Oral Toxicity of Pseudomonas protegens against Muscoid Flies



Abstract
:1. Introduction
2. Results
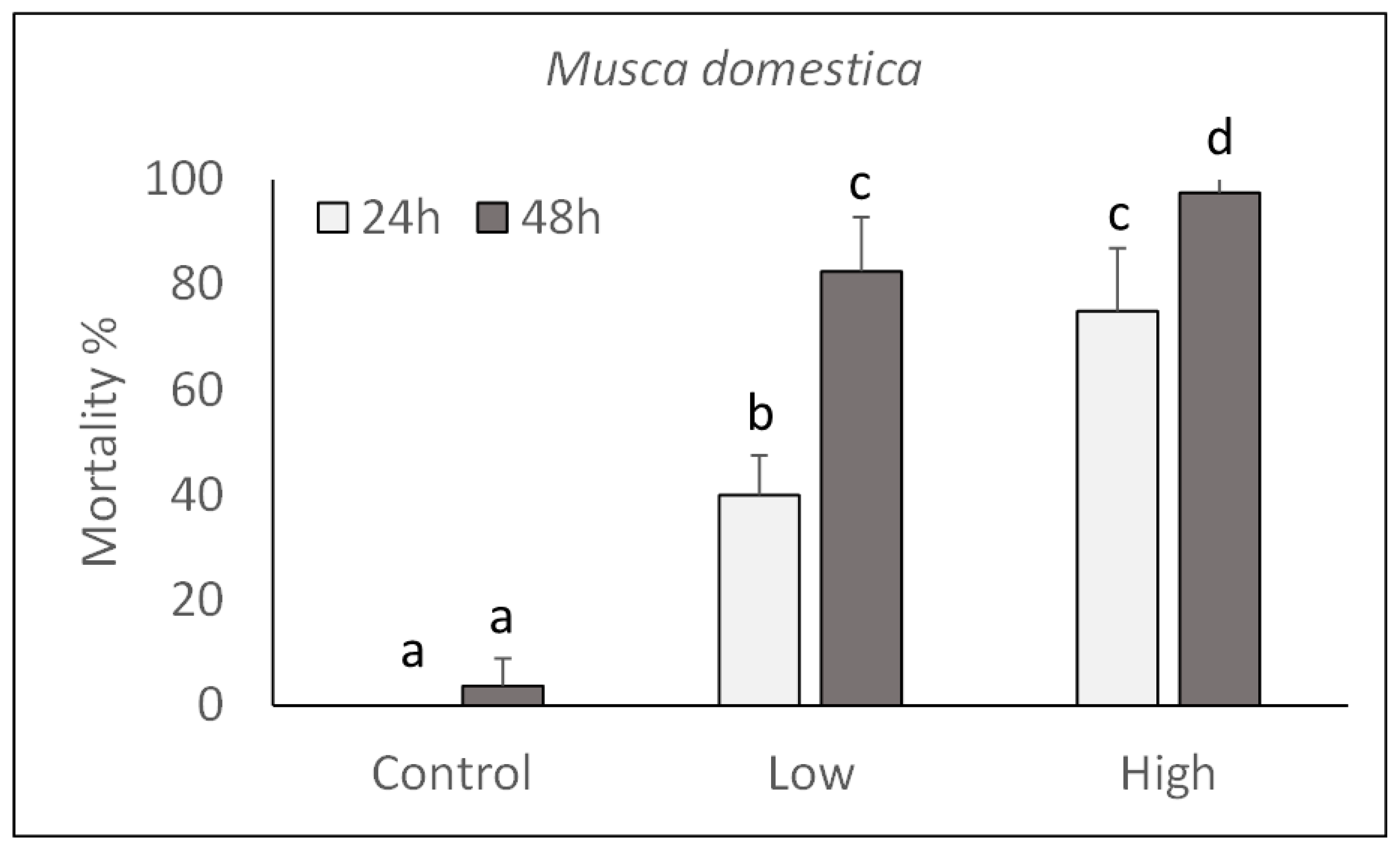
2.1. Oral Toxicity
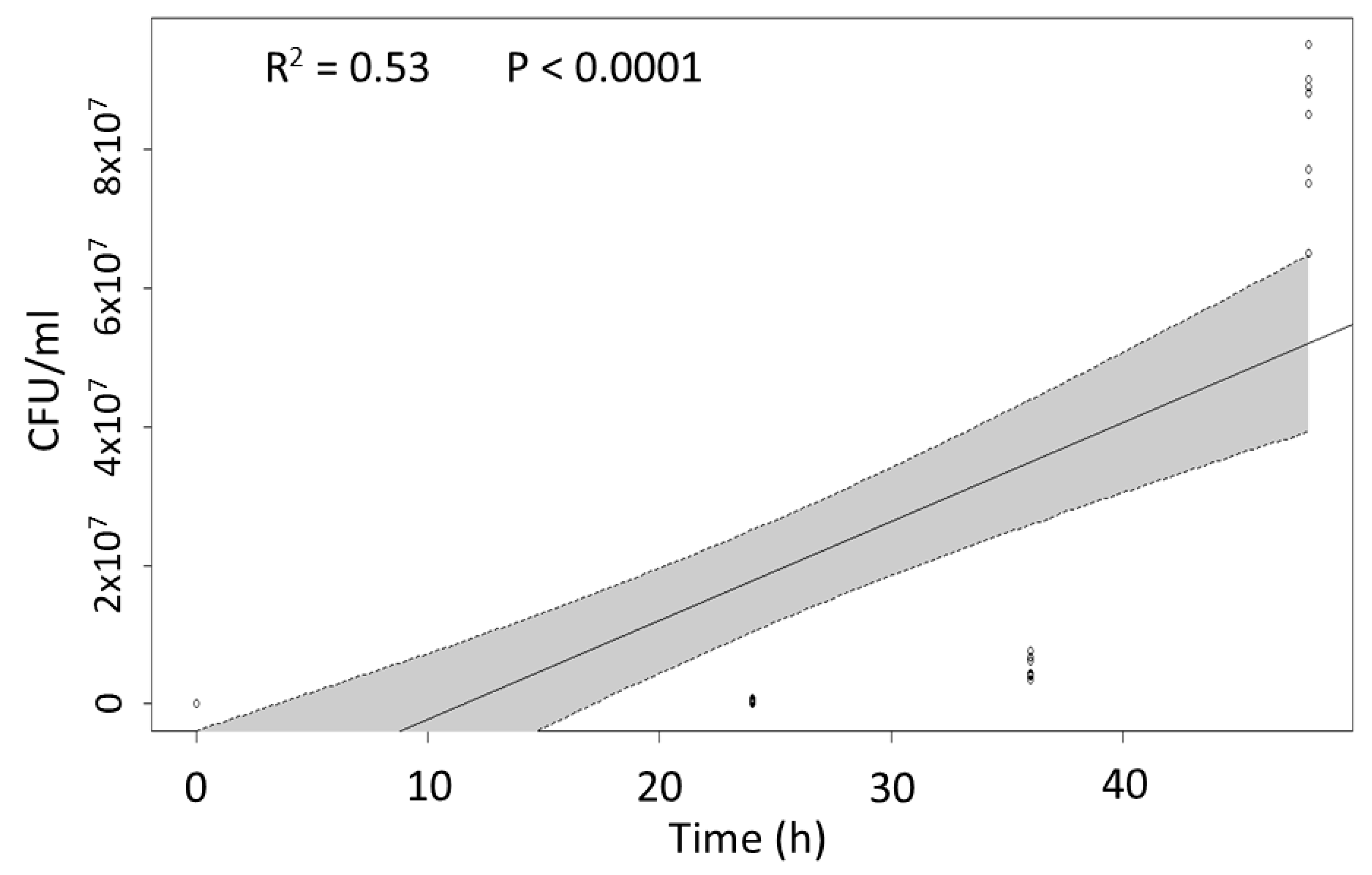
2.2. Post-Injection Pathogenicity
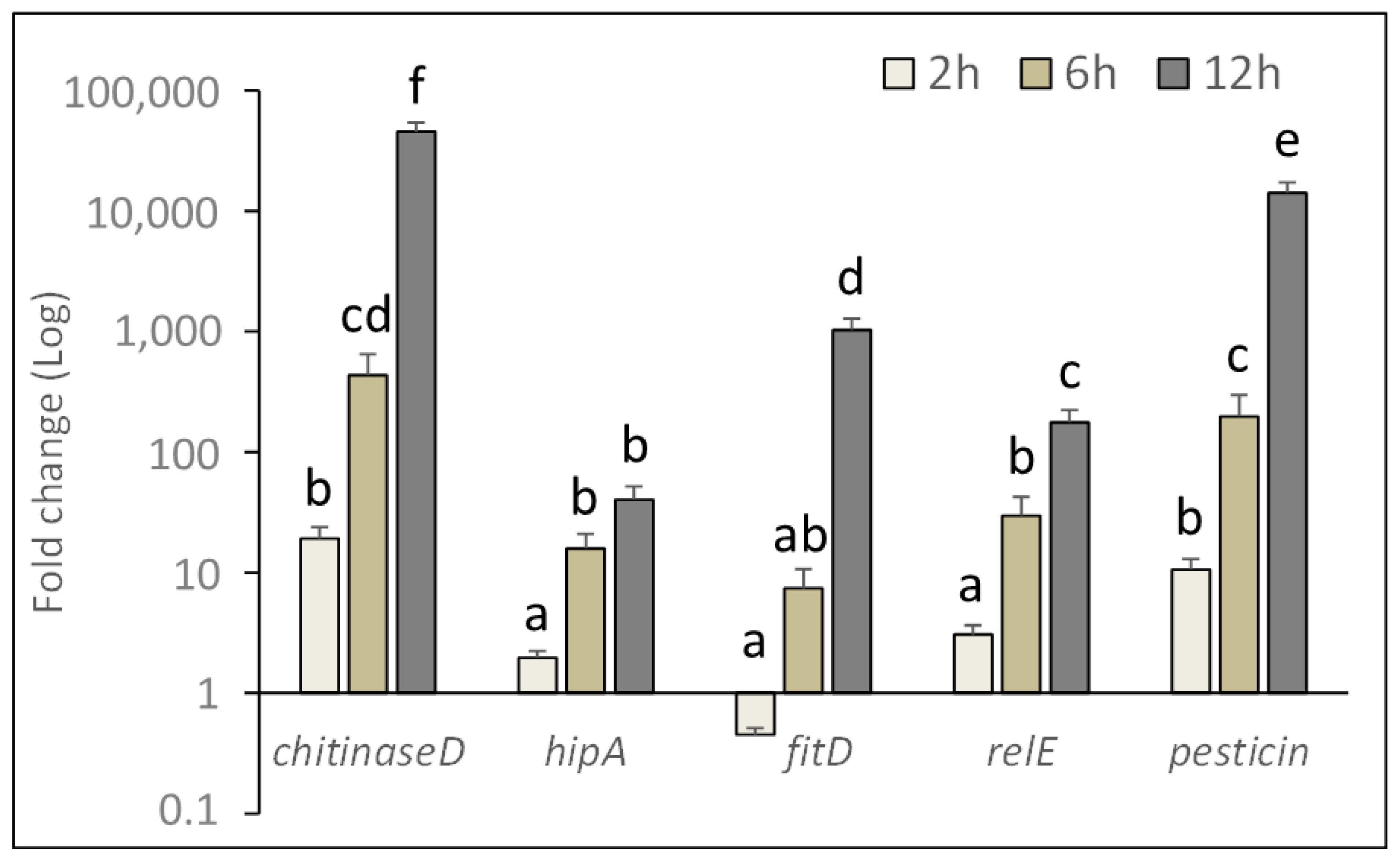
2.3. In Vivo Expression of Bacterial Genes
3. Discussion
4. Materials and Methods
4.1. Bacterial Strain and Preparations
4.2. Bioassays
4.2.1. Ingestion Bioassays
4.2.2. Injection Bioassays
4.3. Insecticidal Protein Gene Expression at Gut Level
4.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ruiu, L. Plant-growth-promoting Bacteria (PGPB) against insects and other agricultural pests. Agronomy 2020, 10, 861. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; Paulitz, T.C.; Steinberg, C.; Alabouvette, C.; Moënne-Loccoz, Y. The rhizosphere: A playground and battlefield for soilborne pathogens and beneficial microorganisms. Plant Soil 2009, 321, 341–361. [Google Scholar] [CrossRef] [Green Version]
- Philmus, B.; Shaffer, B.T.; Kidarsa, T.A.; Yan, Q.; Raaijmakers, J.M.; Begley, T.P.; Loper, J.E. Investigations into the biosynthesis, regulation, and self-resistance of toxoflavin in Pseudomonas protegens Pf-5. ChemBioChem 2015, 16, 1782–1790. [Google Scholar] [CrossRef]
- Vodovar, N.; Vallenet, D.; Cruveiller, S.; Rouy, Z.; Barbe, V.; Acosta, C.; Cattolico, L.; Jubin, C.; Lajus, A.; Segurens, B.; et al. Complete genome sequence of the entomopathogenic and metabolically versatile soil bacterium Pseudomonas entomophila. Nat. Biotechnol. 2006, 24, 673–679. [Google Scholar] [CrossRef] [Green Version]
- Ruiu, L. Microbial biopesticides in agroecosystems. Agronomy 2018, 8, 235. [Google Scholar] [CrossRef] [Green Version]
- Flury, P.; Aellen, N.; Ruffner, B.; Péchy-Tarr, M.; Fataar, S.; Metla, Z.; Dominguez-Ferreras, A.; Bloemberg, G.; Frey, J.; Goesmann, A.; et al. Insect pathogenicity in plant-beneficial pseudomonads: Phylogenetic distribution and comparative genomics. ISME J. 2016, 10, 2527–2542. [Google Scholar] [CrossRef] [PubMed]
- Péchy-Tarr, M.; Bruck, D.J.; Maurhofer, M.; Fischer, E.; Vogne, C.; Henkels, M.D.; Donahue, K.M.; Grunder, J.; Loper, J.E.; Keel, C. Molecular analysis of a novel gene cluster encoding an insect toxin in plant-associated strains of Pseudomonas fluorescens. Environ. Microbiol. 2008, 10, 2368–2386. [Google Scholar] [CrossRef] [PubMed]
- Loper, J.E.; Henkels, M.D.; Rangel, L.I.; Olcott, M.H.; Walker, F.L.; Bond, K.L.; Kidarsa, T.A.; Hesse, C.N.; Sneh, B.; Stockwell, V.O.; et al. Rhizoxin analogs, orfamide A and chitinase production contribute to the toxicity of Pseudomonas protegens strain Pf-5 to Drosophila melanogaster. Environ. Microbiol. 2016, 18, 3509–3521. [Google Scholar] [CrossRef] [PubMed]
- Ruiu, L.; Virdis, B.; Mura, M.E.; Floris, I.; Satta, A.; Tarasco, E. Oral insecticidal activity of new bacterial isolates against insects in two orders. Biocontrol Sci. Technol. 2017, 27, 886–902. [Google Scholar] [CrossRef]
- Ruffner, B.; Péchy-Tarr, M.; Ryffel, F.; Hoegger, P.; Obrist, C.; Rindlisbacher, A.; Keel, C.; Maurhofer, M. Oral insecticidal activity of plant-associated pseudomonads. Environ. Microbiol. 2013, 15, 751–763. [Google Scholar] [CrossRef]
- Glare, T.R.; Jurat-Fuentes, J.L.; O’callaghan, M. Basic and applied research: Entomopathogenic bacteria. In Microbial Control of Insect and Mite Pests, 1st ed.; Lacey, L., Ed.; Academic Press: New York, NY, USA, 2017; pp. 47–67. [Google Scholar]
- Bedini, S.; Muniz, E.R.; Tani, C.; Conti, B.; Ruiu, L. Insecticidal potential of Brevibacillus laterosporus against dipteran pest species in a wide ecological range. J. Invertebr. Pathol. 2020, 177, 107493. [Google Scholar] [CrossRef] [PubMed]
- Adang, M.J.; Crickmore, N.; Jurat-Fuentes, J.L. Diversity of Bacillus thuringiensis crystal toxins and mechanism of action. Adv. Insect Physiol. 2014, 47, 39–87. [Google Scholar]
- Waterfield, N.R.; Bowen, D.J.; Fetherston, J.D.; Perry, R.D.; Ffrench-Constant, R.H. The tc genes of Photorhabdus: A growing family. Trends Microbiol. 2001, 9, 185–191. [Google Scholar] [CrossRef]
- Rangel, L.I.; Henkels, M.D.; Shaffer, B.T.; Walker, F.L.; Ii, E.W.D.; Stockwell, V.O.; Bruck, D.; Taylor, B.J.; Loper, J.E. Characterization of toxin complex gene clusters and insect toxicity of bacteria representing four subgroups of Pseudomonas fluorescens. PLoS ONE 2016, 11, e0161120. [Google Scholar] [CrossRef] [Green Version]
- Keel, C. A look into the toolbox of multi-talents: Insect pathogenicity determinants of plant-beneficial pseudomonads. Environ Microbiol. 2016, 18, 3207–3209. [Google Scholar] [CrossRef] [Green Version]
- Landsberg, M.J.; Jones, S.A.; Rothnagel, R.; Busby, J.N.; Marshall, S.D.G.; Simpson, R.M.; Lott, J.S.; Hankamer, M.; Hurst, M.R.H. 3D structure of the Yersinia entomophaga toxin complex and implications for insecticidal activity. Proc. Natl. Acad. Sci. USA 2011, 108, 20544–20549. [Google Scholar] [CrossRef] [Green Version]
- Mura, M.E.; Ruiu, L. Brevibacillus laterosporus pathogenesis and local immune response regulation in the house fly midgut. J. Invertebr. Pathol. 2017, 145, 55–61. [Google Scholar] [CrossRef]
- Vesga, P.; Flury, P.; Vacheron, J.; Keel, C.; Croll, D.; Maurhofer, M. Transcriptome plasticity underlying plant root colonization and insect invasion by Pseudomonas protegens. ISME J. 2020, 14, 2766–2782. [Google Scholar] [CrossRef]
- Ruiu, L.; Satta, A.; Floris, I. Comparative applications of azadirachtin-and Brevibacillus laterosporus-based formulations for house fly management experiments in dairy farms. J. Medical Entomol. 2011, 48, 345–350. [Google Scholar] [CrossRef] [Green Version]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RTPCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using realtime quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Toxin Gene | Abbreviation | Acc. Number | Primer Sequence | |
|---|---|---|---|---|
| Sense 5′-3′ | Antisense 5′-3′ | |||
| chitinase D | ChiD | NC_021237.1 | 5′ CATATCGAATTGCACAAGGGCAACGAACAG 3′ | 5′ AGGCGCCATGCTGATGATGAAGTGCTT 3′ |
| toxin HipA | HipA | NC_021237.1 | 5′ CTGCGACATGCTCAGAAGCGAGTTCCACTA 3′ | 5′ ACGCAGGTAGTCGGCCACCAGCTC 3′ |
| toxin RelE | RelE | NC_021237.1 | 5′ ATGGCGAAGCCGGAGAGGAACCCA 3′ | 5′ AAGGTCACAAGACCGGCTCGGGCC 3′ |
| pesticin domain protein | Pesticin | NC_021237.1 | 5′ ATGTCACGCTACGCGATTGATTTCAGTTTTATC3′ | 5′ TGATGTTCAAGGGCTGGCCGTCGAGAA 3′ |
| cytotoxin FitD | FitD | EU400157.2 | 5′ CGCCAACACCGAGCCACAGCCGGAGG 3′ | 5′ CGCGTTCAGGCCGTCCACATGCGCCAC 3′ |
| 16s rDNA | 16S rDNA | NR_114749.1 | 5′ TGGGAGGAAGGGCAGTTACCTAATACGTGA 3′ | 5′ TTCCACCACCCTCTACCATACTCTAGC 3′ |
| β-Actin | β-Actin | NW_004765946.1 | 5′ ATGAGGCTCAGAGCAAACGTGGTA 3′ | 5′ AGTCATCTTCTCGCGATTGGCCTT 3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruiu, L.; Mura, M.E. Oral Toxicity of Pseudomonas protegens against Muscoid Flies. Toxins 2021, 13, 772. https://doi.org/10.3390/toxins13110772
Ruiu L, Mura ME. Oral Toxicity of Pseudomonas protegens against Muscoid Flies. Toxins. 2021; 13(11):772. https://doi.org/10.3390/toxins13110772
Chicago/Turabian StyleRuiu, Luca, and Maria Elena Mura. 2021. "Oral Toxicity of Pseudomonas protegens against Muscoid Flies" Toxins 13, no. 11: 772. https://doi.org/10.3390/toxins13110772
APA StyleRuiu, L., & Mura, M. E. (2021). Oral Toxicity of Pseudomonas protegens against Muscoid Flies. Toxins, 13(11), 772. https://doi.org/10.3390/toxins13110772