Protective Effects of the Hydroethanolic Extract of Fridericia chica on Undifferentiated Human Neuroblastoma Cells Exposed to α-Zearalenol (α-ZEL) and β-Zearalenol (β-ZEL)


 , , ,
, , ,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
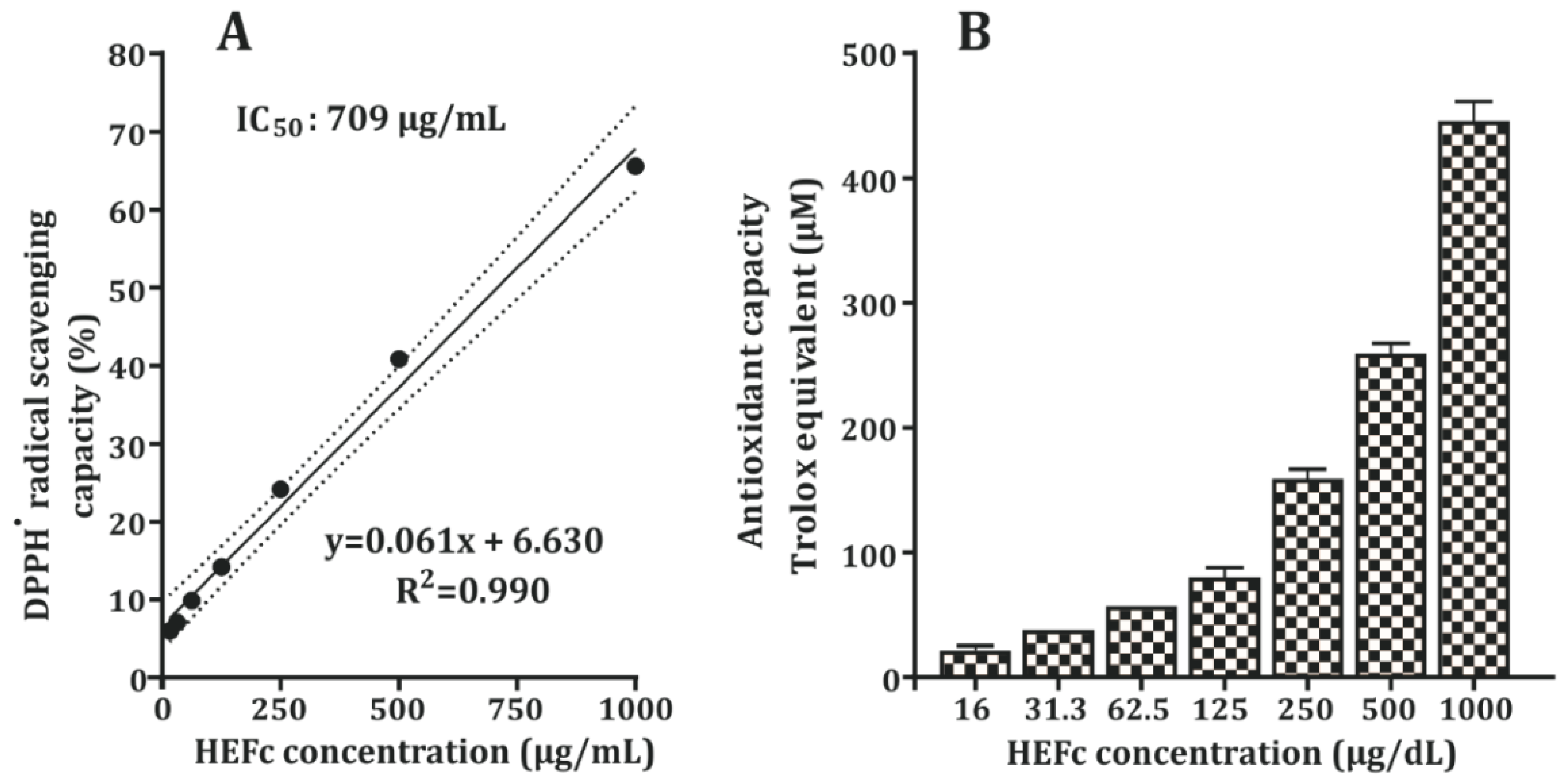
2.1. DPPH Radical Scavenging
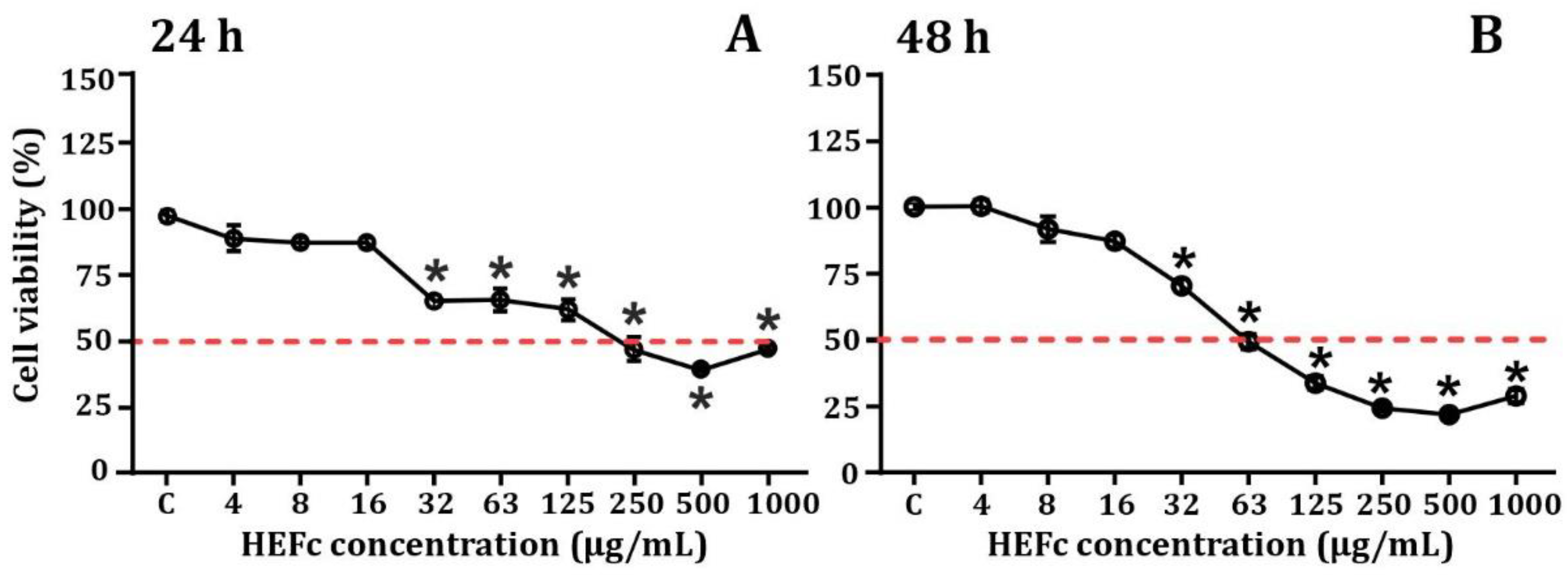
2.2. Cytotoxicity of HEFc and Mycotoxins on SH-SY5Y Cells
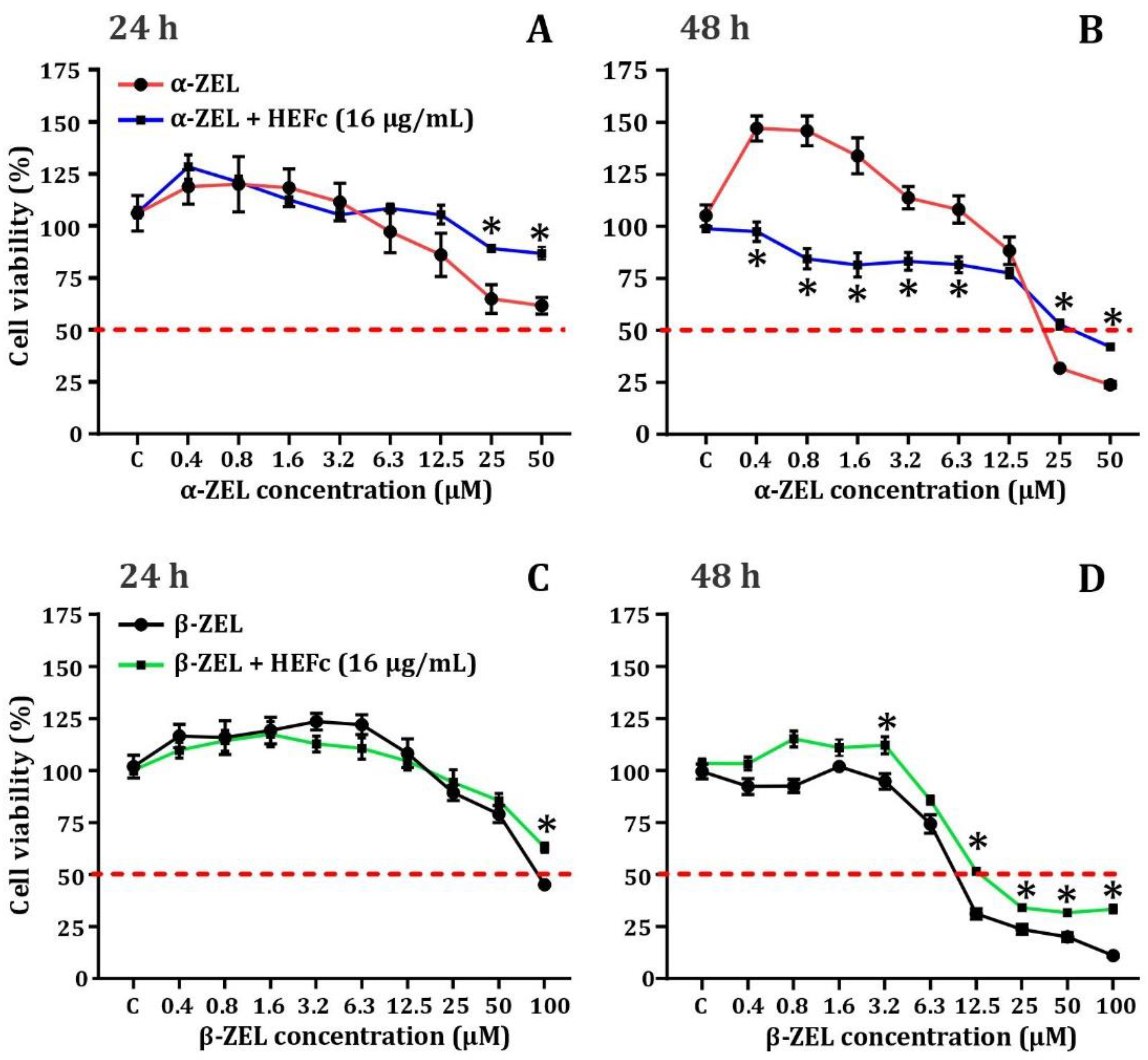
2.3. Cytoprotective Effects of HEFc against Zearalenone Metabolites
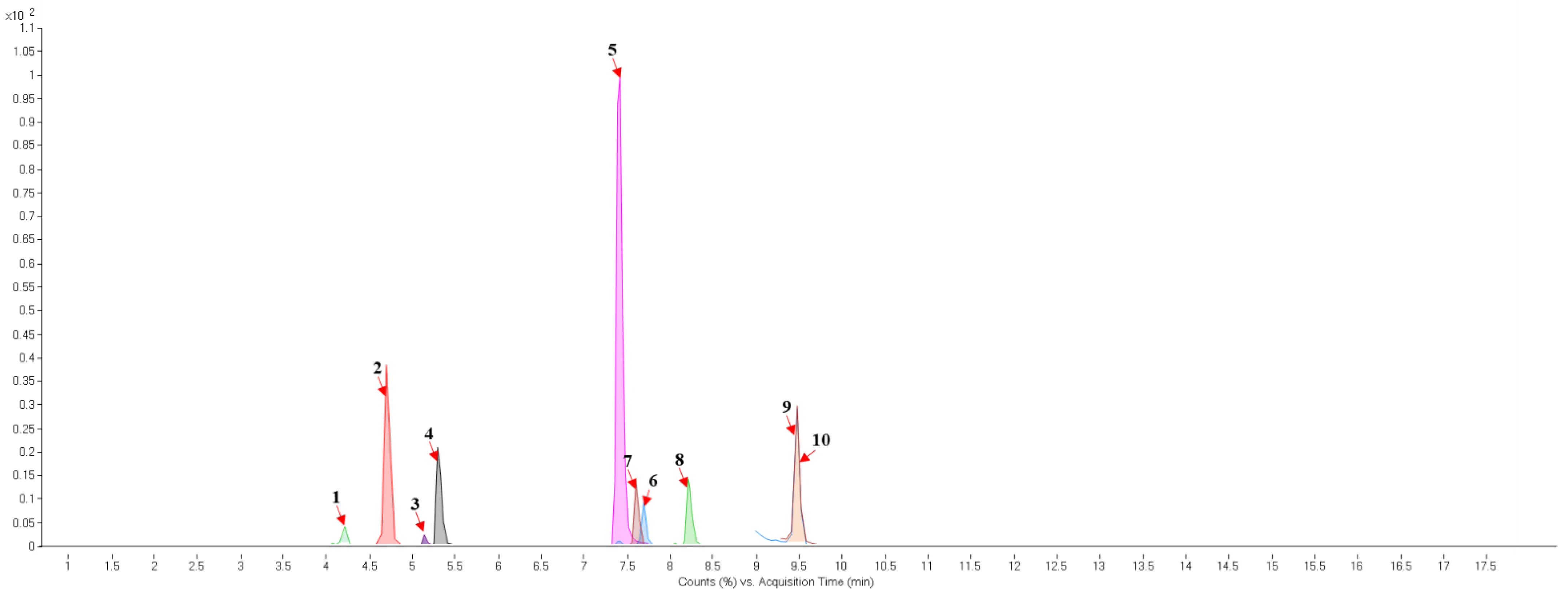
2.4. UPLC-QTOF-MS/MS Analysis
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Material and Extraction
5.2. DPPH Radical-Scavenging Method
5.3. Cellular Exposure to Mycotoxins and HEFc
5.4. Cytoprotective Effects of HEFc against ZEN Metabolites
5.5. MTT Assay
5.6. Analysis of the HEFc by UPLC-QTOF-MS/MS
5.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CI95 | 95% confidence interval |
| IC50 | inhibitory concentration |
| α-ZEL | α-zearalenol |
| β-ZEL | β-zearalenol |
| HEFc | hydroethanolic extract from F. chica leaves |
| SEM | standard error of mean |
| ZEN | zearalenone |
References
- Humboldt. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. 2020. Available online: www.humboldt.org.co (accessed on 19 October 2021).
- Castillo, B.O.D.; Jiménez, Y.M.; Nieves, M.F.F.; Peláez, J.A.M. Composición química de aceites esenciales de hojas de Fridericia florida DC. y Fridericia chica (Bonpl.). Rev. Fac. Cienc. Bás. 2019, 15, 63–70. [Google Scholar] [CrossRef]
- do Amaral, R.R.; Santos, A.A.; Saravia, A.; Botas, G.; Cruz, R.A.; Fernandes, C.P.; Rocha, L.; Boylan, F. Biological activities of Arrabidaea chica (Bonpl.) B. Verl. leaves. Lat. Am. J. Pharm. 2012, 31, 451–455. [Google Scholar]
- Lima, J.C.; de Oliveira, R.G.; Silva, V.C.; de Sousa Jr, P.T.; Violante, I.M.; Macho, A.; Martins, D.T.d.O. Anti-inflammatory activity of 4′, 6, 7-trihydroxy-5-methoxyflavone from Fridericia chica (Bonpl.) LG Lohmann. Nat. Prod. Res. 2020, 34, 726–730. [Google Scholar] [CrossRef] [PubMed]
- Michel, A.F.; Melo, M.M.; Campos, P.P.; Oliveira, M.S.; Oliveira, F.A.; Cassali, G.D.; Ferraz, V.P.; Cota, B.B.; Andrade, S.P.; Souza-Fagundes, E.M. Evaluation of anti-inflammatory, antiangiogenic and antiproliferative activities of Arrabidaea chica crude extracts. J. Ethnopharmacol. 2015, 165, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Jorge, M.P.; Madjarof, C.; Ruiz, A.L.T.G.; Fernandes, A.T.; Rodrigues, R.A.F.; de Oliveira Sousa, I.M.; Foglio, M.A.; de Carvalho, J.E. Evaluation of wound healing properties of Arrabidaea chica Verlot extract. J. Ethnopharmacol. 2008, 118, 361–366. [Google Scholar] [CrossRef]
- Monteiro, F.D.S.; Costa, J.R.D.S.; Martins, L.J.A.; Rocha, C.Q.; Borges, A.C.R.; Borges, M.O.D.R. Hydroalcoholic extract of leaves of Arrabidaea brachypoda (DC.) Bureau present antispasmodic activity mediated through calcium influx blockage. Rev. Ciênc. Farm. Básica Apl. 2020, 41, 1–13. [Google Scholar] [CrossRef]
- Siraichi, J.T.; Pedrochi, F.; Natali, M.R.; Ueda-Nakamura, T.; Filho, B.P.; Bento, A.C.; Baesso, M.L.; Nakamura, C.V. Ultraviolet (UVB and UVA) photoprotector activity and percutaneous penetration of extracts obtained from Arrabidaea chica. Appl. Spectrosc. 2013, 67, 1179–1184. [Google Scholar] [CrossRef]
- Cortez de Sá, J.; Almeida-Souza, F.; Mondêgo-Oliveira, R.; Oliveira Idos, S.; Lamarck, L.; Magalhães Ide, F.; Ataídes-Lima, A.F.; Ferreira Hda, S.; Abreu-Silva, A.L. Leishmanicidal, cytotoxicity and wound healing potential of Arrabidaea chica Verlot. BMC Complement. Altern. Med. 2015, 16, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Siraichi, J.T.; Felipe, D.F.; Brambilla, L.Z.; Gatto, M.J.; Terra, V.A.; Cecchini, A.L.; Cortez, L.E.; Rodrigues-Filho, E.; Cortez, D.A. Antioxidant capacity of the leaf extract obtained from Arrabidaea chica cultivated in Southern Brazil. PLoS ONE 2013, 8, e72733. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.I.; Cheng, C.I.; Kang, Y.F.; Chang, P.C.; Lin, I.P.; Kuo, Y.H.; Jhou, A.J.; Lin, M.Y.; Chen, C.Y.; Lee, C.H. Hispidulin inhibits neuroinflammation in lipopolysaccharide-activated BV2 microglia and attenuates the activation of Akt, NF-κB, and STAT3 pathway. Neurotox Res. 2020, 38, 163–174. [Google Scholar] [CrossRef]
- Juan-García, A.; Juan, C.; Bind, M.A.; Engert, F. Study of locomotion response and development in zebrafish (Danio rerio) embryos and larvae exposed to enniatin A, enniatin B, and beauvericin. Sci. Total Environ. 2021, 777, 146075. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.W.; Wang, J.Q.; Zheng, B.Q.; Li, S.L.; Zhang, Y.D.; Li, F.D.; Zheng, N. Cytotoxicity induced by ochratoxin A, zearalenone, and α-zearalenol: Effects of individual and combined treatment. Food Chem. Toxicol. 2014, 71, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Agahi, F.; Juan, C.; Font, G.; Ana Juan-García, A. In silico methods for metabolomic and toxicity prediction of zearalenone, α-zearalenone and β-zearalenone. Food Chem. Toxicol. 2020, 146, 111818. [Google Scholar] [CrossRef]
- Tatay, E.; Espin, S.; Garcia-Fernandez, A.-J.; Ruiz, M.-J. Oxidative damage and disturbance of antioxidant capacity by zearalenone and its metabolites in human cells. Toxicol. Vitr. 2017, 45, 334–339. [Google Scholar] [CrossRef] [PubMed]
- Tiemann, U.; Viergutz, T.; Jonas, L.; Schneider, F. Influence of the mycotoxins α-and β-zearalenol and deoxynivalenol on the cell cycle of cultured porcine endometrial cells. Reprod. Toxicol. 2003, 17, 209–218. [Google Scholar] [CrossRef]
- Agahi, F.; Juan, C.; Font, G.; Juan-García, A. Neurotoxicity of zearalenone’s metabolites and beauvericin mycotoxins via apoptosis and cell cycle disruption. Toxicology 2021, 456, 152784. [Google Scholar] [CrossRef]
- Shier, W.T.; Shier, A.; Xie, W.; Mirocha, C. Structure-activity relationships for human estrogenic activity in zearalenone mycotoxins. Toxicon 2001, 39, 1435–1438. [Google Scholar] [CrossRef]
- Pizzo, F.; Caloni, F.; Schutz, L.F.; Totty, M.L.; Spicer, L.J. Individual and combined effects of deoxynivalenol and α-zearalenol on cell proliferation and steroidogenesis of granulosa cells in cattle. Environ. Toxicol. Pharmacol. 2015, 40, 722–728. [Google Scholar] [CrossRef]
- Violante, I.M.; Carollo, C.A.; Silva, L.I.; Oliveira, A.Q.; Pardinho, F.C.; Garcez, W.S.; Garcez, F.R.; Oliveira, R.G.d.; Arunachalam, K.; de Oliveira Martins, D.T. Cytotoxicity and antibacterial activity of scutellarein and carajurone-enriched fraction obtained from the hydroethanolic extract of the leaves of Fridericia chica (Bonpl.) LG Lohmann. Nat. Prod. Res. 2020, 1–7. [Google Scholar] [CrossRef]
- Mafioleti, L.; da Silva Junior, I.F.; Colodel, E.M.; Flach, A.; de Oliveira Martins, D.T. Evaluation of the toxicity and antimicrobial activity of hydroethanolic extract of Arrabidaea chica (Humb. & Bonpl.) B. Verl. J. Ethnopharmacol. 2013, 150, 576–582. [Google Scholar] [PubMed]
- Martins, F.J.; Caneschi, C.A.; Vieira, J.L.; Barbosa, W.; Raposo, N.R. Antioxidant activity and potential photoprotective from amazon native flora extracts. J. Photochem. Photobiol. B 2016, 161, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Torres, C.A.; Pérez Zamora, C.M.; Nuñez, M.B.; Gonzalez, A.M. In vitro antioxidant, antilipoxygenase and antimicrobial activities of extracts from seven climbing plants belonging to the Bignoniaceae. J. Integr. Med. 2018, 16, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Paula, J.T.; Paviani, L.C.; Foglio, M.A.; Sousa, I.M.O.; Duarte, G.H.B.; Jorge, M.P.; Eberlin, M.N.; Cabral, F.A. Extraction of anthocyanins and luteolin from Arrabidaea chica by sequential extraction in fixed bed using supercritical CO2, ethanol and water as solvents. J. Supercrit. Fluids 2014, 86, 100–107. [Google Scholar] [CrossRef]
- Olivero-Verbel, J.; De la Parra-Guerra, A.; Caballero-Gallardo, K.; Sierra-Marquez, L.; Fuentes-Lopez, K.; Franco-Marmolejo, J.; Jannasch, A.S.; Sepulveda, M.S.; Stashenko, E. The aqueous extract of Fridericia chica grown in northern Colombia ameliorates toxicity induced by tergitol on Caenorhabditis elegans. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2021, 244, 109026. [Google Scholar] [CrossRef] [PubMed]
- Suffness, M.; Pezzuto, J.M. Methods in Plant Biochemistry: Assays for Bioactivity; Academic Press: London, UK, 1990; pp. 71–133. [Google Scholar]
- Salles, T.H.C.; Volpe-Zanutto, F.; de Oliveira Sousa, I.M.; Machado, D.; Zanatta, A.C.; Vilegas, W.; Lancellotti, M.; Foglio, M.A.; d’Ávila, M.A. Electrospun PCL-based nanofibers Arrabidaea chica Verlot-Pterodon pubescens Benth loaded: Synergic effect in fibroblast formation. Biomed. Mater. 2020, 15, 65001. [Google Scholar] [CrossRef]
- Othmen, Z.O.-B.; El Golli, E.; Abid-Essefi, S.; Bacha, H. Cytotoxicity effects induced by zearalenone metabolites, α-zearalenol and β-zearalenol, on cultured Vero cells. Toxicology 2008, 252, 72–77. [Google Scholar] [CrossRef]
- Pillay, D.; Chuturgoon, A.A.; Nevines, E.; Manickum, T.; Deppe, W.; Dutton, M.F. The quantitative analysis of zearalenone and its derivatives in plasma of patients with breast and cervical cancer. Clin. Chem. Lab. Med. 2002, 40, 946–951. [Google Scholar] [CrossRef]
- Zheng, W.; Wang, B.; Li, X.; Wang, T.; Zou, H.; Gu, J.; Yuan, Y.; Liu, X.; Bai, J.; Bian, J.; et al. Zearalenone promotes cell proliferation or causes cell death? Toxins 2018, 10, 184. [Google Scholar] [CrossRef] [Green Version]
- Gonkowski, S.; Obremski, K.; Calka, J. The influence of low doses of zearalenone on distribution of selected active substances in nerve fibers within the circular muscle layer of porcine ileum. J. Mol. Neurosci. 2015, 56, 878–886. [Google Scholar] [CrossRef] [Green Version]
- Venkataramana, M.; Chandra Nayaka, S.; Anand, T.; Rajesh, R.; Aiyaz, M.; Divakara, S.T.; Murali, H.S.; Prakash, H.S.; Lakshmana Rao, P.V. Zearalenone induced toxicity in SHSY-5Y cells: The role of oxidative stress evidenced by N-acetyl cysteine. Food Chem. Toxicol. 2014, 65, 335–342. [Google Scholar] [CrossRef]
- Liu, K.; Zhao, F.; Yan, J.; Xia, Z.; Jiang, D.; Ma, P. Hispidulin: A promising flavonoid with diverse anti-cancer properties. Life Sci. 2020, 259, 118395. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhao, Q.; Liu, T.; Zhao, H.; Wang, R.; Li, H.; Sun, H. Effect of Vicenin-2 on ovariectomy-induced osteoporosis in rats. Biomed Pharm. 2020, 129, 110474. [Google Scholar] [CrossRef]
- Chen, X.; Yao, Z.; Peng, X.; Wu, L.; Wu, H.; Ou, Y.; Lai, J. Eupafolin alleviates cerebral ischemia/reperfusion injury in rats via blocking the TLR4/NF-κB signaling pathway. Mol. Med. Rep. 2020, 22, 5135–5144. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Liang, J. Pectolinarigenin promotes functional recovery and inhibits apoptosis in rats following spinal cord injuries. Exp. Ther. Med. 2019, 17, 3877–3882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.; Huang, K.; Ning, H. Autophagy induction by hispidulin provides protection against sevoflurane-induced neuronal apoptosis in aged rats. Biomed Pharm. 2018, 98, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Pang, Q.; Zhao, Y.; Chen, X.; Zhao, K.; Zhai, Q.; Tu, F. Apigenin protects the brain against ischemia/reperfusion injury via caveolin-1/VEGF in vitro and in vivo. Oxid. Med. Cell Longev. 2018, 2018, 7017204. [Google Scholar] [CrossRef]
- Yao, H.; Yuan, Z.; Wei, G.; Chen, C.; Duan, J.; Li, Y.; Liu, Y. Thevetiaflavone from Wikstroemia indica ameliorates PC12 cells injury induced by OGD/R via improving ROS-mediated mitochondrial dysfunction. Mol. Med. Rep. 2017, 16, 9197–9202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bu, J.; Shi, S.; Wang, H.Q.; Niu, X.S.; Zhao, Z.F.; Wu, W.D.; Zhu, Y. Acacetin protects against cerebral ischemia-reperfusion injury via the NLRP3 signaling pathway. Neural Regen. Res. 2019, 14, 605. [Google Scholar]
- Montesano, D.; Juan-García, A.; Mañes, J.; Juan, C. Chemoprotective effect of carotenoids from Lycium barbarum L. on SH-SY5Y neuroblastoma cells treated with beauvericin. Food Chem. Toxicol. 2020, 141, 111414. [Google Scholar] [CrossRef]
- Agahi, F.; Font, G.; Juan, C.; Juan-García, A. Individual and combined effect of zearalenone derivates and beauvericin mycotoxins on SH-SY5Y Cells. Toxins 2020, 12, 212. [Google Scholar] [CrossRef] [Green Version]
- Caballero-Gallardo, K.; Olivero-Verbel, J.; Corada-Fernández, C.; Lara-Martín, P.A.; Juan-García, A. Emerging contaminants and priority substances in marine sediments from Cartagena Bay and the Grand Marsh of Santa Marta (Ramsar site), Colombia. Environ. Monit. Assess 2021, 193, 1–18. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | IC50 (CI95) 24 h | IC50 (CI95) 48 h |
|---|---|---|
| HEFc | 61.2 µg/mL (45–83) | 53.8 µg/mL (24.0–116.6) |
| α-ZEL | >50 µM | 17.9 µM (10.4–32.4) |
| β-ZEL | >100 µM | 10.5 µM (7.1–15.7) |
| No. Figure 4 | RT (min) | Tentative Annotation | Structure | Formula | Ion | Experimental Mass | Calculated Mass | Δ ppm |
|---|---|---|---|---|---|---|---|---|
| 1 | 4.204 | Vicenin-2 |  | C27H30O15 | [M+H]+ | 594.1571 | 594.15847 | −2.30 |
| 2 | 4.732 | 6-hydroxyluteolin 7-rhamnoside |  | C21H20O11 | [M+H]+ | 448.10071 | 448.10056 | 0.33 |
| 3 | 5.158 | Scutellarein-O-glucuronide |  | C21H18O12 | [M+H]+ | 462.07918 | 462.07983 | −1.40 |
| 4 | 5.355 | Nepetin |  | C16H12O7 | [M+H]+ | 316.05831 | 316.0583 | 0.03 |
| 5 | 7.443 | Pectolinarigenin |  | C17H14O6 | [M+H]+ | 314.07919 | 314.07904 | 0.47 |
| 6 | 7.578 | Hispidulin |  | C16H12O6 | [M+H]+ | 300.06317 | 300.06339 | −0.73 |
| 7 | 7.631 | Apigenin |  | C15H10O5 | [M+H]+ | 270.05259 | 270.05282 | −0.85 |
| 8 | 8.254 | 5-O-methylscutellarein |  | C16H12O6 | [M+H]+ | 300.06367 | 300.06339 | 0.93 |
| 9 | 9.331 | Thevetiaflavone |  | C16H12O5 | [M+H]+ | 284.0685 | 284.06847 | 0.10 |
| 10 | 9.572 | Acacetin |  | C16H12O5 | [M+H]+ | 284.06872 | 284.06847 | 0.88 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alvarez-Ortega, N.; Caballero-Gallardo, K.; Taboada-Alquerque, M.; Franco, J.; Stashenko, E.E.; Juan, C.; Juan-García, A.; Olivero-Verbel, J. Protective Effects of the Hydroethanolic Extract of Fridericia chica on Undifferentiated Human Neuroblastoma Cells Exposed to α-Zearalenol (α-ZEL) and β-Zearalenol (β-ZEL). Toxins 2021, 13, 748. https://doi.org/10.3390/toxins13110748
Alvarez-Ortega N, Caballero-Gallardo K, Taboada-Alquerque M, Franco J, Stashenko EE, Juan C, Juan-García A, Olivero-Verbel J. Protective Effects of the Hydroethanolic Extract of Fridericia chica on Undifferentiated Human Neuroblastoma Cells Exposed to α-Zearalenol (α-ZEL) and β-Zearalenol (β-ZEL). Toxins. 2021; 13(11):748. https://doi.org/10.3390/toxins13110748
Chicago/Turabian StyleAlvarez-Ortega, Neda, Karina Caballero-Gallardo, María Taboada-Alquerque, Jackeline Franco, Elena E. Stashenko, Cristina Juan, Ana Juan-García, and Jesus Olivero-Verbel. 2021. "Protective Effects of the Hydroethanolic Extract of Fridericia chica on Undifferentiated Human Neuroblastoma Cells Exposed to α-Zearalenol (α-ZEL) and β-Zearalenol (β-ZEL)" Toxins 13, no. 11: 748. https://doi.org/10.3390/toxins13110748
APA StyleAlvarez-Ortega, N., Caballero-Gallardo, K., Taboada-Alquerque, M., Franco, J., Stashenko, E. E., Juan, C., Juan-García, A., & Olivero-Verbel, J. (2021). Protective Effects of the Hydroethanolic Extract of Fridericia chica on Undifferentiated Human Neuroblastoma Cells Exposed to α-Zearalenol (α-ZEL) and β-Zearalenol (β-ZEL). Toxins, 13(11), 748. https://doi.org/10.3390/toxins13110748