Staphylococcus aureus Isolated from Ruminants with Mastitis in Northern Greece Dairy Herds: Genetic Relatedness and Phenotypic and Genotypic Characterization

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Results
2.1. Distribution of Virulence and Resistance Genes
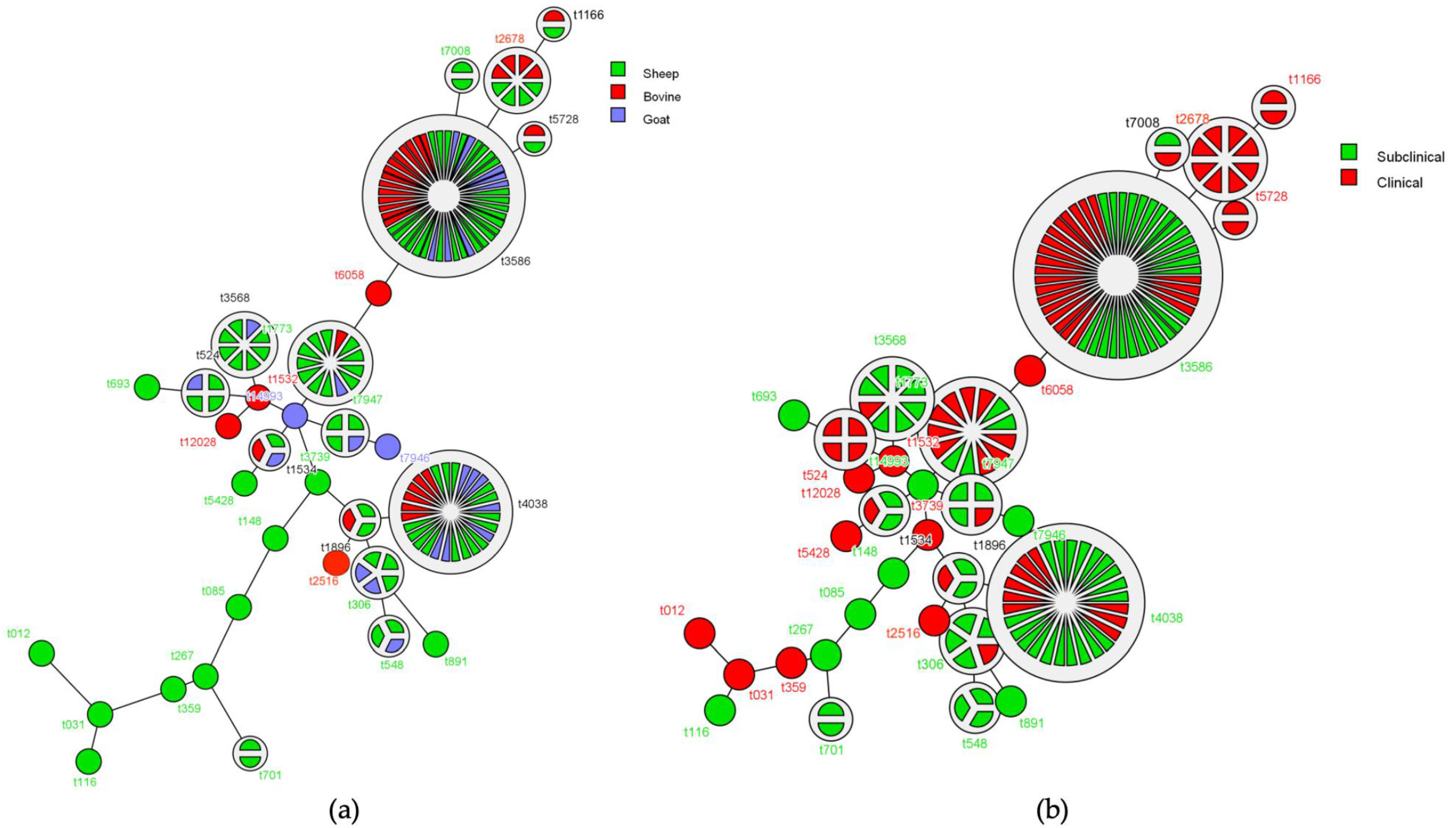
2.2. PFGE Genotyping
2.3. Identification and Distribution of spa Types
2.4. Phenotypic Antimicrobial Resistance
2.5. Biofilm Formation and Cell Surface Traits
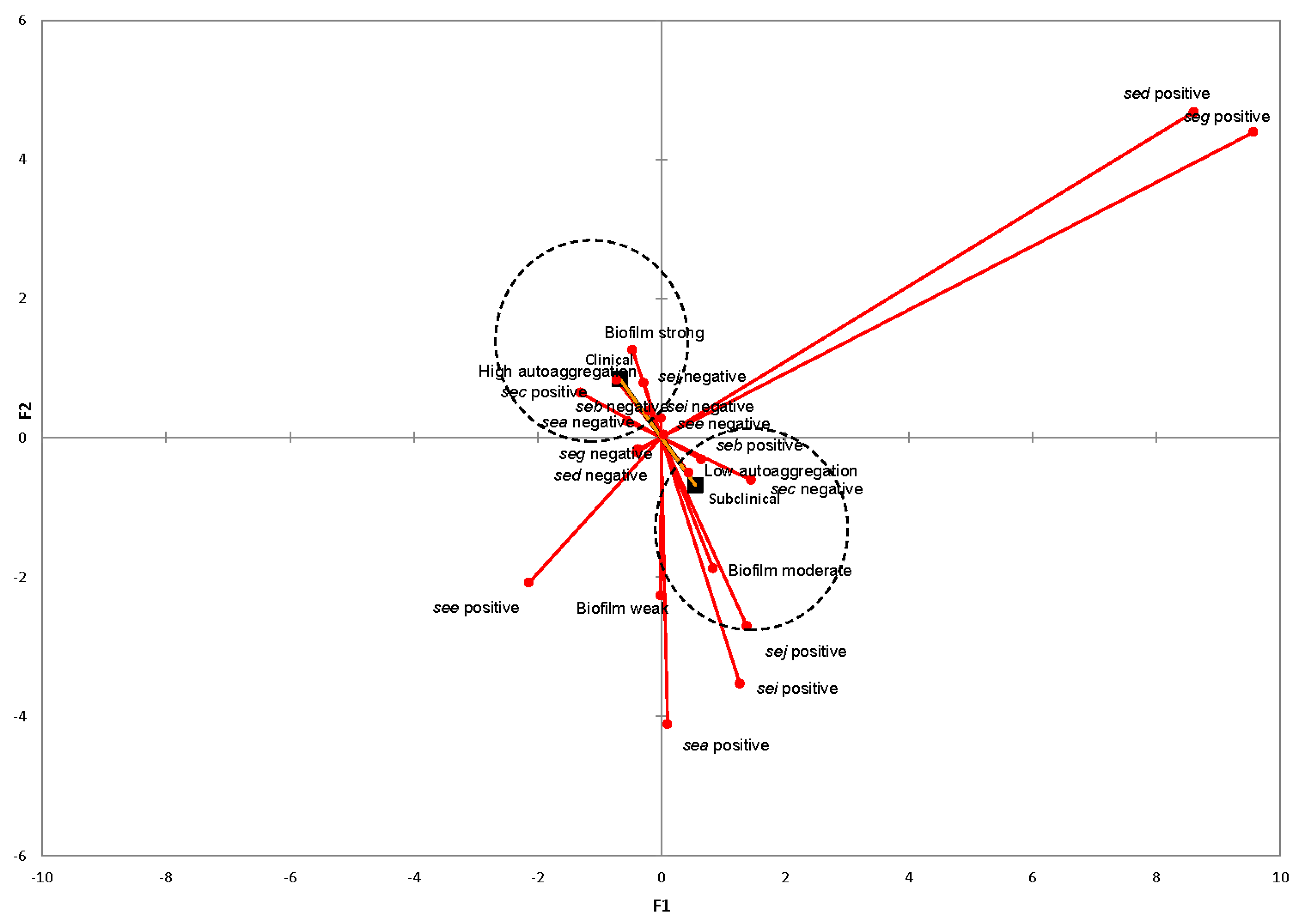
2.6. Associations between SE Genes, Genotypic Traits, and Mastitis
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. DNA Extraction and Species Confirmation
4.3. Detection of Virulence and Resistance Genes
4.4. PFGE
4.5. Spa Typing
4.6. Biofilm Formation and Cell Surface Traits
4.6.1. Biofilm Formation
4.6.2. Autoaggregation Assay
4.6.3. Cell Surface Hydrophobicity
4.7. Antimicrobial Susceptibility Testing
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hogeveen, H.; Huijps, K.; Lam, T.J.G.M. Economic aspects of mastitis: New developments. N. Z. Vet. J. 2011, 59, 16–23. [Google Scholar] [CrossRef]
- Gougoulis, D.A.; Kyriazakis, I.; Fthenakis, G.C. Diagnostic significance of behaviour changes of sheep: A selected review. Small Rumin. Res. 2010, 92, 52–56. [Google Scholar] [CrossRef]
- Wilson, D.J.; Gonzalez, R.N.; Das, H.H. Bovine Mastitis Pathogens in New York and Pennsylvania: Prevalence and Effects on Somatic Cell Count and Milk Production. J. Dairy Sci. 1997, 80, 2592–2598. [Google Scholar] [CrossRef]
- Bergonier, D.; Sobral, D.; Feßler, A.T.; Jacquet, E.; Gilbert, F.B.; Schwarz, S.; Treilles, M.; Bouloc, P.; Pourcel, C.; Vergnaud, G. Staphylococcus aureus from 152 cases of bovine, ovine and caprine mastitis investigated by Multiple-locus variable number of tandem repeat analysis (MLVA). Vet. Res. 2014, 45, 1–8. [Google Scholar] [CrossRef]
- Li, T.; Lu, H.; Wang, X.; Gao, Q.; Dai, Y.; Shang, J.; Li, M. Molecular characteristics of Staphylococcus aureus causing bovine mastitis between 2014 and 2015. Front. Cell. Infect. Microbiol. 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Le Loir, Y.; Baron, F.; Gautier, M. Staphylococcus aureus and food poisoning. Genet. Mol. Res. 2003, 2, 63–76. [Google Scholar]
- Sergelidis, D.; Angelidis, A.S. Methicillin-resistant Staphylococcus aureus: A controversial food-borne pathogen. Lett. Appl. Microbiol. 2017, 64, 409–418. [Google Scholar] [CrossRef] [Green Version]
- McMillan, K.; Moore, S.C.; McAuley, C.M.; Fegan, N.; Fox, E.M. Characterization of Staphylococcus aureus isolates from raw milk sources in Victoria, Australia. BMC Microbiol. 2016, 16, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Piccinini, R.; Tassi, R.; Daprà, V.; Pilla, R.; Fenner, J.; Carter, B.; Anjum, M.F. Study of Staphylococcus aureus collected at slaughter from dairy cows with chronic mastitis. J. Dairy Res. 2012, 79, 249–255. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, L.; Yong, C.; Shen, M.; Ali, T.; Shahid, M.; Han, K.; Zhou, X.; Han, B. Relationships among superantigen toxin gene profiles, genotypes, and pathogenic characteristics of Staphylococcus aureus isolates from bovine mastitis. J. Dairy Sci. 2017, 100, 4276–4286. [Google Scholar] [CrossRef]
- Angelidis, A.S.; Komodromos, D.; Giannakou, R.; Arsenos, G.; Gelasakis, A.I.; Kyritsi, M.; Filioussis, G.; Hadjichristodoulou, C.; Torounidou, P.; Papa, A.; et al. Isolation and characterization of Staphylococcus aureus and methicillin-resistant Staphylococcus aureus (MRSA) from milk of dairy goats under low-input farm management in Greece. Vet. Microbiol. 2020, 247, 108749. [Google Scholar] [CrossRef] [PubMed]
- Trunk, T.; Khalil, H.S.; Leo, J.C. Bacterial autoaggregation. AIMS Microbiol. 2018, 4, 140–164. [Google Scholar] [CrossRef] [PubMed]
- Romanò, A.; Gazzola, A.; Bianchini, V.; Cortimiglia, C.; Maisano, A.M.; Cremonesi, P.; Graber, H.U.; Vezzoli, F.; Luini, M. Staphylococcus aureus From Goats Are Genetically Heterogeneous and Distinct to Bovine Ones. Front. Vet. Sci. 2020, 7, 1–9. [Google Scholar] [CrossRef]
- Zdragas, A.; Papadopoulos, T.; Mitsopoulos, I.; Samouris, G.; Vafeas, G.; Boukouvala, E.; Aikateriniadou, L.; Kyriaki, M.; Alexopoulos, A.; Lagka, V. Prevalence, genetic diversity, and antimicrobial susceptibility profiles of Staphylococcus aureus isolated from bulk tank milk from Greek traditional ovine farms. Small Rumin. Res. 2015, 125, 120–126. [Google Scholar] [CrossRef]
- Pexara, A.; Solomakos, N.; Sergelidis, D.; Angelidis, A.S.; Govaris, A. Occurrence and antibiotic resistance of enterotoxigenic Staphylococcus aureus in raw ovine and caprine milk in Greece. Dairy Sci. Technol. 2016, 96, 345–357. [Google Scholar] [CrossRef]
- Papadopoulos, P.; Angelidis, A.S.; Papadopoulos, T.; Kotzamanidis, C.; Zdragas, A.; Papa, A.; Filioussis, G.; Sergelidis, D. Staphylococcus aureus and methicillin-resistant S. aureus (MRSA) in bulk tank milk, livestock and dairy-farm personnel in north-central and north-eastern Greece: Prevalence, characterization and genetic relatedness. Food Microbiol. 2019, 84. [Google Scholar] [CrossRef]
- Merz, A.; Stephan, R.; Johler, S. Staphylococcus aureus isolates from goat and sheep milk seem to be closely related and differ from isolates detected from bovine milk. Front. Microbiol. 2016, 7, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saei, H.D.; Ahmadi, M.; Mardani, K.; Batavani, R.A. Molecular typing of Staphylococcus aureus isolated from bovine mastitis based on polymorphism of the coagulase gene in the north west of Iran. Vet. Microbiol. 2009, 137, 202–206. [Google Scholar] [CrossRef]
- Van Leeuwen, W.B.; Melles, D.C.; Alaidan, A.; Al-Ahdal, M.; Boelens, H.A.M.; Snijders, S.V.; Wertheim, H.; Van Duijkeren, E.; Peeters, J.K.; Van Der Spek, P.J.; et al. Host- and tissue-specific pathogenic traits of Staphylococcus aureus. J. Bacteriol. 2005, 187, 4584–4591. [Google Scholar] [CrossRef] [Green Version]
- Mørk, T.; Tollersrud, T.; Kvitle, B.; Jørgensen, H.J.; Waage, S. Comparison of Staphylococcus aureus genotypes recovered from cases of bovine, ovine, and caprine mastitis. J. Clin. Microbiol. 2005, 43, 3979–3984. [Google Scholar] [CrossRef] [Green Version]
- Aires-de-Sousa, M.; Parente, C.E.S.R.; Vieira-da-Motta, O.; Bonna, I.C.F.; Silva, D.A.; De Lencastre, H. Characterization of Staphylococcus aureus isolates from buffalo, bovine, ovine, and caprine milk samples collected in Rio de Janeiro State, Brazil ∇. Appl. Environ. Microbiol. 2007, 73, 3845–3849. [Google Scholar] [CrossRef] [Green Version]
- Concepción Porrero, M.; Hasman, H.; Vela, A.I.; Fernández-Garayzábal, J.F.; Domínguez, L.; Aarestrup, F.M. Clonal diversity of Staphylococcus aureus originating from the small ruminants goats and sheep. Vet. Microbiol. 2012, 156, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, P.; Papadopoulos, T.; Angelidis, A.S.; Kotzamanidis, C.; Zdragas, A.; Papa, A.; Filioussis, G.; Sergelidis, D. Prevalence, antimicrobial susceptibility and characterization of Staphylococcus aureus and methicillin-resistant Staphylococcus aureus isolated from dairy industries in north-central and north-eastern Greece. Int. J. Food Microbiol. 2019, 291. [Google Scholar] [CrossRef] [PubMed]
- Aires-De-Sousa, M.; Boye, K.; De Lencastre, H.; Deplano, A.; Enright, M.C.; Etienne, J.; Friedrich, A.; Harmsen, D.; Holmes, A.; Huijsdens, X.W.; et al. High interlaboratory reproducibility of DNA sequence-based typing of bacteria in a multicenter study. J. Clin. Microbiol. 2006, 44, 619–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jørgensen, H.J.; Mørk, T.; Caugant, D.A.; Kearns, A.; Rørvik, L.M. Genetic variation among Staphylococcus aureus strains from Norwegian bulk milk. Appl. Environ. Microbiol. 2005, 71, 8352–8361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lather, P.; Mohanty, A.K.; Jha, P.; Garsa, A.K. Contribution of Cell Surface Hydrophobicity in the Resistance of Staphylococcus aureus against Antimicrobial Agents. Biochem. Res. Int. 2016, 2016. [Google Scholar] [CrossRef] [Green Version]
- Käppeli, N.; Morach, M.; Corti, S.; Eicher, C.; Stephan, R.; Johler, S. Staphylococcus aureus related to bovine mastitis in Switzerland: Clonal diversity, virulence gene profiles, and antimicrobial resistance of isolates collected throughout 2017. J. Dairy Sci. 2019, 102, 3274–3281. [Google Scholar] [CrossRef] [Green Version]
- Hoekstra, J.; Zomer, A.L.; Rutten, V.P.M.G.; Benedictus, L.; Stegeman, A.; Spaninks, M.P.; Bennedsgaard, T.W.; Biggs, A.; De Vliegher, S.; Mateo, D.H.; et al. Genomic analysis of European bovine Staphylococcus aureus from clinical versus subclinical mastitis. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Rocha, L.S.; Silva, D.M.; Silva, M.P.; Vidigal, P.M.P.; Silva, J.C.F.; Guerra, S.T.; Ribeiro, M.G.; De Mendes, T.A.O.; De Ribon, A.O.B. Comparative genomics of Staphylococcus aureus associated with subclinical and clinical bovine mastitis. PLoS ONE 2019, 14, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naushad, S.; Nobrega, D.B.; Naqvi, S.A.; Barkema, H.W.; De Buck, J. Genomic Analysis of Bovine Staphylococcus aureus Isolates from Milk To Elucidate Diversity and Determine the Distributions of Antimicrobial and Virulence Genes and Their Association with Mastitis. mSystems 2020, 5, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Haveri, M.; Roslöf, A.; Rantala, L.; Pyörälä, S. Virulence genes of bovine Staphylococcus aureus from persistent and nonpersistent intramammary infections with different clinical characteristics. J. Appl. Microbiol. 2007, 103, 993–1000. [Google Scholar] [CrossRef]
- Pichette-Jolette, S.; Millette, G.; Demontier, E.; Bran-Barrera, D.; Cyrenne, M.; Ster, C.; Haine, D.; Keefe, G.; Malouin, F.; Roy, J.P. Partial prediction of the duration and the clinical status of Staphylococcus aureus bovine intramammary infections based on the phenotypic and genotypic analysis of isolates. Vet. Microbiol. 2019, 228, 188–195. [Google Scholar] [CrossRef]
- Fang, R.; Cui, J.; Cui, T.; Guo, H.; Ono, H.K.; Park, C.H.; Okamura, M.; Nakane, A.; Hu, D.L. Staphylococcal enterotoxin C is an important virulence factor for mastitis. Toxins 2019, 11, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etter, D.; Schelin, J.; Schuppler, M.; Johler, S. Staphylococcal Enterotoxin C—An Update on SEC Variants, Their Structure and Properties, and Their Role in Foodborne Intoxications. Toxins 2020, 12, 584. [Google Scholar] [CrossRef]
- Veh, K.A.; Klein, R.C.; Ster, C.; Keefe, G.; Lacasse, P.; Scholl, D.; Roy, J.P.; Haine, D.; Dufour, S.; Talbot, B.G.; et al. Genotypic and phenotypic characterization of Staphylococcus aureus causing persistent and nonpersistent subclinical bovine intramammary infections during lactation or the dry period. J. Dairy Sci. 2015, 98, 155–168. [Google Scholar] [CrossRef] [Green Version]
- Grunert, T.; Stessl, B.; Wolf, F.; Sordelli, D.O.; Buzzola, F.R.; Ehling-Schulz, M. Distinct phenotypic traits of Staphylococcus aureus are associated with persistent, contagious bovine intramammary infections. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, L.K.; Zadoks, R.N.; Gaskins, C.T. Biofilm production by Staphylococcus aureus associated with intramammary infection. Vet. Microbiol. 2005, 107, 295–299. [Google Scholar] [CrossRef]
- Fragkou, I.A.; Boscos, C.M.; Fthenakis, G.C. Diagnosis of clinical or subclinical mastitis in ewes. Small Rumin. Res. 2014, 118, 86–92. [Google Scholar] [CrossRef]
- Hookey, J.V.; Richardson, J.F.; Cookson, B.D. Molecular typing of Staphylococcus aureus based on PCR restriction fragment length polymorphism and DNA sequence analysis of the coagulase gene. J. Clin. Microbiol. 1998, 36, 1083–1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hygiene, F.; Sudagidan, M.; Aydin, A. Screening virulence properties of staphylococci. Wien. Tierarztl. Mon. 2008, 95, 128–134. [Google Scholar]
- Løvseth, A.; Loncarevic, S.; Berdal, K.G. Modified multiplex PCR method for detection of pyrogenic exotoxin genes in staphylococcal isolates. J. Clin. Microbiol. 2004, 42, 3869–3872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stegger, M.; Andersen, P.S.; Kearns, A.; Pichon, B.; Holmes, M.A.; Edwards, G.; Laurent, F.; Teale, C.; Skov, R.; Larsen, A.R. Rapid detection, differentiation and typing of methicillin-resistant Staphylococcus aureus harbouring either mecA or the new mecA homologue mecALGA251. Clin. Microbiol. Infect. 2012, 18, 395–400. [Google Scholar] [CrossRef] [Green Version]
- McDougal, L.K.; Steward, C.D.; Killgore, G.E.; Chaitram, J.M.; McAllister, S.K.; Tenover, F.C. Pulsed-Field Gel Electrophoresis Typing of Oxacillin-Resistant Staphylococcus aureus Isolates from the United States: Establishing a National Database. J. Clin. Microbiol. 2003, 41, 5113–5120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, P.R.; Gaston, M.A. Numerical index of the discriminatory ability of typing systems: An application of Simpson’s index of diversity. J. Clin. Microbiol. 1988, 26, 2465–2466. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Yu, F.; Yang, L.; Li, Q.; Zhang, X.; Zeng, Y. Prevalence of virulence genes and biofilm formation among Staphylococcus aureus clinical isolates associated with lower respiratory infection. African J. Microbiol. Res. 2010, 4, 2566–2569. [Google Scholar]
- Borges, S.; Silva, J.; Teixeira, P. Survival and biofilm formation by Group B streptococci in simulated vaginal fluid at different pHs. Antonie Van Leeuwenhoek 2012, 677–682. [Google Scholar] [CrossRef]
- Kos, B.; Šušković, J.; Vuković, S.; Sǐmpraga, M.; Frece, J.; Matošić, S. Adhesion and aggregation ability of probiotic strain Lactobacillus acidophilus M92. J. Appl. Microbiol. 2003, 94, 981–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.M.; Kim, W.S.; Kumura, H.; Shimazaki, K.I. Autoaggregation and surface hydrophobicity of bifidobacteria. World J. Microbiol. Biotechnol. 2008, 24, 1593–1598. [Google Scholar] [CrossRef]
- Crow, V.L.; Gopal, P.K.; Wicken, A.J. Cell surface differences of lactococcal strains. Int. Dairy J. 1995, 5, 45–68. [Google Scholar] [CrossRef]
- Niederle, M.V.; Bosch, J.; Ale, C.E.; Nader-Macías, M.E.; Aristimuño Ficoseco, C.; Toledo, L.F.; Valenzuela-Sánchez, A.; Soto-Azat, C.; Pasteris, S.E. Skin-associated lactic acid bacteria from North American bullfrogs as potential control agents of Batrachochytrium dendrobatidis. PLoS ONE 2019, 14. [Google Scholar] [CrossRef] [Green Version]
- Clinical Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically; Approved Standard—Ninth Edition. CLSI document M07-A9. Clin. Lab. Standars Inst. 2018, 32, 18. [Google Scholar]
- Magiorakos, A.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F. Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Microbiology 2011, 18, 268–281. [Google Scholar]
- Kaklamanos, E.G.; Charalampidou, M.; Menexes, G.; Topitsoglou, V.; Kalfas, S. Transient oral microflora in Greeks attending day centres for the elderly and residents in homes for the elderly. Gerodontology 2005, 22, 158–167. [Google Scholar] [CrossRef] [PubMed]


 ) represent clinical and subclinical mastitis cases.
) represent clinical and subclinical mastitis cases.
) represent clinical and subclinical mastitis cases.
) represent clinical and subclinical mastitis cases.

{kind=link}
{kind=link}
{kind=link}
| No. of Isolates (%) | ||||||
|---|---|---|---|---|---|---|
| Ovine (n = 97) | Caprine (n = 29) | Bovine (n = 36) | Total (n = 162) | |||
| Clinical (n = 34) | Subclinical (n = 63) | Clinical (n = 4) | Subclinical (n = 25) | Clinical | ||
| Toxin genes | ||||||
| sea | 0 (0) | 9 (14.3) | 0 (0) | 1 (4.0) | 0 (0) | 10 (6.2) |
| seb | 5 (14.7) | 19 (30.2) | 1 (25.0) | 7 (28.0) | 12 (33.3) | 44 (27.1) |
| sec | 11 (32.3) | 18 (28.5) | 0 (0) | 8 (32.0) | 16 (44.4) | 53 (32.7) |
| sed | 0 (0) | 4 (6.3) | 0 (0) | 8 (0) | 1 (2.8) | 7 (4.3) |
| see | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| seh | 0 (0) | 3 (4.8) | 0 (0) | 0 (0) | 0 (0) | 3 (1.9) |
| seg | 1 (2.9) | 3 (4.8) | 0 (0) | 2 (8.0) | 0 (0) | 6 (3.7) |
| sei | 4 (11.8) | 18 (28.5) | 0 (0) | 7 (28.0) | 0 (0) | 29 (17.9) |
| sej | 1 (2.9) | 8 (12.7) | 1 (25.0) | 3 (12.0) | 8 (22.2) | 21 (13.0) |
| pvl | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| tsst | 1 (2.9) | 0 (0) | 0 (0) | 0 (0) | 1 (2.8) | 2 (1.2) |
| Toxin gene patterns | ||||||
| sea | 0 (0) | 2 (3.2) | 0 (0) | 0 (0) | 0 (0) | 2 (1.2) |
| seb | 2 (5.9) | 5 (7.9) | 1 (25.0) | 1 (4.0) | 2 (5.5) | 11 (6.7) |
| sec | 11 (32.3) | 10 (15.9) | 0 (0) | 0 (0) | 10 (27.8) | 34 (21.0) |
| sed | 0 (0) | 1 (1.6) | 0 (0) | 0 (0) | 0 (0) | 1 (0.6) |
| seh | 0 (0) | 1 (1.6) | 0 (0) | 0 (0) | 0 (0) | 1 (0.6) |
| sei | 0 (0) | 6 (9.5) | 0 (0) | 0 (0) | 0 (0) | 6 (3.7) |
| sej | 0 (0) | 0 (0) | 1 (25.0) | 1 (4.0) | 5 (13.9) | 7 (4.3) |
| tsst | 1 (2.9) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 1 (0.6) |
| sea, seb | 0 (0) | 1 (1.6) | 0 (0) | 0 (0) | 0 (0) | 1 (0.6) |
| sea, sei | 0 (0) | 0 (0) | 0 (0) | 1 (4.0) | 0 (0) | 1 (0.6) |
| sea, sej | 0 (0) | 1 (1.6) | 0 (0) | 0 (0) | 0 (0) | 1 (0.6) |
| seb, sec | 0 (0) | 2 (3.2) | 0 (0) | 1 (4.0) | 6 (16.7) | 9 (5.6) |
| seb, sei | 2 (5.9) | 4 (6.3) | 0 (0) | 2 (8.0) | 0 (0) | 8 (4.9) |
| seb, sej | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 3 (8.3) | 3 (1.9) |
| sec, sei | 0 (0) | 0 (0) | 0 (0) | 1 (4.0) | 0 (0) | 1 (0.6) |
| sec, sej | 0 (0) | 0 (0) | 0 (0) | 1 (4.0) | 0 (0) | 1 (0.6) |
| sed, seg | 0 (0) | 0 (0) | 0 (0) | 1 (4.0) | 0 (0) | 1 (0.6) |
| sei, sej | 2 (5.9) | 3 (4.8) | 0 (0) | 0 (0) | 0 (0) | 4 (2.5) |
| sea, sec, seh | 0 (0) | 2 (3.2) | 0 (0) | 0 (0) | 0 (0) | 2 (1.2) |
| sea, sei, sej | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 1 (2.8) | 1 (0.6) |
| seb, sec, sei | 0 (0) | 1 (1.6) | 0 (0) | 2 (8.0) | 0 (0) | 3 (1.9) |
| seb, sed, seg | 0 (0) | 2 (3.2) | 0 (0) | 1 (4.0) | 0 (0) | 3 (1.9) |
| seb, sed, tsst | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 1 (2.8) | 1 (0.6) |
| seb, seg, sei | 1 (2.9) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 1 (0.6) |
| sed, seg, sej | 0 (0) | 1 (1.6) | 0 (0) | 0 (0) | 0 (0) | 1 (0.6) |
| sea, seb, sec, sei | 0 (0) | 1 (1.6) | 0 (0) | 0 (0) | 0 (0) | 1 (0.6) |
| sea, seb, sei, sej | 0 (0) | 1 (1.6) | 0 (0) | 0 (0) | 0 (0) | 1 (0.6) |
| seb, sec, sei, sej | 0 (0) | 2 (3.2) | 0 (0) | 0 (0) | 0 (0) | 2 (1.2) |
| spa Type | No. of Isolates (%) | |||||
|---|---|---|---|---|---|---|
| Ovine (n = 97) | Caprine (n = 29) | Bovine (n = 36) | Total (n = 162) | |||
| Clinical (n = 34) | Subclinical (n = 63) | Clinical (n = 4) | Subclinical (n = 25) | Clinical | ||
| t012 | 1 (2.9) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 1 (0.6) |
| t031 | 1 (2.9) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 1 (0.6) |
| t085 | 0 (0) | 1 (1.6) | 0 (0) | 0 (0) | 0 (0) | 1 (0.6) |
| t116 | 0 (0) | 1 (1.6) | 0 (0) | 0 (0) | 0 (0) | 1 (0.6) |
| t1166 | 1 (2.9) | 0 (0) | 0 (0) | 0 (0) | 1 (2.8) | 2 (1.2) |
| t12028 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 1 (2.8) | 1 (0.6) |
| t148 | 0 (0) | 1 (1.6) | 0 (0) | 0 (0) | 0 (0) | 1 (0.6) |
| t14993 | 0 (0) | 1 (1.6) | 0 (0) | 0 (0) | 0 (0) | 1 (0.6) |
| t1532 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 1 (2.8) | 1 (0.6) |
| t1534 | 0 (0) | 1 (1.6) | 0 (0) | 1 (4.0) | 1 (2.8) | 3 (1.8) |
| t1773 | 7 (20.6) | 4 (6.3) | 1 (25.0) | 0 (0) | 1 (2.8) | 13 (8.0) |
| t1896 | 0 (0) | 2 (3.2) | 0 (0) | 0 (0) | 1 (2.8) | 3 (1.8) |
| t2516 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 1 (2.8) | 1 (0.6) |
| t267 | 0 (0) | 1 (1.6) | 0 (0) | 0 (0) | 0 (0) | 1 (0.6) |
| t2678 | 4 (11.8) | 0 (0) | 0 (0) | 0 (0) | 4 (11.1) | 8 (4.9) |
| t306 | 1 (2.9) | 2 (3.2) | 0 (0) | 2 (8.0) | 0 (0) | 5 (3.1) |
| t3568 | 1 (2.9) | 6 (9.5) | 0 (0) | 1 (4.0) | 0 (0) | 8 (4.9) |
| t3586 | 8 (23.5) | 18 (28.6) | 0 (0) | 8 (32.0) | 14 (38.9) | 48 (30.0) |
| t359 | 1 (2.9) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 1 (0.6) |
| t3739 | 1 (2.9) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 1 (0.6) |
| t4038 | 2 (5.9) | 14 (22.2) | 1 (25.0) | 5 (20.0) | 6 (16.7) | 28 (17.3) |
| t524 | 3 (8.8) | 0 (0) | 1 (25.0) | 0 (0) | 0 (0) | 4 (2.5) |
| t5428 | 1 (2.9) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 1 (0.6) |
| t548 | 0 (0) | 2 (3.2) | 0 (0) | 1 (4.0) | 0 (0) | 3 (1.8) |
| t5728 | 1 (2.9) | 0 (0) | 0 (0) | 0 (0) | 1 (2.8) | 2 (1.2) |
| t6058 | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 1 (2.8) | 1 (0.6) |
| t693 | 0 (0) | 1 (1.6) | 0 (0) | 0 (0) | 0 (0) | 1 (0.6) |
| t7008 | 1 (2.9) | 1 (1.6) | 0 (0) | 0 (0) | 0 (0) | 2 (1.2) |
| t701 | 0 (0) | 2 (3.2) | 0 (0) | 0 (0) | 0 (0) | 2 (1.2) |
| t7946 | 0 (0) | 0 (0) | 0 (0) | 1 (4.0) | 0 (0) | 1 (0.6) |
| t7947 | 0 (0) | 3 (4.8) | 1 (25.0) | 0 (0) | 0 (0) | 4 (2.5) |
| t891 | 0 (0) | 1 (1.6) | 0 (0) | 0 (0) | 0 (0) | 1 (0.6) |
| NT | 0 (0) | 1 (1.6) | 0 (0) | 6 (24.0) | 3 (8.3) | 10 (6.2) |
| Phenotype | No. of Isolates (%) | |||||
|---|---|---|---|---|---|---|
| Ovine (n = 97) | Caprine (n = 29) | Bovine (n = 36) | Total (n = 162) | |||
| Clinical (n = 34) | Subclinical (n = 63) | Clinical (n = 4) | Subclinical (n = 25) | Clinical | ||
| Antimicrobial | ||||||
| resistance | ||||||
| Penicillin (P) | 10 (29.4) | 6 (9.6) | 1 (25.0) | 1 (4.0) | 9 (25.0) | 27 (16.7) |
| Erythromycin (E) | 1 (2.9) | 1 (1.6) | 0 (0) | 0 (0) | 1 (2.8) | 3 (1.9) |
| Tetracycline (TET) | 11 (32.4) | 2 (3.2) | 1 (25.0) | 0 (0) | 9 (25.0) | 23 (14.2) |
| Streptomycin (S) | 2 (5.9) | 10 (15.9) | 0 (0) | 3 (12.0) | 2 (5.6) | 17 (10.5) |
| Antimicrobial resistance | ||||||
| patterns | ||||||
| P | 2 (5.9) | 4 (6.3) | 1 (25.0) | 1 (4.0) | 5 (13.9) | 13 (8.0) |
| E | 1 (2.9) | 1 (1.6) | 0 (0) | 0 (0) | 1 (2.8) | 3 (4.8) |
| TET | 5 (14.7) | 0 (0) | 1 (25.0) | 0 (0) | 5 (13.9) | 11 (6.8) |
| S | 0 (0) | 8 (12.7) | 0 (0) | 3 (12.0) | 2 (5.6) | 13 (8.0) |
| P-S | 2 (5.9) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 2 (1.2) |
| P-TET | 6 (17.6) | 0 (0) | 0 (0) | 0 (0) | 4 (11.1) | 10 (6.2) |
| P-TET-S | 0 (0) | 2 (3.2) | 0 (0) | 0 (0) | 0 (0) | 2 (1.2) |
| Biofilm production | ||||||
| ability | ||||||
| No biofilm | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| Weak | 0 (0) | 6 (9.5) | 0 | 3 (12.0) | 0 | 9 (5.6) |
| Moderate | 14 (41.2) | 25 (39.7) | 2 (50.0) | 9 (36.0) | 5 (13.9) | 55 (34.0) |
| Strong | 20 (58.8) | 32 (50.8) | 2 (50.0) | 13 (52.0) | 31 (86.1) | 98 (60.4) |
| Hydrophobicity | ||||||
| Low | 34 (100) | 63 (1000 | 4 (100) | 25 (100) | 36 (100) | 162 (100) |
| Medium | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| High | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) | 0 (0) |
| Autoaggregation | ||||||
| ability | ||||||
| Low | 13 (38.2) | 55 (87.3) | 0 (0) | 23 (92.0) | 9 (25.0) | 100 (61.7) |
| High | 21 (61.8) | 8 (12.7) | 4 (100) | 2 (8.0) | 27 (75.0) | 62 (38.3) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kotzamanidis, C.; Vafeas, G.; Giantzi, V.; Anastasiadou, S.; Mygdalias, S.; Malousi, A.; Loukia, E.; Daniel, S.; Zdragas, A. Staphylococcus aureus Isolated from Ruminants with Mastitis in Northern Greece Dairy Herds: Genetic Relatedness and Phenotypic and Genotypic Characterization. Toxins 2021, 13, 176. https://doi.org/10.3390/toxins13030176
Kotzamanidis C, Vafeas G, Giantzi V, Anastasiadou S, Mygdalias S, Malousi A, Loukia E, Daniel S, Zdragas A. Staphylococcus aureus Isolated from Ruminants with Mastitis in Northern Greece Dairy Herds: Genetic Relatedness and Phenotypic and Genotypic Characterization. Toxins. 2021; 13(3):176. https://doi.org/10.3390/toxins13030176
Chicago/Turabian StyleKotzamanidis, Charalampos, George Vafeas, Virginia Giantzi, Sofia Anastasiadou, Stavros Mygdalias, Andigoni Malousi, Ekateriniadou Loukia, Sergelidis Daniel, and Antonios Zdragas. 2021. "Staphylococcus aureus Isolated from Ruminants with Mastitis in Northern Greece Dairy Herds: Genetic Relatedness and Phenotypic and Genotypic Characterization" Toxins 13, no. 3: 176. https://doi.org/10.3390/toxins13030176
APA StyleKotzamanidis, C., Vafeas, G., Giantzi, V., Anastasiadou, S., Mygdalias, S., Malousi, A., Loukia, E., Daniel, S., & Zdragas, A. (2021). Staphylococcus aureus Isolated from Ruminants with Mastitis in Northern Greece Dairy Herds: Genetic Relatedness and Phenotypic and Genotypic Characterization. Toxins, 13(3), 176. https://doi.org/10.3390/toxins13030176