A 65-Day Fumonisin B Exposure at High Dietary Levels Has Negligible Effects on the Testicular and Spermatological Parameters of Adult Rabbit Bucks

 ,
,  , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
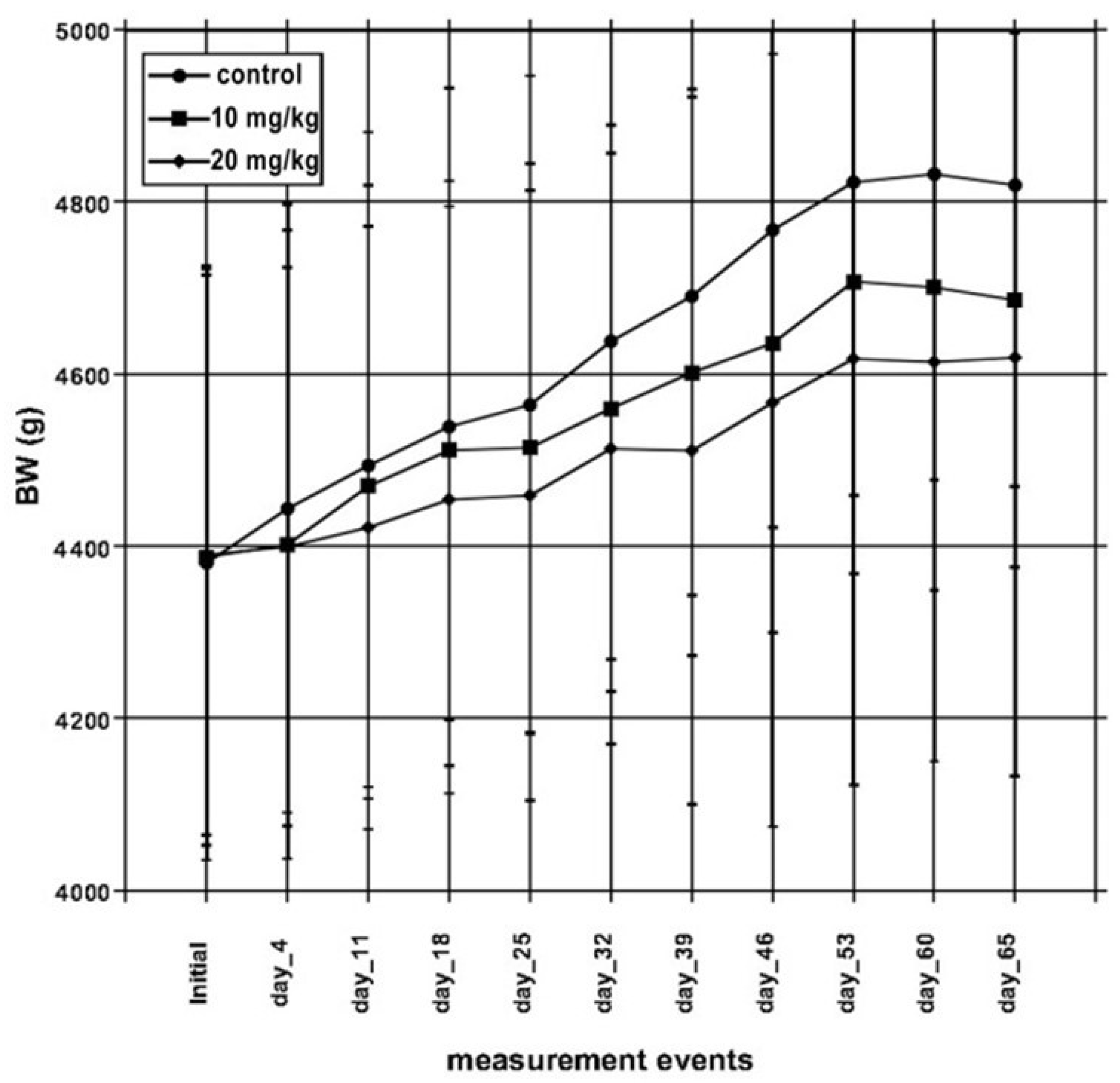
2.1. Animal Performance
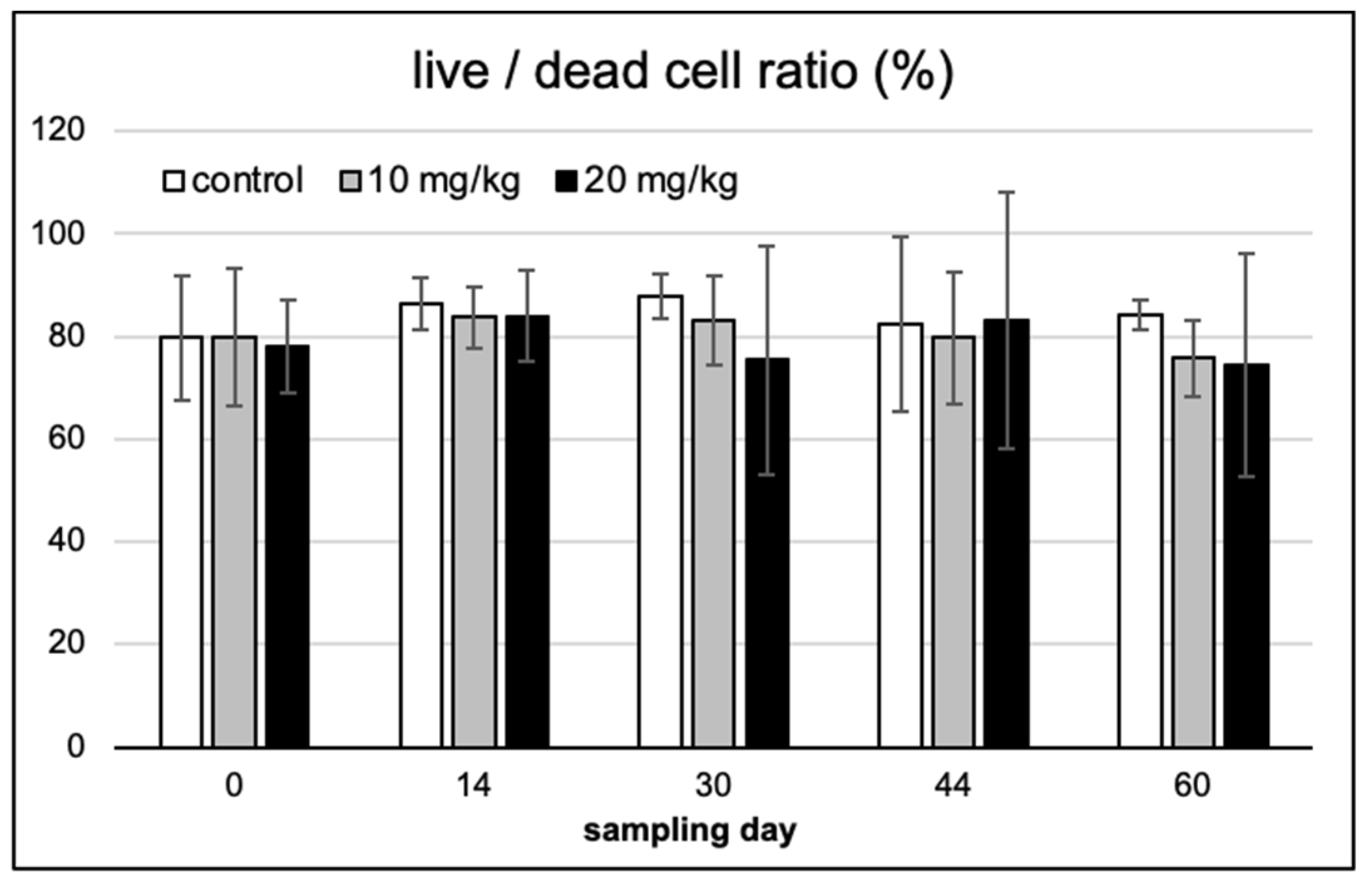
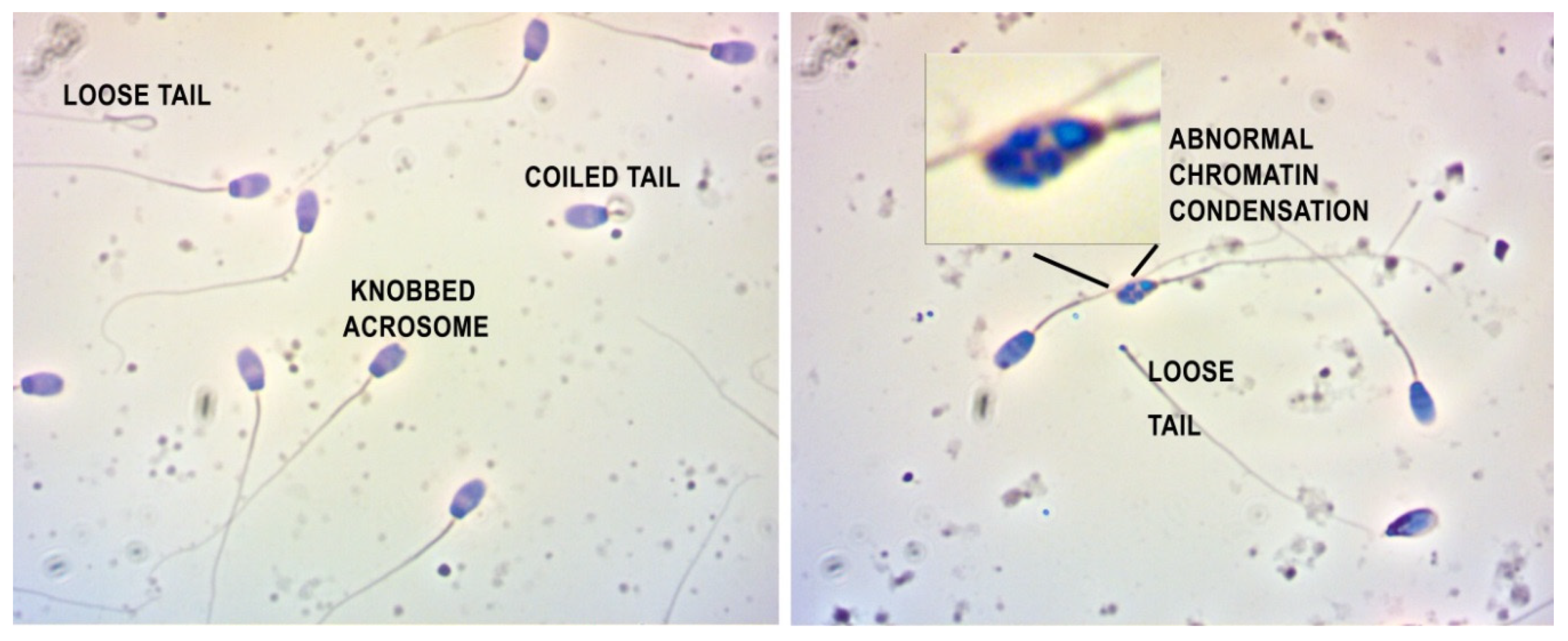
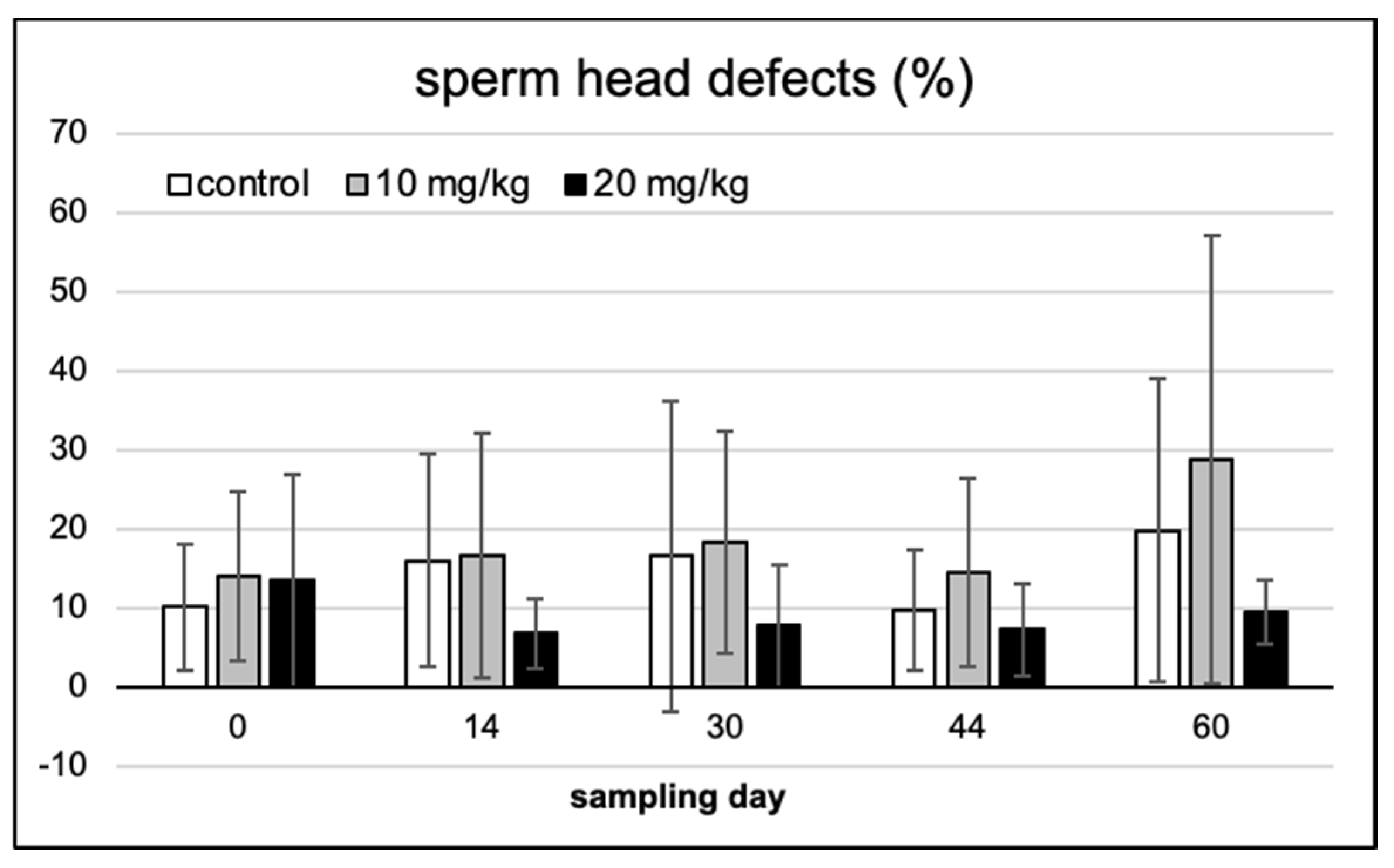
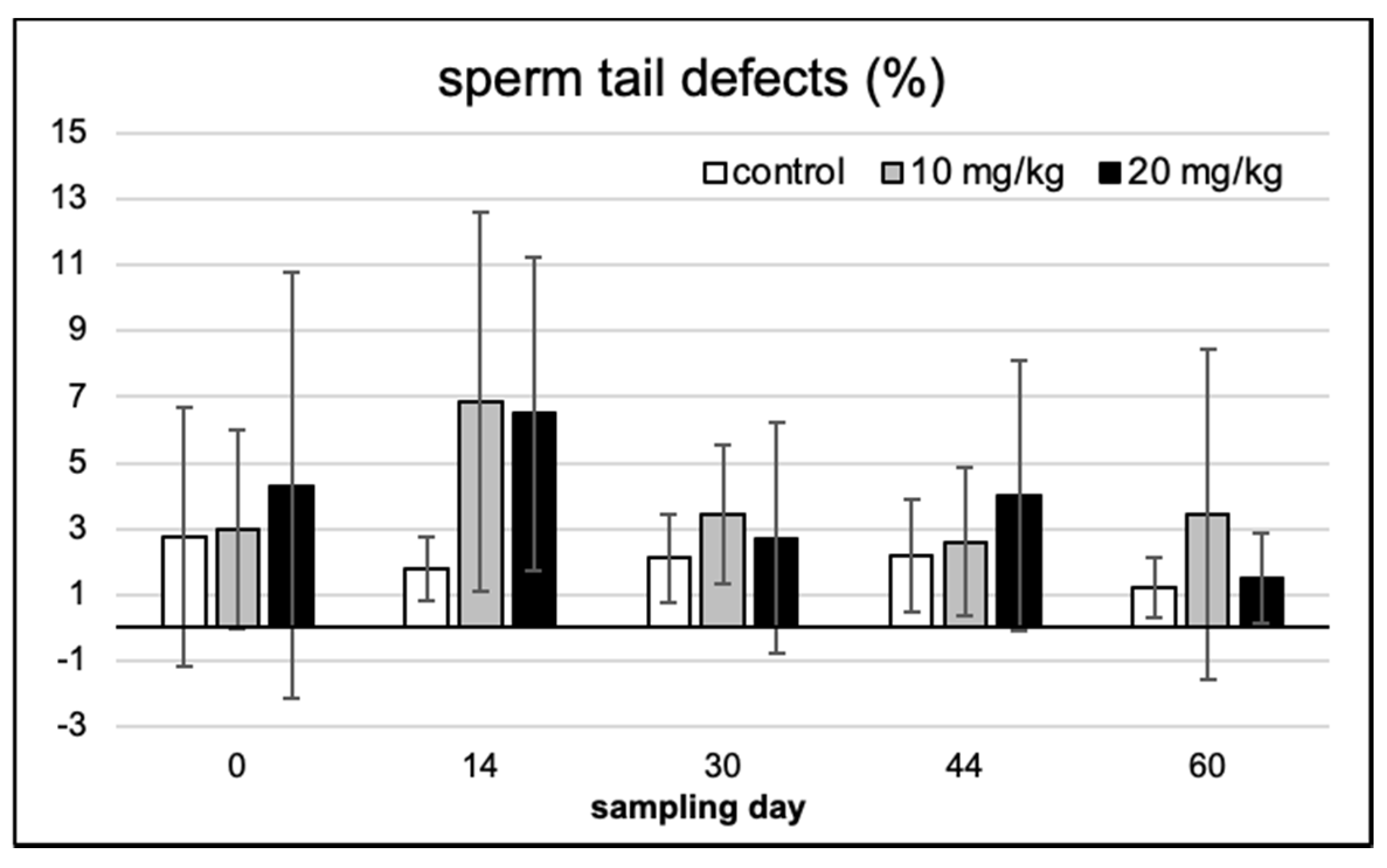
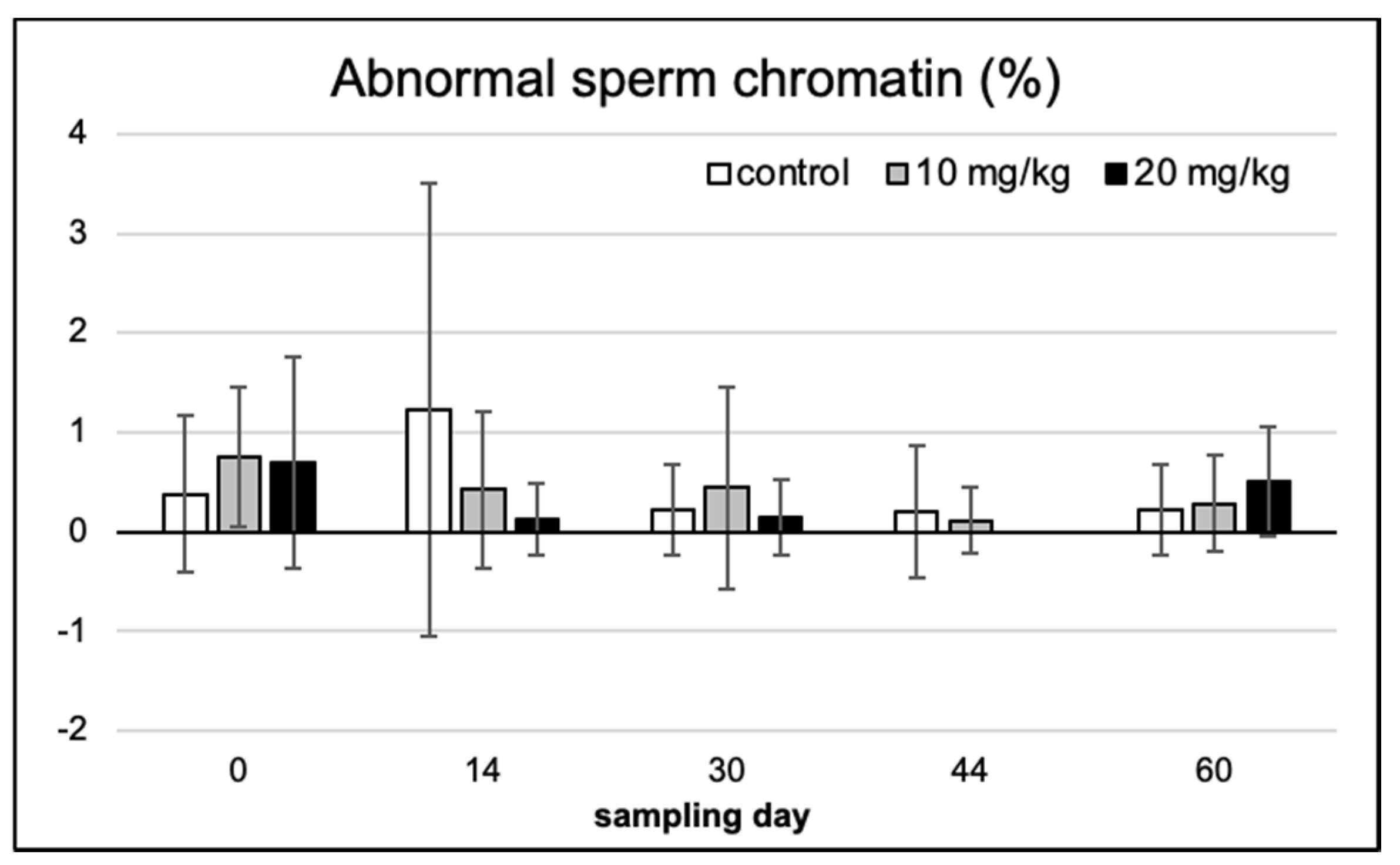
2.2. Spermium Morphology, Chromatin Integrity and Viability
2.3. Spermium Mophology and Distribution
2.4. Testicular Phospholipid Fatty Acid Composition
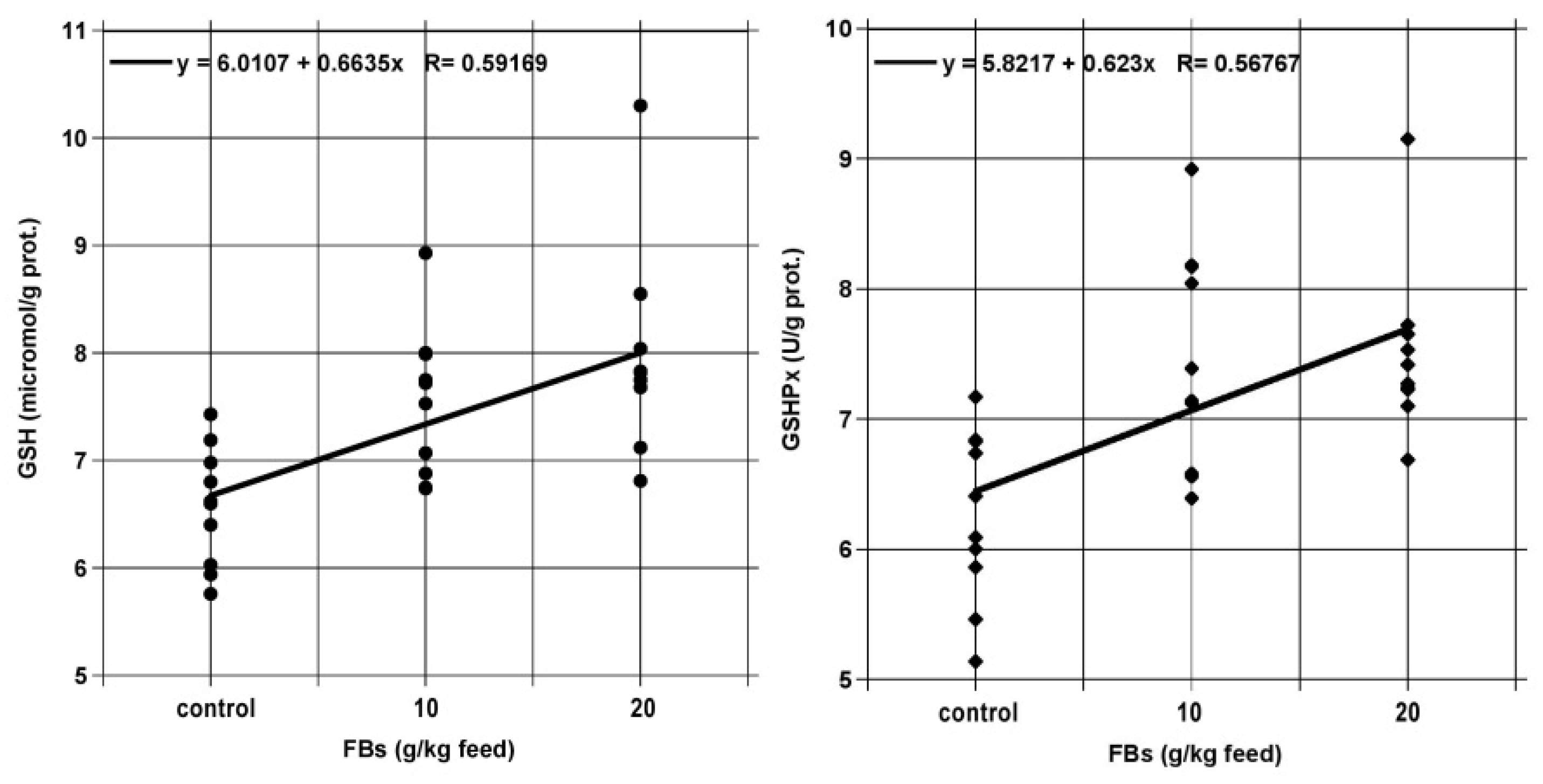
2.5. Testicular Lipid Peroxidation and Antioxidants
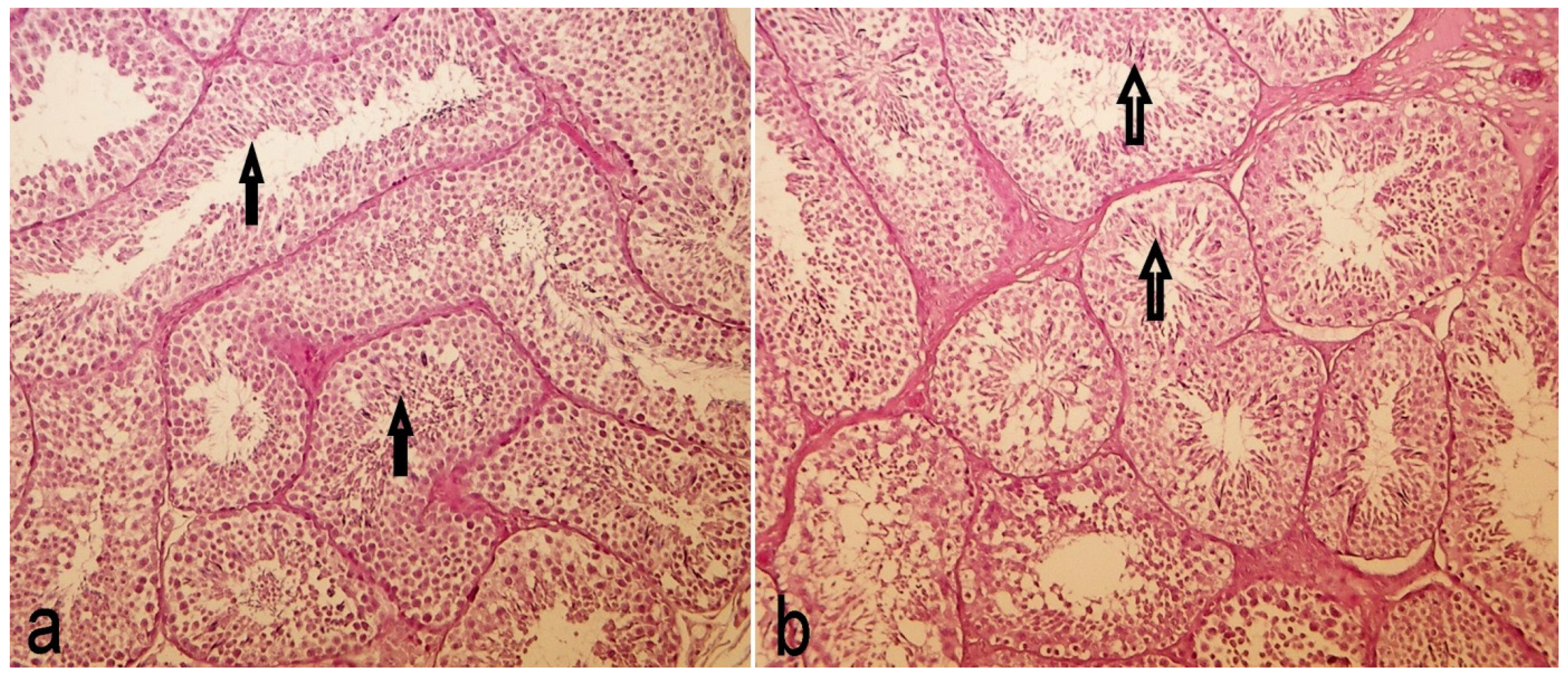
2.6. Testis Histology
2.7. Spermium Membrane Fatty Acid Profile
3. Discussion
3.1. Animal Performance
3.2. Testicular Phospholipid Fatty Acid Composition
3.3. Testicular Lipid Peroxidation and antioxidants
3.4. Testis Histology
3.5. Spermium Phospholipid Fatty Acid Profile
4. Conclusions
5. Materials and Methods
5.1. Animals and keeping
5.2. Feed Mycotoxin Contamination
5.3. Evaluation of Sperm Morphology, Chromatin Integrity, and Viability
5.4. Determination of the Testicular and Spermium Phospholipid Fatty Acid Composition
5.5. Testicular Antioxidant Status and Lipid Peroxidation
5.6. Histopathology
5.7. Statistical Analysis
5.8. Ethical Issues
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rheeder, J.P.; Marasas, W.F.O.; Vismer, H.F. Production of fumonisin analogs by Fusarium species. Appl. Environ. Microbiol. 2002, 68, 2101–2105. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Fan, L.; Yin, S.; Chen, H.; Hu, H. Molecular mechanisms of fumonisin B1-induced toxicities and its applications in the mechanism-based interventions. Toxicon 2019, 167, 1–5. [Google Scholar] [CrossRef]
- Mézes, M.; Balogh, K. Mycotoxins in rabbit feed: A review. World Rabbit Sci. 2009, 17, 53–62. [Google Scholar]
- BIOMIN Holding GmbH, Getzersdorf, Austria. Available online: https://www.biomin.net/science-hub/world-mycotoxin-survey-impact-2020/ (accessed on 15 March 2021).
- Diaz, G.J.; Boermans, H.J. Fumonisin toxicosis in domestic animals: A review. Vet. Hum. Toxicol. 1994, 36, 548–555. [Google Scholar]
- Riley, R.T.; Enongene, E.; Voss, K.A.; Norred, W.P.; Meredith, F.I.; Sharma, R.P.; Spitsbergen, J.; Williams, D.E.; Carlson, D.B.; Merrill, A.H. Sphingolipid perturbations as mechanisms for fumonisin carcinogenesis. Environ. Health Perspect. 2001, 109, 301–308. [Google Scholar] [CrossRef] [Green Version]
- Gelderblom, W.C.A.; Snyman, S.D.; Abel, S.; Lebepe-Mazur, S.; Smuts, C.M.; Van Der Westhuizen, L.; Marasas, W.F.O.; Victor, T.C.; Knasmüller, S.; Huber, W. Hepatotoxicity and -carcinogenicity of the fumonisins in rats: A review regarding mechanistic implications for establishing risk in humans. In Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 1996; Volume 392, pp. 279–296. [Google Scholar]
- Stockmann-Juvala, H.; Mikkola, J.; Naarala, J.; Loikkanen, J.; Elovaara, E.; Savolainen, K. Oxidative stress induced by fumonisin B1 in continuous human and rodent neural cell cultures. Free Radic. Res. 2004, 38, 933–942. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.; Jiang, Y.; Fan, Y.; Ma, Y.; Lei, H.; Su, J. Fumonisin B1 Induces Oxidative Stress and Breaks Barrier Functions in Pig Iliac Endothelium Cells. Toxins 2019, 11, 387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabó, A.; Szabó-Fodor, J.; Fébel, H.; Romvári, R.; Kovács, M. Individual and combined haematotoxic effects of fumonisin B1 and T-2 mycotoxins in rabbits. Food Chem. Toxicol. 2014, 72, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Domijan, A.M.; Abramov, A.Y. Fumonisin B1 inhibits mitochondrial respiration and deregulates calcium homeostasis—Implication to mechanism of cell toxicity. Int. J. Biochem. Cell Biol. 2011, 43, 897–904. [Google Scholar] [CrossRef]
- Minervini, F.; Lacalandra, G.M.; Filannino, A.; Garbetta, A.; Nicassio, M.; Dell’Aquila, M.E.; Visconti, A. Toxic effects induced by mycotoxin fumonisin B1 on equine spermatozoa: Assessment of viability, sperm chromatin structure stability, ROS production and motility. Toxicol. Vitr. 2010, 24, 2072–2078. [Google Scholar] [CrossRef] [PubMed]
- Ewuola, E.O. Organ traits and histopathology of rabbits fed varied levels of dietary fumonisin B1. J. Anim. Physiol. Anim. Nutr. 2009, 93, 726–731. [Google Scholar] [CrossRef]
- Ewuola, E.O.; Egbunike, G.N. Effects of dietary fumonisin B1 on the onset of puberty, semen quality, fertility rates and testicular morphology in male rabbits. Reproduction 2010, 139, 439–445. [Google Scholar] [CrossRef] [Green Version]
- Ewuola, E.O.; Egbunike, G.N. Gonadal and extra-gonadal sperm reserves and sperm production of pubertal rabbits fed dietary fumonisin B1. Anim. Reprod. Sci. 2010, 119, 282–286. [Google Scholar] [CrossRef] [PubMed]
- Foote, R.H.; Carney, E.W. The rabbit as a model for reproductive and developmental toxicity studies. Reprod. Toxicol. 2000, 14, 477–493. [Google Scholar] [CrossRef]
- Knutsen, H.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Edler, L.; Grasl-Kraupp, B.; et al. Risks for animal health related to the presence of fumonisins, their modified forms and hidden forms in feed. EFSA J. 2018, 16, 5242. [Google Scholar]
- Szabó, A.; Szabó-Fodor, J.; Fébel, H.; Mézes, M.; Bajzik, G.; Kovács, M. Oral administration of fumonisin B1 and T-2 individually and in combination affects hepatic total and mitochondrial membrane lipid profile of rabbits. Physiol. Int. 2016, 103, 321–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bondy, G.S.; Suzuki, C.A.M.; Mueller, R.W.; Fernie, S.M.; Armstrong, C.L.; Hierlihy, S.L.; Savard, M.E.; Barker, M.G. Gavage administration of the fungal toxin fumonisin B1 to female sprague-dawley rats. J. Toxicol. Environ. Health Part A 1998, 53, 135–151. [Google Scholar] [CrossRef]
- Gumprecht, L.A.; Marcucci, A.; Weigel, R.M.; Vesonder, R.F.; Riley, R.T.; Showker, J.L.; Beasley, V.R.; Haschek, W.M. Effects of intravenous fumonisin B1 in rabbits: Nephrotoxicityxs and sphingolipid alterations. Nat. Toxins 1995, 3, 395–403. [Google Scholar] [CrossRef]
- Hulbert, A.J. Life, death and membrane bilayers. J. Exp. Biol. 2003, 206, 2303–2311. [Google Scholar] [CrossRef] [Green Version]
- Morin, R.J. In vitro Incorporation of Acetate-l-C14 into Sphingomyelin, Phosphatidyl Choline and Phosphatidyl Ethanolamine of Rabbit Testes. Exp. Biol. Med. 1967, 126, 229–232. [Google Scholar] [CrossRef]
- Retterstøl, K.; Tran, T.N.; Haugen, T.B.; Christophersen, B.O. Metabolism of very long chain polyunsaturated fatty acids in isolated rat germ cells. Lipids 2001, 36, 601–606. [Google Scholar] [CrossRef] [PubMed]
- Xue, S.; He, Z.; Lu, J.; Tao, X.; Zheng, L.; Xie, Y.; Xiao, X.; Peng, R.; Li, H. Effect of growth on fatty acid composition of total intramuscular lipid and phospholipids in Ira rabbits. Korean J. Food Sci. Anim. Resour. 2015, 35, 10–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabó, A.; Szabó-Fodor, J.; Fébel, H.; Mézes, M.; Repa, I.; Kovács, M. Acute hepatic effects of low-dose fumonisin B1 in rats. Acta Vet. Hung. 2016, 64, 436–448. [Google Scholar] [CrossRef] [Green Version]
- Marzouki, Z.M.H.; Coniglio, J.G. Effect of essential fatty acid deficiency on lipids of rat sertoli and germinal cells. Biol. Reprod. 1982, 27, 312–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosny, N.S.; Hashem, N.M.; Morsy, A.S.; Abo-elezz, Z.R. Effects of Organic Selenium on the Physiological Response, Blood Metabolites, Redox Status, Semen Quality, and Fertility of Rabbit Bucks Kept Under Natural Heat Stress Conditions. Front. Vet. Sci. 2020, 7. [Google Scholar] [CrossRef]
- Aydilek, N.; Aksakal, M.; Karakilçik, A.Z. Effects of testosterone and vitamin E on the antioxidant system in rabbit testis. Andrologia 2004, 36, 277–281. [Google Scholar] [CrossRef]
- Ali, O.; Szabó-Fodor, J.; Fébel, H.; Mézes, M.; Balogh, K.; Glávits, R.; Kovács, M.; Zantomasi, A.; Szabó, A. Porcine hepatic response to fumonisin b1 in a short exposure period: Fatty acid profile and clinical investigations. Toxins 2019, 11, 655. [Google Scholar] [CrossRef] [Green Version]
- Sousa, M.C.S.; Galli, G.M.; Bottari, N.B.; Alba, D.F.; Leal, K.W.; Lopes, T.F.; Druzian, L.; Schetinger, M.R.C.; Gloria, E.M.; Mendes, R.E.; et al. Fumonisin-(Fusarium verticillioides)-contaminated feed causes hepatic oxidative stress and negatively affects broiler performance in the early stage: Does supplementation with açai flour residues (Euterpe oleracea) minimize these problems? Microb. Pathog. 2020, 146. [Google Scholar] [CrossRef]
- Zhang, X.; Lv, F.; Tang, J. Protection from ischemia by preconditioning, postconditioning, and combined treatment in rabbit testicular ischemia reperfusion injury. Arch. Biochem. Biophys. 2016, 608, 1–7. [Google Scholar] [CrossRef]
- Sheik Abdul, N.; Marnewick, J.L. Fumonisin B1-induced mitochondrial toxicity and hepatoprotective potential of rooibos: An update. J. Appl. Toxicol. 2020, 40, 1602–1613. [Google Scholar] [CrossRef]
- Wang, X.; Wu, Q.; Wan, D.; Liu, Q.; Chen, D.; Liu, Z.; Martinez-Larrañaga, M.R.; Martínez, M.A.; Anadón, A.; Yuan, Z. Fumonisins: Oxidative stress-mediated toxicity and metabolism in vivo and in vitro. Arch. Toxicol. 2015, 90, 81–101. [Google Scholar] [CrossRef] [PubMed]
- Yayeh, T.; Jeong, H.R.; Park, Y.S.; Moon, S.; Sur, B.; Yoo, H.-S.; Oh, S. Fumonisin B1-Induced Toxicity Was Not Exacerbated in Glutathione Peroxidase-1/Catalase Double Knock Out Mice. Biomol. Ther. 2020. [Google Scholar] [CrossRef]
- Castellini, C.; Lattaioli, P.; Dal Bosco, A.; Minelli, A.; Mugnai, C. Oxidative status and semen characteristics of rabbit buck as affected by dietary vitamin E, C and n-3 fatty acids. Reprod. Nutr. Dev. 2003, 43, 91–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahluwalia, B.; Holman, R.T. Fatty acid composition of lipids of bull, boar, rabbit and human semen. J. Reprod. Fertil. 1969, 18, 431–437. [Google Scholar] [CrossRef] [Green Version]
- Fodor, J.; Kametier, L.; Kovács, M. Practical aspects of fumonisin production under laboratory conditions. Mycotoxin Res. 2006, 22, 211–216. [Google Scholar] [CrossRef]
- Bartók, T.; Tölgyesi, L.; Szekeres, A.; Varga, M.; Bartha, R.; Szécsi, Á.; Bartók, M.; Mesterházy, Á. Detection and characterization of twenty-eight isomers of fumonisin B1 (FB1) mycotoxin in a solid rice culture infected with Fusarium verticillioides by reversed-phase high-performance liquid chromatography/electrospray ionization time-of-flight and ion trap mass spectrometry. Rapid Commun. Mass Spectrom. 2010, 24, 35–42. [Google Scholar] [CrossRef]
- Barth, A.D.; Oko, R.J. Abnormal Morphology of Bovine Spermatozoa; Ames, Iowa State University Press: Ames, IA, USA, 1989. [Google Scholar]
- Nagy, S.; Jansen, J.; Topper, E.K.; Gadella, B.M. A triple-stain flow cytometric method to assess plasma- and acrosome-membrane integrity of cryopreserved bovine sperm immediately after thawing in presence of egg-yolk particles. Biol. Reprod. 2003, 68, 1828–1835. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Leray, C.; Pelletier, X.; Hemmendinger, S.; Cazenave, J.P. Thin-layer chromatography of human platelet phospholipids with fatty acid analysis. J. Chromatogr. B Biomed. Sci. Appl. 1987, 420, 411–416. [Google Scholar] [CrossRef]
- Christie, W.W. A simple procedure for rapid transmethylation of glycerolipids and cholesteryl esters. J. Lipid Res. 1982, 23, 1072–1075. [Google Scholar] [CrossRef]
- Botsoglou, N.A.; Fletouris, D.J.; Papageorgiou, G.E.; Vassilopoulos, V.N.; Mantis, A.J.; Trakatellis, A.G. Rapid, Sensitive, and Specific Thiobarbituric Acid Method for Measuring Lipid Peroxidation in Animal Tissue, Food, and Feedstuff Samples. J. Agric. Food Chem. 1994, 42, 1931–1937. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis (28.054), 14th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 1984. [Google Scholar]
- Sedlak, J.; Lindsay, R.H. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman’s reagent. Anal. Biochem. 1968, 25, 192–205. [Google Scholar] [CrossRef]
- Lawrence, R.A.; Burk, R.F. Species, tissue and subcellular distribution of non Se-dependent glutathione peroxidase activity. J. Nutr. 1978, 108, 211–215. [Google Scholar] [CrossRef]
- OECD Environmental Health and Safety Publications. OECD Principles of Good Laboratory Practice (as Revised in 1997). Available online: https://ntp.niehs.nih.gov/iccvam/suppdocs/feddocs/oecd/oecd_glpcm.pdf (accessed on 12 October 2018).
- SPSS. SPSS for Windows ver. 20; SPSS: Chicago, IL, USA, 2012. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Control | 10 mg/kg | 20 mg/kg | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Organ Weight (g) | Mean | ± | SD | Mean | ± | SD | Mean | ± | SD | |||
| liver | 91.7 | ± | 17.6 | ab | 96.9 | ± | 13.2 | b | 80.9 | ± | 11.6 | a |
| kidney | 19.8 | ± | 1.92 | ab | 20.8 | ± | 1.95 | b | 18.4 | ± | 1.77 | a |
| spleen | 1.80 | ± | 0.41 | 1.58 | ± | 0.47 | 1.62 | ± | 0.30 | |||
| testes | 10.3 | ± | 1.68 | 10.5 | ± | 1.05 | 9.6 | ± | 1.58 | |||
| rel. liver (%) | 1.90 | ± | 0.26 | ab | 2.07 | ± | 0.29 | b | 1.75 | ± | 0.17 | a |
| rel. kidney (%) | 0.41 | ± | 0.04 | 0.44 | ± | 0.04 | 0.40 | ± | 0.04 | |||
| rel. spleen (%) | 0.04 | ± | 0.01 | 0.03 | ± | 0.01 | 0.04 | ± | 0.01 | |||
| rel. testes (%) | 0.22 | ± | 0.04 | 0.22 | ± | 0.02 | 0.21 | ± | 0.03 | |||
| initial BW (g) | 4380 | ± | 345 | 4387 | ± | 335 | 4390 | ± | 325 | |||
| final BW (g) | 4819 | ± | 350 | 4686 | ± | 309 | 4619 | ± | 486 | |||
| Group | Control | 10 mg/kg | 20 mg/kg | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fatty Acid | Mean | SD | Mean | SD | Mean | SD | ||||||
| C14:0 | 0.39 | ± | 0.52 | 0.31 | ± | 0.15 | 0.26 | ± | 0.12 | |||
| C14:1n5 | 0.02 | ± | 0.00 | nd | nd | |||||||
| C15:0 | 0.31 | ± | 0.64 | 0.10 | ± | 0.01 | 0.11 | ± | 0.02 | |||
| C16:0 | 35.3 | ± | 3.01 | 37.0 | ± | 2.32 | 35.3 | ± | 1.39 | |||
| C16:1n7 | 0.20 | ± | 0.07 | 0.23 | ± | 0.05 | 0.21 | ± | 0.03 | |||
| C17:0 | 0.32 | ± | 0.08 | a | 0.40 | ± | 0.09 | a | 0.51 | ± | 0.12 | b |
| C18:0 | 29.0 | ± | 8.17 | 25.0 | ± | 4.82 | 24.6 | ± | 3.16 | |||
| C18:1n9c | 7.95 | ± | 2.45 | 8.79 | ± | 1.38 | 9.04 | ± | 0.85 | |||
| C18:1n7 | 0.83 | ± | 0.25 | 0.96 | ± | 0.15 | 0.88 | ± | 0.12 | |||
| C18:2n6 | 4.91 | ± | 1.24 | 5.62 | ± | 0.99 | 5.47 | ± | 0.76 | |||
| C18:3n6 | 0.12 | ± | 0.05 | 0.15 | ± | 0.02 | 0.15 | ± | 0.04 | |||
| C18:3n3 | 0.02 | ± | 0.01 | 0.08 | ± | 0.18 | 0.02 | ± | 0.00 | |||
| C18:4n3 | 0.43 | ± | 0.45 | 0.27 | ± | 0.27 | 0.30 | ± | 0.14 | |||
| C20:0 | 0.21 | ± | 0.12 | 0.14 | ± | 0.05 | 0.14 | ± | 0.03 | |||
| C20:1n9 | 0.13 | ± | 0.04 | 0.13 | ± | 0.03 | 0.11 | ± | 0.02 | |||
| C20:2n6 | 0.30 | ± | 0.08 | 0.35 | ± | 0.08 | 0.30 | ± | 0.03 | |||
| C20:3n9 (Mead acid) | 0.09 | ± | 0.03 | 0.08 | ± | 0.01 | 0.08 | ± | 0.02 | |||
| C20:3n6 | 7.58 | ± | 2.09 | 8.68 | ± | 1.78 | 8.83 | ± | 1.19 | |||
| C20:4n6 | 11.6 | ± | 3.27 | 11.5 | ± | 2.30 | 13.3 | ± | 1.48 | |||
| C20:5n3 | 0.10 | ± | 0.03 | 0.09 | ± | 0.01 | nd | |||||
| C22:0 | 0.06 | ± | 0.06 | 0.03 | ± | 0.01 | 0.03 | ± | 0.01 | |||
| C22:5n3 | 0.18 | ± | 0.07 | 0.15 | ± | 0.06 | 0.18 | ± | 0.04 | |||
| C24:0 | 0.13 | ± | 0.10 | nd | nd | |||||||
| C22:6n3 | 0.12 | ± | 0.05 | 0.11 | ± | 0.03 | 0.13 | ± | 0.06 | |||
| Saturated | 65.6 | ± | 9.27 | 62.9 | ± | 6.04 | 61.0 | ± | 3.73 | |||
| Unsaturated | 34.4 | ± | 9.27 | 37.1 | ± | 6.04 | 39.0 | ± | 3.73 | |||
| Monounsaturated | 9.09 | ± | 2.80 | 10.1 | ± | 1.55 | 10.2 | ± | 0.97 | |||
| Polyunsaturated | 25.3 | ± | 6.60 | 26.9 | ± | 4.59 | 28.8 | ± | 2.83 | |||
| n3 | 0.77 | ± | 0.48 | 0.58 | ± | 0.33 | 0.62 | ± | 0.14 | |||
| n6 | 24.5 | ± | 6.36 | 26.3 | ± | 4.47 | 28.1 | ± | 2.79 | |||
| n6/n3 | 39.5 | ± | 18.5 | 64.5 | ± | 43.0 | 46.7 | ± | 9.80 | |||
| Odd chain FA | 0.61 | ± | 0.59 | 0.49 | ± | 0.10 | 0.61 | ± | 0.13 | |||
| Unsaturation index | 93.6 | ± | 24.9 | 98.1 | ± | 16.7 | 106.1 | ± | 10.5 | |||
| Average FA chain length | 17.68 | ± | 0.13 | 17.67 | ± | 0.11 | 17.74 | ± | 0.07 | |||
| Group | Control | 10 mg/kg | 20 mg/kg | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GSH (micromol/g prot.) | 6.57 | ± | 0.55 | a | 7.54 | ± | 0.70 | b | 7.90 | ± | 0.98 | b |
| GSHPx (U/g prot.) | 6.25 | ± | 0.66 | a | 7.45 | ± | 0.85 | b | 7.50 | ± | 0.65 | b |
| MDA (nmol/g) | 44.7 | ± | 9.11 | 39.0 | ± | 7.17 | 38.5 | ± | 7.22 | |||
| CD (A232 nm) | 0.34 | ± | 0.02 | b | 0.34 | ± | 0.03 | b | 0.31 | ± | 0.02 | a |
| CT (A268 nm) | 0.16 | ± | 0.01 | b | 0.15 | ± | 0.01 | ab | 0.14 | ± | 0.01 | a |
| Group | Control | 10 mg/kg | 20 mg/kg | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | ||||
| C14:0 | 2.61 | ± | 0.50 | 2.32 | ± | 0.57 | 2.57 | ± | 0.92 |
| C15:0 | 0.19 | ± | 0.07 | 0.49 | ± | 0.34 | 0.46 | ± | 0.35 |
| C16:0 | 25.0 | ± | 0.28 | 26.3 | ± | 1.05 | 26.4 | ± | 0.98 |
| C16:1 | 0.68 | ± | 0.37 | 0.80 | ± | 0.12 | 0.78 | ± | 0.61 |
| C17:0 | 0.74 | ± | 0.49 | 1.13 | ± | 0.61 | 1.04 | ± | 0.12 |
| C17:1n7 | 0.18 | ± | 0.03 | 0.24 | ± | 0.12 | 0.28 | ± | 0.14 |
| C18:0 | 47.5 | ± | 3.76 | 39.0 | ± | 9.26 | 41.1 | ± | 6.85 |
| C18:1n9 | 9.62 | ± | 3.41 | 14.9 | ± | 6.85 | 13.3 | ± | 6.96 |
| C18:1n7 | 0.61 | ± | 0.06 | 0.82 | ± | 0.33 | 0.97 | ± | 0.44 |
| C18:2n6 | 5.01 | ± | 1.98 | 8.54 | ± | 3.90 | 7.16 | ± | 2.45 |
| C18:3n6 | 0.02 | ± | 0.01 | 0.09 | ± | 0.06 | 0.06 | ± | 0.03 |
| C18:3n3 | 0.10 | ± | 0.01 | 0.08 | ± | 0.07 | 0.05 | ± | 0.02 |
| C20:0 | n.d. | 0.15 | ± | 0.03 | 0.17 | ± | 0.15 | ||
| C20:1n9 | 0.08 | ± | 0.02 | 0.08 | ± | 0.05 | 0.02 | ± | 0.01 |
| C20:3n9 (Mead acid) | 0.31 | ± | 0.05 | 0.26 | ± | 0.15 | 0.28 | ± | 0.19 |
| C20:3n6 | 3.18 | ± | 0.80 | 2.21 | ± | 1.14 | 2.64 | ± | 1.28 |
| C20:4n6 | 3.11 | ± | 0.93 | 1.80 | ± | 0.91 | 1.93 | ± | 1.05 |
| C20:5n3 | 0.28 | ± | 0.01 | 0.21 | ± | 0.07 | 0.17 | ± | 0.05 |
| C22:0 | 0.11 | ± | 0.01 | n.d. | 0.05 | ± | 0.04 | ||
| C22:1n9 | 0.08 | ± | 0.01 | n.d. | 0.07 | ± | 0.01 | ||
| C24:0 | 0.41 | ± | 0.01 | 0.11 | ± | 0.06 | n.d. | ||
| C22:6n3 | 0.37 | ± | 0.02 | 0.29 | ± | 0.18 | 0.34 | ± | 0.11 |
| C24:1n9 | 0.19 | ± | 0.16 | 0.19 | ± | 0.07 | 0.14 | ± | 0.13 |
| Saturated | 76.4 | ± | 4.17 | 69.6 | ± | 8.99 | 71.9 | ± | 8.23 |
| Unsaturated | 23.6 | ± | 4.17 | 30.4 | ± | 8.99 | 28.1 | ± | 8.23 |
| Monounsaturated | 11.4 | ± | 4.00 | 17.0 | ± | 7.34 | 15.5 | ± | 8.30 |
| Polyunsaturated | 12.2 | ± | 0.17 | 13.4 | ± | 1.88 | 12.6 | ± | 0.07 |
| n3 | 0.56 | ± | 0.14 | 0.51 | ± | 0.24 | 0.56 | ± | 0.04 |
| n6 | 11.3 | ± | 0.26 | 12.6 | ± | 2.18 | 11.8 | ± | 0.15 |
| n6/n3 | 20.8 | ± | 5.59 | 37.4 | ± | 36.4 | 21.1 | ± | 1.62 |
| Odd chain FA | 0.93 | ± | 0.56 | 1.62 | ± | 0.93 | 1.50 | ± | 0.23 |
| Unsaturation index | 47.4 | ± | 1.15 | 51.7 | ± | 7.28 | 49.5 | ± | 4.37 |
| Average FA chain length | 17.54 | ± | 0.01 | 17.45 | ± | 0.04 | 17.45 | ± | 0.01 |
| Chemical Composition | Fatty Acid (Diet) | Weight % of Total FAME | |
|---|---|---|---|
| Dry material (%) | 89.0 | C12:0 | 0.05 |
| Crude protein (%) | 14.5 | C14:0 | 0.18 |
| Ether extract (%) | 2.4 | C15:0 | 0.14 |
| Crude fibre (%) | 17.1 | C16:0 | 14.3 |
| Ash (%) | 7.5 | C16:1n7 | 0.20 |
| Lysine (%) | 0.90 | C17:0 | 0.12 |
| Methionine (%) | 0.41 | C18:0 | 2.73 |
| Calcium (%) | 0.88 | C18:1n9 | 36.8 |
| Phosphorus (%) | 0.52 | C18:1n7 | 0.84 |
| Sodium (%) | 0.19 | C18:2n6 | 38.6 |
| Vitamin A (IU/kg) | 14000 | C18:3n3 | 3.83 |
| Vitamin D3 (IU/kg) | 1300 | C20:0 | 0.42 |
| Vitamin E (mg/kg) | 107 | C20:1n9 | 0.50 |
| Digestible energy (MJ/kg) | 9.7 | C20:2n6 | 0.04 |
| C21:0 | 0.04 | ||
| C20:4n6 | 0.07 | ||
| C22:0 | 0.61 | ||
| C24:0 | 0.45 | ||
| C22:6n3 | 0.08 | ||
| Saturated | 19.0 | ||
| Unsaturated | 80.9 | ||
| Monounsaturated | 38.3 | ||
| Polyunsaturated | 42.6 | ||
| n3 | 3.91 | ||
| n6 | 38.7 | ||
| n6/n3 | 9.89 | ||
| Odd chain FA | 0.26 | ||
| Unsaturation index | 127.8 | ||
| Average FA chain length | 17.75 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szabó, A.; Nagy, S.; Ali, O.; Gerencsér, Z.; Mézes, M.; Balogh, K.M.; Bartók, T.; Horváth, L.; Mouhanna, A.; Kovács, M. A 65-Day Fumonisin B Exposure at High Dietary Levels Has Negligible Effects on the Testicular and Spermatological Parameters of Adult Rabbit Bucks. Toxins 2021, 13, 237. https://doi.org/10.3390/toxins13040237
Szabó A, Nagy S, Ali O, Gerencsér Z, Mézes M, Balogh KM, Bartók T, Horváth L, Mouhanna A, Kovács M. A 65-Day Fumonisin B Exposure at High Dietary Levels Has Negligible Effects on the Testicular and Spermatological Parameters of Adult Rabbit Bucks. Toxins. 2021; 13(4):237. https://doi.org/10.3390/toxins13040237
Chicago/Turabian StyleSzabó, András, Szabolcs Nagy, Omeralfaroug Ali, Zsolt Gerencsér, Miklós Mézes, Krisztián Milán Balogh, Tibor Bartók, Levente Horváth, Aziz Mouhanna, and Melinda Kovács. 2021. "A 65-Day Fumonisin B Exposure at High Dietary Levels Has Negligible Effects on the Testicular and Spermatological Parameters of Adult Rabbit Bucks" Toxins 13, no. 4: 237. https://doi.org/10.3390/toxins13040237
APA StyleSzabó, A., Nagy, S., Ali, O., Gerencsér, Z., Mézes, M., Balogh, K. M., Bartók, T., Horváth, L., Mouhanna, A., & Kovács, M. (2021). A 65-Day Fumonisin B Exposure at High Dietary Levels Has Negligible Effects on the Testicular and Spermatological Parameters of Adult Rabbit Bucks. Toxins, 13(4), 237. https://doi.org/10.3390/toxins13040237