Phytotoxic Secondary Metabolites from Fungi



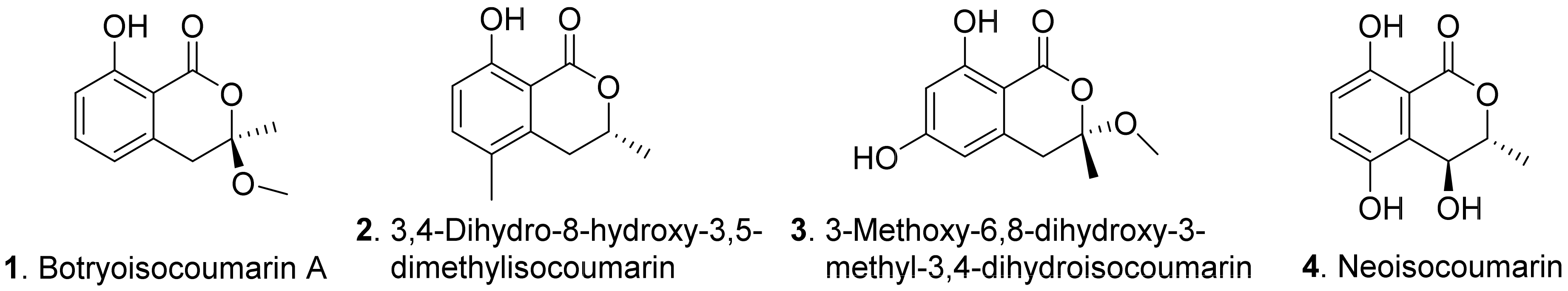{kind=link}
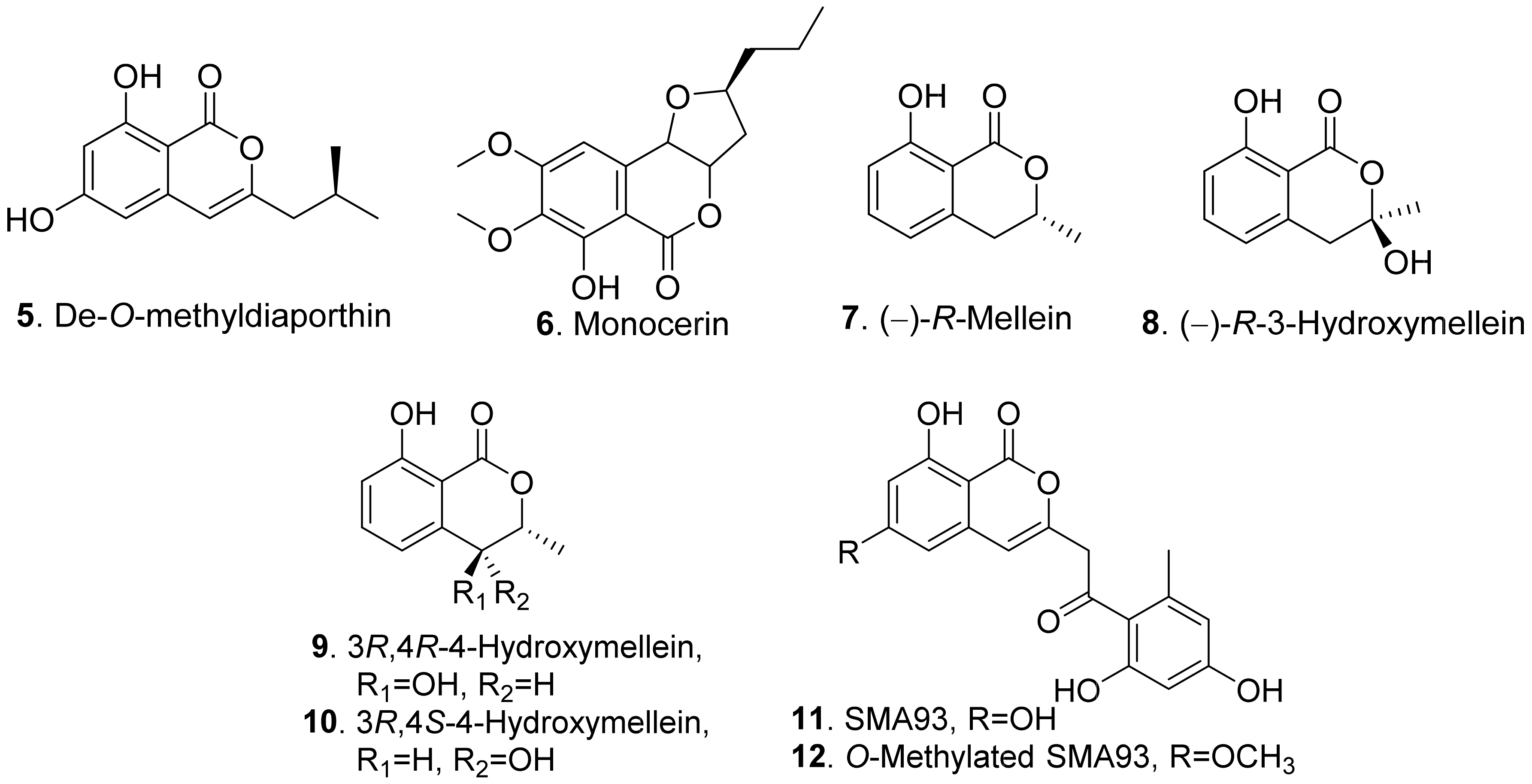{kind=link}
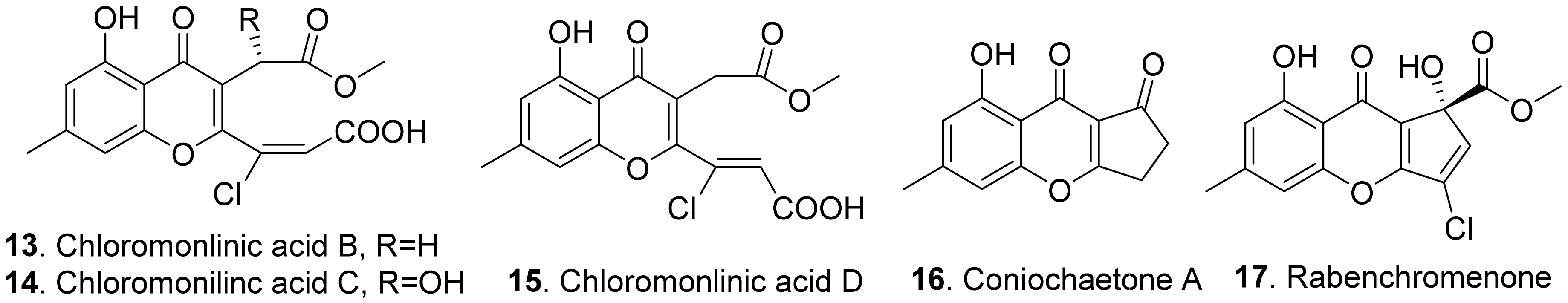{kind=link}
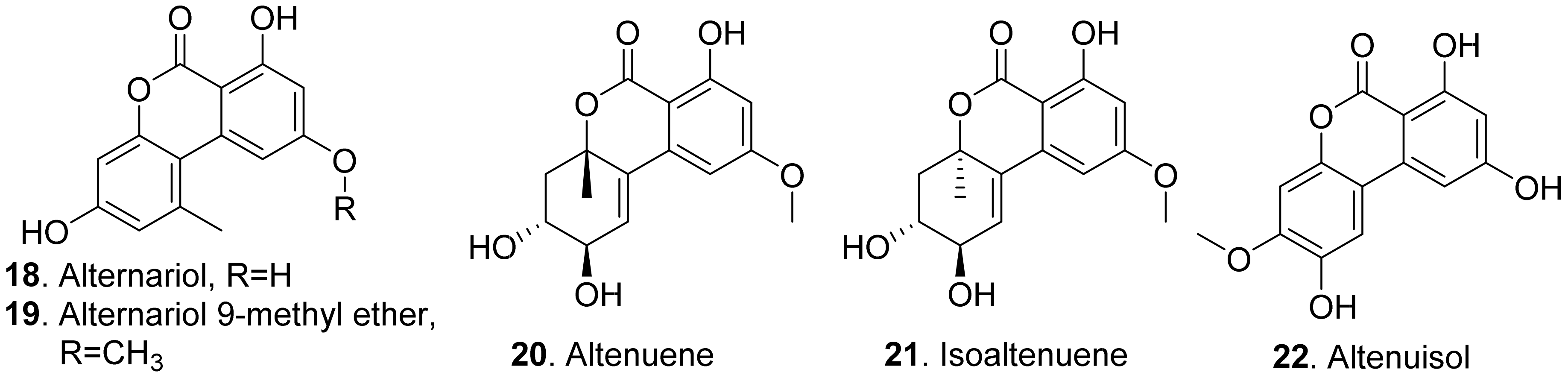{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
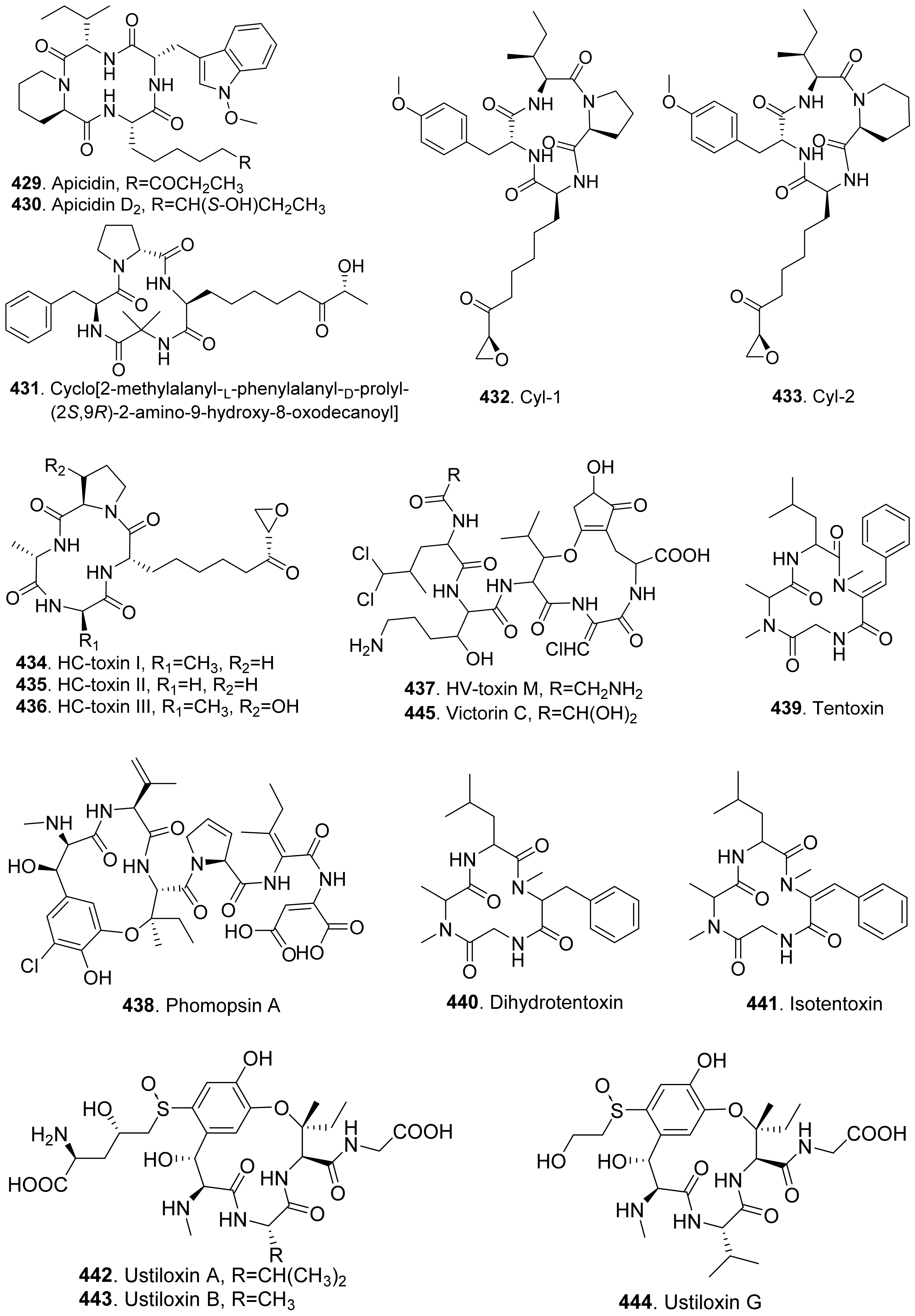{kind=link}
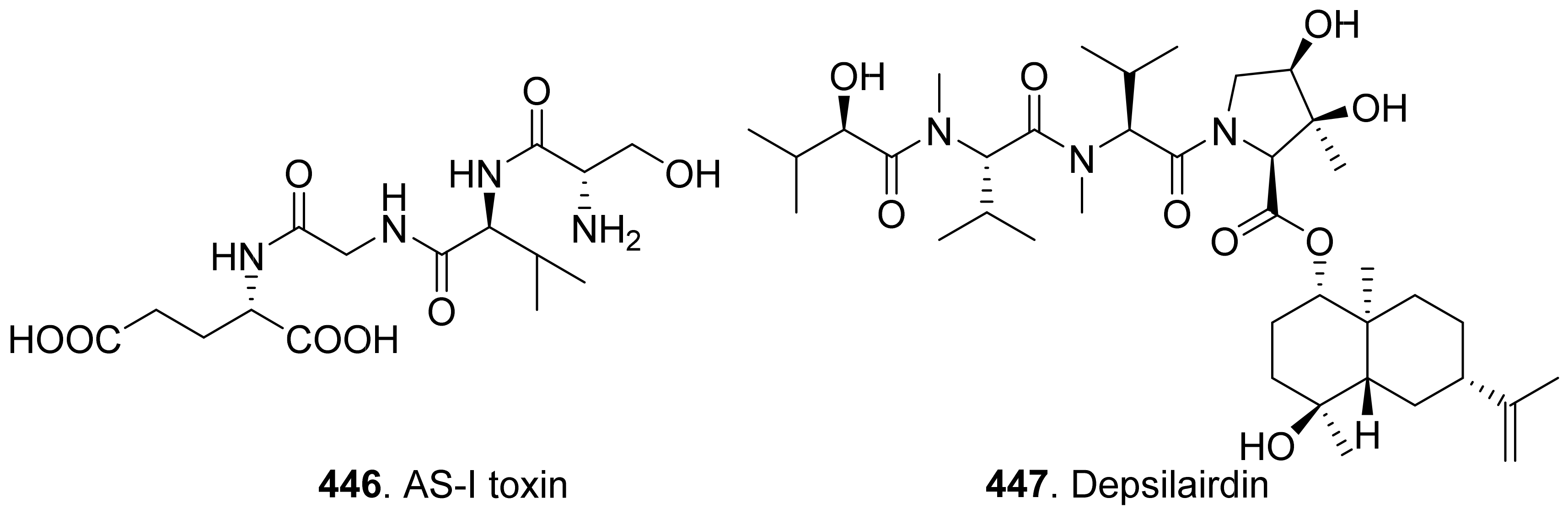{kind=link}
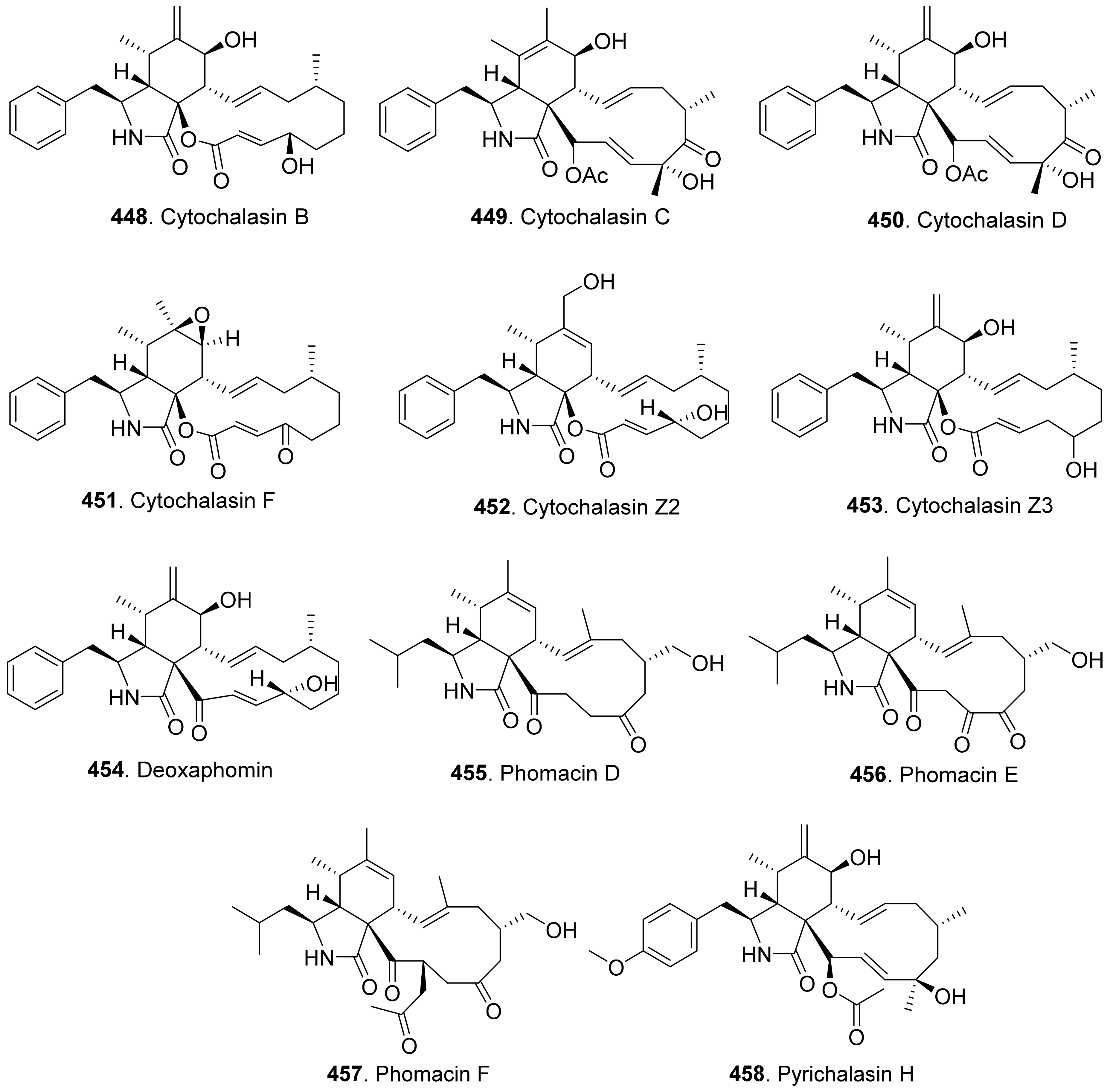{kind=link}
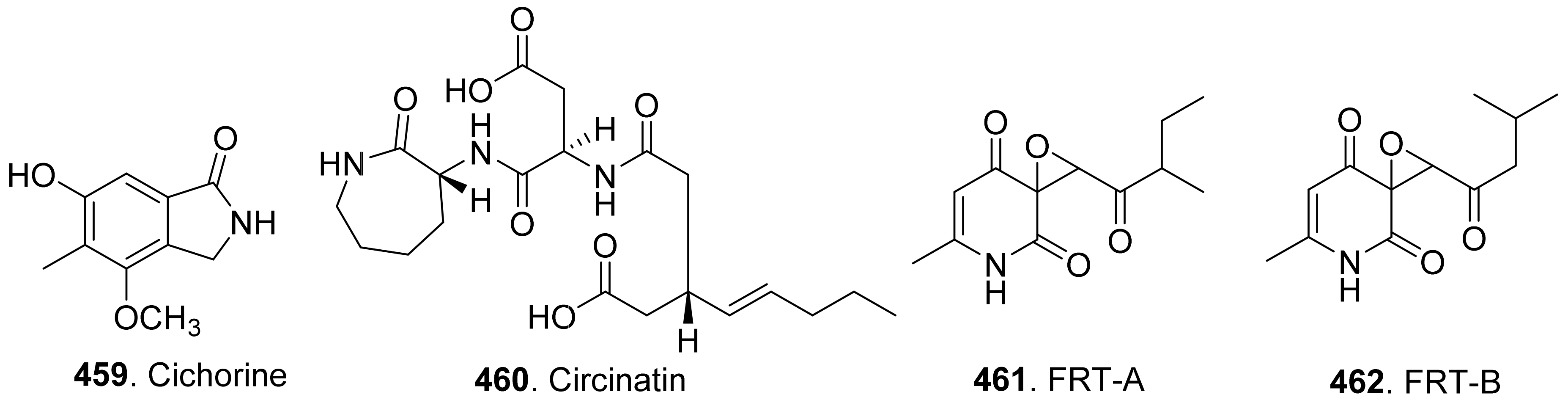{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Polyketides
2.1. Aromatic Polyketides
2.1.1. Benzopyrones
2.1.2. Dibenzopyrones
2.1.3. Benzophenones
2.1.4. Naphthopyrones
2.1.5. Azaphilones
2.1.6. Naphthalene Derivatives
2.1.7. Anthraquinones
2.1.8. Perylenequinonoids
2.1.9. Aromatic Macrolides
2.2. Aliphatic Polyketides
2.2.1. Simple Furan and Furanone Analogues
2.2.2. Aromatic-Free Pyrones
2.2.3. Furopyran and Pyranopyran Analogues
2.2.4. Macrolide Analogues
2.2.5. Sorbicillinoids
2.2.6. Linear Polyketides
3. Phenols and Phenolic Acids
4. Terpenoids
4.1. Monoterpenoids
4.2. Sesquiterpenoids
4.3. Diterpenoids
4.4. Sesterterpenoids
4.5. Triterpenoids
4.6. Meroterpenoids
4.6.1. Meroterpenoids Containing Monoterpene Biosynthetic Pathways
4.6.2. Meroterpenoids Containing Sesquiterpene Biosynthetic Pathways
4.6.3. Meroterpenoids Containing Diterpene Biosynthetic Pathways
5. Nitrogen-Containing Metabolites
5.1. Cyclic Peptides
5.1.1. Cyclic Depsipeptides
5.1.2. Cyclic Peptides without Ester Bond
5.2. Noncyclic Oligopeptides
5.3. Cytochalasin Congeners
5.4. Lactams
5.5. Indole Derivatives
5.6. Pyridine Derivatives
5.7. Amines and Noncyclic Amides
5.8. Other Nitrogen-Containing Metabolites
6. Miscellaneous
7. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Evidente, A.; Cimino, A.; Masi, M. Phytotoxins produced by pathogenic fungi of agrarian plants. Phytochem. Rev. 2019, 18, 843–870. [Google Scholar] [CrossRef]
- Chen, H.; Singh, H.; Bhardwaj, N.; Bhardwaj, S.K.; Khatri, M.; Kim, K.-H.; Peng, W. An exploration on the toxicity mechanisms of phytotoxins and their potential utilities. Cret. Rev. Environ. Sci. Technol. 2020. [Google Scholar] [CrossRef]
- Pontes, J.G.M.; Fernandes, L.S.; Dos Santos, R.V.; Tasic, L.; Fill, T.P. Virulence factors in the phytopathogen-host interactions: An overview. J. Agric. Food Chem. 2020, 68, 7555–7570. [Google Scholar] [CrossRef]
- Meena, M.; Samal, S. Alternaria host-specific (HSTs) toxins: An overview of chemical characterization, target sites, regulation and their toxic effects. Toxicol. Rep. 2019, 6, 745–758. [Google Scholar] [CrossRef]
- Walton, J.D.; Panaccione, D.G. Host-selective toxins and disease specificity: Perspectives and progress. Annu. Rev. Phytopathol. 1993, 31, 275–303. [Google Scholar] [CrossRef] [PubMed]
- Tsuge, T.; Harimoto, Y.; Akimitsu, K.; Ohtani, K.; Kodama, M.; Akagi, Y.; Egusa, M.; Yamamoto, M.; Otani, H. Host-selective toxins produced by the plant pathogenic fungus Alternaria alternata. FEMS Microbiol. Rev. 2013, 37, 44–66. [Google Scholar] [CrossRef] [PubMed]
- Wolpert, T.J.; Dunkle, L.D.; Ciuffetti, L.M. Hostselective toxins and avirulence determinants: What’s in a name? Annu. Rev. Phytopathol. 2002, 40, 251–285. [Google Scholar] [CrossRef] [Green Version]
- Strange, R.N. Phytotoxins produced by microbial plant pathogens. Nat. Prod. Rep. 2007, 24, 127–144. [Google Scholar] [CrossRef] [PubMed]
- Strobel, G.A. Phytotoxins. Annu. Rev. Biochem. 1982, 51, 309–333. [Google Scholar] [CrossRef] [PubMed]
- Ballio, A.; Graniti, A. Phytotoxins and their involvement in plant disease. Experientia 1991, 47, 751–864. [Google Scholar]
- Cimmino, A.; Masi, M.; Evidente, M.; Superchi, S.; Evidente, A. Fungal phytotoxins with potential herbicidal activity: Chemical and biological characterization. Nat. Prod. Rep. 2015, 32, 1629–1653. [Google Scholar] [CrossRef] [PubMed]
- Mobius, N.; Hertweck, C. Fungal phytotoxins as mediators of virulence. Curr. Opin. Plant Biol. 2009, 12, 390–398. [Google Scholar] [CrossRef]
- Varejao, E.V.V.; Demuner, A.J.; Barbosa, L.C.A.; Barreto, R.W. The search for new natural herbicides—Strategic approaches for discovering fungal phytotoxins. Crop Prot. 2013, 48, 41–50. [Google Scholar] [CrossRef]
- McLean, M. The phytotoxicity of Fusarium metabolites: An update since 1989. Mycopathologia 1996, 133, 163–179. [Google Scholar] [CrossRef] [PubMed]
- Lou, J.; Fu, L.; Peng, Y.; Zhou, L. Metabolites from Alternaria fungi and their bioactivities. Molecules 2013, 18, 5891–5935. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Chen, W. Phytotoxic metabolites produced by legume-associated Ascochyta and its related genera in the Dothidemycetes. Toxins 2019, 11, 627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Kistler, H.C.; Ma, Z. Fusarium graminearum trichothecene mycotoxins: Biosynthesis, regulation and management. Annu. Rev. Phytopathol. 2019, 57, 15–39. [Google Scholar] [CrossRef] [Green Version]
- Masi, M.; Cimmino, A.; Reveglia, P.; Mugnai, L.; Surico, G.; Evidente, A. Advances on fungal phytotoxins and their role in grapevine trunk diseases. J. Agric. Food Chem. 2018, 66, 5948–5958. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Shan, T.; Mou, Y.; Zhou, L. Plant-derived bioactive compounds produced by endophytic fungi. Mini Rev. Med. Chem. 2011, 11, 159–168. [Google Scholar] [CrossRef]
- Piyasena, K.G.N.P.; Wickramarachchi, W.A.R.T.; Kumar, N.S.; Jayasinghe, L.; Fujimoto, Y. Two phytotoxic azaphilone derivatives from Chaetomium globosum, a fungal endophyte isolated from Amaranthus viridis leaves. Mycology 2015, 6, 158–160. [Google Scholar] [CrossRef] [Green Version]
- Ma, K.-L.; Wei, W.-J.; Li, H.-Y.; Song, Q.-Y.; Dong, S.-H.; Gao, K. Meteroterpenoids with diverse ring systems and dioxolanone-type secondary metabolites from Phyllosticta capitalensis and their phytotoxic activity. Tetrahedron 2019, 75, 4611–4619. [Google Scholar] [CrossRef]
- Li, S.; Shao, M.-W.; Lu, Y.-H.; Kong, L.-C.; Jiang, D.-H.; Zhang, Y.-L. Phytotoxic and antibacterial metabolites from Fusarium proliferatum ZS07 isolated from the gut of long-horned grasshoppers. J. Agric. Food Chem. 2014, 62, 8997–9001. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-Y.; Tan, X.-M.; Wang, Y.-D.; Yang, J.; Zhang, Y.-G.; Sun, B.-D.; Gong, T.; Guo, L.-P.; Ding, G. Bioactive seco-sativene sesquiterpenoids from an Artemisia desertorum endophytic fungus, Cochliobolus sativus. J. Nat. Prod. 2020, 83, 1488–1494. [Google Scholar] [CrossRef]
- Lu, Y.; Li, S.; Shao, M.; Xiao, X.; Kong, L.; Jiang, D.; Zhang, Y. Isolation, identification, derivatization and phytotoxic activity of secondary metabolites produced by Cladosporium oxysporum DH14, a locust-associated fungus. J. Integr. Agric. 2016, 15, 832–839. [Google Scholar] [CrossRef] [Green Version]
- Mallik, M.A.B. Selective isolation and screening of soil microorganisms for metabolites with herbicidal potential. J. Crop Prot. 2001, 4, 219–236. [Google Scholar] [CrossRef]
- Li, H.; Wei, J.; Pan, S.-Y.; Gao, J.-M.; Tian, J.-M. Antifungal, phytotoxic and toxic metabolites produced by Penicillium purpurogenum. Nat. Prod. Res. 2014, 28, 2358–2361. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.-H.; Gou, J.-Y.; Zhao, D.-L.; Wang, D.; Liu, J.; Ma, G.-Y.; Li, Y.-Q.; Zhang, C.-S. Phytotoxicity and anti-phytopathogenic activities of marine-derived fungi and their secondary metabolites. RSC Adv. 2018, 8, 37573–37580. [Google Scholar] [CrossRef] [Green Version]
- Du, F.-Y.; Li, X.-M.; Sun, Z.-C.; Meng, L.-H.; Wang, B.-G. Secondary metabolites with agricultural antagonistic potentials from Beauveria felina, a marine-derived entomopathogenic fungus. J. Agric. Food Chem. 2020, 68, 14824–14831. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, R.; Chen, X.; Sun, X.; Yan, Y.; Shen, X.; Yuan, Q. Biosynthesis of aromatic polyketides in microorganisms using type II polyketide synthases. Microb. Cell Fact. 2020, 19, 110. [Google Scholar] [CrossRef]
- Masi, M.; Aloi, F.; Nocera, P.; Cacciola, S.O.; Surico, G.; Evidente, A. Phytotoxic metabolites isolated from Neufusicoccum batangarum, the causal agent of the scabby canker of cactus pear (Opuntia ficus-indica L.). Toxins 2020, 12, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meepagala, K.M.; Briscoe, W.E.; Techen, N.; Johnson, R.D.; Clausen, B.M.; Duke, S.O. Isolation of a phytotoxic isocoumarin from Diaporthe eres-infected Hedera helix (English ivy) and synthesis of its phytotoxic analogs. Pest Manag. Sci. 2018, 74, 37–45. [Google Scholar] [CrossRef] [Green Version]
- Masi, M.; Santoro, E.; Clement, S.; Meyer, S.; Scafato, P.; Superchi, S.; Evidente, A. Further secondary metabolites produced by the fungus Pyricularia grisea isolated from buffelgrass (Cenchrus ciliaris). Chirality 2020, 32, 1234–1242. [Google Scholar] [CrossRef] [PubMed]
- Hallock, Y.F.; Clardy, J.; Kenfield, D.S.; Strobel, G. De-O-methyl-diaporthin, a phytotoxin from Drechslera siccans. Phytochemistry 1988, 27, 3123–3125. [Google Scholar] [CrossRef]
- Robeson, D.J.; Strobel, G.A. Monocerin, a phytotoxin from Exserohilum turicum (Drechslera turcica). Agric. Biol. Chem. 1982, 46, 2681–2683. [Google Scholar]
- Parisi, A.; Piattelli, M.; Tringali, C.; Di San Lio, G.M. Indentificaton of the phytotoxin mullein in culture fluids of Phoma tracheiphila. Phytochemistry 1993, 32, 865–867. [Google Scholar] [CrossRef]
- Cimmino, A.; Maddau, L.; Masi, M.; Linaldeddu, B.T.; Evidente, A. Secondary metabolites produced by Sardiniella urbana, a new emerging pathogen on European hackberry. Nat. Prod. Res. 2019, 33, 1862–1869. [Google Scholar] [CrossRef] [PubMed]
- Masi, M.; Reveglia, P.; Baaijens-Billones, R.; Gorecki, M.; Pescitelli, G.; Savocchia, S.; Evidente, A. Phytotoxic metabolites from three Neofusicoccum species causal agents of Botryosphaeria dieback in Australia, luteopyroxin, neoanthraquinone, and luteoxepinone, a disubstituted furo-α-pyrone, a hexasubstituted anthraquinone, and a trisubstituted oxepi-2-one from Neofusicoccum luteum. J. Nat. Prod. 2020, 83, 453–460. [Google Scholar]
- Abou-Mansour, E.; Debieux, J.-L.; Ramirez-Suero, M.; Benard-Gellon, M.; Magnin-Robert, M.; Spagnolo, A.; Chong, J.; Farine, S.; Bertsch, C.; L’Haridon, F.; et al. Phytotoxic metabolites from Neofusicoccum parvum, a pathogen of Botryosphaeria dieback of grapevine. Phytochemistry 2015, 115, 207–215. [Google Scholar] [CrossRef]
- Masi, M.; Reveglia, P.; Femina, G.; Baaijens-Bilones, R.; Savochia, S.; Evidente, A. Luteoethanones A and B two phytotoxic 1-substituted ethanones produced by Neofusicoccum luteum, a causal agent of Botryosphaeria dieback on grapevine. Nat. Prod. Res. 2020. [Google Scholar] [CrossRef]
- Masi, M.; Meyer, S.; Clement, S.; Pescitelli, G.; Cimmino, A.; Crstofaro, M.; Evidente, A. Chloromonilinic acids C and D, phytotoxic tetrasubstituted 3-chromanonacrylic acids isolated from Cochliobolus australiensis with potential herbicidal activity against buffelgrass (Cenchrus ciliaris). J. Nat. Prod. 2017, 80, 2771–2777. [Google Scholar] [CrossRef]
- Bashiri, S.; Abdollahzadeh, J.; Lecce, R.D.; Alioto, D.; Gorecki, M.; Pescitelli, G.; Masi, M.; Evidente, A. Rabenchromenone and rabenzophenone, phytotoxic tetrasubstituted chromenone and hexasubstituted benzophenone constituents produced by the oak-decline-associated fungus Fimetariella rabenhorstii. J. Nat. Prod. 2020, 83, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.; Sun, W.; Fu, L.; Luo, H.; Lai, D.; Zhou, L. Natural dibenzo-α-pyrones and their bioactivities. Molecules 2014, 19, 5088–5108. [Google Scholar] [CrossRef]
- Tang, J.; Huang, L.; Liu, Y.; Toshmatov, Z.; Zhang, C.; Shao, H. Two phytotoxins isolated from the pathogenic fungus of the invasive weed Xanthium italicum. Chem. Biodivers. 2020, 17, e2000043. [Google Scholar] [CrossRef] [PubMed]
- Demuner, A.J.; Barbosa, L.C.A.; Miranda, A.C.M.; Geraldo, G.C.; Da Silva, C.M.; Giberti, S.; Bertazzini, M.; Forlani, G. The fungal phytotoxin alternariol 9-methyl ether and some of its synthetic analogues inhibit the photosynthetic electron transport chain. J. Nat. Prod. 2013, 76, 2234–2245. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.-L.; Han, X.-B.; Wang, M.; Zeng, Y.-T.; Li, Y.-Q.; Ma, G.-Y.; Liu, J.; Zheng, C.-J.; Wen, M.-X.; Zhang, Z.-F.; et al. Herbicidal and antifungal xanthone derivatives from the alga-derived fungus Aspergillus versicolor D5. J. Agric. Food Chem. 2020, 68, 11207–11214. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.-B.; Long, C.; Kennelly, E.J. Structural diversity and bioactivities of natural benzophenones. Nat. Prod. Rep. 2014, 31, 1158–1174. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, T.; Tahara, S.; Takaoka, S.; Tori, M.; Askawa, Y. Structures of a novel binaphthyl and three novel benzophenone derivatives with plant-growth inhibitory activity from the fungus Daldinia concentrica. Chem. Pharm. Bull. 1994, 42, 1528–1530. [Google Scholar] [CrossRef] [Green Version]
- Dalinova, A.; Chisty, L.; Kochura, D.; Garnyuk, V.; Petrova, M.; Prokofieva, D.; Yurchenko, A.; Dubovik, V.; Ivanov, A.; Smirnov, S.; et al. Isolation and bioactivity of secondary metabolites from solid culture of the fungus, Alternaria sonchi. Biomolecules 2020, 10, 81. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Tian, J.; Sun, W.; Meng, J.; Wang, X.; Fu, X.; Wang, A.; Lai, D.; Liu, Y.; Zhou, L. Bis-naphtho-γ-pyrones form fungi and their bioactivities. Molecules 2014, 19, 7169–7188. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Sun, W.; Meng, J.; Wang, A.; Wang, X.; Tian, J.; Fu, X.; Dai, J.; Liu, Y.; Lai, D.; et al. Bioactive bis-naphtho-γ-pyrones from rice false smut pathogen Ustilaginoidea virens. J. Agric. Food Chem. 2015, 63, 3501–3508. [Google Scholar] [CrossRef]
- Sun, W.; Wang, A.; Xu, D.; Wang, W.; Meng, J.; Dai, J.; Liu, Y.; Lai, D.; Zhou, L. New ustilaginoidins from rice false smut balls caused by Villosiclava virens and their phytotoxic and cytotoxic activities. J. Agric. Food Chem. 2017, 65, 5151–5160. [Google Scholar] [CrossRef]
- Gao, J.-M.; Yang, Y.-X.; Qin, J.-C. Azaphilones: Chemistry and biology. Chem. Rev. 2013, 113, 4755–4811. [Google Scholar] [CrossRef] [PubMed]
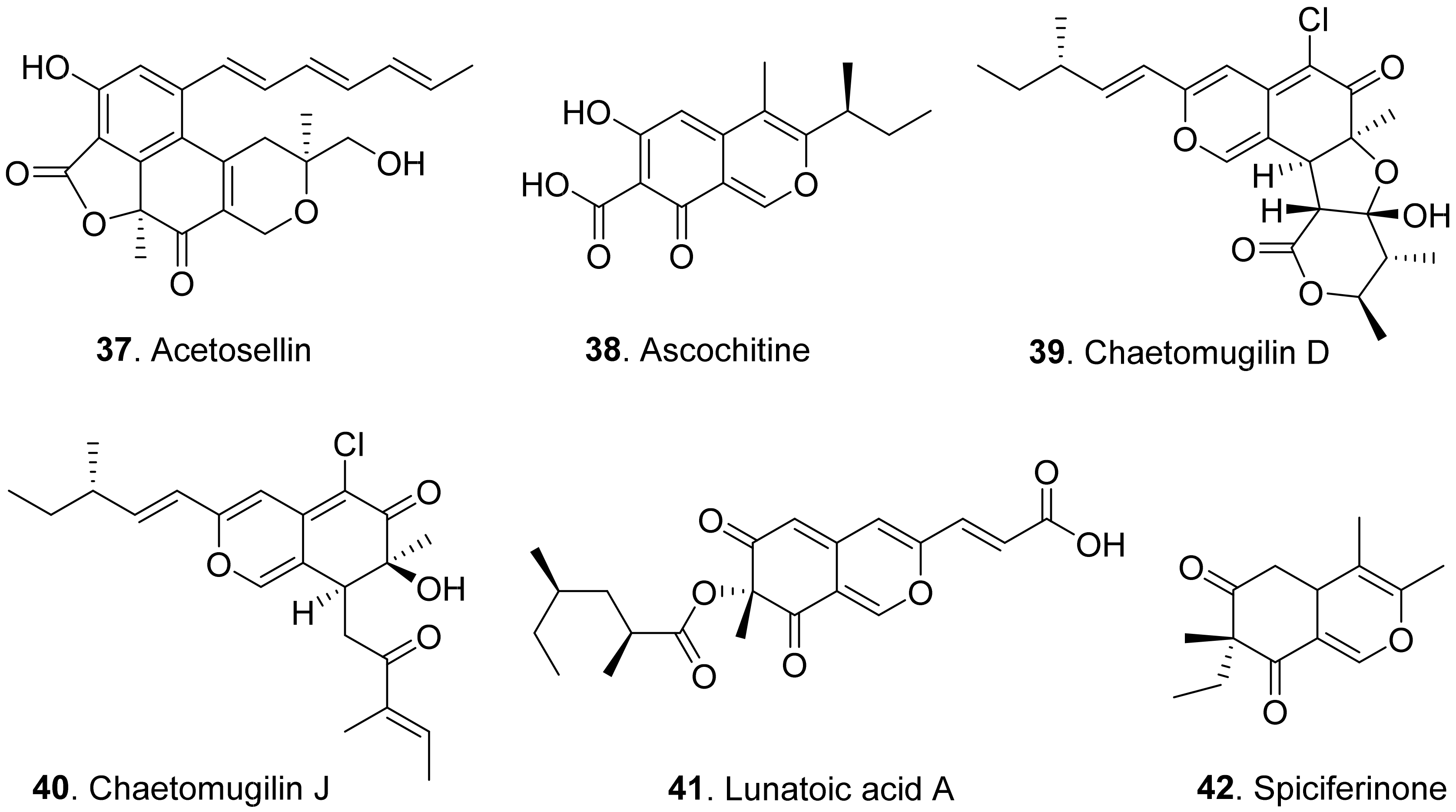
- Nasini, G.; Arnone, A.; Assante, G.; Candiani, G.; Vajna de Pava, O. Structure and absolute configuration of acetosellin, a new polyketide from a phytotoxic strain of Cercosporella acetosella. Tetrahedron Lett. 2002, 43, 1665–1668. [Google Scholar] [CrossRef] [Green Version]
- Iwai, I.; Mishima, H. Consitution of ascochitine. Chem. Ind. 1965, 73, 186–187. [Google Scholar]
- Smith, G.R.; Munro, M.H.G.; Fineran, B.A.; Cole, A.L.J. Evidence for the involvement of asochitine in phoma leafspot-wilt disease of Clematis. Physiol. Mol. Plant Pathol. 1994, 45, 333–348. [Google Scholar] [CrossRef]
- Nakajima, H.; Kimura, Y.; Hamasaki, T. Spiciferinone, an azaphilone phytotoxin produced by the fungus Cochliobolus spicifer. Phytochemistry 1992, 31, 105–107. [Google Scholar] [CrossRef]
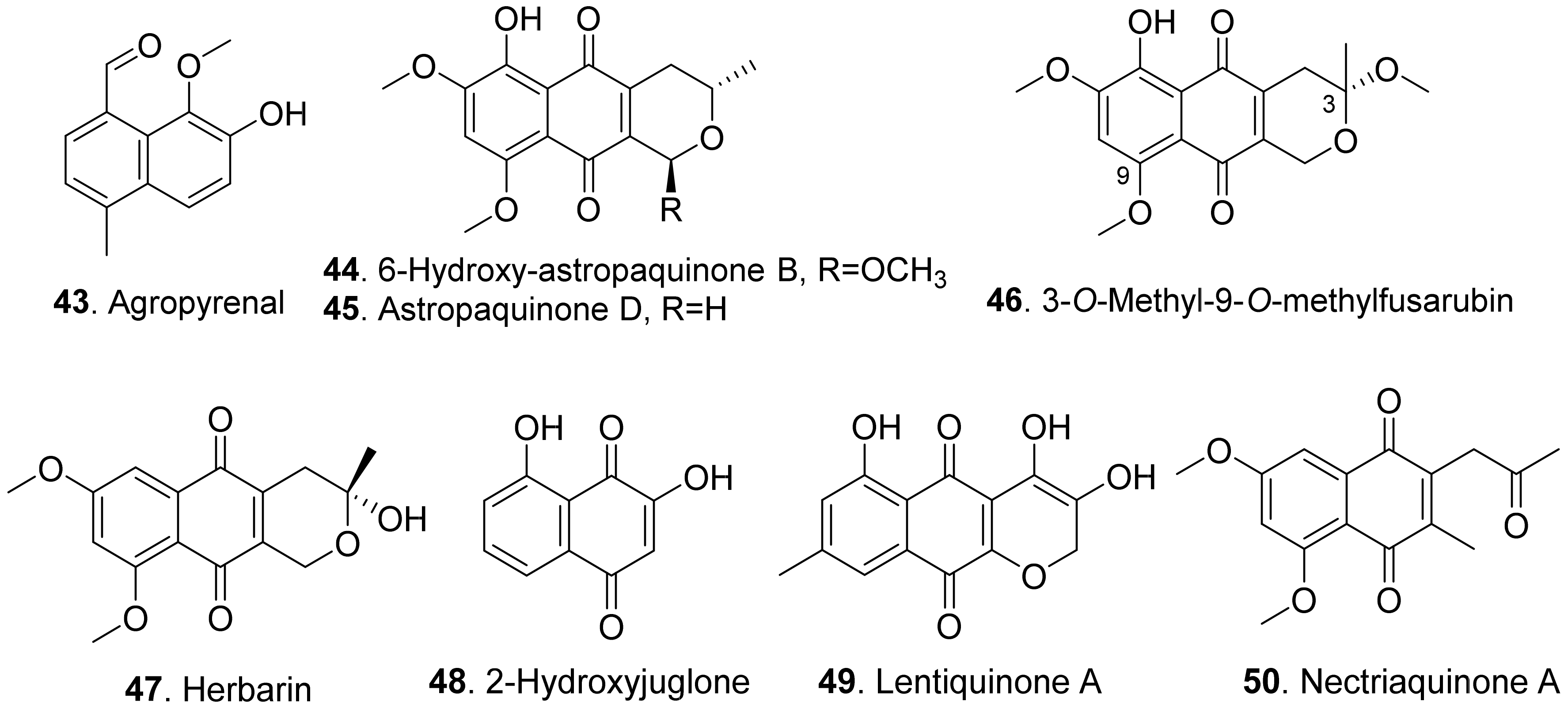
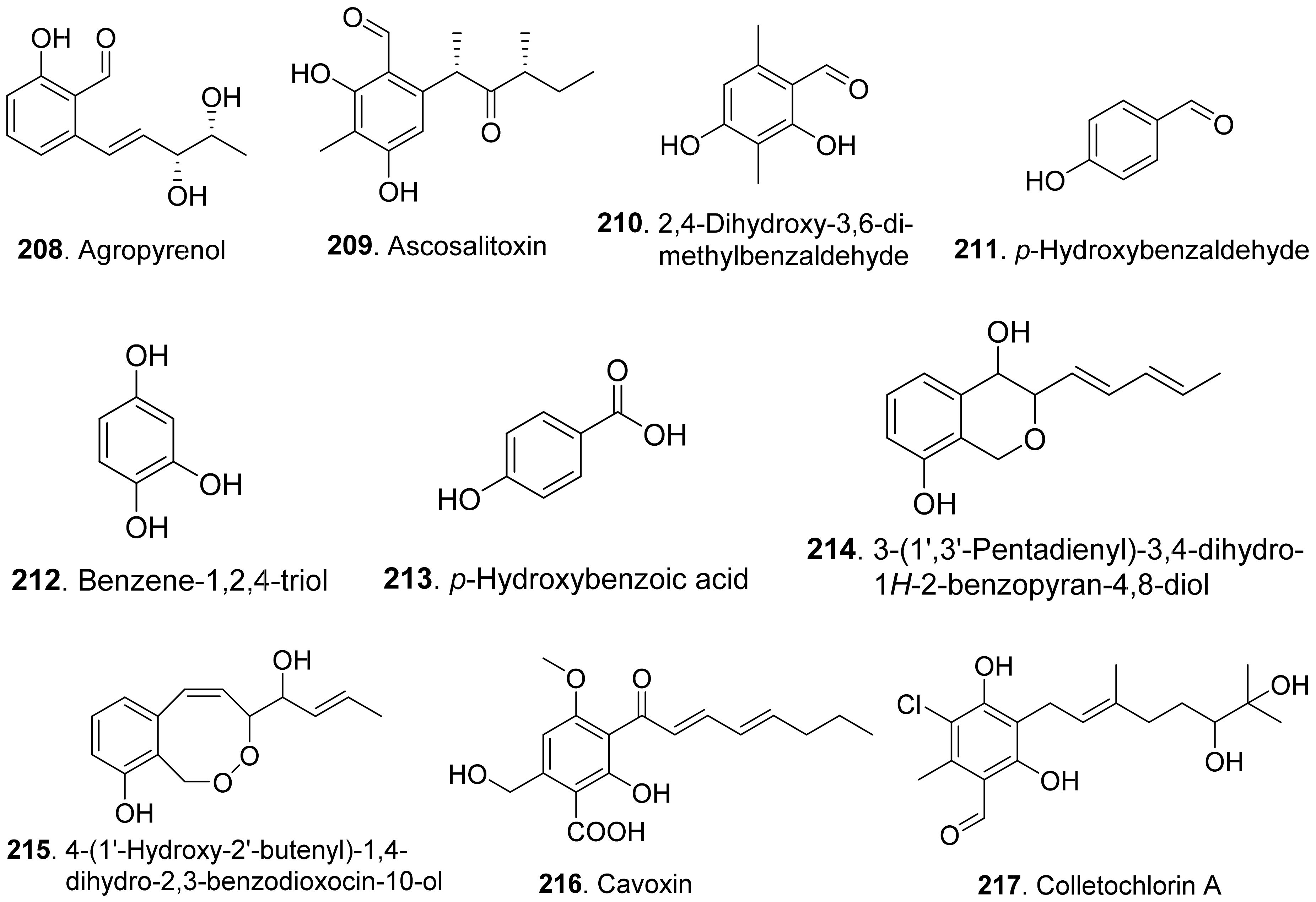
- Andolfi, A.; Cimmino, A.; Vurro, M.; Berestetskiy, A.; Troise, C.; Zonno, M.C.; Evidente, A. Agropyrenol and agropyrenal, phytotoxins from Ascochyta agropyrina var. nana, a fungal pathogen of Elitrigia repens. Phytochemsitry 2012, 79, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Supratman, U.; Hirai, N.; Sato, S.; Watanabe, K.; Malik, A.; Annas, S.; Harneti, D.; Maharani, R.; Koseki, T.; Shiono, Y. New naphthoquinone derivatives from Fusarium napiforme of a mangrove plant. Nat. Prod. Res. 2019. [Google Scholar] [CrossRef]
- Ariefta, N.R.; Kristiana, P.; Aboshi, T.; Murayama, T.; Tawaraya, K.; Koseki, T.; Kurisawa, N.; Kimura, K.; Shiono, Y. New isocoumarins, naphthoquinones, and a cleistanthane-type from Nectria pseudotrichia 120-1NP. Fitoterapia 2018, 127, 356–361. [Google Scholar] [CrossRef]
- Burki, N.; Michel, A.; Tabacchi, R. Naphthalenones and isocoumarins of the fungus Ceratocystis fimbriata f.sp. platani. Phytopathol. Mediterr. 2003, 42, 191–198. [Google Scholar]
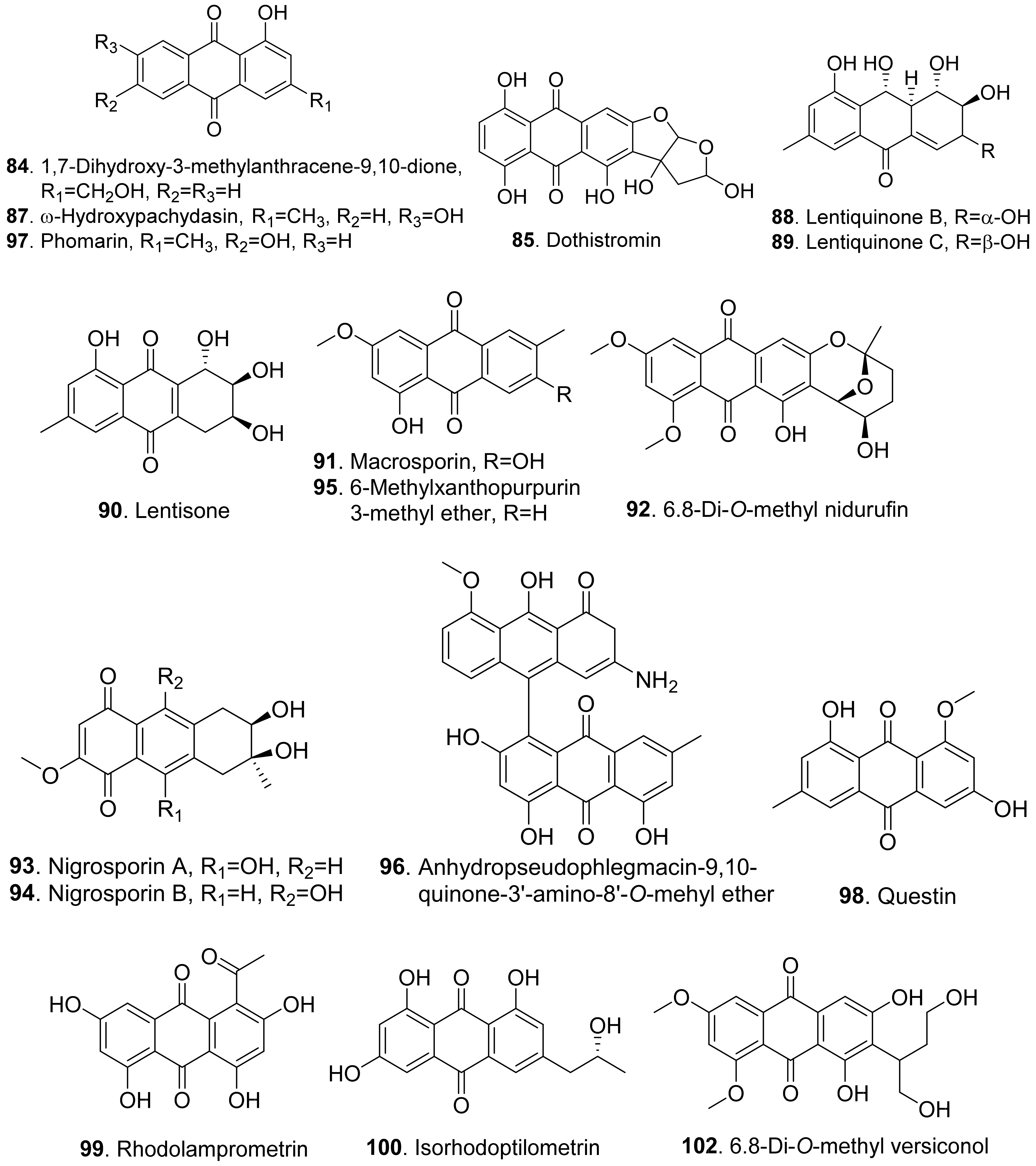
- Masi, M.; Nocera, P.; Zonno, M.C.; Tuzi, A.; Pescitelli, G.; Cimmino, A.; Boari, A.; Infantino, A.; Vurro, M.; Evidente, A. Lentiquinones A, B, and C, phytotoxic anthraquinone derivatives isolated from Ascochyta lentis, a pathogen of lentil. J. Nat. Prod. 2018, 81, 2700–2709. [Google Scholar] [CrossRef] [PubMed]
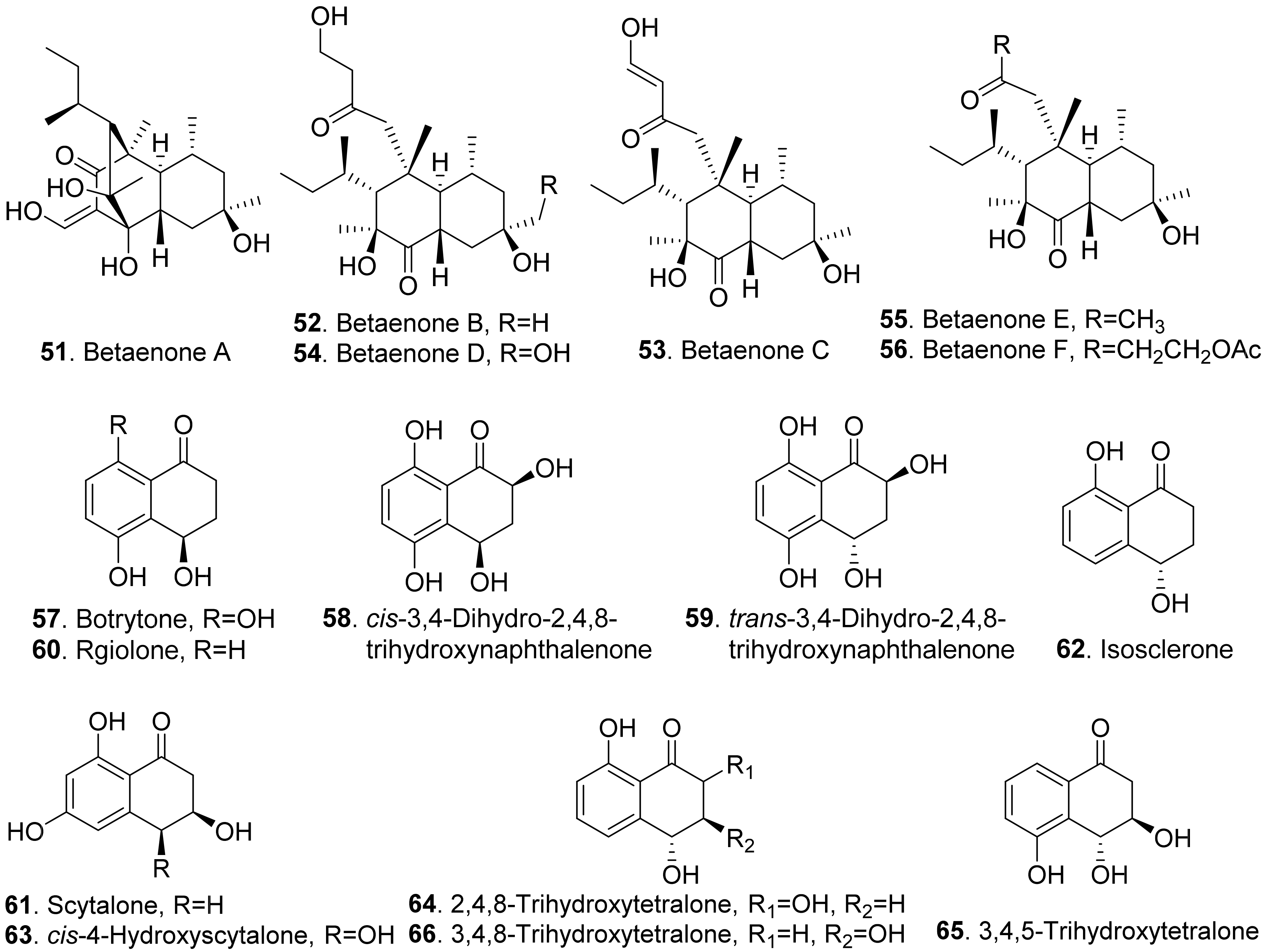
- Ichihara, A.; Oikawa, H.; Hayashi, K.; Sakamura, S. Structures of betaenones A and B, novel phytotoxins from Phoma betae Fr. J. Am. Chem. Soc. 1983, 105, 2907–2908. [Google Scholar] [CrossRef]
- Ichihara, A.; Oikawa, H.; Hashimoto, M.; Sakamura, S.; Haraguchi, T.; Nagano, H. A phytotoxin, betaenone C, and its related metabolites of Phoma betae Fr. Agric. Biol. Chem. 1983, 47, 2965–2967. [Google Scholar] [CrossRef]
- Cimmino, A.; Villegas-Fernandez, A.M.; Andolfi, A.; Melck, D.; Rubiales, D.; Evidente, A. Botrytone, a new naphthalenone pentaketide produced by Botrytis fabae, the causal agent of chocolate spot disease on Vicia faba. J. Agric. Food Chem. 2011, 59, 9201–9206. [Google Scholar] [CrossRef] [PubMed]
- Evidente, A.; Sparapano, L.; Andolfi, A.; Bruno, G. Two naphthalenone pentaketides from liquid cultures of Phaeoacremonium aleophilum, a fungus associated with esca of grapevine. Phytopathol. Mediterr. 2000, 39, 162–168. [Google Scholar]
- Nakamura, T.; Supratman, U.; Harneti, D.; Maharani, R.; Koseki, T.; Shiono, Y. New compounds from Japanese oak wilt disease-associated fungus Raffaelea quercivora. Nat. Prod. Res. 2020. [Google Scholar] [CrossRef]
- Tabacchi, R.; Fkyerat, A.; Poliart, C.; Dubin, G.M. Phytotoxins from fungi of esca of grapevine. Phytopathol. Mediterr. 2000, 39, 156–161. [Google Scholar]
- Abou-Mansour, A.; Couche, E.; Tabacchi, R. Do fungal naphthalenones have a role in the development of esca symptoms? Phytopathol. Mediterr. 2004, 43, 75–82. [Google Scholar]
- Borgschulte, K.; Rebuffat, S.; Trowitzsch-Kienast, W.; Schomburg, D.; Pinon, J.; Bodo, B. Isolation and structure elucidation of hymatoxins B-E and other phytotoxins from Hypoxylon mammatum fungal pathogen of leuce poplars. Tetrahedron 1991, 47, 8351–8360. [Google Scholar] [CrossRef]
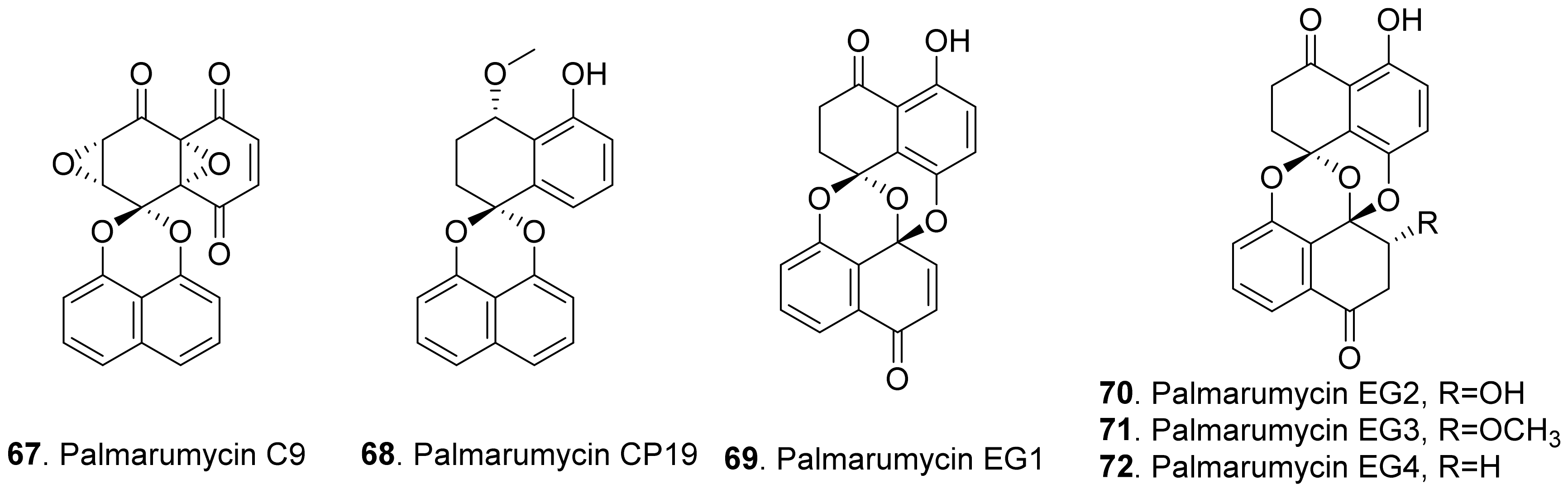
- Zhou, L.; Zhao, J.; Shan, T.; Cai, X.; Peng, Y. Spirobisnaphthalenes from fungi and their biological activities. Mini Rev. Med. Chem. 2010, 10, 977–989. [Google Scholar] [CrossRef]
- Liu, X.; Zhao, Y.; Wang, W.; Wang, M.; Zhou, L. Recent progress of natural product spirobisnaphthalenes. Chinese J. Org. Chem. 2017, 37, 2883–2894. [Google Scholar] [CrossRef]
- Krohn, K.; Michel, A.; Florke, U.; Aust, H.-J.; Draeger, S.; Schulz, B. Palmarumycins C1-C16 form Coniothyrium sp.: Isolation, structure elucidation, and biological activity. Liebigs Ann. Chem. 1994, 11, 1099–1108. [Google Scholar] [CrossRef]
- Macias-Rubalcava, M.; Sobrino, M.E.R.-V.; Melendez-Gonzalez, C.; Hernandez-Ortega, S. Naphthoquinone spiroketals and organic extracts from the endophytic fungus Ednia gomezpompae as potential herbicides. J. Agric. Food Chem. 2014, 62, 3553–3562. [Google Scholar] [CrossRef] [PubMed]
- Masi, M.; Evidente, A. Fungal bioactive anthraquinones and analogues. Toxins 2020, 12, 714. [Google Scholar] [CrossRef]
- Evidente, A.; Rodeva, R.; Andolfi, A.; Stoyanova, Z.; Perrone, C.; Motta, A. Phytotoxic polyketides produced by Phomopsis foeniculi, a strain isolated from diseased Bulgarian fennel. Eur. J. Plant Pathol. 2011, 130, 173–182. [Google Scholar] [CrossRef]
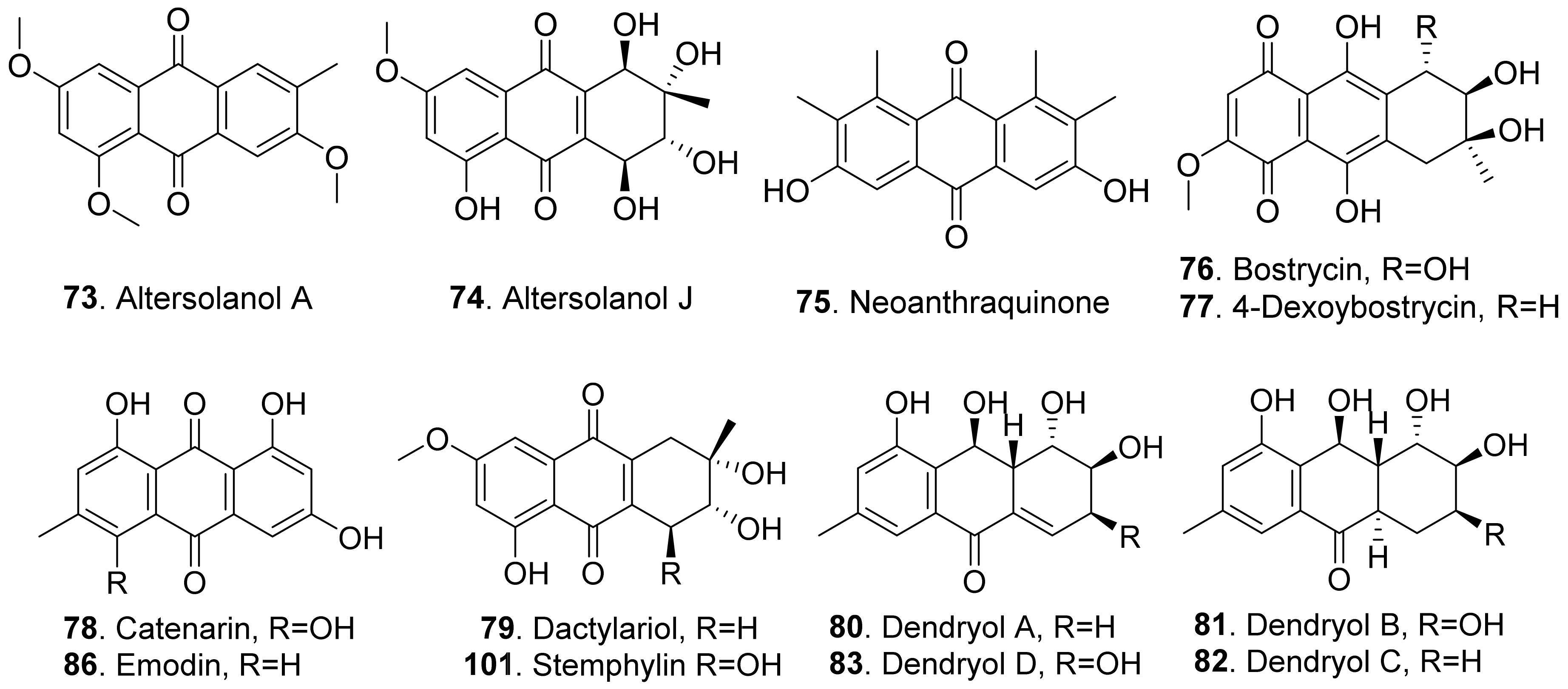
- Suemitsu, R.; Yamada, Y.; Sano, T.; Yamashita, K. Phytotoxic activities of altersolanol A, B and dactylariol, and activities of altersolanol A against some microorganism. Agric. Biol. Chem. 1984, 48, 2383–2384. [Google Scholar]
- Charudattan, R.; Rao, K.V. Bostrycin and 4-deoxybostrycin: Two nonspecific phytotoxins produced by Alternaria eichhorniae. Appl. Environ. Microbiol. 1982, 43, 846–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babu, R.M.; Saeena, A.; Seetharaman, K.; Ebenezar, E.G.; Raja, A.S.; Biji, K.R.; Prakash, M.S. Isolation and characterisation of phytotoxic substance from Alternaria alternata—A leaf blight pathogen on Eichhornia crassipes. J. Ecobiol. 2003, 15, 315–318. [Google Scholar]
- Bouras, N.; Strelkov, S.E. The anthraquinone catenarin is phytotoxic and produced in leaves and kernels of wheat infected by Pyrenophora tritici-repentis. Physiol. Mol. Plant Pathol. 2008, 72, 87–95. [Google Scholar] [CrossRef]
- Assante, G.; Nasini, G. Identity of the phytotoxin stemphylin from Stemphylium botryosum with altersolanol A. Phytochemistry 1987, 26, 703–705. [Google Scholar] [CrossRef]
- Tanaka, M.; Ohra, J.; Tsujino, Y.; Fujimori, T.; Ago, H.; Tsuge, H. Dendryol A, B, C, and D, four new compounds produced by a weed pathogenic fungus Dendryphiella sp. Z. Naturfursch. C 1995, 50, 751–756. [Google Scholar] [CrossRef]
- Gallagher, R.; Hodeges, R. The chemistry of dothistromin a difuronanthraquinone from Dothistroma pini. Aust. J. Chem. 1972, 25, 2399–2407. [Google Scholar] [CrossRef]
- Schor, R.; Cox, R. Classic fungal natural products in the genomic age: The molecular legacy of Harold Raistrick. Nat. Prod. Rep. 2018, 35, 230–256. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Fu, J.; Yin, X.; Cao, S.; Li, X.; Lin, L.; Qi, H.; Ni, J. Emodin: A review of its pharmacology, toxicity and pharmacokinetics. Phytother. Res. 2016, 30, 1207–1218. [Google Scholar] [CrossRef] [PubMed]
- Izhaki, I. Emodin: A secondary metabolite with multiple ecological functions in higher plants. New Phytol. 2002, 155, 205–217. [Google Scholar] [CrossRef] [Green Version]
- Macias, M.; Ulloa, M.; Gamboa, A.; Mata, R. Phytotoxic compounds from the new coprophilous fungus Guanomyces polythrix. J. Nat. Prod. 2000, 63, 757–761. [Google Scholar] [CrossRef]
- Andolfi, A.; Cimmino, A.; Villegas-Fernadez, A.M.; Tuzi, A.; Santini, A.; Melck, D.; Rubiales, D.; Evidente, A. Lentisone, a new phytotoxic anthraquinone produced by Aschyta lentis, the causal agent of Ascochyta blight in Lens culinaris. J. Agric. Food Chem. 2013, 61, 7301–7308. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, S. Metabolites of phytotoxic fungi. I. Isolation of macrosporin and 6-methylxanthopurpurin 3-methyl ether from Alternaria bataticola. Chem. Pharm. Bull. 1973, 21, 2083–2085. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Fukushima, T.; Tsujino, Y.; Fujimori, T. Nigrosporins A and B, new phytotoxic and antibacterial metabolites produced by a fungus Nigrospora oryzae. Biosci. Biotechnol. Biochem. 1997, 61, 1848–1852. [Google Scholar] [CrossRef] [Green Version]
- Quereshi, S.; Khan, N.A.; Pandey, A.K. Anthraquinone pigment with herbicidal potential from Phoma herbarum FGCC# 54. Chem. Nat. Compd. 2011, 47, 521–523. [Google Scholar]
- Gui, R.-Y.; Xu, L.; Kuang, Y.; Chung, I.-M.; Qin, J.-C.; Liu, L.; Yang, S.-X.; Zhao, L.-C. Chaetominine, (+)-alantrypinone, questin, isorhodoptilometrin, and 4-hydroxybenzaldehyde produced by the endophytic fungus Aspergillus sp. YL-6 inhibit wheat (Triticum aestivum) and radish (Raphanus sativus) germination. J. Plant Interact. 2015, 10, 87–92. [Google Scholar]
- Daub, M.E.; Herrero, S.; Chung, K.-R. Photoactivated perylenequinone toxins in fungal pathogenesis of plants. FEMS Microbiol. Lett. 2005, 252, 197–206. [Google Scholar] [CrossRef] [Green Version]
- Stierle, A.C.; Cardellina II, J.H.; Strobel, G.A. Phytotoxins from Alternaria alternata, a pathogen of spotted knapweed. J. Nat. Prod. 1989, 52, 42–47. [Google Scholar] [CrossRef]
- Hradil, C.M.; Hallock, Y.F.; Clardy, J.; Kenfield, D.S.; Strobel, G. Phytotoxins from Alternaria cassiae. Phytochemistry 1989, 28, 73–75. [Google Scholar] [CrossRef]
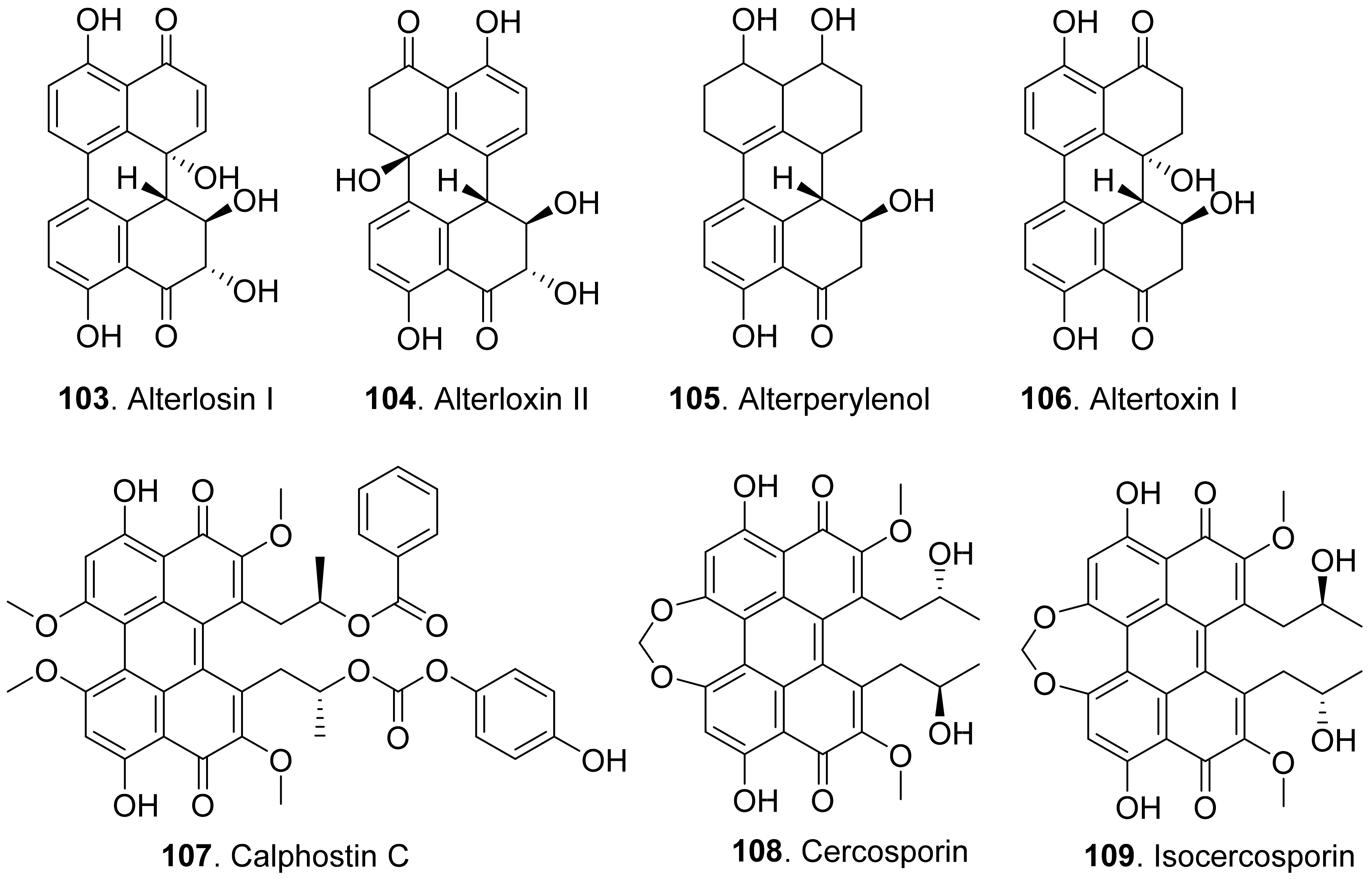
- Bruns, R.F.; Miller, F.D.; Merriman, R.L.; Howbert, J.J.; Heath, W.F.; Kobayashi, E.; Takahashi, I.; Tamaoki, T.; Nakano, H. Inhibition of protein kinase C by calphostin C is light-dependent. Biochem. Biophys. Res. Commun. 1991, 176, 288–293. [Google Scholar] [CrossRef]
- Daub, M.E. Cercosporin, a photosensitizing toxin from Cercospora pecies. Phytopathology 1982, 72, 370–374. [Google Scholar] [CrossRef]
- Daub, M.E.; Ehrenshaft, M. The photoactivated Cercospora toxin cercosporin: Contributions to plant disease and fundamental biology. Annu. Rev. Phytopathol. 2000, 38, 461–490. [Google Scholar] [CrossRef] [Green Version]
- Tabuchi, H.; Tajimi, A.; Ichihara, A. Phytotoxic metabolites isolated from Scolecotrichum graminis Fuckel. Biosci. Biotechnol. Biochem. 1994, 58, 1956–1959. [Google Scholar] [CrossRef]
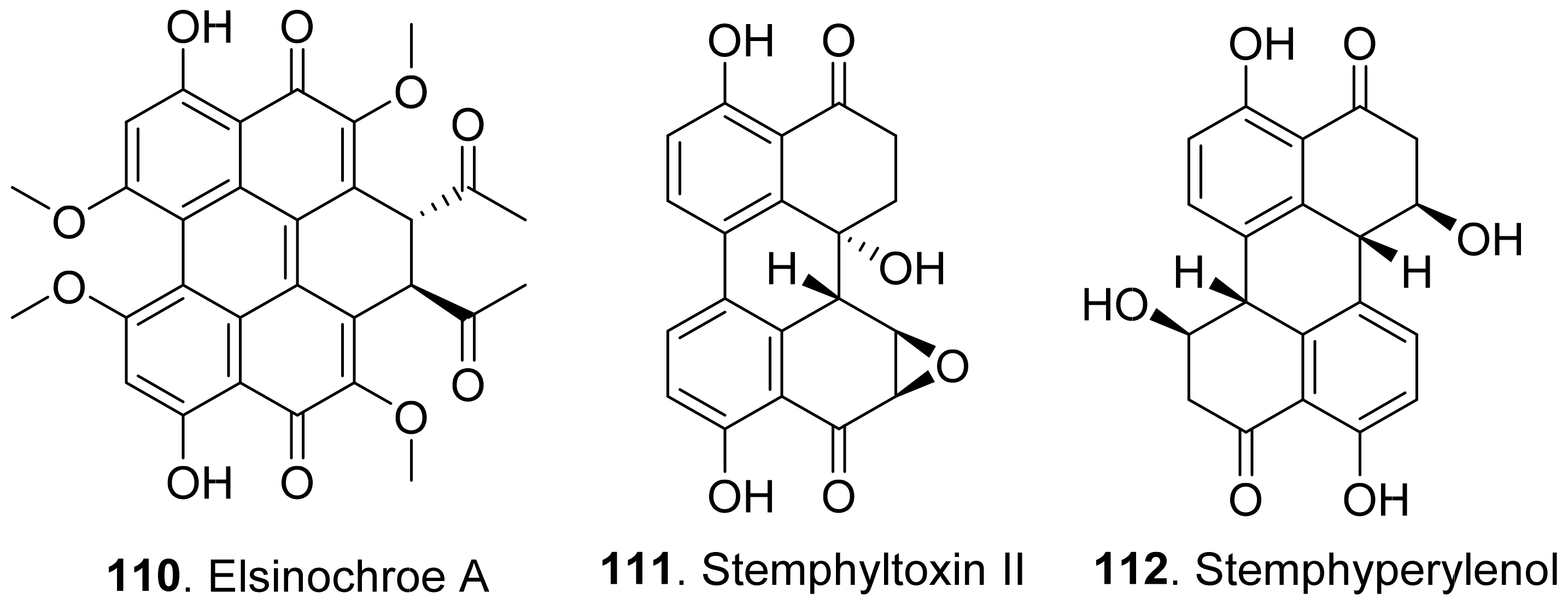
- Boss, D.; Maurhofer, M.; Schlapfer, E.; Defago, G. Elsinochrome A production by the bindweed biocontrol fungus Stagonospora convolvuli LA39 does not pose a risk to the environmentor the consumerof treated crops. FEMS Microbiol. Ecol. 2007, 59, 194–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
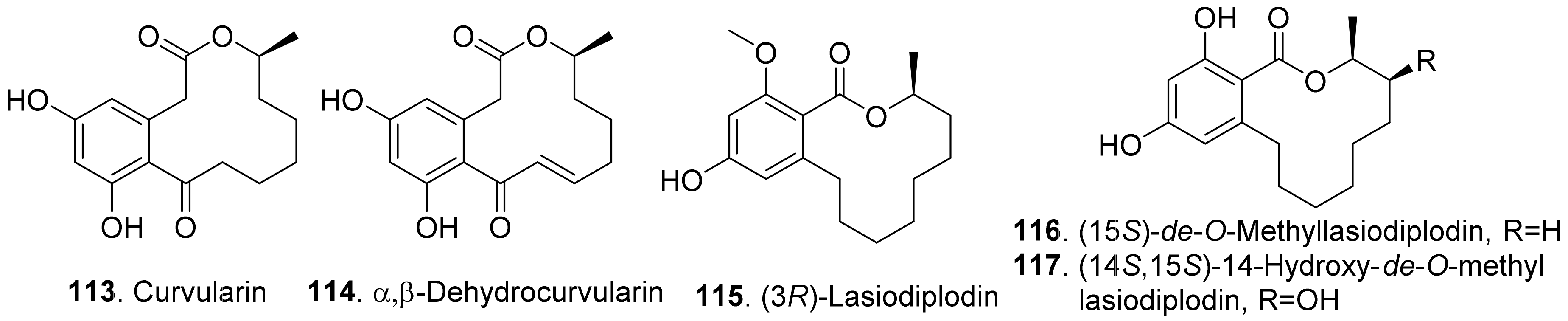
- Meepagala, K.M.; Johnson, R.D.; Robert, D.; Duke, S.O. Curvularin and dehydrocurvularin as phytotoxic constituents from Curvularia intermedia infecting Pandanus amaryllifolius. J. Agric. Chem. Environ. 2016, 5, 12–22. [Google Scholar]
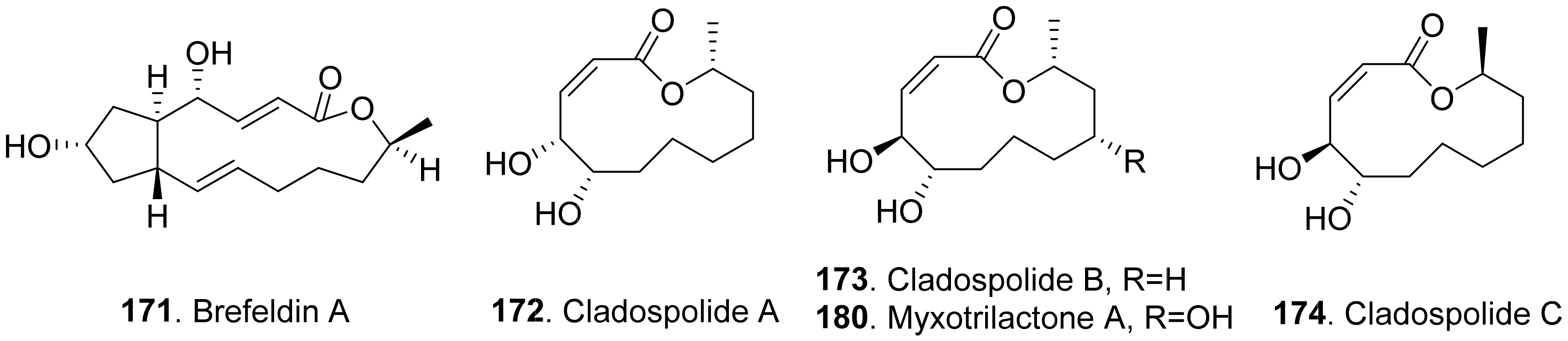
- Vurro, M.; Evidente, A.; Andolfi, A.; Zonno, M.C.; Giordano, F.; Motta, A. Brefeldin A and α,β-dehydrocurvularin, two phytotoxins from Alternaria zinniae, a biocontrol agent of Xanthium occidentale. Plant Sci. 1998, 138, 67–79. [Google Scholar] [CrossRef]
- Gutierrez, M.; Thoduloz, C.; Rodriguez, J.; Lolas, M.; Schmeda-Hirschmann, G. Bioactive metabolites from the fungus Nectria galligena, the main apple canker agent in Chile. J. Agric. Food Chem. 2005, 53, 7701–7708. [Google Scholar] [CrossRef]
- Veiga, T.A.M.; Silva, S.C.; Francisco, A.-C.; Filho, E.R.; Vieira, P.C.; Fernandes, J.B.; Silva, M.F.G.F.; Muller, M.W.; Lotina-Hennen, B. Inhibition of photophosphorylation and electron transport chain in thyakoids by lasiodiplodin, a natural product from Botryosphaeria rhodina. J. Agric. Food Chem. 2007, 55, 4217–4221. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Sofian, F.F.; Suehiro, W.; Harneti, D.; Maharani, R.; Supratman, U.; Abdullah, F.F.A.; Koseki, T.; Shiono, Y. β-Resorcylic acid derivatives, with their phytotoxic activities, from the endophytic fungus Lasiodiplodia theobromae in the mangrove plant Xylocarpus granatum. Chem. Biodivers. 2021, 18, e2000928. [Google Scholar] [CrossRef] [PubMed]
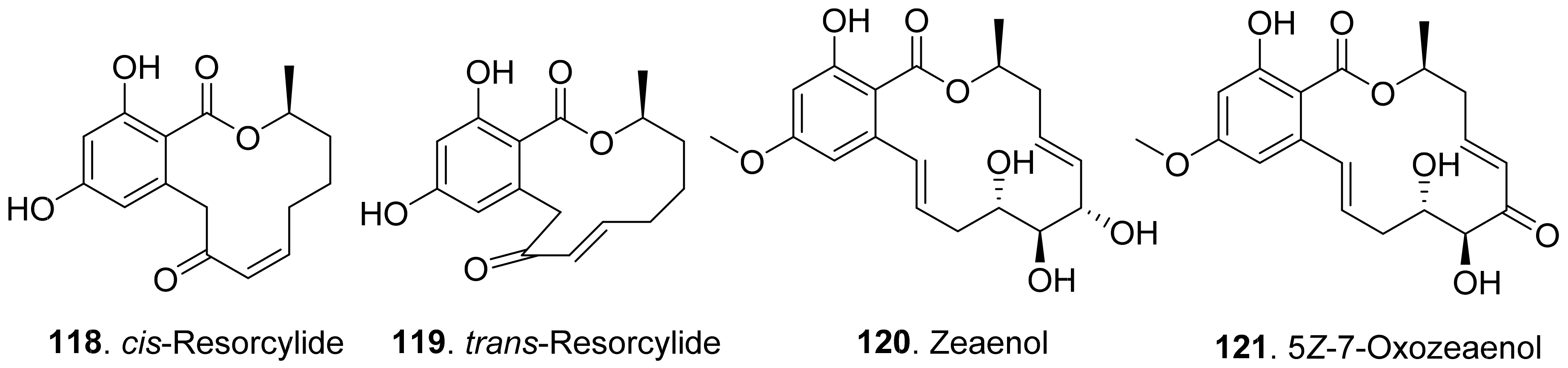
- Oyama, H.; Sassa, T.; Ikeda, M. Structures of new plant growth inhibitors, trans- and cis-resorcylide. Agric. Biol. Chem. 1978, 42, 2407–2409. [Google Scholar] [CrossRef]
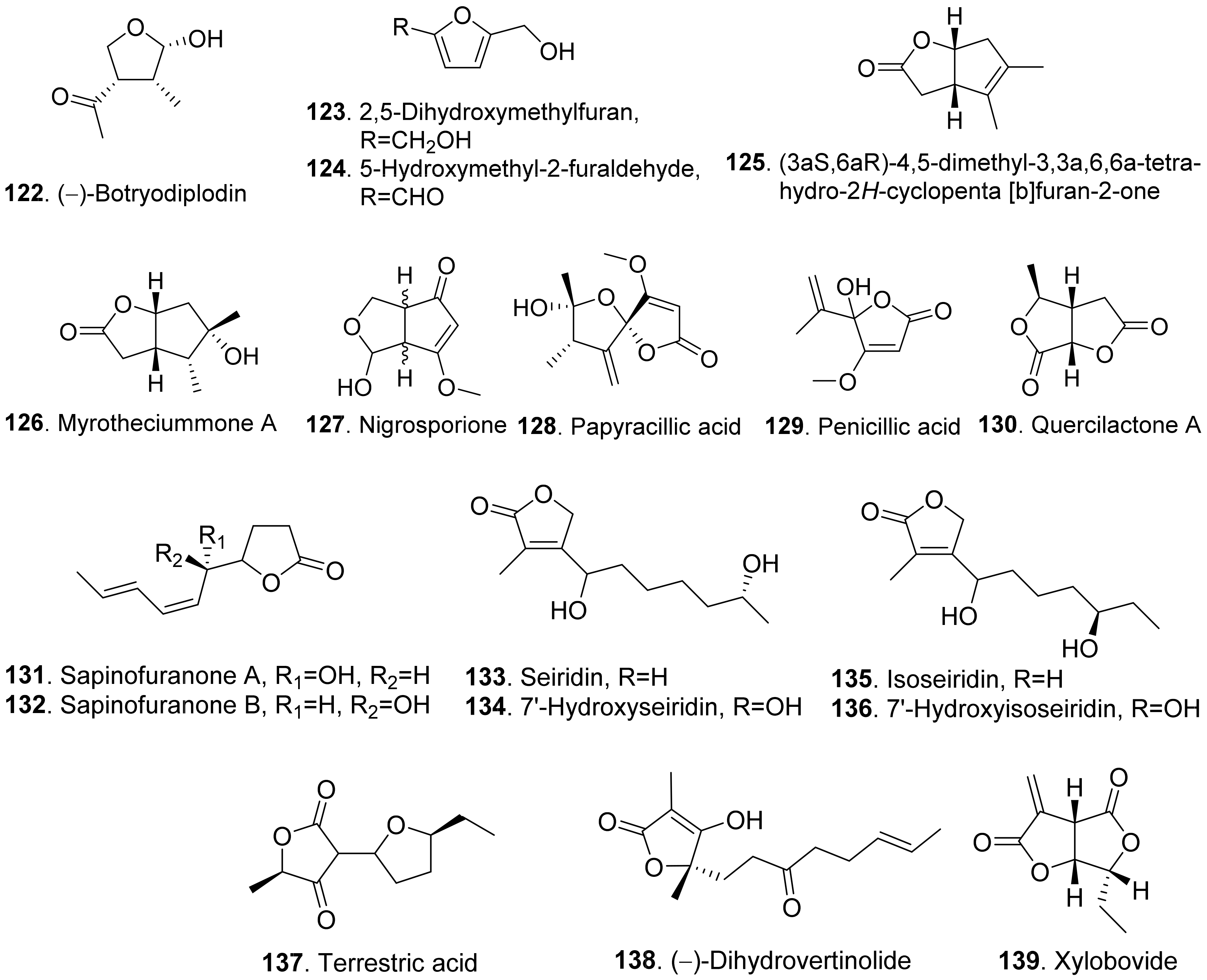
- Ramezani, M.; Shier, W.T.; Abbas, H.K.; Tonos, J.L.; Baird, R.E.; Sciumbato, G.L. Soybean charcoal rot disease fungus Macrophomina phaseolina in Mississippi produces the phytotoxin (−)-botryodiplodin but no detectable phaseolinone. J. Nat. Prod. 2007, 70, 128–129. [Google Scholar] [CrossRef]
- Nouguier, R.; Gastaldi, S.; Stien, D.; Bertrand, M.; Villar, F.; Andrey, O.; Renaud, P. Synthesis of (±)- and (−)-botryodiplodin using stereoselective radical cyclizations of acysclic esters and acetals. Tetrahedron Asymmetry 2003, 14, 3005–3018. [Google Scholar] [CrossRef]
- Di Lecce, R.; Bashiri, S.; Masi, M.; Alioto, D.; Tuzi, A.; Abdollahzadeh, J.; Evidente, A. Phytotoxic metabolites from Stilbocrea macrostoma, a fungal pathogen of Quercus brantii in Iran. Nat. Prod. Res. 2020. [Google Scholar] [CrossRef]
- Tchoukoua, A.; Ota, T.; Akanuma, R.; Ju, Y.-M.; Supratman, U.; Murayama, T.; Koseki, T.; Shiono, Y. A phytotoxic bicyclic lctones and other compounds from endophyte Xylaria curta. Nat. Prod. Res. 2017, 31, 2113–2118. [Google Scholar] [CrossRef]
- Evidente, A.; Berestetskiy, A.; Cimmino, A.; Tuzi, A.; Superchi, S.; Melck, D.; Andolfi, A. Papyracillic acid, a phytotoxic 1,6-dioxaspiro[4,4]nonene produced by Ascochyta agropyrina var. nana, a potential mycoherbicide for Elytrigia repens biocontrol. J. Agric. Food Chem. 2009, 57, 11168–11173. [Google Scholar] [CrossRef]
- Martinez-Luis, S.; Gonalez, M.C.; Ulloa, M.; Mata, R. Phytotoxins from the fungus Malbranchea aurantiaca. Phytochemistry 2005, 66, 1012–1016. [Google Scholar] [CrossRef]
- Evidente, A.; Sparapano, L.; Fierro, O.; Bruno, G.; Motta, A. Sapinofuranones A and B, two new 2(3H)-dihydrofuranones produced by Sphaeropsis sapinea, a common pathogen of conifers. J. Nat. Prod. 1999, 62, 253–256. [Google Scholar] [CrossRef] [PubMed]
- Sparapano, L.; Evidente, A. Studies on structure-activity relationship of seiridins, phytotoxins produced by three species of Seiridium. Nat. Toxins 1995, 3, 166–173. [Google Scholar] [CrossRef]
- Evidente, A.; Sparapano, L. 7′-Hydroxyseiridin and 7′-hydroxyisoseiridin two new phytotoxic Δα,β-butanolides from three species of Seiridium pathogenic to cypresses. J. Nat. Prod. 1994, 57, 1720–1725. [Google Scholar] [CrossRef]
- Nukina, M. Terrestric acid as a phytotoxic metabolite from Pyricularia oryzae Cavara. Agric. Biol. Chem. 1988, 52, 2357–2358. [Google Scholar] [CrossRef]
- Supratman, U.; Suzuki, T.; Nakamura, T.; Yokoyama, Y.; Harneti, D.; Maharani, R.; Salam, S.; Abdullah, F.F.; Koseki, T.; Shiono, Y. New metabolites produced by endophytic Clonostachys rosea B5-2. Nat. Prod. Res. 2019. [Google Scholar] [CrossRef] [PubMed]
- Abate, D.; Abraham, W.-R.; Meyer, H. Cytochalasins and phytotoxins from the fungus Xylaria obovata. Phytochemistry 1997, 44, 1443–1448. [Google Scholar] [CrossRef]
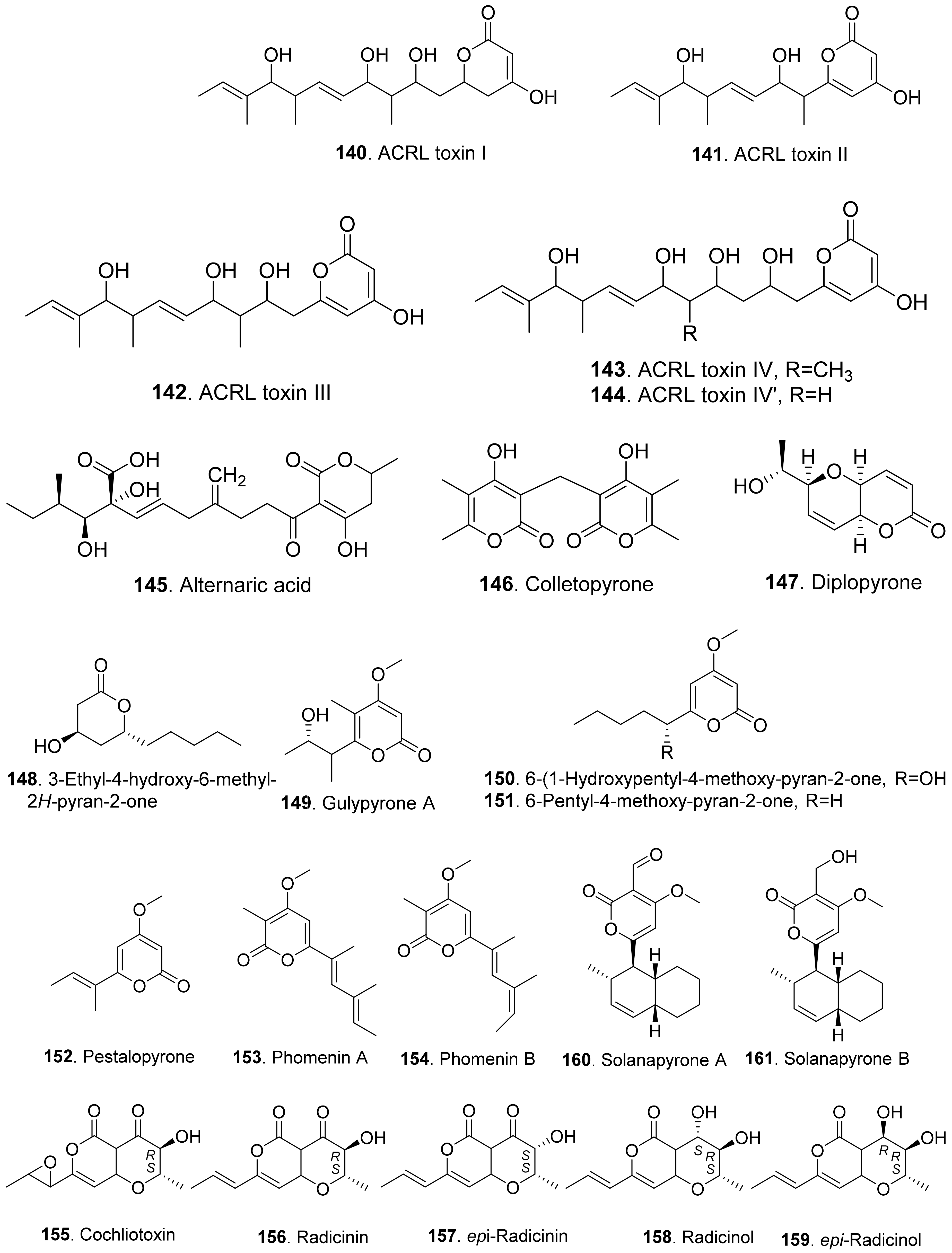
- Gardner, J.M.; Kono, Y.; Tatum, J.H.; Suzuki, Y.; Takeuchi, S. Plant pathotoxins from Alternaria citri: The major toxin specific for rough lemon plants. Phytochemistry 1985, 24, 2861–2867. [Google Scholar] [CrossRef]
- Kono, Y.; Gardner, J.M.; Suzuki, Y.; Takeuchi, S. Plant pathotoxins from Alternaria citri: The minor ACRL toxins. Phytochemistry 1985, 24, 2869–2874. [Google Scholar] [CrossRef]
- Maiero, M.; Bean, G.A.; Ng, T.J. Toxin production by Alternaria solani and its related phytotoxicity to tomato breeding lines. Phytopathology 1991, 81, 1030–1033. [Google Scholar] [CrossRef]
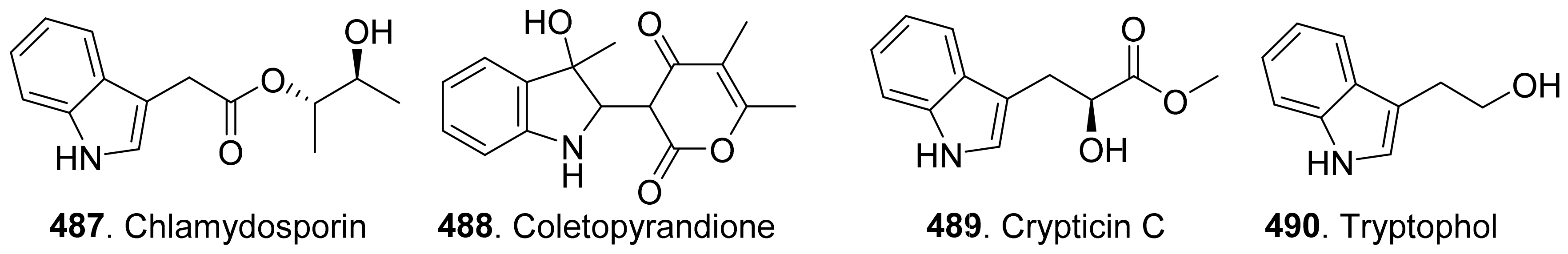
- Gohbara, M.; Hyeon, S.B.; Suzuki, A.; Tamura, S. Isolation and structure elucidation of colletopyrone from Colletotrichum nicotianae. Agric. Biol. Chem. 1976, 40, 1453–1455. [Google Scholar] [CrossRef]
- Evidente, A.; Maddau, L.; Spanu, E.; Franceschini, A.; Lazzaroni, S.; Motta, A. Diplopyrone, a new phytotoxic tetrahydropyranpyran-2-one produced by Diplodia mutila, a fungus pathogen of cork oak. J. Nat. Prod. 2003, 66, 313–315. [Google Scholar] [CrossRef]
- Masi, M.; Maddau, L.; Linaldeddu, B.T.; Cimmino, A.; D’Amico, W.; Scanu, B.; Evidente, M.; Tuzi, A.; Evidente, A. Bioactive secondary metabolites produced by the oak pathogen Diplodia corticola. J. Agric. Food Chem. 2016, 64, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Fuse, M.; Mazzeo, G.; Longhi, G.; Abbate, S.; Masi, M.; Evident, A.; Puzarini, C.; Barone, V. Unviased determination of absolute configurations by vis-à-vis comparison of experimental and simulated spectra: The challenging case of diplopyrone. J. Phys. Chem. B 2019, 123, 9230–9237. [Google Scholar] [CrossRef] [PubMed]
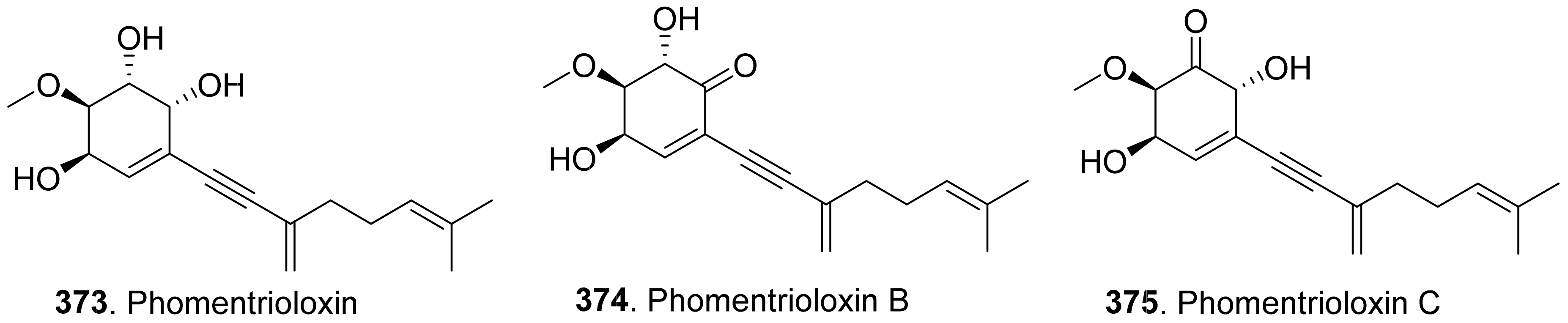
- Andolfi, A.; Boari, A.; Evidente, M.; Cimmino, A.; Vurro, M.; Ash, G.; Evidente, A. Gulypyrones A and B and phomentrioloxins B and C produced by Diaporthe gulyae, a potential mycoherbicide for saffron thistle (Carthamus lanatus). J. Nat. Prod. 2015, 78, 623–629. [Google Scholar] [CrossRef]
- Evidente, A.; Zonno, M.C.; Andolfi, A.; Troise, C.; Cimmino, A.; Vurro, M. Phytotoxic α-pyrones produced by Pestalotiopsis guepinii, the causal agent of hazelnut twig blight. J. Antibiot. 2012, 65, 203–206. [Google Scholar] [CrossRef]
- Turkkan, M.; Andolfi, A.; Zonno, M.C.; Erper, I.; Perrone, C.; Cimmino, A.; Vurro, M.; Evidente, A. Phytotoxins produced by Pestalotiopsis guepinii, the causal agent of hazelnut twig blight. Phytopathol. Mediterr. 2011, 50, 154–158. [Google Scholar]
- Tringali, C.; Parisi, A.; Piattelli, M.; Di San Lio, G.M. Phomenins A and B, bioactive polypropinoate pyrones from culture fluids of Phoma tracheiphila. Nat. Prod. Lett. 1993, 3, 101–106. [Google Scholar] [CrossRef]
- Solfrizzo, M.; Vitti, C.; De Girolamo, A.; Visconti, A.; Logrieco, A.; Fanizzi, F.P. Radicinols and radicinin phytotoxins produced by Alternaria radicina on carrots. J. Agric. Food Chem. 2004, 52, 3655–3660. [Google Scholar] [CrossRef]
- Masi, M.; Meyer, S.; Clement, S.; Cimmino, A.; Cristofaro, M.; Evidente, A. Cochliotoxin, a dihydropyranopyran-4,5-dione, and its analogues produced by Cochliobolus australiensis display phytotoxic activity against buffelgrass (Cenchrus ciliaris). J. Nat. Prod. 2017, 80, 1241–1247. [Google Scholar] [CrossRef]
- Zhang, Y.; Kong, L.; Jiang, D.; Yin, C.; Cai, Q.; Chen, Q.; Zheng, J. Phytotoxic and antifungal metabolites from Curvularia sp. FH01 isolated from the gut of Atractomorpha sinensis. Bioresource Technol. 2011, 102, 3575–3577. [Google Scholar] [CrossRef] [PubMed]
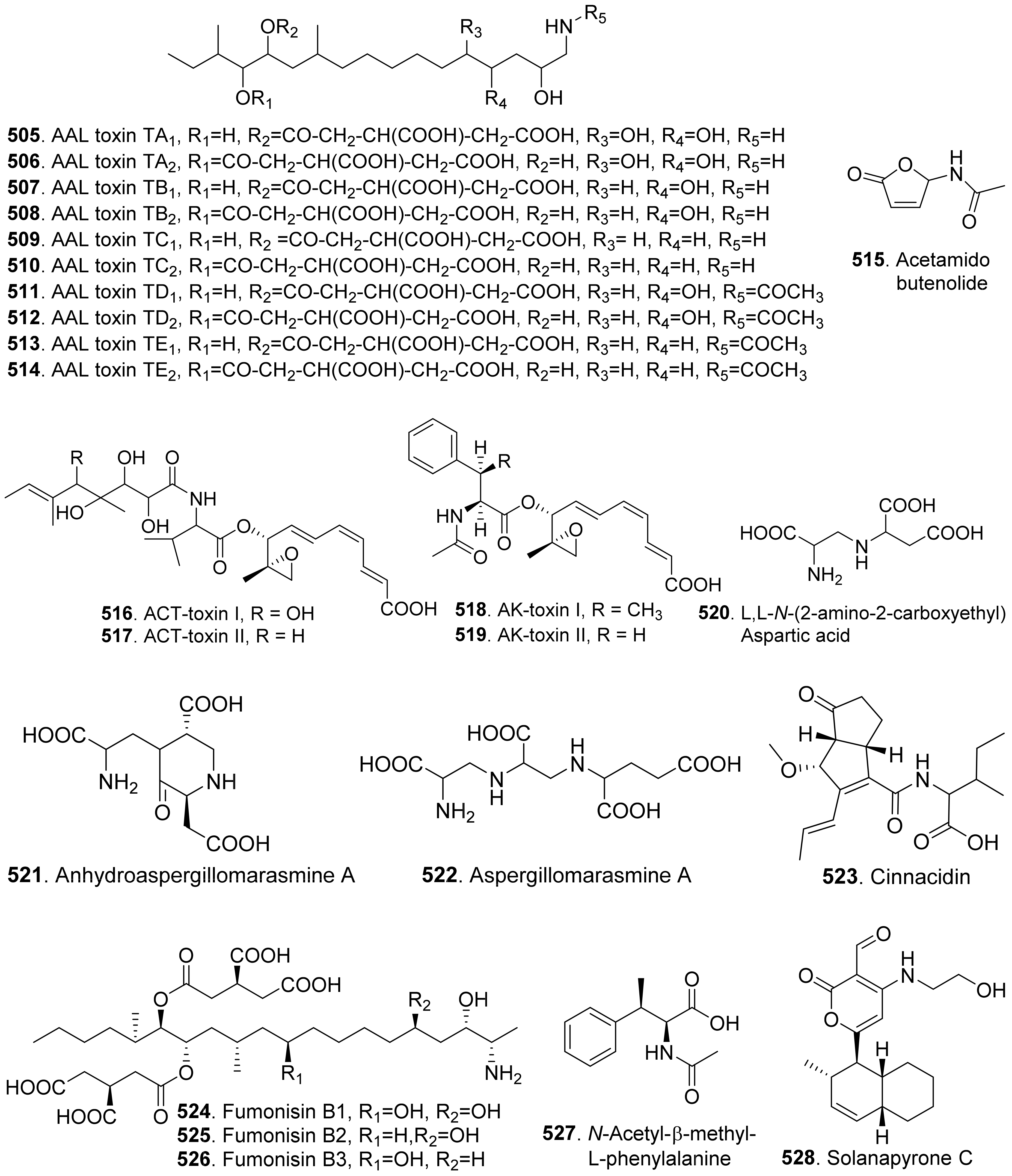
- Ichihara, A.; Tazaki, H.; Sakamura, S. Solanapyrones A, B and C, phytotoxic etabolites from the fungus Alternaria solani. Tetrahedron Lett. 1983, 24, 5373–5376. [Google Scholar] [CrossRef]
- Alam, S.S.; Bilton, J.N.; Slawin, M.Z.; Williams, D.J.; Sheppard, R.N.; Strange, R.N. Chickpea blight: Production of the phytotoxins solanapyrones A and C by Asochyta rabiei. Phytochemistry 1989, 28, 2627–2630. [Google Scholar] [CrossRef]
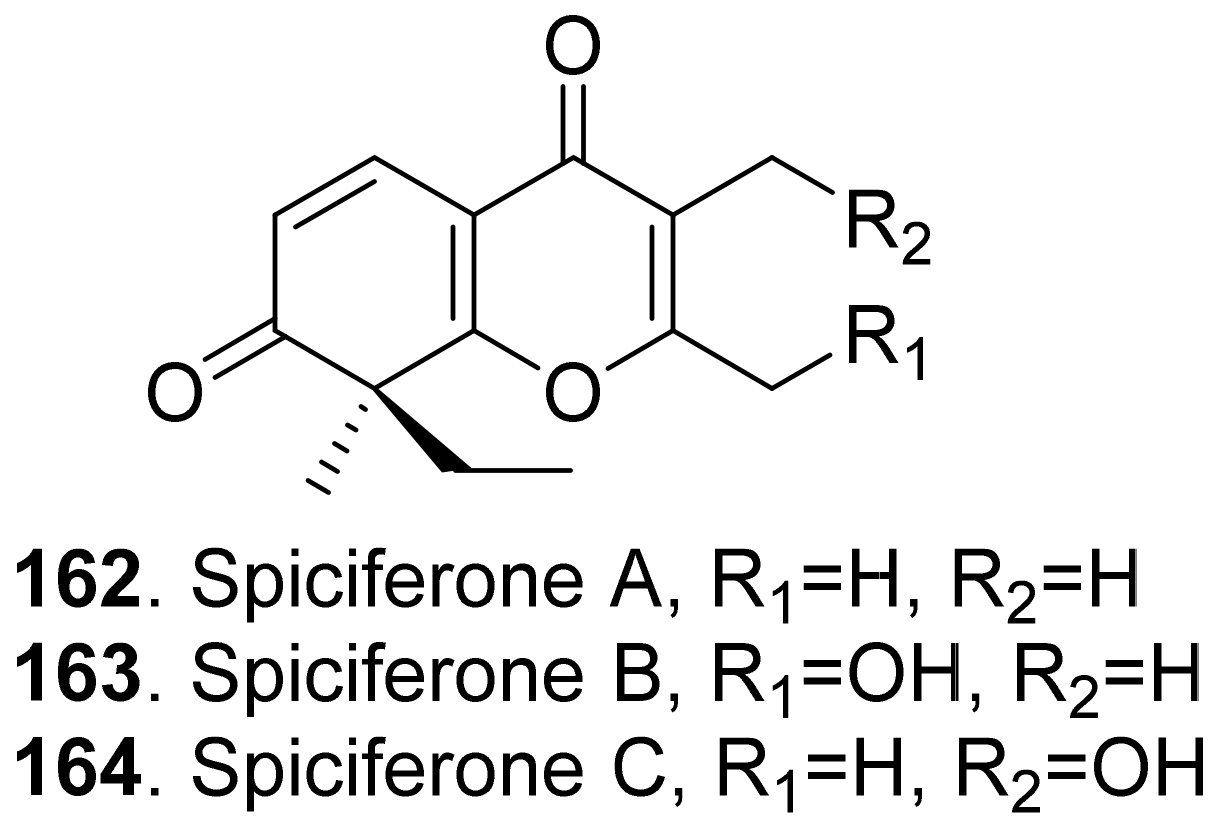
- Nakajima, H.; Hamasaki, T.; Kohno, M.; Kimura, Y. Spiciferones B and C, minor phytotoxins from the fungus Cochliobolus spicifer. Phytochemistry 1991, 30, 2563–2565. [Google Scholar] [CrossRef]
- Nakajima, H.; Fujimoto, H.; Kimura, Y.; Hamasaki, T. Importance of the ketone function for the phytotoxicity of spiciferone A, a phytotoxin produced by the fungus Cochliobolus spicifer. Biosci. Biotechnol. Biochem. 1993, 57, 1938–1939. [Google Scholar] [CrossRef] [Green Version]
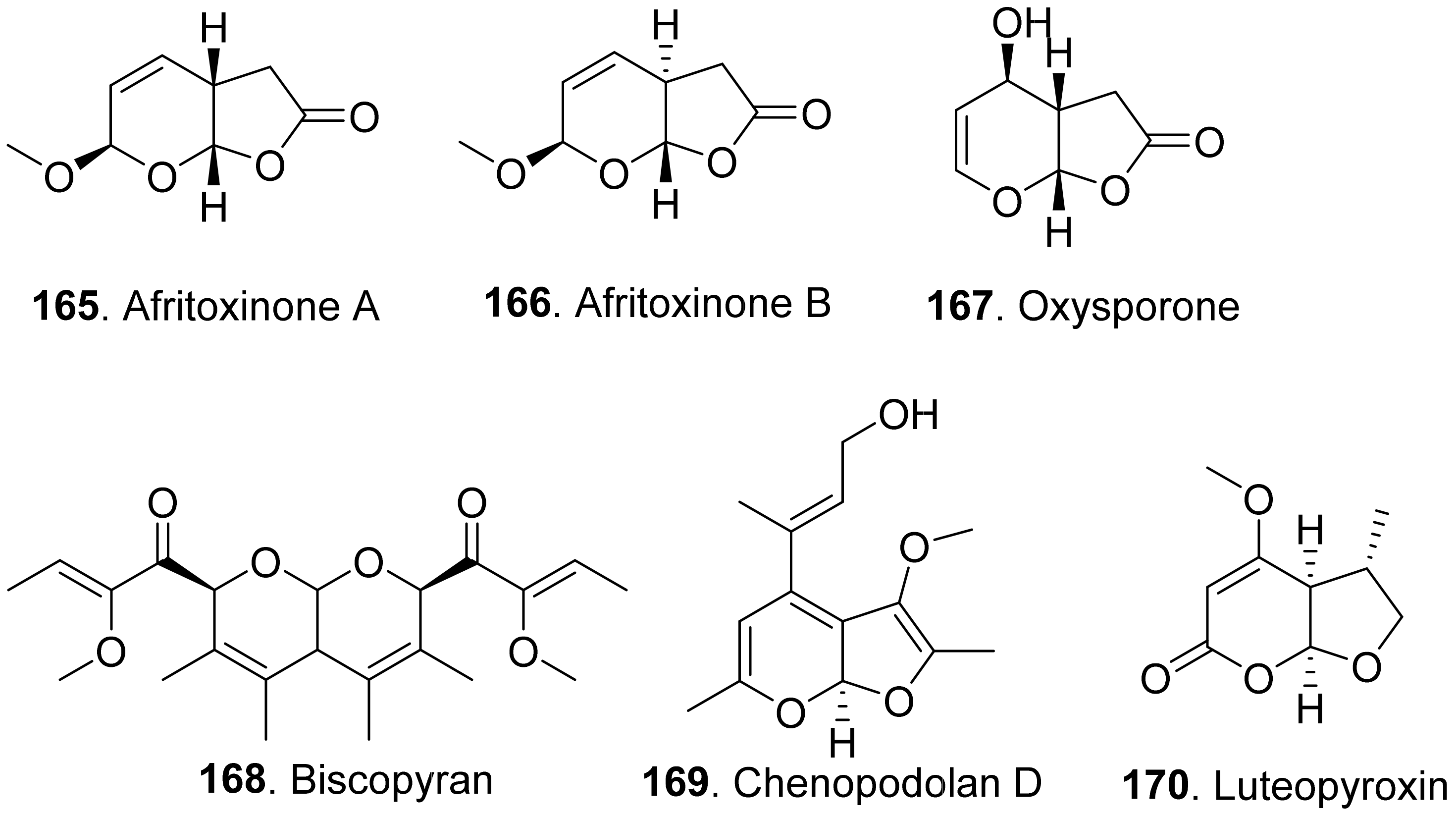
- Evidente, A.; Masi, M.; Linaldeddu, B.T.; Franceschini, A.; Scanu, B.; Cimmino, A.; Andolfi, A.; Motta, A.; Maddau, L. Afritoxinones A and B, dihydrofuropyran-2-ones produced by Diplodia africana the causal agent of branch dieback on Juniperus phoenicea. Phytochemistry 2012, 77, 245–250. [Google Scholar] [CrossRef]
- Evidente, A.; Andolfi, A.; Madau, L.; Franceschini, A.; Marras, F. Biscopyran, a phytotoxic hexasubstituted pyranopyran produced by Biscogniauxia mediterranea, a fungus pathogen of cork oak. J. Nat. Prod. 2005, 68, 568–571. [Google Scholar] [CrossRef] [PubMed]
- Evidente, M.; Cimmino, A.; Zonno, M.C.; Masi, M.; Berestetskyi, A.; Santoro, E.; Superchi, S.; Vurro, M.; Evidente, A. Phytotoxins produced by Phoma chenopodiicola, a fungal pathogen of Chenopodium album. Phytochemistry 2015, 117, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Hirota, A.; Sakai, H.; Isogai, A. New plant growth regulators, cladospolide A and B, macrolides produced by Cladosporium cladosporioides. Agric. Biol. Chem. 1985, 49, 731–735. [Google Scholar] [CrossRef]
- Fujii, Y.; Fukuda, A.; Hamasaki, T.; Ichimoto, I.; Hiromitsu, N. Twelve-membered lactones produced by Cladosporium tenuissimum and the plant growth retardant activity of cladospolide B. Phytochemistry 1995, 40, 1443–1446. [Google Scholar] [CrossRef]
- Yuan, C.; Ding, G.; Wang, H.; Guo, Y.; Ma, X.; Zou, Z. Phytotoxic secondary metabolites from the endolichenic fungus Myxotrichum sp. Chem. Nat. Compt. 2018, 54, 638–641. [Google Scholar] [CrossRef]
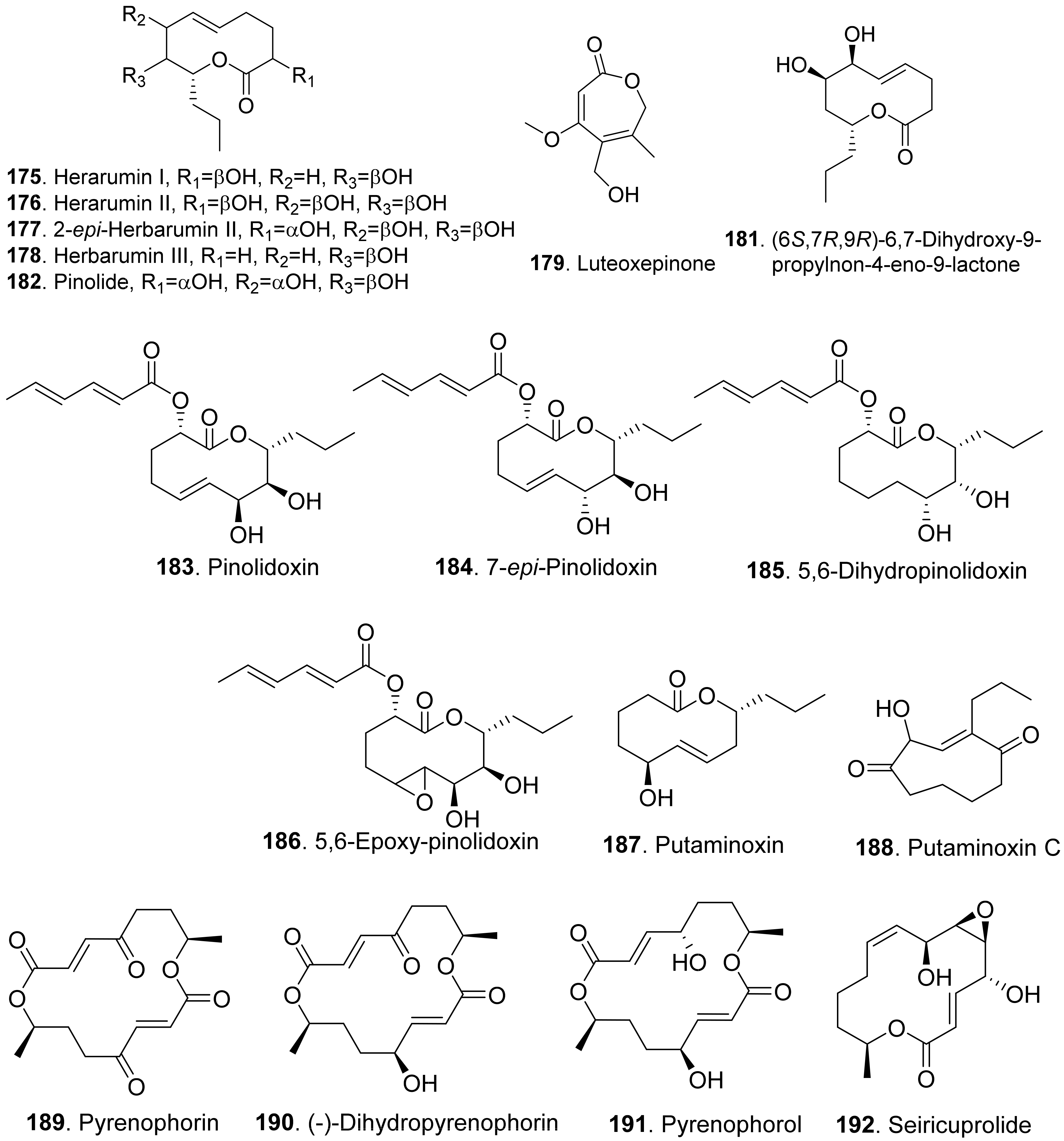
- Rivero-Cruz, J.F.; Garcia-Aguirre, G.; Cerda-Garcia-Rojas, C.M.; Mata, R. Conformational behavior and absolute stereostructure of two phytotoxic nonenolides from the fungus Phoma herbarum. Tetrahedron 2000, 56, 5337–5344. [Google Scholar] [CrossRef]
- Rivero-Cruz, J.F.; Macias, M.; Cerda-Garcia-Rojas, C.M.; Mata, R. A new phytotoxic nonenolide from Phoma herbarum. J. Nat. Prod. 2003, 66, 511–514. [Google Scholar] [CrossRef]
- Cimmino, A.; Andolfi, A.; Fondevilla, S.; Abouzeid, M.A.; Rubiales, D.; Evidente, A. Pinolide, a new noneolide produced by Didymella pinodes, the causal agent of Ascochyta blight on Pisum sativum. J. Agric. Food Chem. 2012, 60, 5273–5278. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Ge, M.; Yin, Y.; Chen, Y.; Luo, M.; Chen, D. A novel phytotoxic nonenolide from Phomopsis sp. HCCB03520. Chem. Biodivers. 2012, 9, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Evidente, A.; Lanzetta, R.; Capasso, R.; Vurro, M.; Bottalico, A. Pinlidoxin, a phytotoxic nonenolide from Ascochyta pinodes. Phytochemistry 1993, 34, 999–1003. [Google Scholar] [CrossRef]
- Evidente, A.; Capasso, R.; Abouzeid, M.A. Three new toxic pinolidoxins from Ascochyta pinodes. J. Nat. Prod. 1993, 56, 1937–1943. [Google Scholar] [CrossRef]
- Evidente, A.; Lanzetta, R.; Capasso, R.; Andolfi, A.; Bottalico, A.; Vurro, M.; Zonno, M.C. Putaminoxin, a phytotoxic nonenolide from Phoma putaminum. Phytochemistry 1995, 40, 1637–1641. [Google Scholar] [CrossRef]
- Evidente, A.; Lanzetta, R.; Capasso, R.; Andolfi, A.; Vurro, M.; Zonno, M.C. Putaminoxins B and C from Phoma putaminum. Phytochemistry 1997, 44, 1041–1045. [Google Scholar] [CrossRef]
- Lerario, P.; Graniti, A. Phytotoxic activity of pyrenophorin and its production in cultures of Pyrenophora avenae Ito et Kurib. Phytoathol. Mediterr. 1985, 24, 280–283. [Google Scholar]
- Sugawara, F.; Strobel, G.A. (−)-Dihydropyrenophorin, a novel and selective phytotoxin produced by Drechslera avenae. Plant Sci. 1986, 43, 1–5. [Google Scholar] [CrossRef]
- Kastanias, M.A.; Chrysayi-Tokousbalides, M. Herbicidal potential of pyrenophorol isolated from a Drechslera avenae pathotype. Pest Manag. Sci. 2000, 56, 227–232. [Google Scholar] [CrossRef]
- Ballio, A.; Evidente, A.; Graniti, A.; Randazzo, G.; Sparapano, L. Phytotoxins produced by species of Seiridium causing canker diseases of cypress. Part 3. Seiricuprolide, a new phytotoxic macrolide from a strain of Seiridium cupressi infecting cypress. Phytochemistry 1988, 27, 3117–3121. [Google Scholar] [CrossRef]
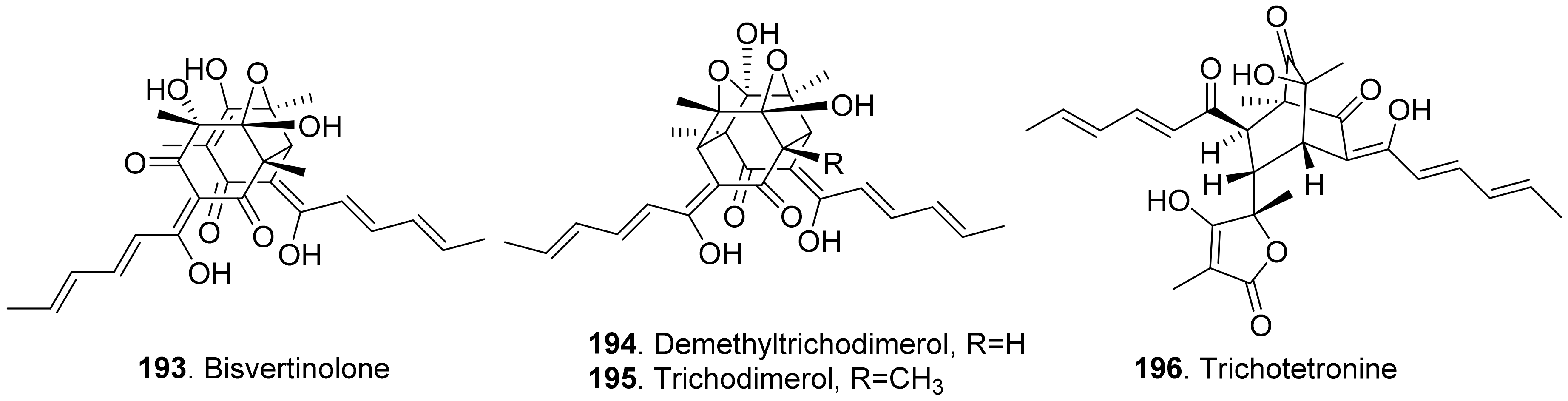
- Harned, A.M.; Volp, K.A. The sorbicillinoid family of natural products: Isolation, biosynthesis and synthetic studies. Nat. Prod. Rep. 2011, 28, 1790–1810. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Wang, X.; Xu, D.; Fu, X.; Zhang, X.; Lai, D.; Zhou, L.; Zhang, G. Sorbicillinoids from fungi and their bioactivities. Molecules 2016, 21, 715. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Gu, G.; Dang, P.; Zhang, X.; Wang, W.; Dai, J.; Liu, Y.; Lai, D.; Zhou, L. Sorbicilinoids from the fungus Ustiaginoidea virens and their phytotoxic, cytotoxic, and antimicrobial activities. Front. Chem. 2019, 7, 435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
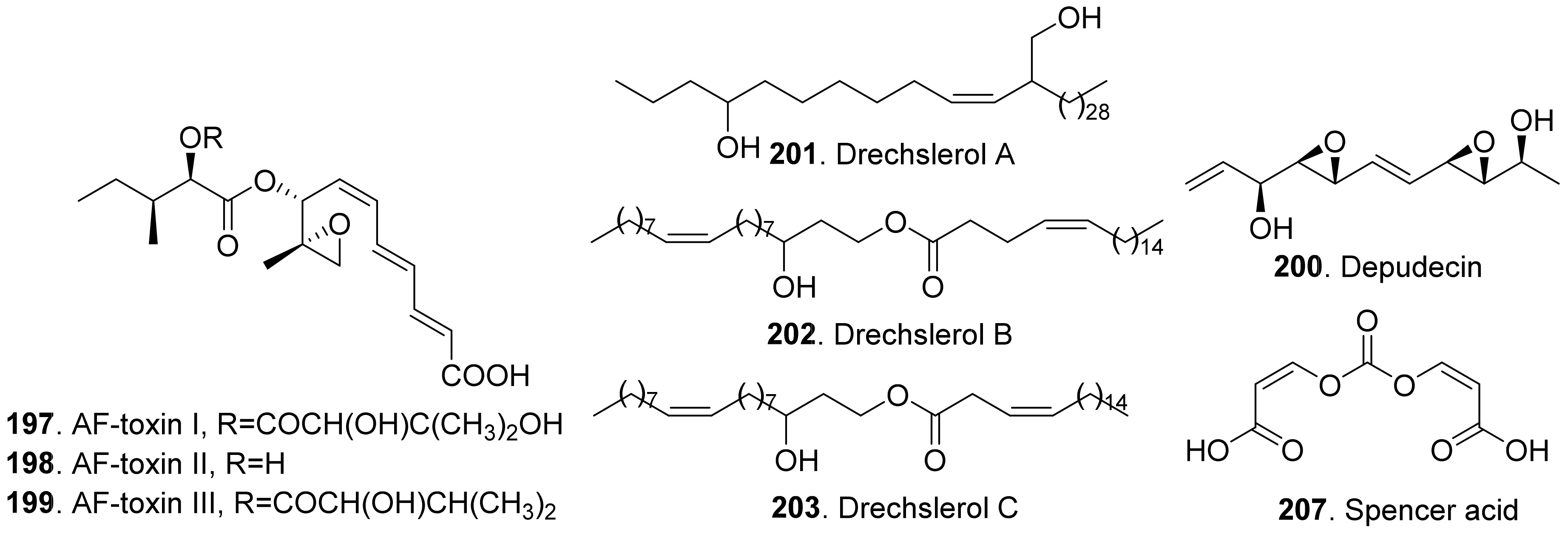
- Maekawa, N.; Yamamoto, M.; Nishimura, S.; Kohmoto, K.; Kuwada, K.; Watanabe, Y. Studies on host-specific AF-toxins produced by Alternaria alternata strawberry pathotype causing Alternaria black spot of strawberry. (1) Production of hostspecific toxins and their biological activities. Ann. Phytopathol. Soc. Jpn. 1984, 50, 600–609. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Ohra, J.; Tsujino, Y.; Sawaji, Y.; Fujimori, T. Phytotoxin produced by Nimbya scirpicola. Biosci. Biotechnol. Biochem. 1994, 58, 565–566. [Google Scholar] [CrossRef] [Green Version]
- Shukla, R.S.; Agrawal, P.K.; Husain, A. Drechslerol A, a new phytotoxic metabolite produced by Drechslera maydis, a strain from Costus speciosus. Plant Sci. 1987, 48, 159–163. [Google Scholar] [CrossRef]
- Shukla, R.S.; Agrawal, P.K.; Thakur, R.S.; Husain, A. Drechslerol B, a host-selective phytotoxin produced by Drechslera maydis. Phytochemistry 1989, 28, 2089–2091. [Google Scholar] [CrossRef]
- Shukla, R.S.; Agrawal, P.K.; Thakur, R.S.; Husain, A. Drechslerol C, a phytotoxin produced by Drechslera maydis, the causal organism of leaf blight of Costus speciosus. Plant Sci. 1990, 66, 43–49. [Google Scholar] [CrossRef]
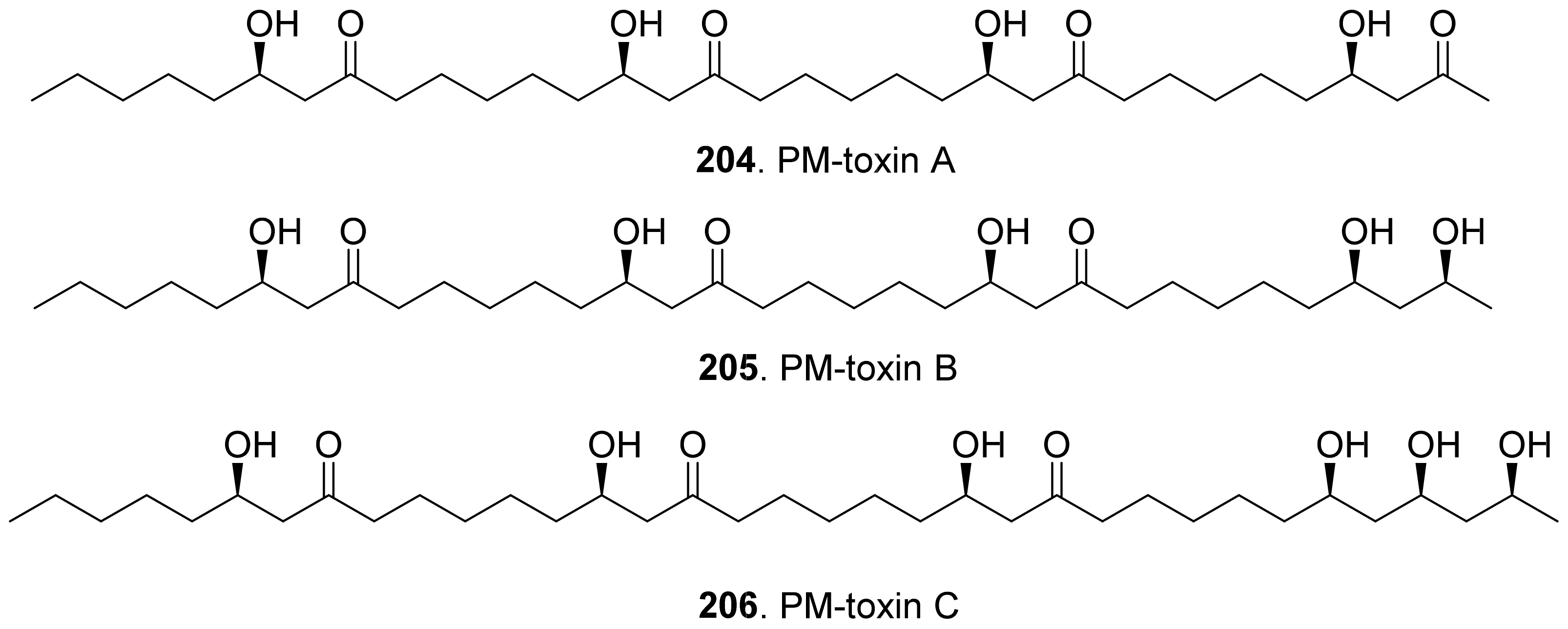
- Danko, S.J.; Kono, Y.; Daly, J.M.; Suzuki, Y.; Takeuchi, S.; McCrery, D.A. Structure and biological activity of a host-specific toxin produced by the fungal corn pathogen Phyllosticta maydis. Biochemistry 1984, 23, 759–766. [Google Scholar] [CrossRef]
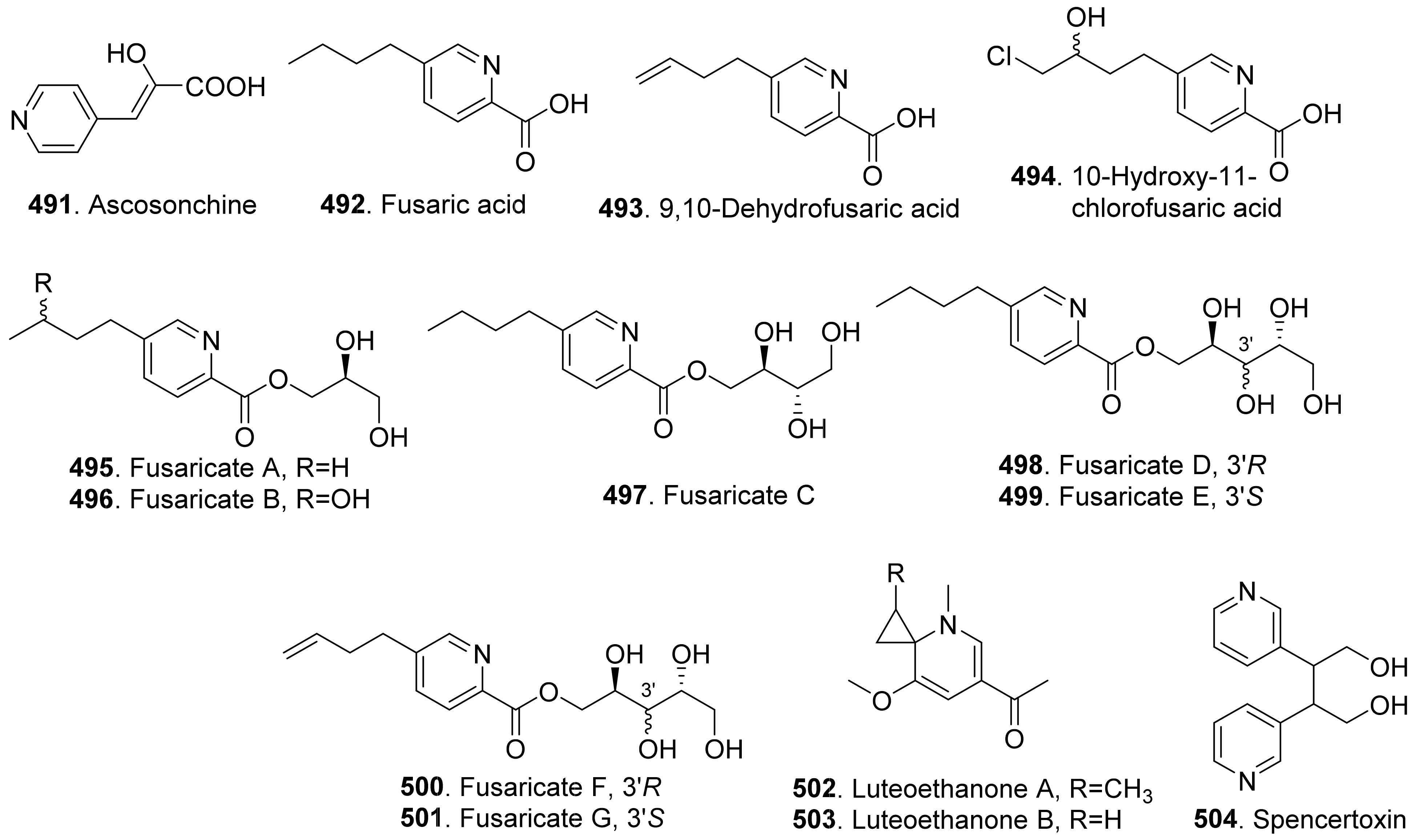
- Reveglia, P.; Savocchia, S.; Billones-Baaijens, R.; Masi, M.; Evidente, A. Spencertoxin and spencer acid, new phytotoxic derivatives of diacrylic acid and dipyridinbutan-1,4-diol produced by Spencermartinsia viticola, a causal agent of grapevine Botryosphaeria dieback in Australia. Arab. J. Chem. 2020, 13, 1803–1808. [Google Scholar] [CrossRef]
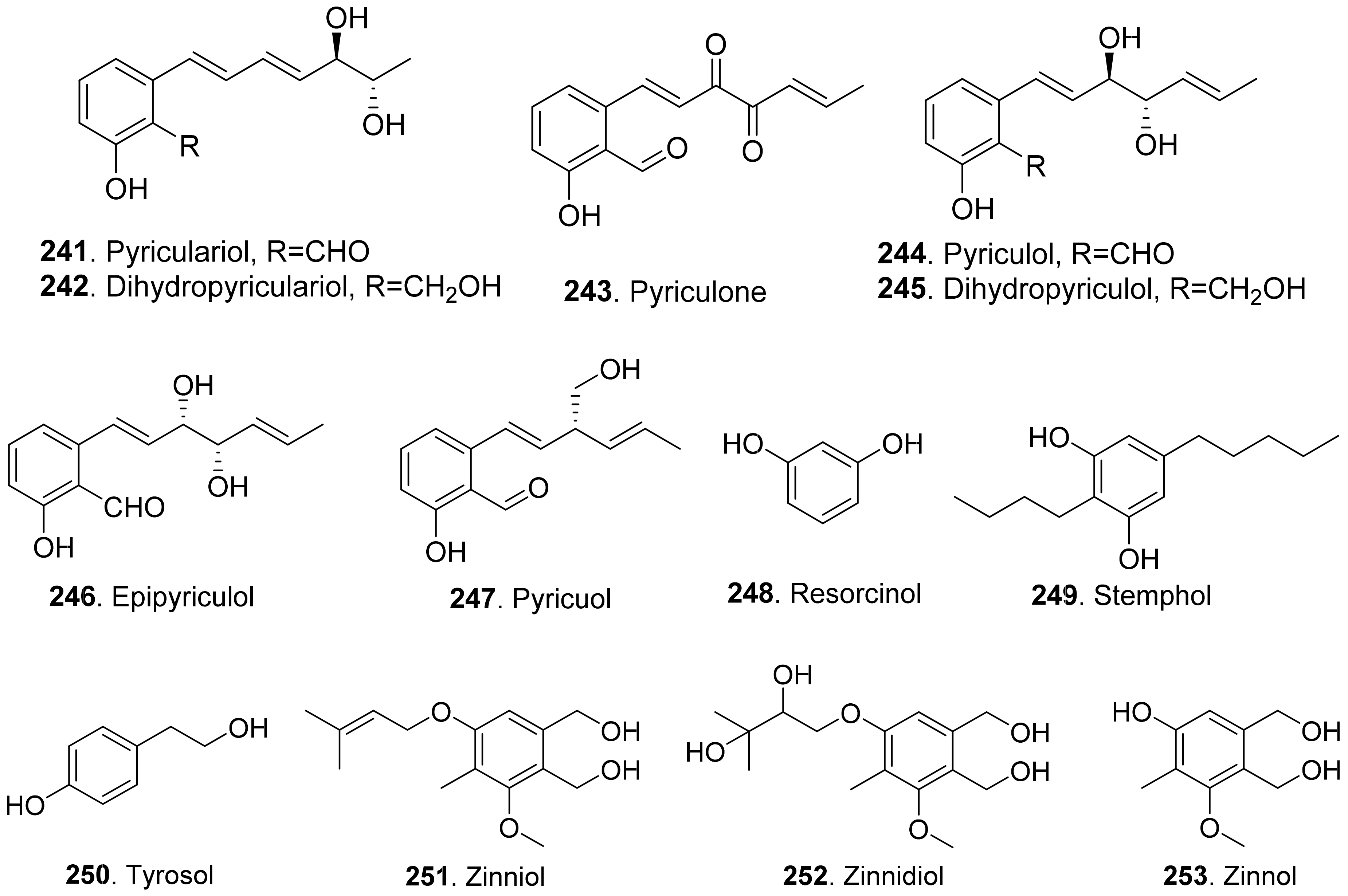
- Jacob, S.; Groetsch, T.; Foster, A.J.; Schueffler, A.; Rieger, P.H.; Sandjo, L.P.; Lirmann, J.C.; Opatz, T.; Thines, E. Unravelling the biosynthesis of pyriculol in the rice blast fungus Magnaporthe oryzae. Microbiology 2017, 163, 541–553. [Google Scholar] [CrossRef] [PubMed]
- Mafezoli, J.; Xu, Y.; Hilario, F.; Fredhof, B.; Espinosa-Artiles, P.; Dos Santos, L.C.; De Oliveira, M.C.F.; Gunatilaka, A.A.L. Modulation of polyketide biosynthetic pathway of the endophytic fungus Anteaglonium sp. FL0768, by copper (II) and anacardic acid. Phytochem. Lett. 2018, 28, 157–163. [Google Scholar] [CrossRef]
- Evidente, A.; Capasso, R.; Vurro, M.; Bottalico, A. Ascosalitoxin, a phytotoxic trisubstituted salicylic aldehyde from Ascochyta pisi. Phytochemistry 1993, 34, 995–998. [Google Scholar] [CrossRef]
- Pedras, M.S.C.; Yu, Y. Phytotoxins, elicitors and other secondary metabolites from phytopathogenic “blackleg” fungi: Structure, phytotoxicity and biosynthesis. Nat. Prod. Commun. 2009, 4, 1291–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reveglia, P.; Savocchia, S.; Billones-Baaijens, R.; Cimmino, A.; Evidente, A. Isolation of phytotoxic phenols and characterization of a new 5-hydroxymethyl-2-isopropoxyphenol from Dothiorella vidmadera, a causal agent of grapevine trunk disease. J. Agric. Food Chem. 2018, 66, 1760–1764. [Google Scholar] [CrossRef]
- Leyte-Lugo, M.; Richomme, P.; Poupard, P.; Pena-Rodriguez, L.M. Identification and quantification of a phytotoxic metabolite from Alternaria dauci. Molecules 2020, 25, 4003. [Google Scholar] [CrossRef] [PubMed]
- Kono, Y.; Sekido, S.; Yamaguchi, I.; Kondo, H.; Suzuki, Y.; Neto, G.C.; Sakurai, A.; Yaegashi, H. Structures of two novel pyriculol-related compounds and identification of naturally produced epipyriculol from Pyricularia oryzae. Agric. Biol. Chem. 1991, 55, 2785–2791. [Google Scholar] [CrossRef] [Green Version]
- Evidente, A.; Randazzo, G.; Iacobellis, N.S.; Bottalico, A. Structure of cavoxin, a new phytotoxin from Phoma cava and cavoxone, its related chroman-4-one. J. Nat. Prod. 1985, 48, 916–923. [Google Scholar] [CrossRef]
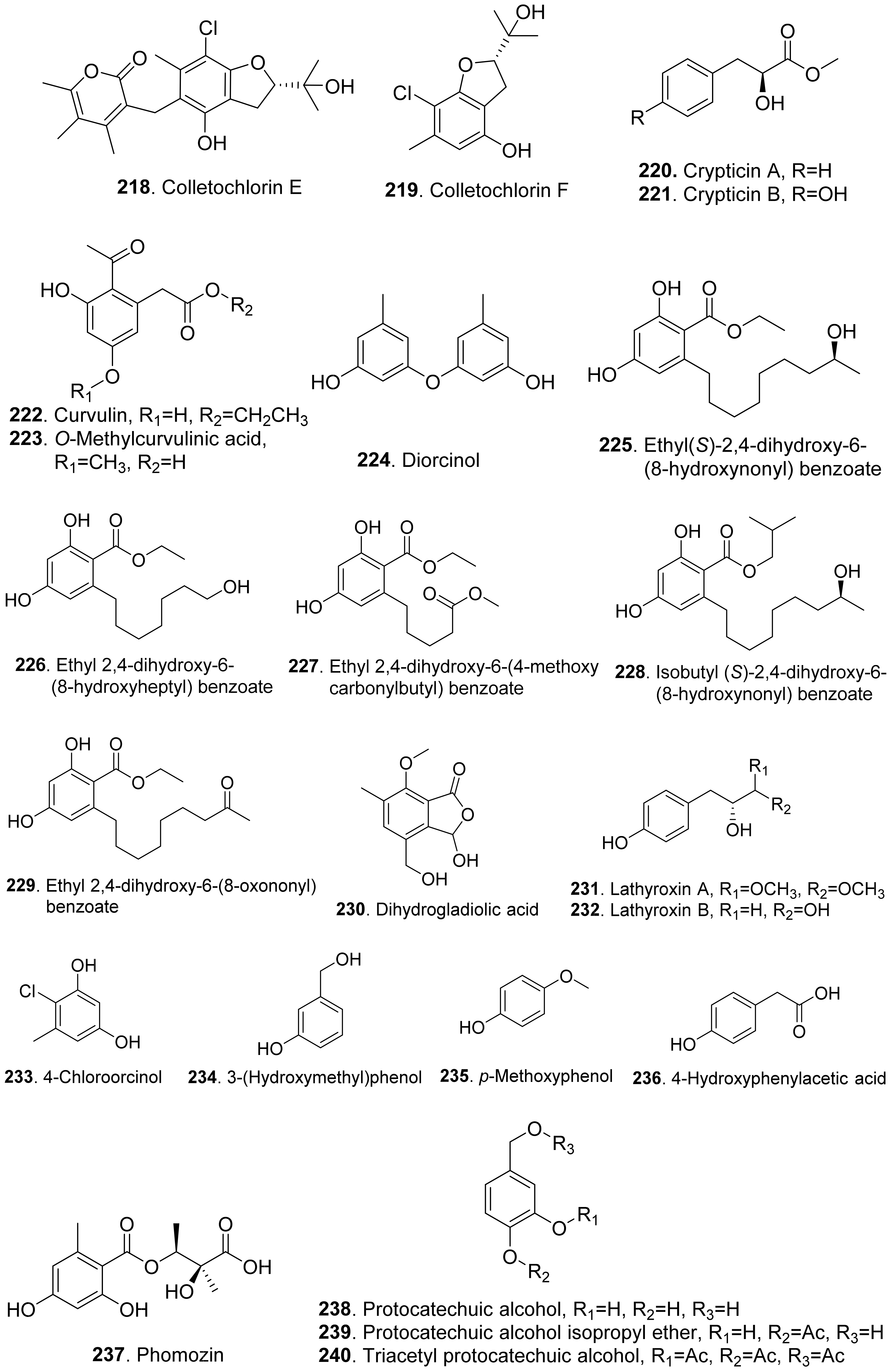
- Masi, M.; Cimmino, A.; Boari, A.; Tuzi, A.; Zonno, M.C.; Baroncelli, R.; Vurro, M.; Evidente, A. Colletochlorins E and F, new phytotoxic tetrasubstituted pyran-2-one and dihydrobenzofuran, isolated from Colletotrichum higginsianum with potential herbicidal activity. J. Agric. Food Chem. 2017, 65, 1124–1130. [Google Scholar] [CrossRef]
- Cimmino, A.; Nocera, P.; Linaldeddu, B.T.; Masi, M.; Gorecki, M.; Pescitelli, G.; Montecchio, L.; Maddau, L.; Evidente, A. Phytotoxic metabolites produced by Diaporthella cryptica, the causal agent of hazelnut branch canker. J. Agric. Food Chem. 2018, 66, 3435–3442. [Google Scholar] [CrossRef] [PubMed]
- Kenfield, D.; Hallock, Y.; Clardy, J.; Strobel, G. Curvulin and O-methylcurvulinic acid: Phytotoxic metabolites of Drechslera indica which cause necroses on purslane and spiny amaranth. Plant Sci. 1989, 60, 123–127. [Google Scholar] [CrossRef]
- Cimmino, A.; Maddau, L.; Masi, M.; Evidente, M.; Linaldeddu, B.T.; Evidente, A. Further secondary metabolites produced by Diplodia corticola, a fungal pathogen involved in cork oak decline. Tetrahedron 2016, 72, 6788–6793. [Google Scholar] [CrossRef]
- Song, Q.-Y.; Nan, Z.-B.; Gao, K.; Song, H.; Tian, P.; Zhang, X.-X.; Li, C.-J.; Xu, W.-B.; Li, X.-Z. Antifungal, phytotoxic, and cytotoxic activities of metabolites from Epichloë bromicola, a fungus obtained from Elymus tangutorum grass. J. Agric. Food Chem. 2015, 63, 8787–8792. [Google Scholar] [CrossRef] [PubMed]
- Ichihara, A.; Sawamura, S.; Kawakami, Y.; Sakamura, S. Dihydrogladiolic acid, another phytotoxin from Phoma asparagi Sacc. Agric. Biol. Chem. 1985, 49, 1891–1892. [Google Scholar]
- Masi, M.; Nocera, P.; Boari, A.; Cimmino, A.; Zonno, M.C.; Infantino, A.; Vurro, M.; Evidente, A. Lathyroxins A and B, phytotoxic monosubstituted phenols isolated from Ascochyta lentis var. lathyri, a fungal pathogen of grass pea (Lathyrus sativus). J. Nat. Prod. 2018, 81, 1093–1097. [Google Scholar] [CrossRef] [PubMed]
- Mazars, C.; Rossignol, M.; Auriol, P.; Klaebe, A. Phomozin, a phytotoxin from Phomopsis helianthi, the causal agent of stem canker of sunflower. Phytochemistry 1990, 29, 3441–3444. [Google Scholar] [CrossRef]
- Iwasaki, S.; Muro, H.; Sasaki, K.; Nozoe, S.; Okuda, S.; Sato, Z. Isolations of phytotoxic substances produced by Pyricularia oryzae Cavara. Tetrahedron Lett. 1973, 14, 3537–3542. [Google Scholar] [CrossRef]
- Tanaka, K.; Nakamura, Y.; Sasaki, A.; Ueda, R.; Suzuki, Y.; Kuwahara, S.; Kiyota, H. Synthesis and plant growth inhibitory activity of both enantiomers of pyricuol, a phytotoxin isolated from rive blast disease fungus Magnaporthe grisea. Tetrahedron 2009, 65, 6115–6122. [Google Scholar] [CrossRef]
- Kim, J.-C.; Min, J.-Y.; Kim, H.-T.; Cho, K.-Y.; Yu, S.-H. Pyricuol, a new phytotoxin from Magnaporthe grisea. Biosci. Biotechnol. Biochem. 1998, 62, 173–174. [Google Scholar] [CrossRef] [PubMed]
- Motoyama, T. Secondary metabolites of the rice blast fungus Pyricularia oryzae: Biosynthesis and biological function. Int. J. Mol. Sci. 2020, 21, 8691. [Google Scholar] [CrossRef]
- Solfrizzo, M.; Strange, R.N.; Sabia, C.; Visconti, A. Production of a toxin stemphol by Stemphylium species. Nat. Toxins 1994, 2, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Stierle, A.; Hershenhorn, J.; Strobel, G. Zinniol-related phytotoxins from Alternaria cichorii. Phytochemistry 1993, 32, 1145–1149. [Google Scholar] [CrossRef]
- Ulloa-Benitez, A.; Medina-Romero, Y.M.; Sanchez-Fernandez, R.E.; Lappe-Oliveras, P.; Roque-Flores, G.; Duarte Lisci, G.; Herrera Suarez, T.; Macias-Rubalcava, M.L. Phytotoxic and antimicrobial activity of volatile and semi-volatile organic compounds from the endophyte Hypoxylon anthochroum strain Blaci isolated from Bursera lancifolia (Burseraceae). J. Appl. Microbiol. 2016, 121, 380–400. [Google Scholar] [CrossRef]
- Ichihara, A.; Sawamura, S.; Sakamura, S. Structures of altiloxins A and B, phytotoxins from Phoma asparagi Sacc. Tetrahedron Lett. 1984, 25, 3209–3212. [Google Scholar] [CrossRef]
- Shimada, A.; Kusano, M.; Takeuchi, S.; Fujioka, S.; Inokuchi, T.; Kimura, Y. Asterric acid and 6-hydroxymellein, inhibitors of pollen development in Arabidopsis thaliana, produced by Aspergillus terreus. Z. Naturforsch. C 2002, 57, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Liu, Q.; Zhang, X.; Yuan, S.; Bat-Erdene, U.; Nguyen, C.; Gan, J.; Zhou, J.; Jacobsen, S.E.; Tang, Y. Resistance-gene-directed discovery of a natural-product herbicide with a new mode of action. Nature 2018, 559, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, F.; Strobel, G.; Fisher, L.E.; Van Duyne, G.D.; Clardy, J. Bipolaroxin, a selective phytotoxin produced by Bipolaris cynodontis. Proc. Natl. Acad. Sci. USA 1985, 82, 8291–8294. [Google Scholar] [CrossRef] [Green Version]
- Jahani, M.; Aggarwal, R.; Gupta, S.; Sharma, S.; Dureja, P. Purification and characterization of a novel toxin from Bipolaris sorokiniana, causing spot blotch of wheat and analysis of variability in the pathogen. Cereal Res. Commun. 2014, 42, 252–261. [Google Scholar] [CrossRef]
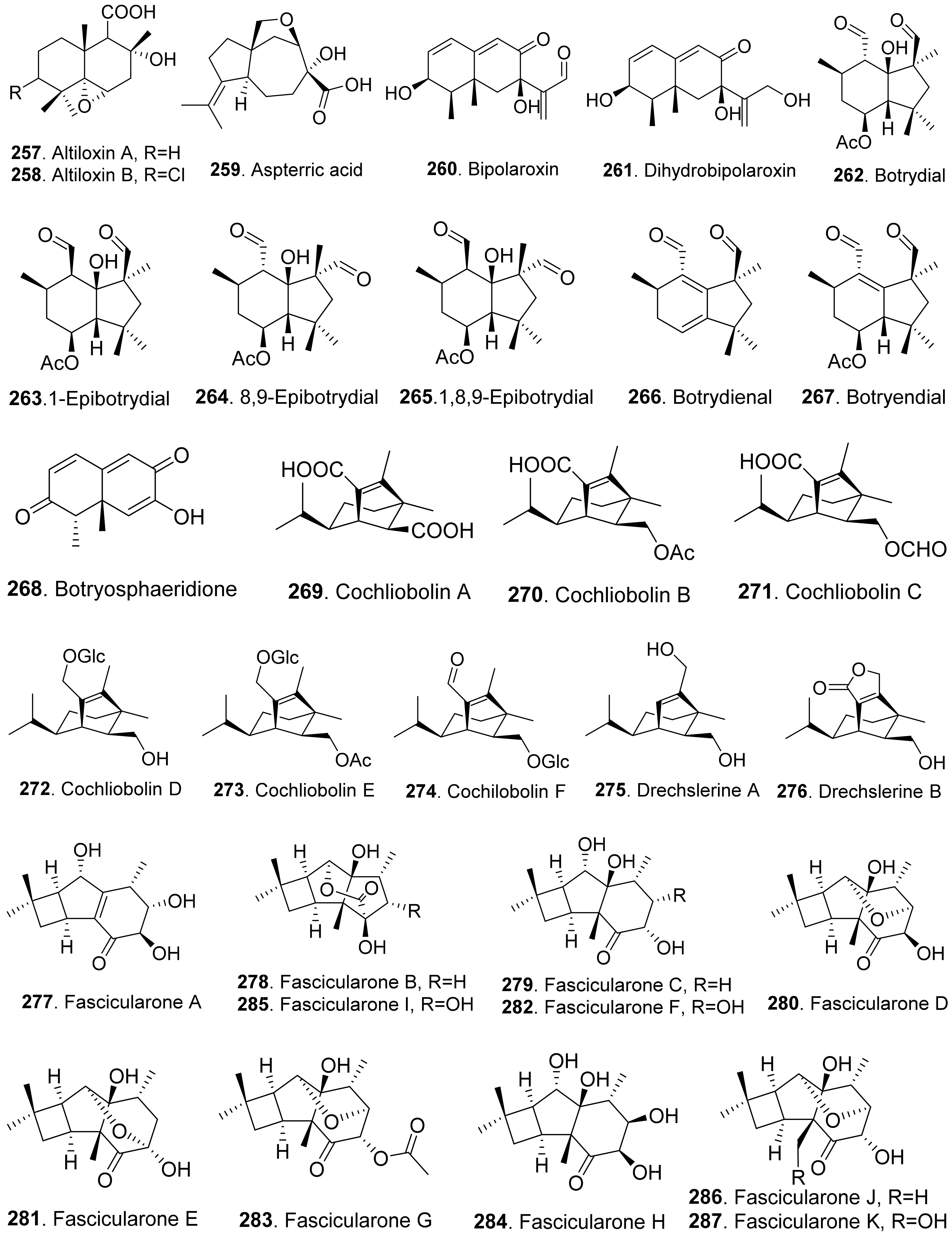
- Duran-Patron, R.; Hernandez-Galan, R.; Rebordinos, L.G.; Cantoral, J.M.; Collado, I.G. Structure-activity relationships of new phytotoxic metabolites with the botryane skeleton from Botrytis cinerea. Tetrahedron 1999, 55, 2389–2400. [Google Scholar] [CrossRef]
- D’Ambrosio, J.M.; Gonorazky, G.; Sueldo, D.J.; Moraga, J.; Di Palma, A.A.; Lamattina, L.; Collado, I.G.; Laxalt, A.M. The sesquiterpene botrydial from Botrytis cinerea induces phosphatidic acid production in tomato cell suspensions. Planta 2018, 247, 1001–1009. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, S.-Q.; Li, X.-J.; Zhang, A.-L.; Zhang, Q.; Gao, J.-M. New insight into the stereochemistry of botryosphaeridione from a Phoma endophyte. J. Mol. Struct. 2012, 1016, 72–75. [Google Scholar] [CrossRef]
- Qader, M.M.; Kumar, N.S.; Jayasinghe, L.; Araya, H.; Fujimoto, Y. Bioactive sesquiterpenes from an endophytic fungus Bipolaris sorokiniana isolated from a popular medicinal plant Costus speciosus. Mycology 2017, 8, 17–20. [Google Scholar] [CrossRef] [Green Version]
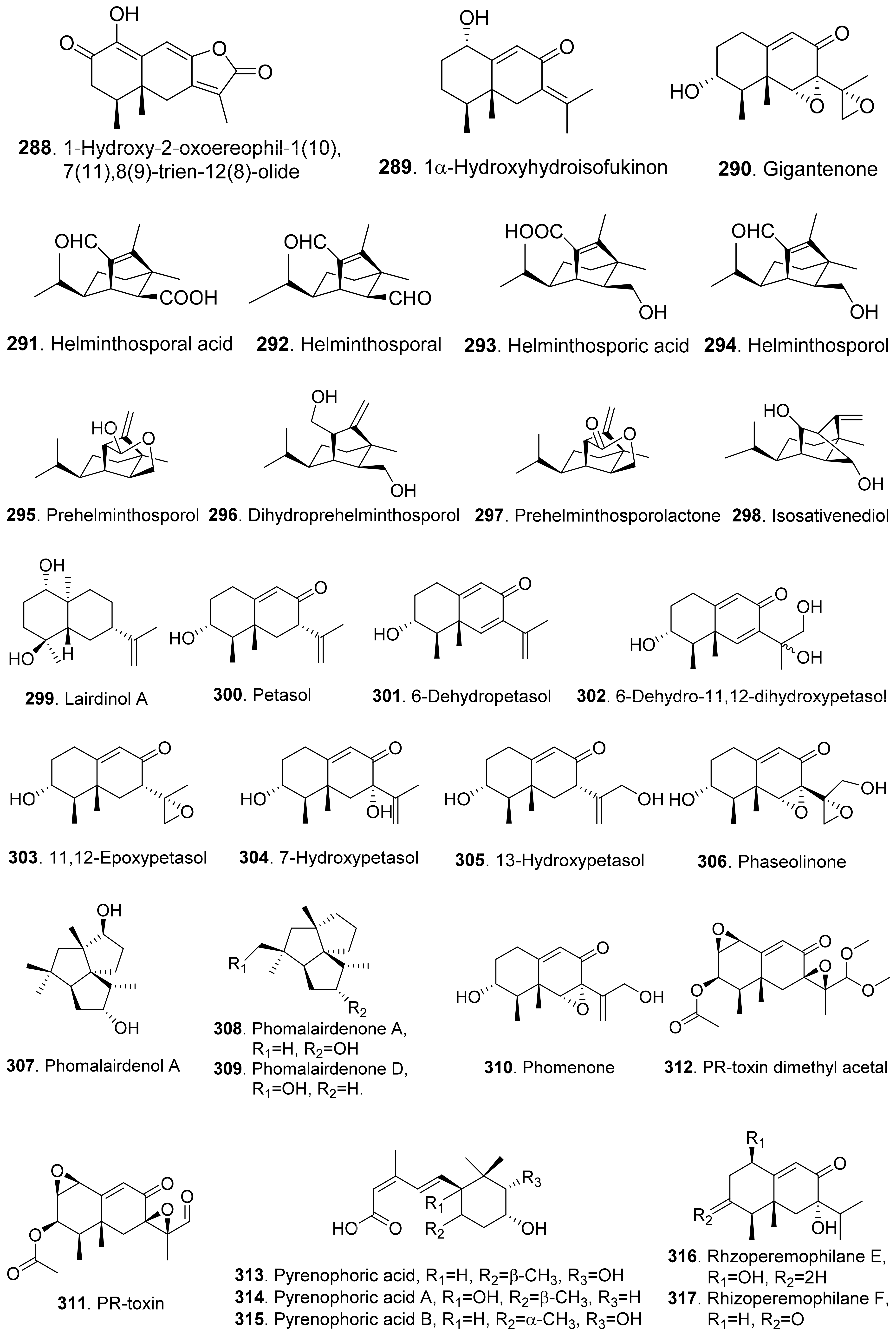
- Taniguchi, E.; White, G.A. The mode of action of helminthosporal. II. Effect on the permeability of plant cell membrane. Can. J. Bot. 1972, 50, 1415–1420. [Google Scholar]
- Shiono, Y.; Matsuzaka, R.; Wakamatsu, H.; Muneta, K.; Murayama, T.; Ikeda, M. Fascicularones A and B from a mycelial culture of Naematoloma fasciculare. Phytochemistry 2004, 65, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Shiono, Y.; Wakamatsu, H.; Murayama, T.; Ikeda, M. Fascicularones C and D, tricycle[5.4.0.02.5]undecane sesquiterpenoids from the liquid culture of Naematolama fasciculare. Z. Naturforsch. B Chem. Sci. 2004, 59, 119–123. [Google Scholar] [CrossRef]
- Shiono, Y.; Akasaka, H.; Hiramatsu, F.; Sato, K.; Murayama, T.; Ikeda, M. Three sesquiterpenoids, fascicularones E, F, and G produced by the fungus Hypholoma fasciculare. Z. Naturforsch. B Chem. Sci. 2005, 60, 880–884. [Google Scholar] [CrossRef]
- Akasaka, H.; Shiono, Y.; Murayama, T.; Ikeda, M. Fascicularones H-K, four new sesquiterpenoids from the cultured mycelia of the fungus Hypholoma fasciculare. Helv. Chim. Acta 2005, 88, 2944–2950. [Google Scholar] [CrossRef]
- Cutler, H.; Crumley, F.G.; Cox, R.H.; Davis, E.E.; Harper, J.L.; Cole, R.J.; Sumner, D.R. Prehelminthosporol and prehelminthosporol acetate: Plant growth regulating properties. J. Agric. Food Chem. 1982, 30, 658–662. [Google Scholar] [CrossRef]
- Pena-Rodriguez, L.M.; Armingeon, N.A.; Chilton, W.S. Toxins from weed pathogens, I. Phytotoxins from a Bipolaris pathogen of Johnson grass. J. Nat. Prod. 1988, 51, 821–828. [Google Scholar] [CrossRef] [PubMed]
- Pena-Rodriguez, L.M.; Chilton, W.S. Victoxinine and prehelminthosporolactone, two minor phytotoxic metabolites produced by Bipolaris sp. a pathogen of Johnson grass. J. Nat. Prod. 1989, 52, 899–901. [Google Scholar]
- Sugawara, F.; Hallock, Y.F.; Bunkers, G.D.; Kenfield, D.S.; Strobel, G.; Yoshida, S. Phytotoxic eremophilanes produced by the weed pathogen Drechslera gigantean. Biosci. Biotech. Biochem. 1993, 57, 236–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, D.; Siddiqui, K.A.; Ali, E. Phytotoxic metabolites of Macrophomina phaseolina. Indian J. Mycol. Plant Pathol. 1992, 22, 54–57. [Google Scholar]
- Pedras, M.S.C.; Erosa-Lopez, C.C.; Quail, J.W.; Taylor, J.L. Phomalairdenone: A new host-selective phytotoxin from a virulent type of the blackleg fungus Phoma lingam. Bioorg. Med. Chem. Lett. 1999, 9, 3291–3294. [Google Scholar] [CrossRef]
- Pedras, M.S.C.; Chumala, P.B.; Venkatesham, U. New sesquiterpenic phytotoxins establish unprecedented relationship between different groups of blackleg fungal isolates. Bioorg. Med. Chem. 2005, 13, 2469–2475. [Google Scholar] [CrossRef]
- Capasso, R.; Iacobellis, N.S.; Bottalico, A.; Randazzo, G. Structure-toxicity relationships of the eremophilane phomenone and PR-toxin. Phytochemistry 1984, 23, 2781–2784. [Google Scholar] [CrossRef]
- Masi, M.; Meyer, S.; Cimmino, A.; Andolfi, A.; Evidente, A. Pyrenophoric acid, a phytotoxic sesquiterpenoid penta-2,4-dienoic acid produced by a potential mycoherbicide, Pyrenophora semeniperda. J. Nat. Prod. 2014, 77, 925–930. [Google Scholar] [CrossRef]
- Masi, M.; Meyer, S.; Cimmino, A.; Clement, S.; Black, B.; Evidente, A. Pyrenophoric acids B and C, two new phytotoxic sesquiterpenoids produced by Pyrenophora semeniperda. J. Agric. Food Chem. 2014, 62, 10304–10311. [Google Scholar] [CrossRef]
- Lozano-Juste, J.; Masi, M.; Cimmino, A.; Clement, S.; Fernandez, M.A.; Antoni, R. The fungal sesquiterpenoid pyrenophoric acid B uses the plant ABA biosynthetic pathway to inhibit seed germination. J. Exp. Bot. 2019, 70, 5487–5494. [Google Scholar] [CrossRef] [Green Version]
- Wang, A.; Yin, R.; Zhou, Z.; Gu, G.; Dai, J.; Lai, D.; Zhou, L. Eremophilane-type sesquiterpenoids from the endophytic fungus Rhizopycnis vagum and their antibacterial, cytotoxic, and phytotoxic activities. Front. Chem. 2020, 8, 596889. [Google Scholar] [CrossRef] [PubMed]
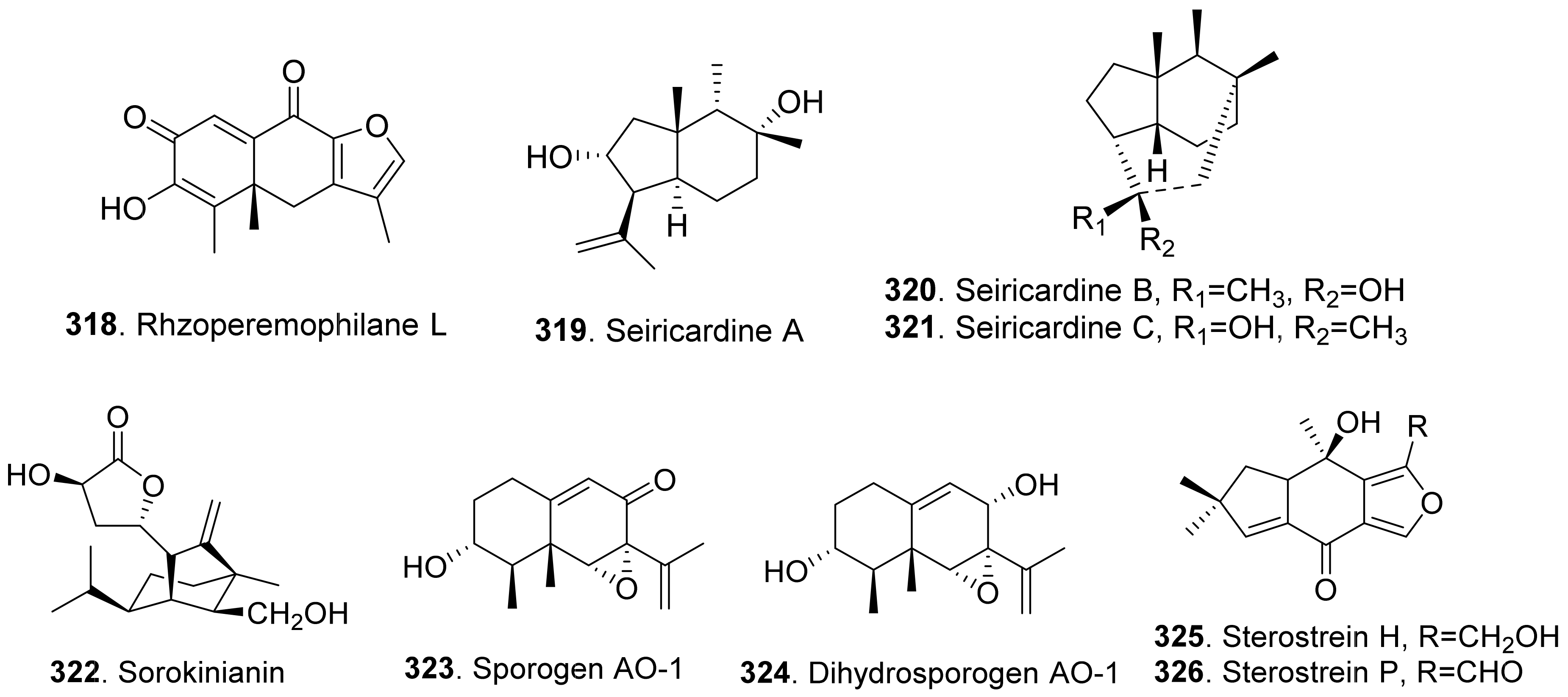
- Ballio, A.; Morelli, M.A.C.; Evidente, A.; Graniti, A.; Randazzo, G.; Sparapano, L. Seiricardine A, a phytotoxic sesquiterpene from three Seiridium species pathogenic for cypress. Phytochemistry 1991, 30, 131–136. [Google Scholar] [CrossRef]
- Evidente, A.; Motta, A.; Sparapano, L. Seicardines B and C, phytotoxic sesquiterpenes from three species of Seiridium pathogenic for cypress. Phytochemistry 1993, 33, 69–78. [Google Scholar] [CrossRef]
- Nakajima, H.; Isomi, K.; Hamasaki, T.; Ichinoe, M. Sorokinianin: A novel phytotoxin produced by the phytopathogenic fungus Bipolaris sorokiniana. Tetrahedron Lett. 1994, 35, 9597–9600. [Google Scholar] [CrossRef]
- Del Valle, P.; Figueroa, M.; Mata, R. Phytotoxic eremophilane sesquiterpenes from the coprophilous fungus Penicillium sp. G1-a14. J. Nat. Prod. 2015, 78, 339–342. [Google Scholar] [CrossRef] [PubMed]
- Schneider, G.; Anke, H.; Sterner, O. New secondary metabolites from a mycophilic Hansfordia species. Nat. Prod. Lett. 1997, 10, 133–138. [Google Scholar] [CrossRef]
- Perera, W.H.; Meepagala, K.M.; Wedge, D.E.; Duke, S.O. Sesquiterpenoids from culture of the fungus Stereum complicatum (Steraceae): Structural diversity, antifungal and phytotoxic activities. Phytochem. Lett. 2020, 37, 51–58. [Google Scholar] [CrossRef]
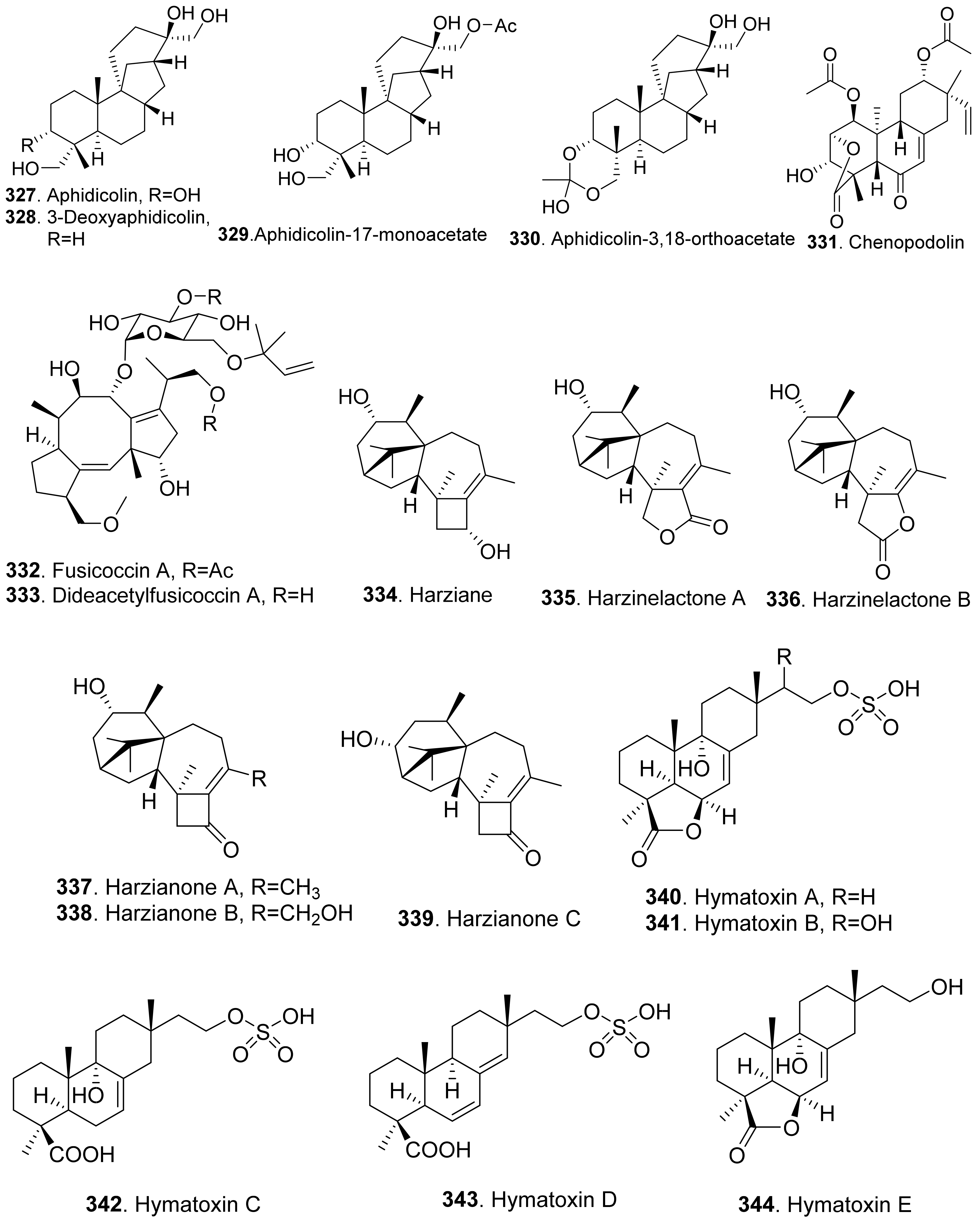
- Ichihara, A.; Oikawa, H.; Hayashi, K.; Hashimoto, M.; Sakamura, S.; Sakai, R. 3-Deoxyaphidicolin and aphidicolin analogues as phytotoxins from Phoma betae. Agric. Biol. Chem. 1984, 48, 1687–1689. [Google Scholar] [CrossRef] [Green Version]
- Cimmino, A.; Andolfi, A.; Zonno, M.C.; Avolio, F.; Santini, A.; Tuzi, A.; Berestetskyi, A.; Vurro, M.; Evidente, A. Chenopodolin: A phytotoxic unrearranged ent-pimaradiene diterpene produced by Phoma chenopodicola, a fungal pathogen for Chenopodium album biocontrol. J. Nat. Prod. 2013, 76, 1291–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Aparicio, M.; Andolfi, A.; Cimmino, A.; Rubiales, D.; Evidente, A. Stimulation of seed germination of Orobanche species by ophobolin A and fusicoccin derivatives. J. Agric. Food Chem. 2008, 56, 8343–8347. [Google Scholar] [CrossRef]
- Ohkanda, J. Fusicoccin: A chemical modulator for 14-3-3 proteins. Chem. Lett. 2021, 50, 57–67. [Google Scholar] [CrossRef]
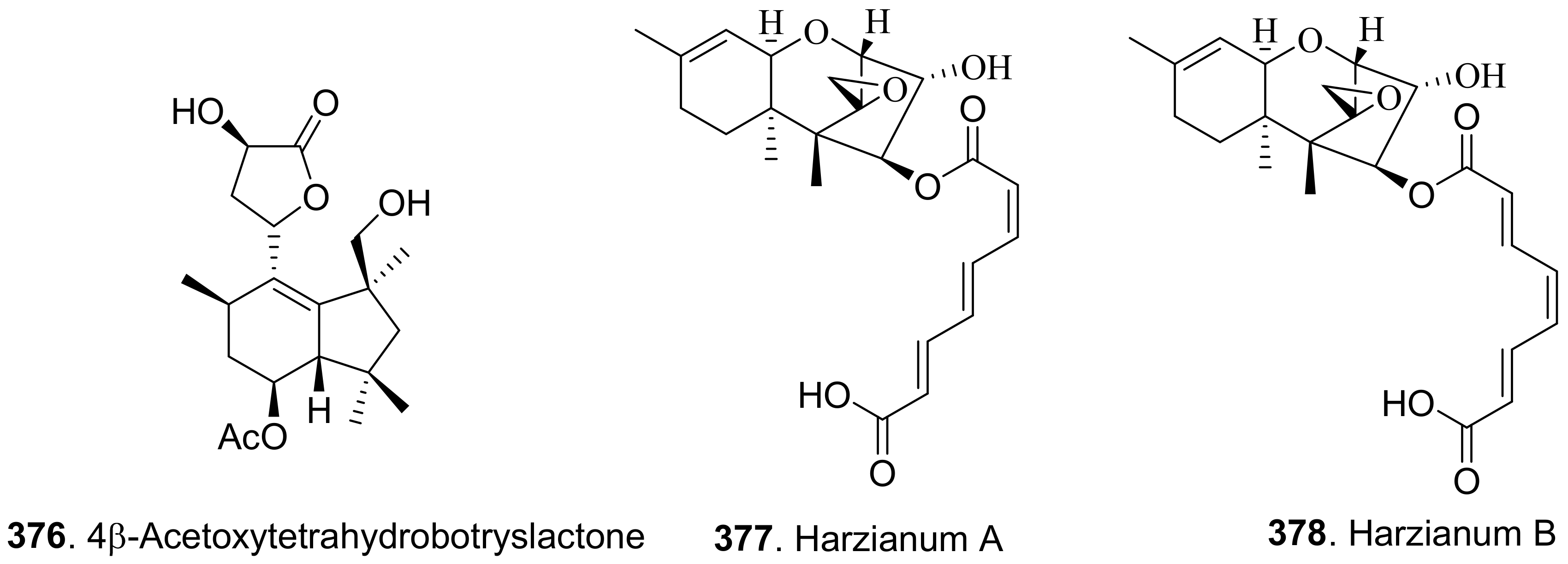
- Zhao, D.-L.; Yang, L.-J.; Shi, T.; Wang, C.-Y.; Shao, C.-L.; Wang, C.-Y. Potent phytotoxic harziane diterpenes from a soft coral-derived strain of the fungus Trichoderma harzianum XS-20090075. Sci. Rep. 2019, 9, 13345. [Google Scholar] [CrossRef] [Green Version]
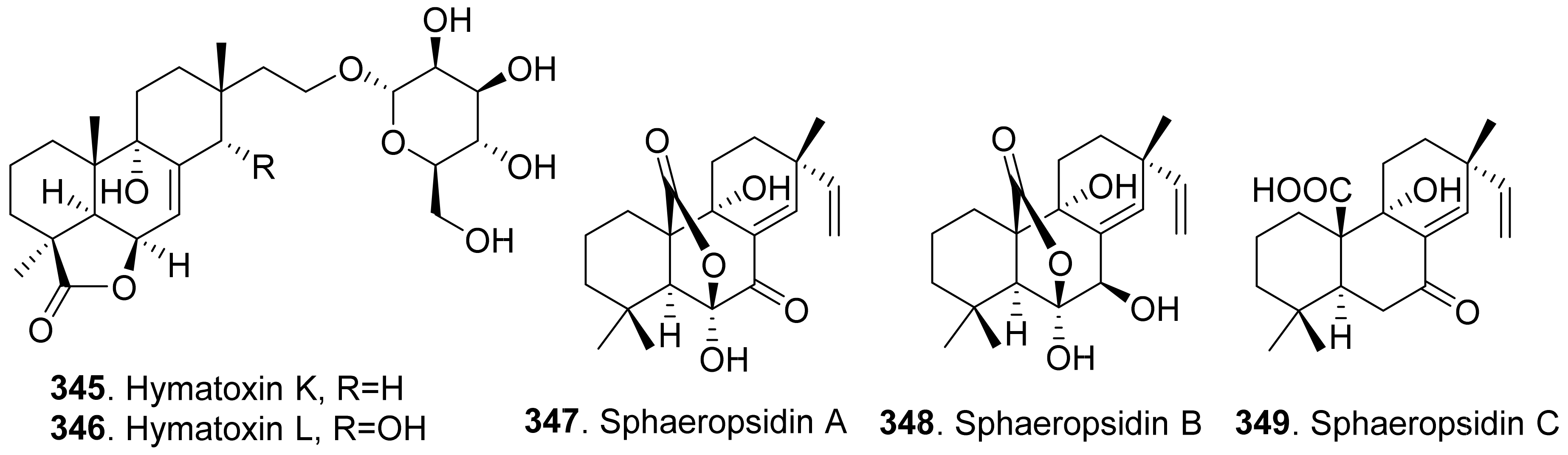
- Bodo, B.; Davoust, D.; Lecommandeur, D.; Rebuffat, S.; Genetet, I.; Pinon, J. Hymatoxin A, a diterpene sulfate phytotoxin of Hypoxylon mammatum, parasite of aspen. Tetrahedron Lett. 1987, 28, 2355–2358. [Google Scholar] [CrossRef]
- Jossang, A.; Mbeminack, B.; Pinon, J.; Bodo, B. Hyatoxins K and L, novel phytotoxins from Hypoxylon mammatum, fungal pathogen of aspens. Nat. Prod. Lett. 1995, 6, 37–42. [Google Scholar] [CrossRef]
- Evidente, A.; Sparapano, L.; Motta, A.; Giordano, F.; Fierro, O.; Frisullo, S. A phytotoxic pimarane diterpene of Sphaeropsis sapinea f. sp. cupressi, the pathogen of a canker disease of cypress. Phytochemistry 1996, 42, 1541–1546. [Google Scholar] [CrossRef]
- Evidente, A.; Sparapano, L.; Fierro, O.; Bruno, G.; Giordano, F.; Motta, A. Sphaeropsidins B and C, phytotoxic pimarane diterpenes from Sphaeropsis sapinea f. sp. cupressi and Diplodia mutila. Phytochemistry 1997, 45, 705–713. [Google Scholar] [CrossRef]
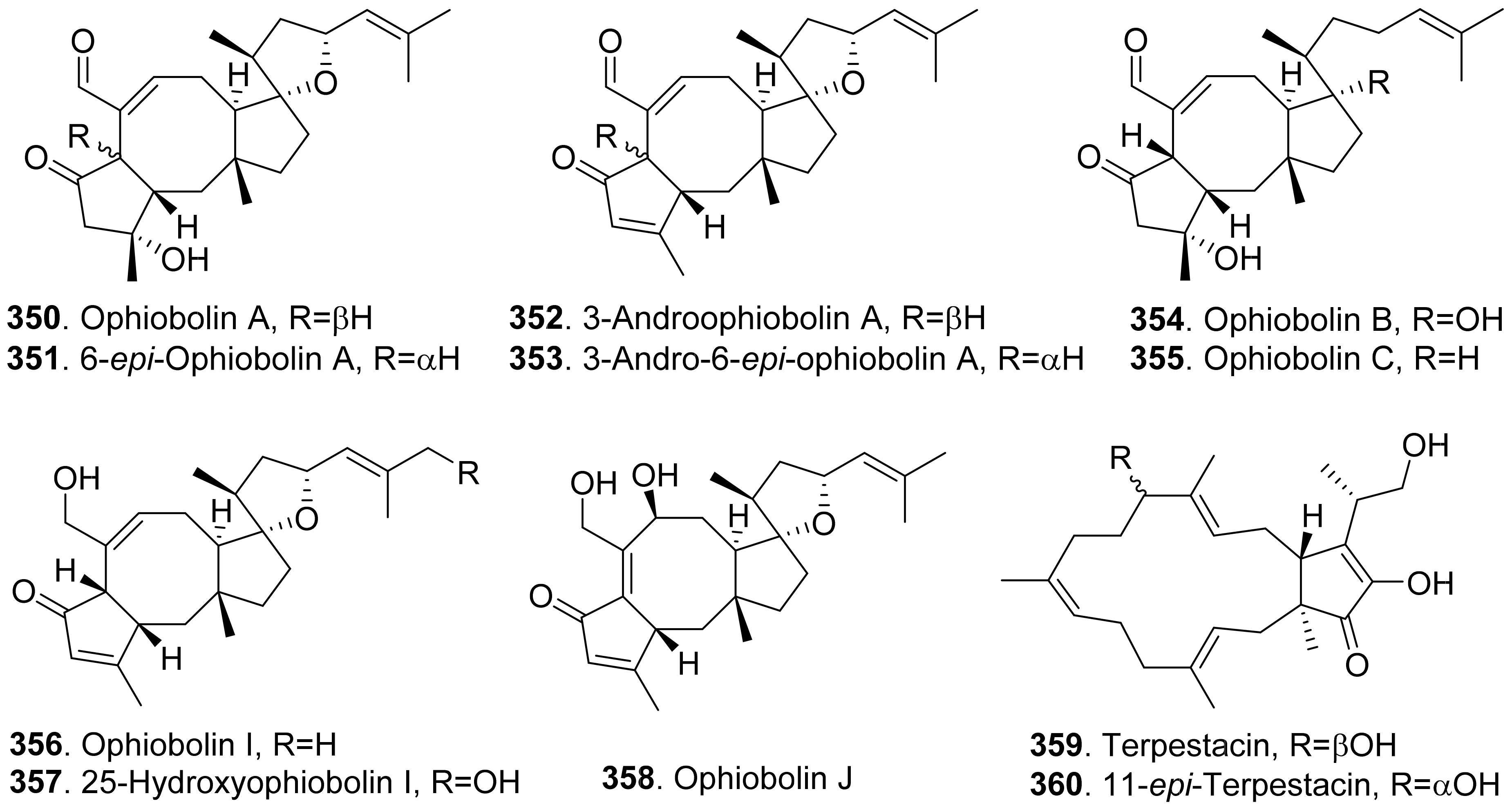
- Au, T.K.; Chick, W.S.H.; Leung, P.C. The biology of ophiobolins. Life Sci. 2000, 67, 733–742. [Google Scholar] [CrossRef]
- Tian, W.; Deng, Z.; Hong, K. The biological activities of sesterterpenoid-type ophiobolins. Mar. Drugs 2017, 15, 229. [Google Scholar] [CrossRef] [Green Version]
- Masi, M.; Dasari, R.; Evidente, A.; Mathieu, V.; Kornienko, A. Chemistry and biology of ophiobolin A and its congeners. Bioorg. Med. Chem. Lett. 2019, 29, 859–869. [Google Scholar] [CrossRef]
- Kim, J.-M.; Hyeon, S.-B.; Isogai, A.; Suzuki, A. Isotation of ophiobolin A and its analogs as inhibitors to photosynthesis. Agric. Biol. Chem. 1984, 48, 803–805. [Google Scholar]
- Evidente, A.; Andolfi, A.; Cimmino, A.; Vurro, M.; Fracchiolla, M.; Charudattan, R. Herbicidal potential of ophiobolins produced by Drechslera gigantean. J. Agric. Food Chem. 2006, 54, 1779–1783. [Google Scholar] [CrossRef]
- Pena-Rodriguez, L.M.; Chilton, W.S. 3-Anhydroophiobolin A and 3-anhydro-6-epi-ophiobolin A, phytotoxic metabolites of the Johnson grass pathogen Bipolaris sorghicola. J. Nat. Prod. 1989, 52, 1170–1172. [Google Scholar] [CrossRef]
- Sugawara, F.; Strobel, G.; Strange, R.N.; Siedow, J.N.; Van Duyne, G.D.; Clardy, J. Phytotoxins from the pathogenic fungi Drechslera maydis and Drechelera sorghicola. Proc. Nat. Acad. Sci. USA 1987, 84, 3081–3085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.J.; Kim, J.C.; Kim, B.S.; Kim, H.G.; Cho, K.Y. Antibiotic and phytotoxic activities of ophiobolins from Helminthosporium species. Plant Pathol. J. 1999, 15, 14–20. [Google Scholar]
- Sugawara, F.; Takahashi, N.; Strobel, G.; Yun, C.; Gray, G.; Fu, Y.; Clardy, J. Some new phytotoxic ophiobolins produced by Drechslera oryzae. J. Org. Chem. 1988, 53, 2170–2172. [Google Scholar] [CrossRef]
- Nihashi, Y.; Lim, C.-H.; Tanaka, C.; Miyagawa, H.; Ueno, T. Phytotoxic sesterterpene, 11-epiterpestacin, from Bipolaris sorokiniana NSDR-011. Biosci. Biotechnol. Biochem. 2002, 66, 685–688. [Google Scholar] [CrossRef] [Green Version]
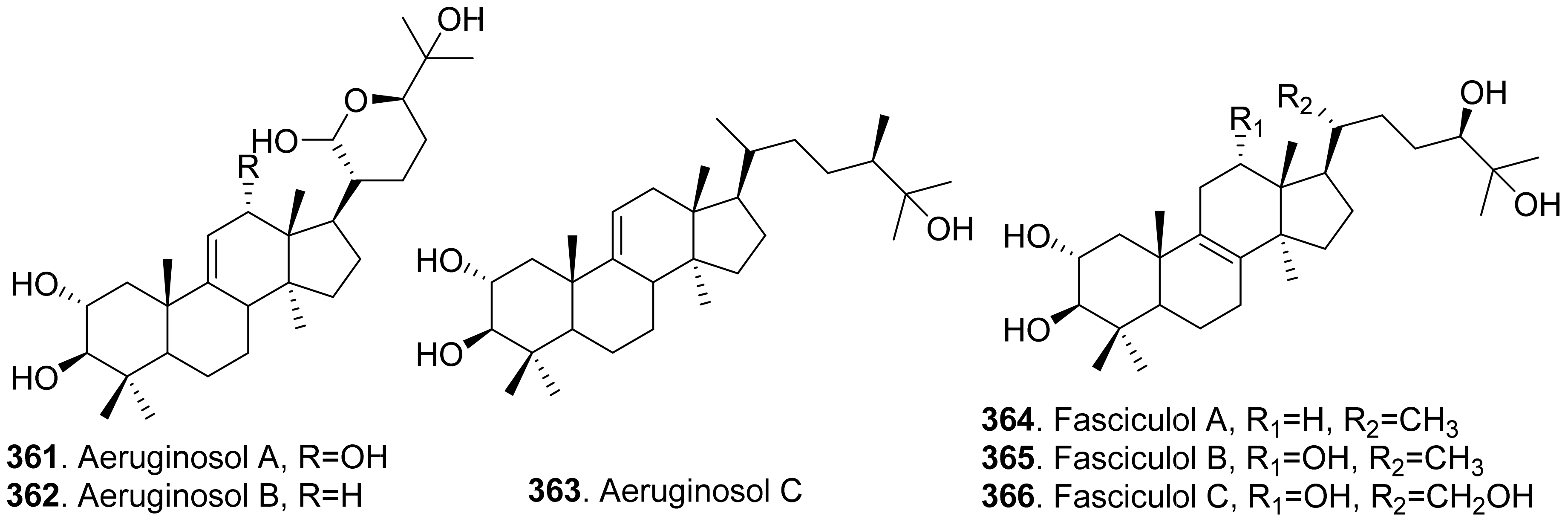
- Shiono, Y.; Sugawara, H.; Nazarova, M.; Mrayama, T.; Takahashi, K.; Ikeda, M. Three lanostane triterpenoids, aeruginosols A, B and C, from the fruiting bodies of Stropharia aeruginosa. J. Asian Nat. Prod. Res. 2007, 9, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, M.; Stato, Y.; Izawa, M.; Sassa, T.; Miura, Y. Isolation and structure of fasciculol A, a new plant growth inhibitor from Neamatoloma jasciculare. Agric. Biol. Chem. 1977, 41, 1539–1541. [Google Scholar]
- Ikeda, M.; Watanabe, H.; Hayakawa, A.; Sato, K.; Sassa, T.; Miura, Y. Structures of fasciculol B and its depsipepide, new biologically active substances from Neamatoloma jasciculare. Agric. Biol. Chem. 1977, 41, 1543–1545. [Google Scholar]
- Ikeda, M.; Niwa, G.; Tohyama, K.; Sassa, T.; Miura, Y. Structures of fasciculol C and its depsipeptides, new biologically active substances from Neamatoloma jasciculare. Agric. Biol. Chem. 1977, 41, 1803–1805. [Google Scholar]
- Evidente, A.; Lanzetta, R.; Abouzeid, M.A.; Corsaro, M.M.; Mugnai, L.; Surico, G. Foeniculoxin, a new phytotoxic geranylhydroquinone from Phomopsis foeniculi. Tetrahedron 1994, 50, 10371–10378. [Google Scholar] [CrossRef]
- Masi, M.; Sautua, F.; Zatout, R.; Castaldi, S.; Arrico, L.; Isticato, R.; Pescitelli, G.; Carmona, M.A.; Evidente, A. Phaseocyclopentenones A and B, phytotoxic penta- and tetrasubstituted cyclopentenones produced by Macrophomina phaseolina, the causal agent of charcoal rot of soybean in Argentina. J. Nat. Prod. 2021, 84, 459–465. [Google Scholar] [CrossRef]
- Cimmino, A.; Andolfi, A.; Zonno, M.C.; Troise, C.; Santini, A.; Tuzi, A.; Vurro, M.; Ash, G.; Evidente, A. Phomentrioloxin: A phytotoxic pentasubstituted geranylcyclohexentriol produced by Phomopsis sp., a potential mycoherbicide for Carthamus lanatus biocontrol. J. Nat. Prod. 2012, 75, 1130–1137. [Google Scholar] [CrossRef]
- Cimmino, A.; Andolfi, A.; Zonno, M.C.; Boari, A.; Troise, C.; Motta, A.; Vurro, M.; Ash, G.; Evidente, A. Phomentrioloxin, a fungal phytotoxin with potential herbicidal activity, and its derivatives: A structure−activity relationship study. J. Agric. Food Chem. 2013, 61, 9645–9649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colmenares, A.J.; Duran-Patron, R.M.; Hernandez-Galan, R.; Collando, I.G. Four new lactones from Botrytis cinera. J. Nat. Prod. 2002, 65, 1724–1726. [Google Scholar] [CrossRef] [PubMed]
- Miyagawa, H.; Nagai, S.; Tsurushima, T.; Sato, M.; Ueno, T.; Fukami, H. Phytotoxins produced by the plant pathogenic fungus Bipolaris bicolor El-1. Biosci. Biotechnol. Biochem. 1994, 58, 143–1145. [Google Scholar] [CrossRef] [Green Version]
- Long, Y.; Tang, T.; Wang, L.-Y.; He, B.; Gao, K. Absolute configuration and biological activities of meroterpenoids from an endophytic fungus of Lycium barbarum. J. Nat. Prod. 2019, 82, 2229–2237. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Zhang, J.; Song, Z.; Zhu, G.; Liu, M.; Dai, H.; Hsiang, T.; Liu, X.; Zhang, L.; Quinn, R.J.; et al. Genome-based mining of new antimicrobial meroterpenoids from the phytopathogenic fungus Bipolaris sorokiniana strain 11134. Appl. Microbiol. Biotechnol. 2020, 104, 3835–3846. [Google Scholar] [CrossRef]
- Yin, M.; Fasoyin, O.E.; Wang, C.; Yue, Q.; Zhang, Y.; Dun, B.; Xu, Y.; Zhang, L. Herbicidal efficacy of harzianums produced by the biofertilizer fungus, Trichoderma brevicompactum. AMB Expr. 2020, 10, 118. [Google Scholar] [CrossRef]
- Gohbara, M.; Kosuge, Y.; Yamasaki, S.; Kimura, Y.; Suzuki, A.; Tamura, S. Isolation, structures and biological activities of colletotrichins, phytotoxic substances from Colletotrichum nicotianae. Agric. Biol. Chem. 1978, 42, 1037–1043. [Google Scholar] [CrossRef]
- Wang, X.; Gong, X.; Li, P.; Lai, D.; Zhou, L. Structural diversity and biological activities of cyclic depsipeptides from fungi. Molecules 2018, 23, 169. [Google Scholar] [CrossRef] [Green Version]
- Ueno, T.; Nakashima, T.; Hayashi, Y.; Fukami, H. Structures of AM-toxin I and II, host specific phytotoxic metabolites produced by Alternaria mali. Agric. Biol. Chem. 1975, 39, 1115–1122. [Google Scholar] [CrossRef]
- Ueno, T.; Nakashima, T.; Hayashi, Y.; Fukami, H. Isolation and structure of AM-toxin III, a host specific phytotoxic metabolite produced by Alternaria mali. Agric. Biol. Chem. 1975, 39, 2081–2082. [Google Scholar] [CrossRef] [Green Version]
- Miyashita, M.; Nakamori, T.; Miyagawa, H.; Akamatsu, M.; Ueno, T. Inhibitor activity of analogs of AM-toxin, a host-specific phytotoxin from the Alternaria alternata apple pathotype, on photosynthetic O2 evolution in apple leaves. Biosci. Biotechnol. Biochem. 2003, 67, 635–638. [Google Scholar] [CrossRef] [PubMed]
- Evans, N.; Mcroberts, N.; Hill, R.A.; Marshall, G. Phytotoxin production by Alternaria linicola and phytoalexin production by the linseed host. Ann. Appl. Biol. 1996, 129, 415–431. [Google Scholar] [CrossRef]
- Bains, P.S.; Tewari, J.P. Purificaiton, chemical characterization and host-specificity of the toxin produced by Alternaria brassicae. Physiol. Mol. Plant Pathol. 1987, 30, 259–271. [Google Scholar] [CrossRef]
- Pedras, M.S.C.; Zaharia, I.L.; Gai, Y.; Smith, K.C.; Ward, D.E. Metabolism of the hsot-selective toxins destruxin B and homodestruxin B: Probing a plant disease resistance trait. Org. Lett. 1999, 1, 1655–1658. [Google Scholar] [CrossRef]
- Hornbogen, T.; Glinski, M.; Zocher, R. Biosynthesis of depsipeptide mycotoxins in Fusarium. Eur. J. Plant Pathol. 2002, 108, 713–718. [Google Scholar] [CrossRef]
- Burmeister, H.R.; Plattner, R.D. Enniatin production by Fusarium tricinctum and its effect on germinating wheat seeds. Phytopathology 1987, 77, 1483–1487. [Google Scholar] [CrossRef]
- Herrmann, M.; Zocher, R.; Haese, A. Enniatin production by Fusarium strains and its effect on potato tuber tissue. Appl. Environ. Microbiol. 1996, 62, 393–398. [Google Scholar] [CrossRef] [Green Version]
- Hershenhorn, J.; Park, S.H.; Stierle, A.; Strobel, G.A. Fusarium avenaceum as a novel pathogen of spotted knapweed and its phytotoxins, acetamido-butenolide and enniatin B. Plant Sci. 1992, 86, 155–160. [Google Scholar] [CrossRef]
- Bahadoor, A.; Brauer, E.K.; Bosnich, W.; Schneiderman, D.; Johnston, A.; Aubin, Y.; Blackwell, B.; Melanson, J.E.; Harris, L.J. Gramillin A and B: Cyclic lipopeptides identified as the nonribosomal biosynthetic products of Fusarium graminearum. J. Am. Chem. Soc. 2018, 140, 16783–16791. [Google Scholar] [CrossRef]
- Pedras, M.S.C.; Biesenthal, C.J. Production of the host-selective phytotoxin phomalideby isolates of Leptosphaeria maculans and its correlation with sirodesmin PL production. Can. J. Microbiol. 1998, 44, 547–553. [Google Scholar] [CrossRef]
- Zabka, M.; Drastichova, K.; Jegorov, A.; Soukupova, J.; Nedbal, L. Direct evidence of plant-pathogenic activity of fungal metabolites of Trichothecium roseum on apple. Mycopathologia 2006, 162, 65–68. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, Y.; Zhang, X.; Lai, D.; Zhou, L. Structrual diversity and biological activities of the cyclodipeptides from fungi. Molecules 2017, 22, 2026. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Lin, M.; Xu, D.; Lai, D.; Zhou, L. Structural diversity and biological activities of fungal cyclic peptides, excluding cyclodipeptides. Molecules 2017, 22, 2069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Wang, S.-Q.; Tang, H.-Y.; Li, X.-J.; Zhang, L.; Xiao, J.; Gao, Y.-Q.; Zhang, A.-L.; Gao, J.-M. Potential allelopathic indole diketopiperazines produced by the plant endophytic Aspergillus fumigatus using the one strain-many compounds method. J. Agric. Food Chem. 2013, 61, 11447–11452. [Google Scholar] [CrossRef] [PubMed]
- Stierle, A.C.; Cardellina II, J.H.; Strobel, G.A. Maculosin, a host-specific phytotoxin for spotted knapweed from Alternaria alternata. Proc. Natl. Acad. Sci. USA 1988, 85, 8008–8011. [Google Scholar] [CrossRef] [Green Version]
- Pedras, M.S.C.; Abrams, S.R.; Séguin-Swartz, G.; Quail, J.W.; Jia, Z.C. Phomalirazine, a novel toxin from the phytopathogenic fungus Phoma lingam. J. Am. Chem. Soc. 1989, 111, 1904–1905. [Google Scholar] [CrossRef]
- Pedras, M.S.C.; Biesenthal, C.J. Isolation, structure determination, and phytotoxicity of unusual dioxopiperazines from the phytopathogenic fungus Phoma lingam. Phytochemistry 2001, 58, 905–909. [Google Scholar] [PubMed]
- Pedras, M.S.C.; Séguin-Swartz, G.; Abrams, S.R. Minor phytotoxins from the blackleg fungus Phoma lingam. Phytochemistry 1990, 29, 777–782. [Google Scholar] [CrossRef]
- Mitrovic, P.M.; Orcic, D.Z.; Sakac, V.O.; Marjanovic-Jeromela, A.M.; Grahovac, N.L.; Milosevic, D.M.; Marisavljevic, D.P. Characerization of sirodesmins isolated from the phytopathogenic fungus Leptosphaeria macuans. J. Serb. Chem. Soc. 2012, 77, 1363–1379. [Google Scholar] [CrossRef]
- Jin, J.; Baek, S.-R.; Lee, K.-R.; Lee, J.; Yun, S.-H.; Kang, S.; Lee, Y.-W. Purification and phytotoxicity of apicidins produced by the Fusarium semitectum KCTC16676. Plant Pathol. J. 2008, 24, 417–422. [Google Scholar] [CrossRef]
- Gupta, S.; Peiser, G.; Nkajima, T.; Hwang, Y.-S. Characterization of a phytotoxic cyclotetrapeptide, a novel chlamydocin analogue, from Verticillium coccosporum. Tetrahedron Lett. 1994, 35, 6009–6012. [Google Scholar] [CrossRef]
- Isogai, A.; Takayama, S.; Hirota, A.; Suzuki, A. Determination of the absolute stereochemistry of the epoxide of aoe (2-amino-8-oxa-9,10-epoxy-decanoic acid) in Cyl-1 and Cyl-2 by CD spectra. Agric. Biol. Chem. 1986, 50, 517–518. [Google Scholar]
- Takayama, S.; Lsogai, A.; Nakata, M.; Suzuki, H.; Susuki, A. Structure of Cyl-1, a novel cyclic tetrapetide from Cylindrocladium scoparium. Agric. Biol. Chem. 1984, 48, 839–842. [Google Scholar]
- Hirota, A.; Suzuki, A.; Aizawa, K.; Tamura, S. Structure of Cyl-2, a novel cyclopeptide from Cylindrocladium scoparium. Agric. Biol. Chem. 1973, 37, 955–956. [Google Scholar] [CrossRef]
- Liesch, J.M.; Sweeley, C.C.; Staffeld, G.D.; Anderson, M.S.; Weber, D.J.; Scheffer, R.P. Structure of HC-toxin, a cyclic tetrapeptide from Helminthosporium carbonum. Tetrahedron 1982, 38, 45–48. [Google Scholar] [CrossRef]
- Rasmussen, J.B.; Scheffer, R.P. Isolation and biological activities of four selective toxins from Helminthosporium carbonum. Plant Physiol. 1988, 86, 187–191. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, T.; Kono, Y.; Takeuchi, S.; Daly, J.M. Structure of HV-toxin M, a host-specific toxin-related compound produced by Helminthosporium victoriae. Agric. Biol. Chem. 1989, 53, 1283–1290. [Google Scholar] [CrossRef]
- Allen, J.G.; Hancock, G.R. Evidence that phomopsins A and B are not the only toxic metabolites produced by Phomopsis leptostromiformis. J. Appl. Toxicol. 1989, 9, 83–89. [Google Scholar] [CrossRef]
- Culvenor, C.C.J.; Edgar, J.A.; Mackay, M.F.; Gorst-Allman, C.P.; Marasas, W.F.O.; Steyn, P.S.; Vleggaar, R.; Wessels, P.L. Structure elucidation and absolute configuration of phomopsin A, a hexapeptide mycotoxin produced by Phomopsis leptostromiformis. Tetrahedron 1989, 45, 2351–2372. [Google Scholar] [CrossRef]
- Liebermann, B.; Koelblin, R. A new phytotoxic activity of the cyclic peptides tentoxin and dihydrotentoxin. J. Phytopathol. 1992, 135, 245–250. [Google Scholar] [CrossRef]
- Liebermann, B.; Ellinger, R.; Pinet, E. Isotentoxin, a conversion product of the phytotoxin tentoxin. Phytochemistry 1996, 42, 1537–1540. [Google Scholar] [CrossRef]
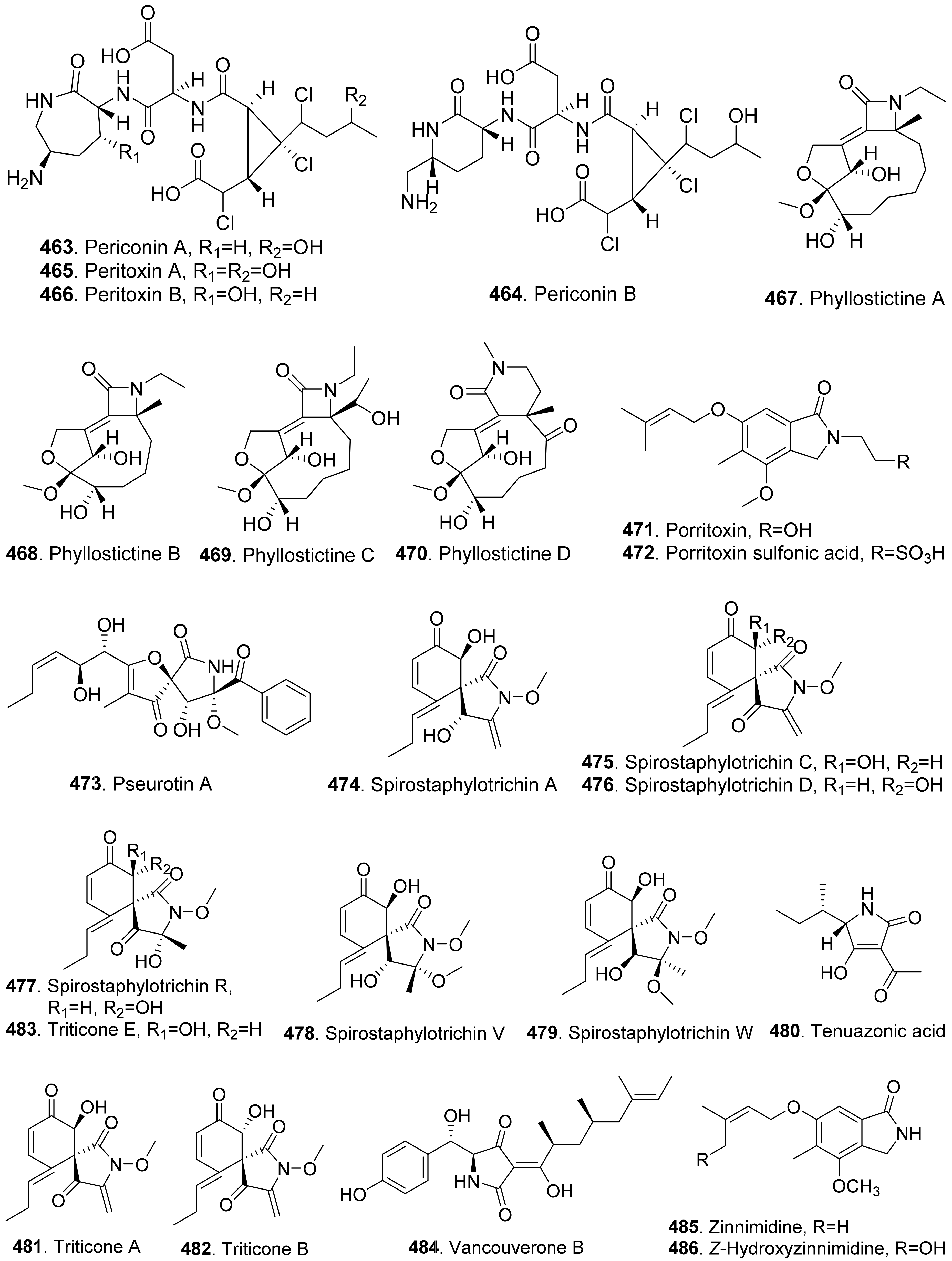
- Horiuchi, M.; Ohnishi, K.; Iwase, N.; Nakajima, Y.; Tounai, K.; Yamashita, M.; Yamada, Y. A novel isoindoline, porritoxin sulfonic acid, from Alternaria porri and the structure-phytotoxicity correlation of its related compounds. Biosci. Biotechnol. Biochem. 2003, 67, 1580–1583. [Google Scholar] [CrossRef] [Green Version]
- Wolpert, T.J.; Macko, V.; Acklin, W.; Jaun, B.; Seibl, J.; Meili, J.; Arigoni, D. Structure of victorin C, the major host-selective toxin from Cochliobolus victoriae. Experientia 1985, 41, 1524–1529. [Google Scholar] [CrossRef]
- Liakopoulou-Kyriakides, M.; Lagopodi, A.L.; Thanassoulopoulos, C.C.; Stavropoulos, G.S.; Magafa, V. Isolation and synthesis of a host-selective toxin produced by Alternaria alternata. Phytochemistry 1997, 45, 37–40. [Google Scholar] [CrossRef]
- Pedras, M.S.C.; Chumala, P.B.; Quail, J.W. Chemical mediators: The remarkable structure and host-selectivity of depsilairdin, a sesquiterpenic depsipeptide containing a new amino acid. Org. Lett. 2004, 6, 4615–4617. [Google Scholar] [CrossRef]
- Capio, E.R.; Tate, M.E.; Wallwork, H. Phytotoxic metabolites from Drechslera wirrenganensis and D. campanulata. Australas. Plant Pathol. 2004, 33, 23–28. [Google Scholar] [CrossRef]
- Biasetto, C.R.; Somensi, A.; Abdalla, V.C.P.; De Abreu, L.M.; Gualtieri, S.C.J.; Pfenning, L.H.; Bolzani, V.S.; Araujo, A.R. Phytotoxic constituents from endophytic fungus Xylaria cubensis associated with Eugenia brasiliensis. Quim. Nova 2019, 42, 485–488. [Google Scholar] [CrossRef]
- Latif, Z.; Strange, R.N.; Bilton, J.; Riazuddin, S. Production of the phytotoxins, solanapyrones A and C and cytochalasin D among nine isolates of Ascochyta rabiei. Plant Pathol. 1993, 42, 172–180. [Google Scholar] [CrossRef]
- Cimmino, A.; Andolfi, A.; Berestetskiy, A.; Evidente, A. Production of phytotoxins by Phoma exigua var. exigua, a potential mycoherbicide against perennial thistles. J. Agric. Food Chem. 2008, 56, 6304–6309. [Google Scholar] [CrossRef]
- Li, H.; Wei, H.; Hu, J.; Lacey, E.; Sobolev, A.N.; Stubbs, K.A.; Solomon, P.S.; Chooi, Y.-H. Genomics-driven discovery of phytotoxic cytochalasans involved in the virulence of the wheat pathogen Parastagonospora nodorum. ACS Chem. Biol. 2020, 15, 226–233. [Google Scholar] [CrossRef]
- Nukina, M. Pyrichalasin H, a new phytotoxic metabolite belonging to the cytochalasans from Pyricularia grisea (Cooke) Saccardo. Agric. Biol. Chem. 1987, 51, 2625–2628. [Google Scholar] [CrossRef]
- Sassa, T.; Onuma, Y. Isolation and identification of fruit rot toxins from the fungus-caused Macrophoma fruit rot of apple. Agric. Biol Chem. 1983, 47, 1155–1157. [Google Scholar] [CrossRef]
- Evidente, A.; Fiore, M.; Bruno, G.; Sparapano, L.; Motta, A. Chemical and biological characterisation of sapinopyridione, a phytotoxic 3,3,6-trisubstituted-2,4-pyridione produced by Sphaeropsis sapinea, a toxigenic pathogen of native and exotic conifers, and its derivatives. Phytochemistry 2006, 67, 1019–1028. [Google Scholar] [CrossRef]
- Kumarihamy, M.; Rosa, L.H.; Techen, N.; Ferreira, D.; Croom, E.M.; Duke, S.O.; Tekwani, B.L.; Khan, S.; Nanayakkara, N.P.D. Antimalarials and phytotoxins from Botryosphaeria dothidea identified from a seed of diseased Torreya taxifolia. Molecules 2021, 26, 59. [Google Scholar] [CrossRef] [PubMed]
- Macko, V.; Stimmel, M.B.; Wopert, T.J.; Dunkle, L.D.; Acklin, W.; Banteli, R.; Jaun, B.; Arigoni, D. Structure of the host-specific toxins produced by the fungal pathogen Periconia circinata. Proc. Natl. Acad. Sci. USA 1992, 89, 9574–9578. [Google Scholar] [CrossRef] [Green Version]
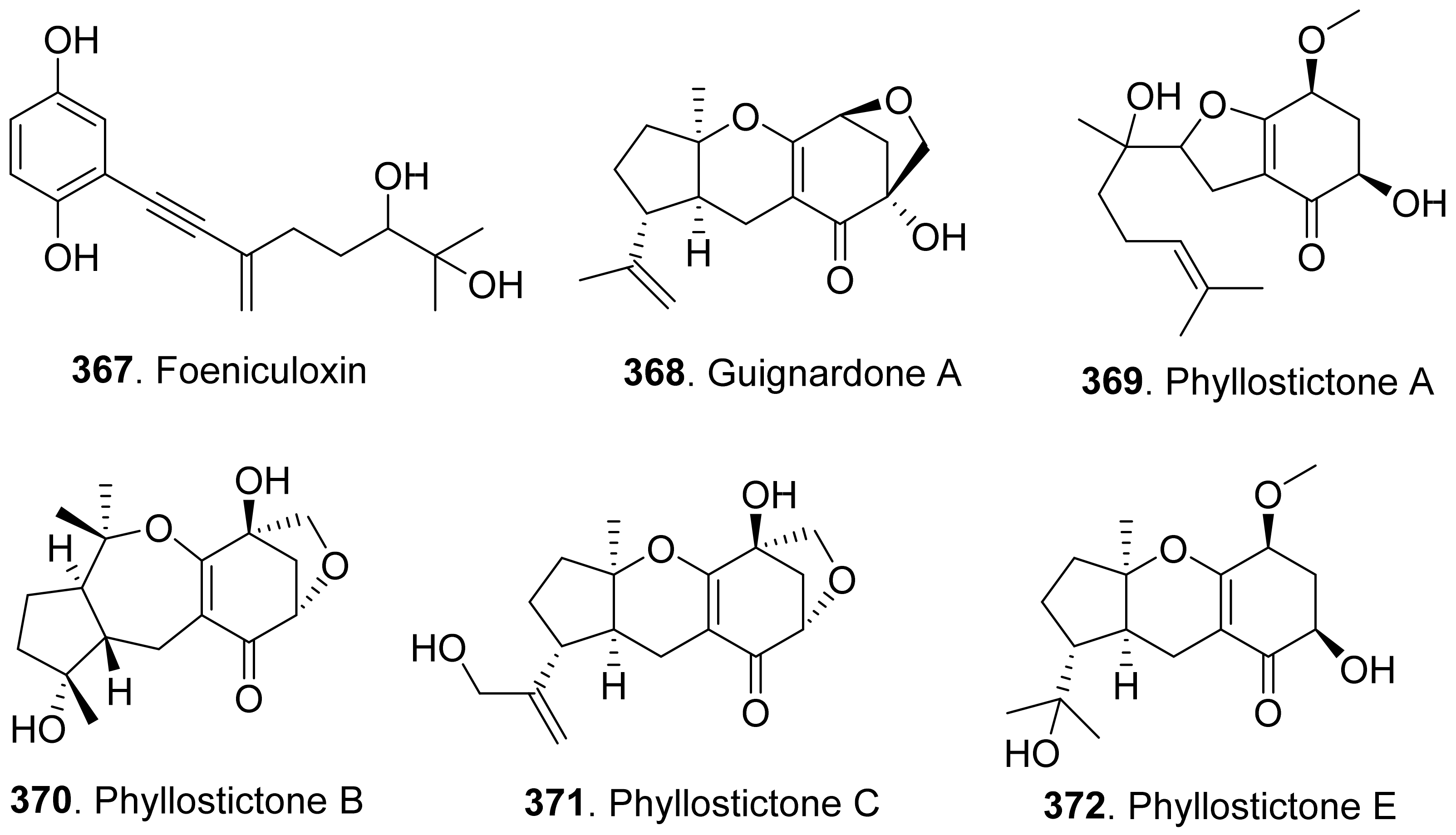
- Evidente, A.; Cimmino, A.; Andolfi, A.; Vurro, M.; Zonno, M.C.; Cantrell, C.L.; Motta, A. Phyllostictines A-D, oxazatricycloalkenones produced by Phyllosticta cirsii, a potential mycoherbicide for Cirsium arvense biocontrol. Tetrahedron 2008, 64, 1612–1619. [Google Scholar] [CrossRef]
- Suemitsu, R.; Ohnishi, K.; Horiuchi, M.; Kitaguchi, A.; Odamura, K. Porritoxin, a phytotoxin of Alternaria porri. Phytochemistry 1992, 31, 2325–2326. [Google Scholar] [CrossRef]
- Horiuchi, M.; Maoka, T.; Iwase, N.; Ohnishi, K. Reinvestigation of structure of porritoxin, a phytotoxin of Alternaria porri. J. Nat. Prod. 2002, 65, 1204–1205. [Google Scholar] [CrossRef] [PubMed]
- Masi, M.; Meyer, S.; Clement, S.; Andolfi, A.; Cimmino, A.; Evidente, A. Spirostaphylotrichin W, a spirocyclic γ-lactam isolated from liquid culture of Pyrenophora semeniperda, a potential mycoherbicide for cheatgrass (Bromus tectorum) biocontrol. Tetrahedron 2014, 70, 1497–1501. [Google Scholar] [CrossRef]
- Liu, Y.-X.; Xu, X.M.; Dai, X.-B.; Qiang, S. Alternaria alternata crofton-weed toxin: A natural inhibitor of photosystem II in Chlamydomonas reinhardtii thylakoids. J. Agric. Food Chem. 2007, 55, 5180–5185. [Google Scholar] [CrossRef]
- Kono, Y.; Gardner, J.M.; Takeuchi, S. Nonselective phytotoxins simultaneously produced with host-selective ACTG-toxins by a pathotype of Alternaria citri causing brown spot disease of mandarins. Agric. Biol. Chem. 1986, 50, 2401–2403. [Google Scholar] [CrossRef]
- Sattar, A.; Alam, M.; Janardhanan, K.K.; Husain, A. Isolation of tenuazonic acid, a phytotoxin from Alternaria crassa (Sacc.) rands causing leaf blight and fruit rot of Datura stramonium Mill. Curr. Sci. 1986, 55, 195–196. [Google Scholar]
- Davis, N.D.; Diener, U.L.; Morgan-Jones, G. Tenuazonic acid production by Alternaria alternata and Alternaria tenuissima isolated from cotton. Appl. Environ. Microbiol. 1977, 34, 155–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umetsu, N.; Kaji, J.; Tamari, K. Investigation on the toxin production by several blast fungus strains and isolation of tenuazonic acid as a novel toxin. Agric. Biol. Chem. 1972, 36, 859–866. [Google Scholar] [CrossRef]
- Steyn, P.S.; Rabie, C.J. Characterization of magnesium and calcium tenuazonate from Phoma sorghina. Phytochemistry 1976, 15, 1977–1979. [Google Scholar] [CrossRef]
- Chen, S.; Xu, X.; Dai, X.; Yang, C.; Qiang, S. Identification of tenuazonic acid as a novel type of natural photosystem II inhibitor binding in Q(B)-site of Chlamydomonas reinhardtii. Biochim. Biophys. Acta 2007, 1767, 306–318. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Yin, C.; Qiang, S.; Zhou, F.; Dai, X. Chloroplastic oxidative burst induced by tenuazonic acid, a natural photosynthesis inhibitor, triggers cell necrosis in Eupatorium adenophorum Spreng. Biochim. Biophys. Acta 2010, 1797, 391–405. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Kim, C.; Lee, J.M.; Lee, H.A.; Fei, Z.; Wang, L.; Apel, K. Blocking the QB-binding site of photosystem II by tenuazonic acid, a non-host-specific toxin of Alternaria alternata, activates singlet oxygen-mediated and EXECUTER-dependent signalling in Arabidopsis. Plant Cell Environ. 2015, 38, 1069–1080. [Google Scholar] [CrossRef]
- Chen, S.; Qiang, S. Recent advances in tenuazonic acid as a potential herbicide. Pestic. Biochem. Physiol. 2017, 143, 252–257. [Google Scholar] [CrossRef]
- Sugawara, F.; Takahashi, N.; Strobel, G.A.; Strobel, S.A.; Lu, H.S.M.; Clardy, J. Triticones A and B, novel phytotoxins from the plant pathogenic fungus Drechelera tritici-repentis. J. Am. Chem. Soc. 1988, 110, 4086–4087. [Google Scholar] [CrossRef]
- Yokoyama, Y.; Koseki, T.; Harneti, D.; Maharani, R.; Supratman, U.; Shiono, Y. Phytotoxic compounds isolated from a sea snail derived fungus, Penicillium vancouverense YY-1. Phytochem. Lett. 2020, 39, 57–63. [Google Scholar] [CrossRef]
- Wang, Z.-F.; Zhang, W.; Xiao, L.; Zhou, Y.-M.; Du, F.-Y. Characterization and bioactive potentials of secondary metabolites from Fusarium chlamydosporum. Nat. Prod. Res. 2020, 34, 889–892. [Google Scholar] [CrossRef] [PubMed]
- Masi, M.; Cimmino, A.; Boari, A.; Zonno, M.C.; Gorecki, M.; Pescitelli, G.; Tuzi, A.; Vurro, M.; Evidente, A. Colletopyrandione, a new phytotoxic tetrasubstituted indolylidenepyran-2,4-dione, and colletochlorins G and H, new tetrasubstituted chroman- and isochroman-3,5-diols isolated from Colletothichum higginsianum. Tetrahedron 2017, 73, 6644–6650. [Google Scholar] [CrossRef]
- Sugawara, F.; Strobel, G.A. Tryptophol, a phytotoxin produced by Drechslera nodulosum. Phytochemistry 1987, 26, 1349–1351. [Google Scholar] [CrossRef]
- Evidente, A.; Andolfi, A.; Abouzeid, M.A.; Vurro, M.; Zonno, M.C.; Motta, A. Ascosonchine, the enol tautomer of 4-pyridylpyruvic acid with herbicidal activity produced by Ascochyta sonchi. Phytochemistry 2004, 65, 475–480. [Google Scholar] [CrossRef]
- Reveglia, P.; Cinelli, T.; Cimmino, A.; Masi, M.; Evidente, A. The main phytotoxic metabolite produced by a strain of Fusarium oxysporum inducing grapevine plant declining in Italy. Nat. Prod. Res. 2018, 32, 2398–2407. [Google Scholar] [CrossRef]
- Wang, X.; Ng, T.B. Phamacological activities of fusaric acid (5-butylpicolinic acid). Life Sci. 1999, 65, 849–856. [Google Scholar] [CrossRef]
- Capasso, R.; Evidente, A.; Cutignano, A.; Vurro, M.; Zonno, M.C.; Bottalico, A. Fusaric and 9,10-dehydrofusaric acids and their methylesters from Fusarium nygamai. Phytochemistry 1996, 41, 1035–1039. [Google Scholar] [CrossRef]
- Liu, S.; Dai, H.; Orfali, R.S.; Lin, W.; Liu, Z.; Proksch, P. New fusaric acid derivatives from the endophytic fungus Fusarium oxysporum and their phytotoxicity to barley leaves. J. Agric. Food Chem. 2016, 64, 3127–3132. [Google Scholar] [CrossRef]
- Bottini, A.T.; Gilchrist, D.G. Phytotoxins. I. A 1-aminodimethylheptadecapentol from Alternaria alternata f.sp. lycopersici. Tetrahedron Lett. 1981, 22, 2719–2722. [Google Scholar] [CrossRef]
- Bottini, A.T.; Bowen, J.R.; Gilchrist, D.G. Phytotoxins. II. Characterization of a phytotoxic fraction from Alternaria alternata f.sp. lycopersici. Tetrahedron Lett. 1981, 22, 2723–2726. [Google Scholar] [CrossRef]
- Caldas, E.D.; Jones, A.D.; Ward, B.; Winter, C.K.; Gilchrist, D.G. Structural characterization of three new AAL toxins produced by Alternaria alternata f.sp. lycopersici. J. Agric. Food Chem. 1994, 42, 327–333. [Google Scholar] [CrossRef]
- Winter, C.K.; Gilchrist, D.G.; Dickman, M.B.; Jones, C. Chemistry and biological activity of AAL toxins. Adv. Exp. Med. Biol. 1996, 392, 307–316. [Google Scholar] [PubMed]
- Brandwagt, B.F.; Mesbah, L.A.; Takken, F.L.W.; Laurent, P.L.; Kneppers, T.J.A.; Hille, J.; Nijkamp, H.J. A longevity assurance gene homolog of tomato mediates resistance to Alternaria alternata f.sp. lycopersici toxins and fumonisin B1. Proc. Natl. Acad. Sci. USA 2000, 97, 4961–4966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohmoto, K.; Itoh, Y.; Shimomura, N.; Kondoh, Y.; Otani, H.; Kodama, M.; Nishimura, S.; Nakatsuka, S. Isolation and biological activities of two host-specific toxins from the tangerine pathotype of Alternaria alternata. Phytopathology 1993, 83, 495–502. [Google Scholar] [CrossRef]
- Ueno, T. Secondary metabolites related to host selection by plant pathogenic fungi. Pure Appl. Chem. 1990, 62, 1347–1352. [Google Scholar] [CrossRef]
- Weiergang, I.; Lyngs Jorgensen, H.J.; Moller, I.M.; Friis, P.; Smedegaard-Petersen, V. Optimization of in vitro growth conditions of Pyrenophora teres for production of the phytotoxin aspergillomarasmine A. Physiol. Mol. Plant Pathol. 2002, 60, 131–140. [Google Scholar] [CrossRef]
- Irvine, N.M.; Yerkes, C.N.; Graupner, P.R.; Roberts, R.E.; Hahn, D.R.; Pearce, C.; Gerwick, B.C. Synthesis and characterization of synthetic analogs of cinnacidin, a novel phytotoxin from Nectria sp. Pest. Manag. Sci. 2008, 64, 891–899. [Google Scholar] [CrossRef]
- Lamprecht, S.C.; Marasas, W.F.O.; Alberts, J.F.; Cawood, M.E.; Gelderblom, W.C.A.; Shephard, G.S.; Thiel, P.G.; Calitz, F.J. Phytotoxicity of fumonisins and TA-toxin to corn and tomato. Phytopathology 1994, 84, 383–391. [Google Scholar] [CrossRef]
- Wang, A.; Li, P.; Han, P.; Gu, G.; Shan, T.; Lai, D.; Zhou, L. New nitrogen-containing metabolites from cultures of rice flase smut pathogen Villosiclava virens. Nat. Prod. Res. 2021, 35, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Pedras, M.S.C.; Chumala, P.B.; Jin, W.; Islam, M.S.; Hauck, D.W. The phytopathogenic fungus Alternaria brassicicola: Phytotoxin production and phytoalexin elicitation. Phytochemistry 2009, 70, 394–402. [Google Scholar] [CrossRef]
- Hershenhorn, J.; Vurro, M.; Zonno, M.C.; Stierle, A.; Strobel, G. Septoria cirsii, a potential biocontrol agent of Canada thistle and its phytotoxin-β-nitropropionic acid. Plant Sci. 1993, 94, 227–234. [Google Scholar] [CrossRef]
- Coval, S.J.; Hradil, C.M.; Lu, H.S.M.; Clardy, J.; Satouri, S.; Strobel, G.A. Pyrenoline-A and –B, two new phytotoxins from Pyrenophora teres. Tetraheron Lett. 1990, 31, 2117–2120. [Google Scholar] [CrossRef]
- Nagata, T.; Ando, Y.; Hirota, A. Phytotoxins from tea gray blight fungi, Pestalotiopsis longiesta and Pestalotiopsis theae. Biosci. Biotechnol. Biochem. 1992, 56, 810–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Demerdash, A.; Genta-Jouve, G.; Barenstrauch, M.; Kunz, C.; Baudouin, E.; Prado, S. High oxygenated isoprenylated cyclohexanoids from the fungus Parastagonospora nodorum SN15. Phytochemistry 2019, 166, 112056. [Google Scholar] [CrossRef] [PubMed]
- Kenfield, D.; Bunkers, G.; Strobel, G.A.; Sugawara, F. Potential new herbicides—phytotoxins from plant pathogens. Weed Technol. 1988, 2, 519–524. [Google Scholar] [CrossRef]
- Abbas, H.K.; Duke, S.O. Phytotoxins from plant pathogens as potential herbicides. Toxin Rev. 1995, 14, 523–543. [Google Scholar] [CrossRef]
- Rossi, M.; Wen, K.; Caruso, F.; Belli, S. Emodin scavenging of superoxide radical includes π–π interaction. X-ray crystal structure, hydrodynamic voltammetry and theoretical studies. Antioxidants 2020, 9, 194. [Google Scholar] [CrossRef] [Green Version]
- Wei, W.-T.; Lin, S.-Z.; Liu, D.-L.; Wang, Z.-H. The distinct mechanisms of the antitumor activity of emodin in different types of cancer (review). Oncol. Rep. 2013, 30, 2555–2562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, X.-F.; Zhao, Z.-M.; Li, J.-C.; Yang, G.-Z.; Liu, Y.-Q.; Dai, L.-X.; Zhang, Z.-J.; Yang, Z.-G.; Miao, X.-L.; Yang, C.-J.; et al. Insecticidal and antifungal activities of Rheum palmatum L. anthraquinones and structurally related compounds. Ind. Crop Prod. 2019, 137, 508–520. [Google Scholar] [CrossRef]
- Xu, Y.; Mafezoli, J.; Oliveira, M.C.F.; U’Ren, J.M.; Arnold, A.E.; Gunatilaka, A.A.L. Anteaglonialides A-F and palmarumycins CE1-CE3 from Anteaglonium sp. FL0768, a fungal endophyte of the spikemoss Selaginella arenicola. J. Nat. Prod. 2015, 78, 2738–2747. [Google Scholar] [CrossRef] [PubMed]
- Paranagama, P.A.; Wijeratne, E.M.K.; Burns, A.M.; Marron, M.T.; Gunatilaka, M.K.; Arnold, A.E.; Gunatilaka, A.A.L. Heptaketides from Corynespora sp. inhabiting the cavern beard lichen, Usnea cavernosa: First report of metabolites of an endolichenic fungus. J. Nat. Prod. 2007, 70, 1700–1705. [Google Scholar] [CrossRef]























































Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, D.; Xue, M.; Shen, Z.; Jia, X.; Hou, X.; Lai, D.; Zhou, L. Phytotoxic Secondary Metabolites from Fungi. Toxins 2021, 13, 261. https://doi.org/10.3390/toxins13040261
Xu D, Xue M, Shen Z, Jia X, Hou X, Lai D, Zhou L. Phytotoxic Secondary Metabolites from Fungi. Toxins. 2021; 13(4):261. https://doi.org/10.3390/toxins13040261
Chicago/Turabian StyleXu, Dan, Mengyao Xue, Zhen Shen, Xiaowei Jia, Xuwen Hou, Daowan Lai, and Ligang Zhou. 2021. "Phytotoxic Secondary Metabolites from Fungi" Toxins 13, no. 4: 261. https://doi.org/10.3390/toxins13040261
APA StyleXu, D., Xue, M., Shen, Z., Jia, X., Hou, X., Lai, D., & Zhou, L. (2021). Phytotoxic Secondary Metabolites from Fungi. Toxins, 13(4), 261. https://doi.org/10.3390/toxins13040261