Fry, B.G.; Meritt, D.J.; King, G.F. The assassin bug Pristhesancus plagipennis produces two distinct venoms in separate gland lumens. Nat. Commun. 2018, 9, 755. [Google Scholar] [CrossRef] [Green Version]
- Robertson, P. A morphological and functional study of the venom apparatus in representatives of some major groups of Hymenoptera. Aust. J. Zool. 1968, 16, 133. [Google Scholar] [CrossRef]
- Kumpanenko, A.S.; Gladun, D.V. Functional morphology of the sting apparatus of the spider wasp Cryptocheilus versicolor (Scopoli, 1763) (Hymenoptera: Pompilidae): The sting of Cryptocheilus versicolor. Entomol. Sci. 2018, 21, 124–132. [Google Scholar] [CrossRef]
- Schendel, V.; Rash, L.D.; Jenner, R.A.; Undheim, E.A.B. The diversity of venom: The importance of behavior and venom system morphology in understanding its ecology and evolution. Toxins 2019, 11, 666. [Google Scholar] [CrossRef] [Green Version]
- Robinson, S.D.; Mueller, A.; Clayton, D.; Starobova, H.; Hamilton, B.R.; Payne, R.J.; Vetter, I.; King, G.F.; Undheim, E.A.B. A comprehensive portrait of the venom of the giant red bull ant, Myrmecia gulosa, reveals a hyperdiverse hymenopteran toxin gene family. Sci. Adv. 2018, 4, eaau4640. [Google Scholar] [CrossRef] [Green Version]
- Jackson, T.N.W.; Koludarov, I. How the toxin got its toxicity. Front. Pharmacol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Martinson, E.O.; Mrinalini.; Kelkar, Y.D.; Chang, C.H.; Werren, J.H. The evolution of venom by co-option of single-copy genes. Curr. Biol. 2017, 27, 2007–2013.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fletcher, J.I.; Smith, R.; O’Donoghue, S.I.; Nilges, M.; Connor, M.; Howden, M.E.H.; Christie, M.J.; King, G.F. The structure of a novel insecticidal neurotoxin, ω-atracotoxin-HV1, from the venom of an Australian funnel web spider. Nat. Struct. Biol. 1997, 4, 559–566. [Google Scholar] [CrossRef]
- King, G.F. Tying pest insects in knots: The deployment of spider-venom-derived knottins as bioinsecticides. Pest Manag. Sci. 2019, 75, 2437–2445. [Google Scholar] [CrossRef]
- King, G.F.; Hardy, M.C. Spider-venom peptides: Structure, pharmacology, and potential for control of insect pests. Annu. Rev. Entomol. 2013, 58, 475–496. [Google Scholar] [CrossRef]
- Herzig, V.; King, G.F. The cystine knot is responsible for the exceptional stability of the insecticidal spider toxin ω-hexatoxin-Hv1a. Toxins 2015, 7, 4366–4380. [Google Scholar] [CrossRef] [Green Version]
- Yaninek, J.S.; Herren, H.R. Introduction and spread of the cassava green mite, Mononychellus tanajoa (Bondar) (Acari: Tetranychidae), an exotic pest in Africa and the search for appropriate control methods: A review. Bull. Entomol. Res. 1988, 78, 1–13. [Google Scholar] [CrossRef]
- Narayanaswamy, K.C.; Geethabai, M.; Raghuraman, R. Mite pests of mulberry—A review. Indian J. Seric. 1996, 35, 1–8. [Google Scholar]
- Collins, D.A. A review of alternatives to organophosphorus compounds for the control of storage mites. J. Stored Prod. Res. 2006, 42, 395–426. [Google Scholar] [CrossRef]
- Van Leeuwen, T.; Vontas, J.; Tsagkarakou, A.; Dermauw, W.; Tirry, L. Acaricide resistance mechanisms in the two-spotted spider mite Tetranychus urticae and other important Acari: A review. Insect Biochem. Mol. Biol. 2010, 40, 563–572. [Google Scholar] [CrossRef] [Green Version]
- Navia, D.; Gondim, M.G.C.; Aratchige, N.S.; de Moraes, G.J. A review of the status of the coconut mite, Aceria guerreronis (Acari: Eriophyidae), a major tropical mite pest. Exp. Appl. Acarol. 2013, 59, 67–94. [Google Scholar] [CrossRef]
- Idris, A.L.; Fan, X.; Muhammad, M.H.; Guo, Y.; Guan, X.; Huang, T. Ecologically controlling insect and mite pests of tea plants with microbial pesticides: A review. Arch. Microbiol. 2020, 202, 1–10. [Google Scholar] [CrossRef]
- Stocker, K.; Barlow, G.H. The coagulant enzyme from Bothrops atrox venom (batroxobin). In Methods in Enzymology; Part B: Proteolytic Enzymes; Academic Press: San Diego, CA, USA, 1976; Volume 45, pp. 214–223. [Google Scholar] [CrossRef]
- Vidt, D.G.; Bravo, E.L.; Fouad, F.M. Captopril. N. Engl. J. Med. 1982, 306, 214–219. [Google Scholar]
- Goa, K.L.; Noble, S. Eptifibatide. Drugs 1999, 57, 439–462. [Google Scholar] [CrossRef]
- Scarborough, R.M. Development of eptifibatide. Am. Heart J. 1999, 138, 1093–1104. [Google Scholar] [CrossRef]
- Curran, M.P.; Keating, G.M. Eptifibatide. Drugs 2005, 65, 2009–2035. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Xiao, P.; Chen, J.; Wei, Y. Are batroxobin agents effective for perioperative hemorrhage in thoracic surgery? A systematic review of randomized controlled trials. Blood Coagul. Fibrinolysis 2009, 20, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Kini, R.M. Toxins in thrombosis and haemostasis: Potential beyond imagination. J. Thromb. Haemost. 2011, 9, 195–208. [Google Scholar] [CrossRef]
- Lazarovici, P.; Marcinkiewicz, C.; Lelkes, P.I. From snake venom’s disintegrins and C-type lectins to anti-platelet drugs. Toxins 2019, 11, 303. [Google Scholar] [CrossRef] [Green Version]
- Mohamed Abd El-Aziz, T.; Soares, A.G.; Stockand, J.D. Snake venoms in drug discovery: Valuable therapeutic tools for life saving. Toxins 2019, 11, 564. [Google Scholar] [CrossRef] [Green Version]
- Davidson, M.B.; Bate, G.; Kirkpatrick, P. Exenatide. Nat. Rev. Drug Discov. 2005, 4, 713–714. [Google Scholar] [CrossRef]
- Warkentin, T.E.; Koster, A. Bivalirudin: A review. Expert Opin. Pharmacother. 2005, 6, 1349–1371. [Google Scholar] [CrossRef]
- Furman, B.L. The development of Byetta (exenatide) from the venom of the Gila monster as an anti-diabetic agent. Toxicon 2012, 59, 464–471. [Google Scholar] [CrossRef]
- Pope, J.E.; Deer, T.R. Ziconotide: A clinical update and pharmacologic review. Expert Opin. Pharmacother. 2013, 14, 957–966. [Google Scholar] [CrossRef]
- Alves, E.M.; Heneine, L.G.D.; Pesquero, J.L.; Merlo, L.D.A. Pharmaceutical Composition Containin an Apitoxin Fraction and Use Thereof. WO2011041865A1, 14 April 2011. [Google Scholar]
- Moreno, M.; Giralt, E. Three valuable peptides from bee and wasp venoms for therapeutic and biotechnological use: Melittin, apamin and mastoparan. Toxins 2015, 7, 1126–1150. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.S.; Pittler, M.H.; Shin, B.C.; Kong, J.C.; Ernst, E. Bee venom acupuncture for musculoskeletal pain: A review. J. Pain 2008, 9, 289–297. [Google Scholar] [CrossRef]
- Lee, J.A.; Son, M.J.; Choi, J.; Jun, J.H.; Kim, J.I.; Lee, M.S. Bee venom acupuncture for rheumatoid arthritis: A systematic review of randomised clinical trials. BMJ Open 2014, 4, e006140. [Google Scholar] [CrossRef] [PubMed]
- Panchal, R.; Smart, M.; Bowser, D.; Williams, D.; Petrou, S. Pore-forming proteins and their application in biotechnology. Curr. Pharm. Biotechnol. 2002, 3, 99–115. [Google Scholar] [CrossRef] [PubMed]
- Aliverdieva, D.A.; Efendieva, M.H.; Mamaev, D.V. Pore forming drugs: Antimicrobial mechanism and clinical applications. In Microbes in the Spotlight: Recent Progress in the Understanding of Beneficial and Harmful Microorganisms; Méndez-Vilas, A., Ed.; BrownWalker Press: Boca Raton, FL, USA, 2016; pp. 302–306. [Google Scholar]
- Ashrafuzzaman, M.; Tseng, C.Y.; Duszyk, M.; Tuszynski, J.A. Chemotherapy drugs form ion pores in membranes due to physical interactions with lipids. Chem. Biol. Drug Des. 2012, 80, 992–1002. [Google Scholar] [CrossRef]
- Elhilali, M.M.; Pommerville, P.; Yocum, R.C.; Merchant, R.; Roehrborn, C.G.; Denmeade, S.R. Prospective, randomized, double-blind, vehicle controlled, multicenter phase iib clinical trial of the pore forming protein PRX302 for targeted treatment of symptomatic benign prostatic hyperplasia. J. Urol. 2013, 189, 1421–1426. [Google Scholar] [CrossRef] [Green Version]
- Roy, A.; Bharadvaja, N. Venom-derived bioactive compounds as potential anticancer agents: A review. Int. J. Pept. Res. Ther. 2021, 27, 129–147. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dashevsky, D.; Rodriguez, J. A Short Review of the Venoms and Toxins of Spider Wasps (Hymenoptera: Pompilidae). Toxins 2021, 13, 744. https://doi.org/10.3390/toxins13110744
Dashevsky D, Rodriguez J. A Short Review of the Venoms and Toxins of Spider Wasps (Hymenoptera: Pompilidae). Toxins. 2021; 13(11):744. https://doi.org/10.3390/toxins13110744
Chicago/Turabian StyleDashevsky, Daniel, and Juanita Rodriguez. 2021. "A Short Review of the Venoms and Toxins of Spider Wasps (Hymenoptera: Pompilidae)" Toxins 13, no. 11: 744. https://doi.org/10.3390/toxins13110744
APA StyleDashevsky, D., & Rodriguez, J. (2021). A Short Review of the Venoms and Toxins of Spider Wasps (Hymenoptera: Pompilidae). Toxins, 13(11), 744. https://doi.org/10.3390/toxins13110744