Characterization and Toxicity of Crude Toxins Produced by Cordyceps fumosorosea against Bemisia tabaci (Gennadius) and Aphis craccivora (Koch)



Abstract
:1. Introduction
2. Results
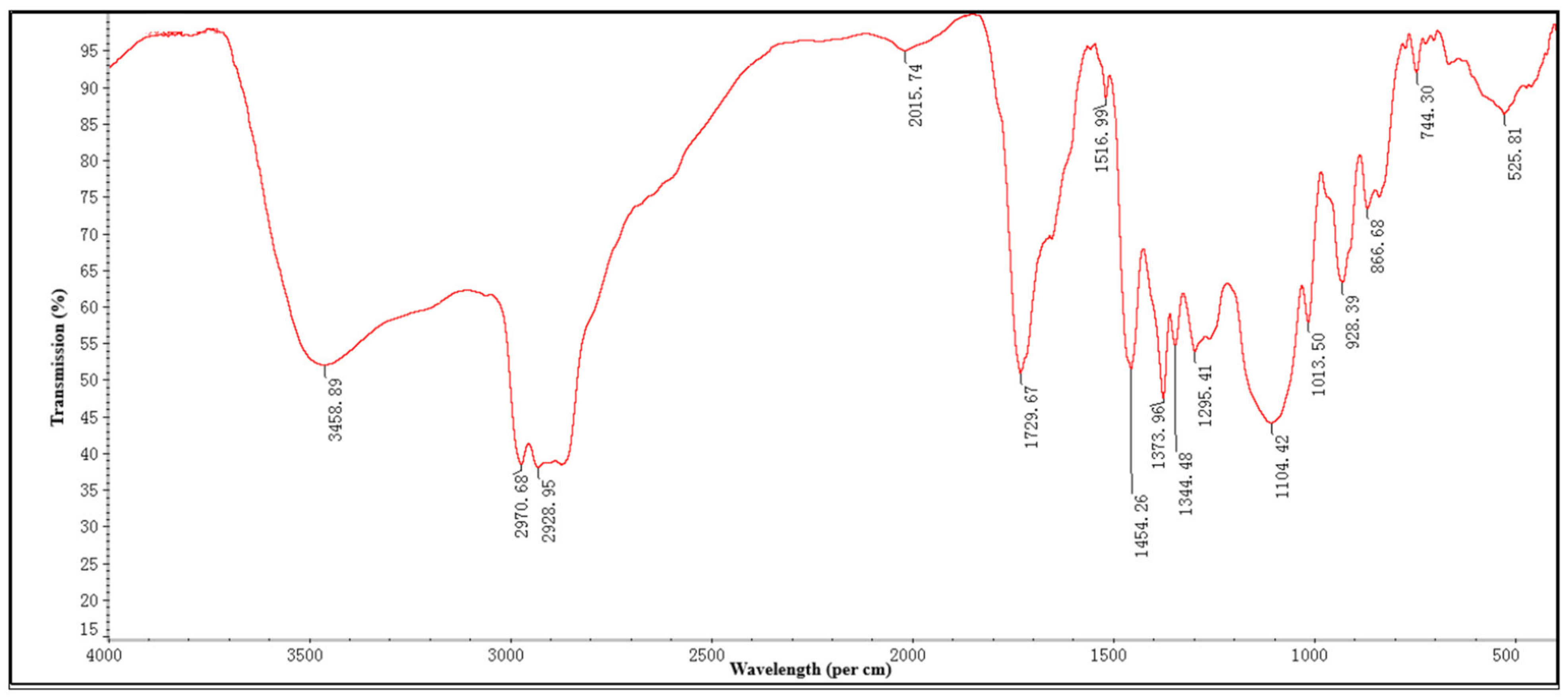
2.1. Fourier Transformed Infrared Spectroscopy (FTIR)
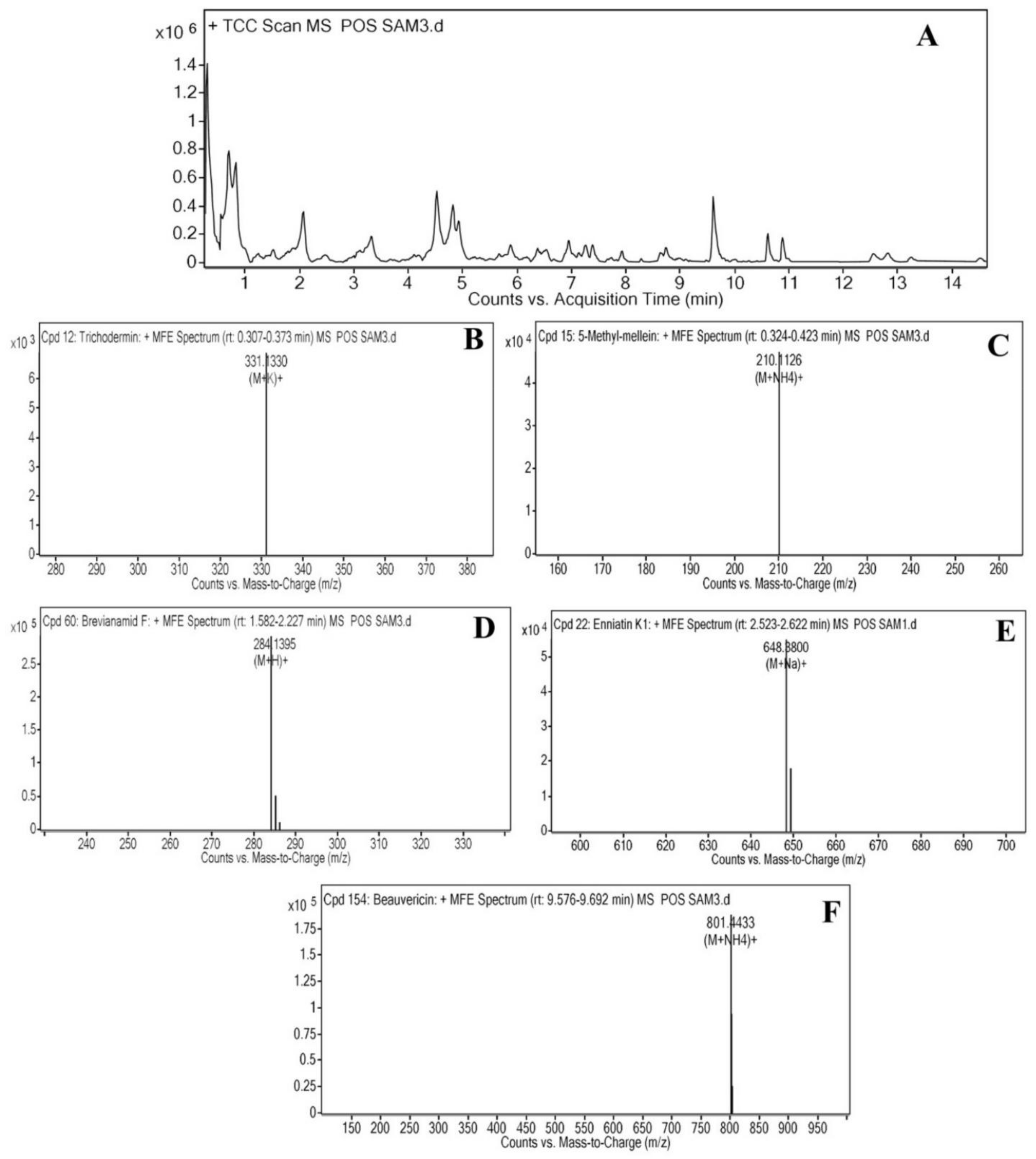
2.2. Liquid Chromatography-Mass Spectrometry Analysis of Cordyceps fumosorosea
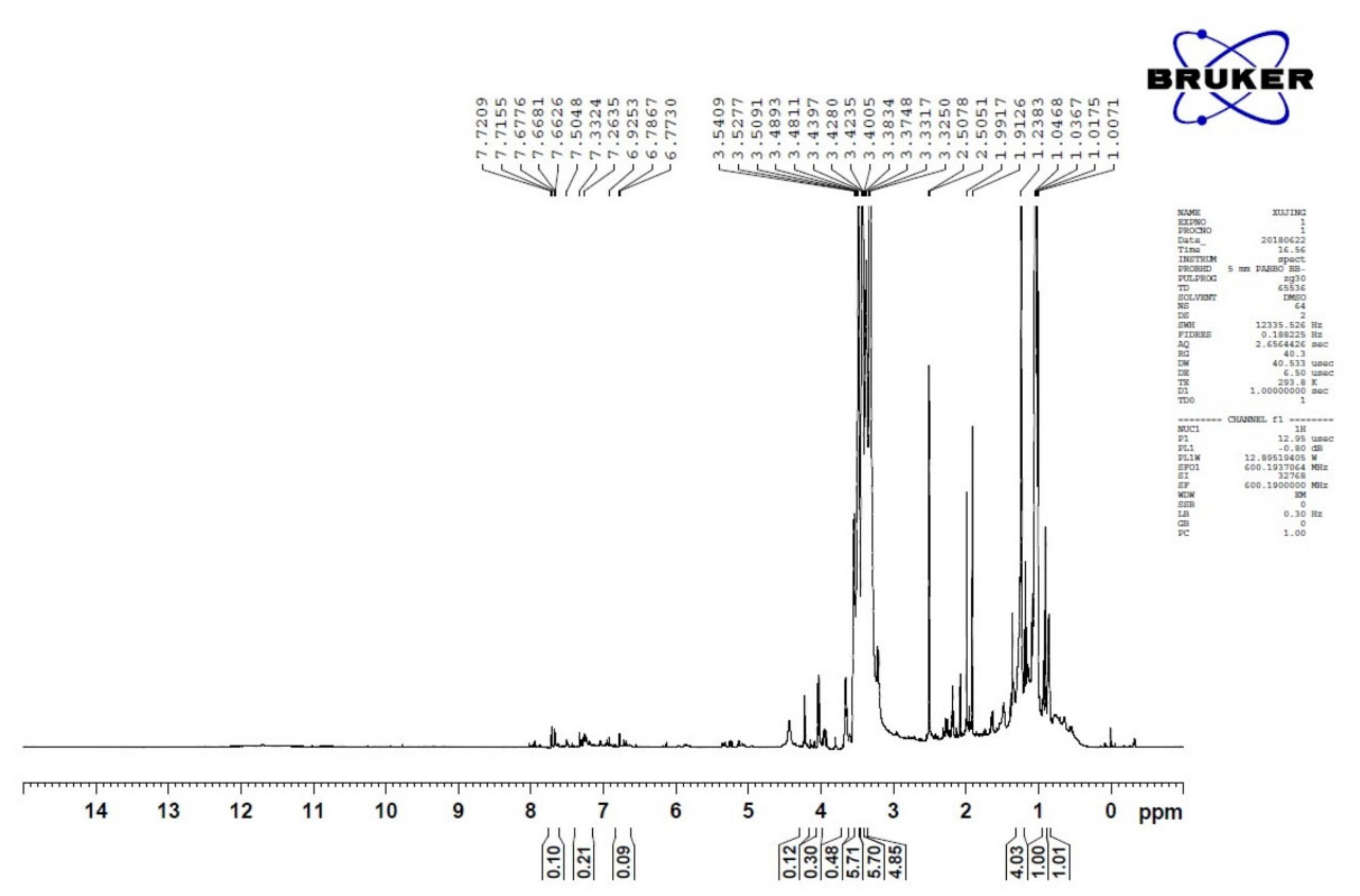
2.3. Nuclear Magnetic Resonance (NMR)
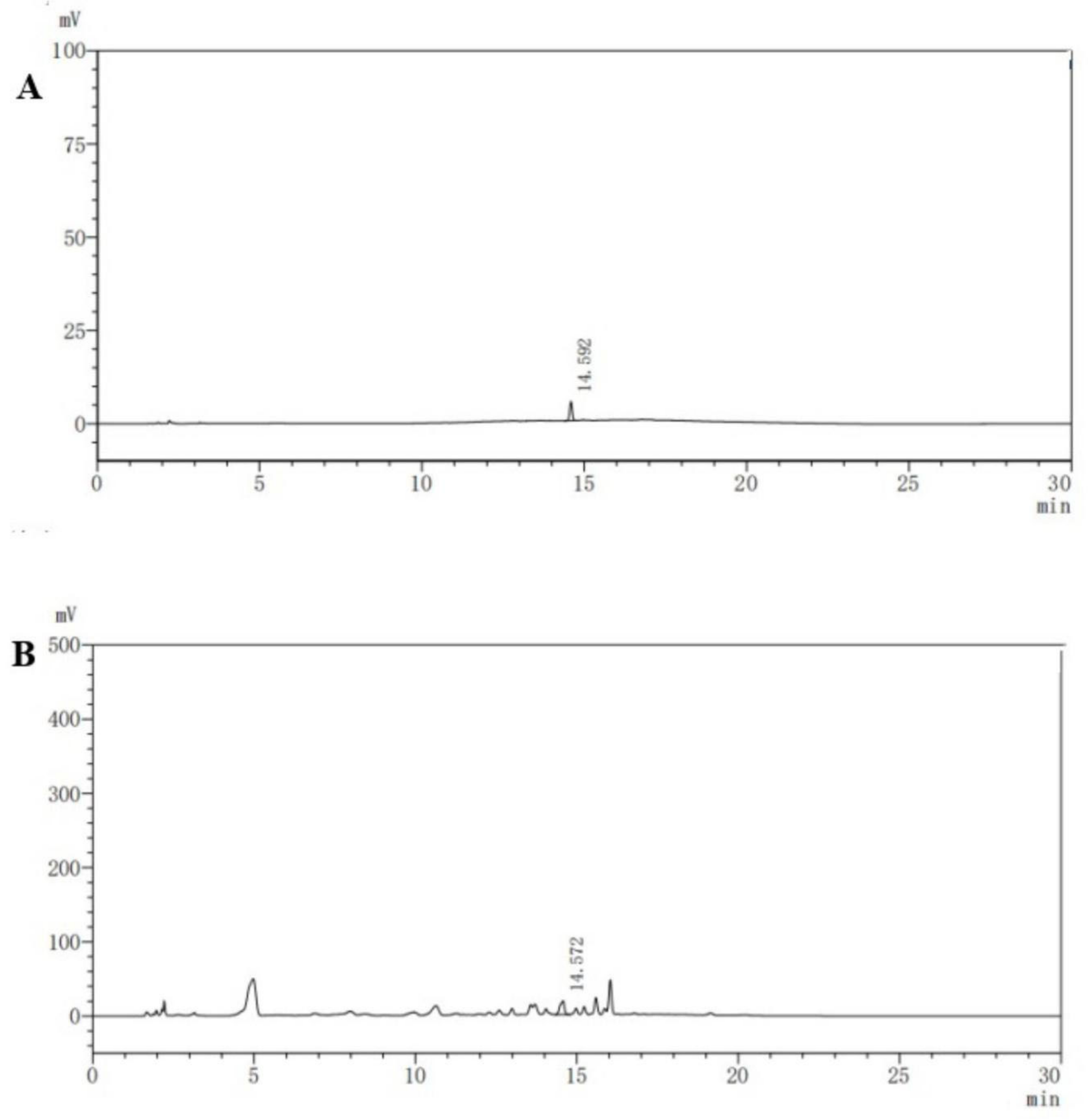
2.4. HPLC Analysis
2.5. Toxicity of C. fumosorosea Secondary Metabolites Against Immature Instars of B. tabaci and A. craccivora
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Insect Cultures
5.2. Fungal Inoculum and Culture Conditions
5.3. Secondary Metabolite Extraction from Cordyceps fumosorosea
5.4. Liquid Chromatography-Mass Spectrophotometry (LC-MS) Analysis
5.5. Fourier Transformed Infrared Spectroscopy (FTIR)
5.6. Nuclear Magnetic Resonance (NMR)
5.7. High Performance Liquid Chromatography (HPLC) Analysis
5.8. Toxicity of C. fumosorosea Secondary Metabolites against Immature Instars of B. tabaci and A. craccivora
5.9. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hu, J.; De Barro, P.; Zhao, H.; Wang, J.; Nardi, F.; Liu, S.S. An extensive field survey combined with a phylogenetic analysis reveals rapid and widespread invasion of two alien whiteflies in China. PLoS ONE. 2011, 6, e16061. [Google Scholar] [CrossRef] [PubMed]
- Firdaus, S.; Vosman, B.; Hidayati, N.; Supena, E.; Darmo, J.; Visser, R.G.F.; van Heusden, A.W. The Bemisia tabaci species complex: Additions from different parts of the world. Insect Sci. 2013, 20, 723–733. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Vänninen, I. The importance of maintaining protected zone status against Bemisia tabaci. Insects 2015, 6, 432–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Ali, S.; Musa, P.D.; Wang, X.M.; Qiu, B.L. Evaluation of the pathogenecity of Aschersonia aleyrodis on Bemisia tabaci in the laboratory and greenhouse. Biocontrol Sci. Technol. 2017, 27, 210–221. [Google Scholar] [CrossRef]
- Perring, T.M. The Bemisia tabaci species complex. Crop Prot. 2001, 20, 725–737. [Google Scholar] [CrossRef]
- Stansly, P.A.; Naranjo, S.E. Bemisia: Bionomics and Management of a Global Pest; Springer: New York, NY, USA, 2010. [Google Scholar]
- Obopile, M.; Ositile, B. Life table and population parameters of cowpea aphid, Aphis craccivora Koch (Homoptera: Aphididae) on five cowpea Vigna unguiculata (L. Walp.) varieties. J. Pest Sci. 2010, 83, 9–14. [Google Scholar] [CrossRef]
- Tang, L.D.; Wu, J.H.; Ali, S.; Ren, S.X. The influence of different aphid prey species on biology and life table parameters of Propylea japonica. Biocontrol Sci. Technol. 2013, 23, 624–636. [Google Scholar] [CrossRef]
- Obopile, M. Economic threshold and injury levels for control of cowpea aphid, Aphis craccivora Linnaeus (Homoptera: Aphididae) on cowpea. Afr. Plant Prot. 2006, 12, 111–115. [Google Scholar]
- Ofuya, T.I. Control of the cowpea aphid, Aphis craccivora Koch (Homoptera: Aphididae) in cowpea, Vigna unguiculata (L.) Walp. Integr. Pest Manag. Rev. 1997, 2, 199–207. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Murchie, A.K. European red spider mite—An environmental consequence of persistent chemical pesticide application. Int. J. Environ. Sci. Technol. 2005, 2, 287–290. [Google Scholar] [CrossRef]
- He, S.; Wu, X.Y.; Zheng, J.Z.; Han, Z.Q.; Zhao, C.X.; Wang, Y.Y.; Chen, H.; Chen, C.; Han, J. A study of control effect of Bemisia tabaci (Gennadius) on greenhouse tomatoes using parasitoid Encarsia formosa Gahan. J. Anhui Agri. Sci. 2013, 41, 6244–6248. (In Chinese) [Google Scholar]
- Cuthbertson, A.G.S. Unnecessary pesticide applications in Northern Ireland apple orchards due to mis-identification of a beneficial mite species. Res. J. Chem. Environ. 2004, 8, 77–78. [Google Scholar]
- Cuthbertson, A.G.S.; Murchie, A.K. The environmental impact of an orchard winter wash and early season pesticide applications on both a beneficial and a pest mite species in Bramley apple orchards. Int. J. Environ. Sci. Technol. 2006, 3, 333–339. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.S.; Xu, J.; Wang, X.M.; Qiu, B.L.; Cuthbertson, A.G.S.; Du, C.L.; Wu, J.H.; Ali, S. Isaria fumosorosea-based-zero-valent iron nanoparticles affect the growth and survival of sweet potato whitefly, Bemisia tabaci (Gennadius). Pest Manag. Sci. 2019, 75, 2174–2181. [Google Scholar] [CrossRef] [PubMed]
- Du, C.L.; Yang, B.; Wu, J.H.; Ali, S. Identification and virulence characterization of two Akanthomyces attenuatus isolates against Megalurothrips usitatus (Thysanoptera: Thripidae). Insects 2019, 10, 168. [Google Scholar] [CrossRef] [Green Version]
- Du, C.L.; Wu, J.H.; Cuthbertson, A.G.S.; Bashir, M.H.; Sun, T.F.; Ali, S. Morphological, molecular and virulence characterisation of six Cordyceps spp. isolates infecting the diamondback moth, Pluttela xylostella. Biocontrol Sci. Technol. 2020. [Google Scholar] [CrossRef]
- Ali, S.; Huang, Z.; Ren, S.X. Production of cuticle degrading enzymes by Isaria fumosorosea and their evaluation as a biocontrol agent against diamondback moth. J. Pest. Sci. 2010, 83, 361–370. [Google Scholar] [CrossRef]
- Huang, Z.; Ali, S.; Ren, S.X.; Wu, J.H. The effect of Isaria fumosorosea on mortality and fecundity of Bemisia tabaci and Plutella xylostella. Insect Sci. 2010, 17, 140–148. [Google Scholar] [CrossRef]
- Pick, D.A.; Avery, P.B.; Hunter, W.B.; Powell, C.A.; Arthurs, S.P. Effect of Isaria fumosorosea (Hypocreales: Cordycipitaceae) and Lysiphlebus testaceipes, (Hymenoptera: Braconidae) on the brown citrus aphid: Preliminary assessment of a compatibility study. Fla. Entomol. 2012, 95, 764–766. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Audsley, N. Further screening of entomopathogenic fungi and nematodes as control agents for Drosophila suzukii. Insects 2016, 24, 7020024. [Google Scholar] [CrossRef]
- Bernardini, M.; Carilli, A.; Pacioini, G.; Santurbano, B. Isolation of beauvericin from Paecilomyces fumosoroseus. Phytochem. Rep. 1975, 14, 1865. [Google Scholar] [CrossRef]
- Grove, J.F.; Pople, M. The insecticidal activity of beauvericin and the enniatin complex. Mycopathologia 1980, 70, 103–105. [Google Scholar] [CrossRef]
- Weng, Q.; Zhang, X.; Chen, W.; Hu, Q. Secondary metabolites and the risks of Isaria fumosorosea and Isaria farinosa. Molecules 2019, 24, 664. [Google Scholar] [CrossRef] [Green Version]
- Qasim, M.; Islam, S.U.; Islam, W.; Noman, A.; Khan, K.A.; Hafeez, M.; Hussain, D.; Das, C.K.; Bamisisle, B.S.; Akutse, K.S.; et al. Characterization of mycotoxins from entomopathogenic fungi (Cordyceps fumosorosea) and their toxic effects to the development of asian citrus psyllid reared on healthy and diseased citrus plants. Toxicon 2020, 188, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Xu, J. Studies on the Extraction, Characterization and Toxicity of Toxins Produced by Different Isolates of Entomopathogenic Fungus Isaria fumosorosea. Master’s Thesis, South China Agricultural University, Guangzhou, China, 2018. [Google Scholar]
- Gupta, S.; Montllor, C.; Hwang, Y.S. Isolation of novel beauvericin analogues from the fungus Beauveria bassiana. J. Natl. Prod. 1995, 58, 733–738. [Google Scholar] [CrossRef]
- Ragavendran, C.; Dubey, N.K.; Natarajan, D. Beauveria bassina (Clavicipitaceae): A potent fungal agent for controlling mosquito vectors of Anopheles stephensi, Culex quinquefasciatus and Aedes aegypti (Diptera: Culicidae). RSC Adv. 2017, 7, 3838–3851. [Google Scholar] [CrossRef] [Green Version]
- Vivekanandhan, P.; Kavitha, T.; Karthi, S.; Senthil-Nathan, S.; Shivakumar, M.S. Toxicity of Beauveria bassiana-28 mycelial extracts on larvae of Culex quinquefasciatus mosquito (Diptera: Culicidae). Int. J. Environ. Res. Public Health 2018, 15, 440. [Google Scholar] [CrossRef] [Green Version]
- Supothina, S.; Srisanoh, U.; Nithithanaslip, S.; Tasanathai, K.; Luangsa-ard, J.J.; Li, C.R.; Isaka, M. Beauvericin production by the lepidoptera pathogenic fungus Isaria tenuipes: Analysis of natural specimens, synnemata from cultivation, and mycelia from liquid-media fermentation. Nat. Prod. Bioprospect. 2011, 1, 112–115. [Google Scholar] [CrossRef] [Green Version]
- Ravindran, K.; Akutse, K.S.; Sivaramakrishnan, S.; Wang, L. Determination and characterization of destruxin production in Metarhizium anisopliae Tk6 and formulations for Aedes aegypti mosquitoes control at the field level. Toxicon 2016, 120, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Huang, J.; You, M.S.; Guan, X.; Liu, B. Toxicity and feeding deterrence of crude toxin extracts of Lecanicillium (Verticillium) lecanii (Hyphomycetes) against sweet potato whitefly. Bemisia tabaci (Homoptera: Aleyrodidae). Pest Manag. Sci. 2017, 63, 381–387. [Google Scholar]
- Ekesi, S.; Akpa, A.D.; Onu, I.; Ogunlana, M.O. Entomopathogenicity of Beauveria bassiana and Metarhizium anisopliae to the cowpea aphid, Aphis craccivora Koch (Homoptera: Aphididae). Arch. Phytopathol. Plant Prot. 2000, 33, 171–180. [Google Scholar] [CrossRef]
- Ali, S.; Huang, Z.; Ren, S.X. Media composition influences on growth, enzyme activity and virulence of the entomopathogen hyphomycete Isaria fumosorosea. Entomol. Experiment. Appl. 2009, 131, 30–38. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Walters, K.F.A.; Northing, P. Susceptibility of Bemisia tabaci immature stages to the entomopathogenic fungus Lecanicillium muscarium on tomato and verbena foliage. Mycopathologia 2005, 159, 23–29. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Blackburn, L.F.; Northing, P.; Luo, W.; Cannon, R.J.C.; Walters, K.F.A. Leaf dipping as an environmental screening measure to test chemical efficacy against Bemisia tabaci on poinsettia plants. Int. J. Environ. Sci. Technol. 2009, 6, 347–352. [Google Scholar] [CrossRef] [Green Version]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Am. Mosq. Control Assoc. 1987, 3, 302–303. [Google Scholar] [CrossRef] [PubMed]
- SAS Institute. SAS User’s Guide; Statistics SAS Institute: Cary, NC, USA, 2000. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Observed Wave Numbers (cm−1) | Functional Group | Bonding Pattern |
|---|---|---|
| 3458.89 | O-H stretch alcohols or phenols | Strong, broad |
| 2970.68 | N-H alkane (Methyl) | Sharp |
| 2928.95 | C-H alkane (Methylene) | Strong, medium |
| 2015.74 | C≡C stretch nitrile | Weak |
| 1729.67 | C=O carboxylic acid | Strong, sharp |
| 1516.99 | C=C stretch alkane | Weak, sharp |
| 1454.26 | C=O trans alkenes | Strong, sharp |
| 1373.96 | C=C trans alkenes | Strong, sharp |
| 1344.48 | C=C trans alkenes | Weak, sharp |
| 1295.41 | C=C ester carbonyl group | Weak, medium |
| 1104.42 | C=C ether/alcohol | Strong, medium |
| 1013.50 | C-O trans-alkenes | Strong, sharp |
| 928.39 | C=C vinyl-alkenes | Strong, sharp |
| 866.68 | C=C aromatics | Strong, medium |
| 744.30 | C=C aromatics (Ring) | Weak, sharp |
| 525.81 | C=C Aromatics (Ring) | Weak, sharp |
| Sr# | Compound Name | Rt | Molecular Formula | Biological Activity |
|---|---|---|---|---|
| 1 | Trichodermin | 0.340 | C17H24O4 | Fungicide; Pesticide |
| 2 | 5-Methylmellein | 0.344 | C11H13O3 | Antimicrobial activity |
| 3 | Brevianamide F | 2.062 | C16H17N3 O2 | Antioxidant activity, mycotoxin |
| 4 | Enniatin | 2.565 | C32H55N3 O9 | Antimicrobial activity |
| 5 | Beauvericin | 9.615 | C45H57N3 O9 | Mycotoxin |
| Nymphal Instar | Concentration (mg/L) | Mortality (%) | LC50 (LCL–UCL) (mg/L) | LC90 (LCL–UCL) (mg/L) | χ2 (df)4 |
|---|---|---|---|---|---|
| 1st instar | ddH2O | 3.27 | 46.35 (30.89–66.65) | 346.83 (203.44–865.38) | 24.29 |
| Ethyl acetate | 3.67 | ||||
| Matrine | 51.16 | ||||
| 10 | 14.29 | ||||
| 20 | 36.67 | ||||
| 50 | 51.67 | ||||
| 100 | 62.97 | ||||
| 200 | 78.99 | ||||
| 500 | 97.84 | ||||
| 2nd instar | ddH2O | 2.34 | 54.55 (42.11–69.94) | 419.39 (280.43–747.78) | 11.28 |
| Ethyl acetate | 3.33 | ||||
| Matrine | 52.34 | ||||
| 10 | 12.91 | ||||
| 20 | 31.06 | ||||
| 50 | 48.21 | ||||
| 100 | 60.37 | ||||
| 200 | 77.04 | ||||
| 500 | 94.67 | ||||
| 3rd instar | ddH2O | 2.33 | 68.94 (56.14–84.77) | 539.42 (377.61–873.66) | 7.55 |
| Ethyl acetate | 3.19 | ||||
| Matrine | 48.67 | ||||
| 10 | 10.97 | ||||
| 20 | 24.67 | ||||
| 50 | 41.87 | ||||
| 100 | 58.09 | ||||
| 200 | 70.33 | ||||
| 500 | 92.33 | ||||
| 4th instar | ddH2O | 2.11 | 81.92 (73.47–91.52) | 714.74 (575.59–921.65) | 3.46 |
| Ethyl acetate | 3.56 | ||||
| Matrine | 47.92 | ||||
| 10 | 10.02 | ||||
| 20 | 23.17 | ||||
| 50 | 37.92 | ||||
| 100 | 54.08 | ||||
| 200 | 68.23 | ||||
| 500 | 87.67 |
| Nymphal Instar | Concentration (mg/L) | Mortality (%) | LC50 (LCL–UCL) (mg/L) | LC90 (LCL–UCL) (mg/L) | χ2 (df)4 |
|---|---|---|---|---|---|
| 1st instar | ddH2O | 2.01 | 62.67 (39.26–100.48) | 339.31 (185.15–1129.67) | 9.26 |
| Ethyl acetate | 3.16 | ||||
| Matrine | 54.23 | ||||
| 10 | 13.06 | ||||
| 20 | 21.23 | ||||
| 50 | 38.54 | ||||
| 100 | 53.67 | ||||
| 200 | 82.33 | ||||
| 500 | 100 | ||||
| 2nd instar | ddH2O | 1.06 | 72.84 (58.88–90.51) | 442.00 (311.92–712.57) | 4.06 |
| Ethyl acetate | 2.14 | ||||
| Matrine | 53.18 | ||||
| 10 | 12.18 | ||||
| 20 | 19.36 | ||||
| 50 | 35.87 | ||||
| 100 | 51.67 | ||||
| 200 | 78.23 | ||||
| 500 | 95.08 | ||||
| 3rd instar | ddH2O | 1.37 | 77.40 (61.89–97.56) | 535.70 (365.12–913.75) | 4.52 |
| Ethyl acetate | 2.23 | ||||
| Matrine | 46.72 | ||||
| 10 | 11.5 | ||||
| 20 | 18.29 | ||||
| 50 | 33.33 | ||||
| 100 | 51.33 | ||||
| 200 | 75.67 | ||||
| 500 | 94.33 | ||||
| 4th instar | ddH2O | 1.24 | 94.40 (75.48–119.81) | 653.27 (439.06–1139.34) | 2.31 |
| Ethyl acetate | 2.87 | ||||
| Matrine | 45.91 | ||||
| 10 | 10.6 | ||||
| 20 | 15.06 | ||||
| 50 | 29.33 | ||||
| 100 | 48.16 | ||||
| 200 | 72.34 | ||||
| 500 | 89.17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, J.; Yang, B.; Xu, J.; Cuthbertson, A.G.S.; Ali, S. Characterization and Toxicity of Crude Toxins Produced by Cordyceps fumosorosea against Bemisia tabaci (Gennadius) and Aphis craccivora (Koch). Toxins 2021, 13, 220. https://doi.org/10.3390/toxins13030220
Wu J, Yang B, Xu J, Cuthbertson AGS, Ali S. Characterization and Toxicity of Crude Toxins Produced by Cordyceps fumosorosea against Bemisia tabaci (Gennadius) and Aphis craccivora (Koch). Toxins. 2021; 13(3):220. https://doi.org/10.3390/toxins13030220
Chicago/Turabian StyleWu, Jianhui, Bo Yang, Jing Xu, Andrew G. S. Cuthbertson, and Shaukat Ali. 2021. "Characterization and Toxicity of Crude Toxins Produced by Cordyceps fumosorosea against Bemisia tabaci (Gennadius) and Aphis craccivora (Koch)" Toxins 13, no. 3: 220. https://doi.org/10.3390/toxins13030220
APA StyleWu, J., Yang, B., Xu, J., Cuthbertson, A. G. S., & Ali, S. (2021). Characterization and Toxicity of Crude Toxins Produced by Cordyceps fumosorosea against Bemisia tabaci (Gennadius) and Aphis craccivora (Koch). Toxins, 13(3), 220. https://doi.org/10.3390/toxins13030220