Effect of Indian Polyvalent Antivenom in the Prevention and Reversal of Local Myotoxicity Induced by Common Cobra (Naja naja) Venom from Sri Lanka In Vitro


 ,
,  and
and 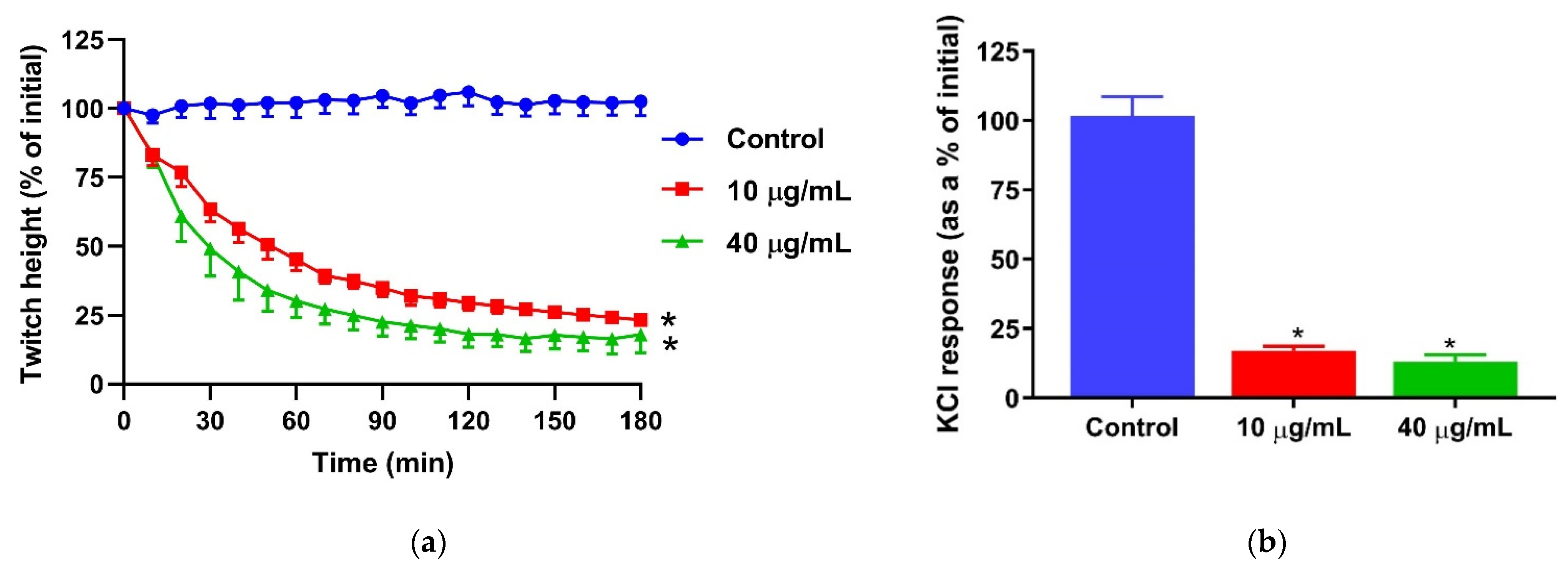{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. In Vitro Muscle Injury Caused by Naja naja Venom
2.2. Prevention of the Naja naja Venom-Mediated In Vitro Muscle Injury by Indian Polyvalent Antivenoms
2.3. Reversibility of Naja naja Venom-Mediated In Vitro Muscle Injury by Indian Polyvalent Antivenoms
2.4. Reversibility of Naja naja Venom-Mediated In Vitro Muscle Injury by Washing the Preparation
3. Discussion
4. Materials and Methods
4.1. Venom
4.2. Antivenom
4.3. Chick Biventer Cervicis Nerve-Muscle Preparation
4.4. Data Analysis and Statistics
4.5. Animal Ethics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gutiérrez, J.M.; Calvete, J.J.; Habib, A.G.; Harrison, R.A.; Williams, D.J.; Warrell, D.A. Snakebite envenoming. Nat. Rev. Dis. Prim. 2017, 3, 17063. [Google Scholar] [CrossRef]
- Kasturiratne, A.; Wickremasinghe, A.R.; De Silva, N.; Gunawardena, N.K. The Global Burden of Snakebite: A Literature Analysis and Modelling Based on Regional Estimates of Envenoming and Deaths. PLoS Med. 2008, 5, e218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ralph, R.; Sharma, S.K.; Faiz, M.A.; Ribeiro, I.; Rijal, S.; Chappuis, F.; Kuch, U. The timing is right to end snakebite deaths in South Asia. BMJ 2019, 364, k5317. [Google Scholar] [CrossRef] [Green Version]
- Waiddyanatha, S.; Silva, A.; Siribaddana, S.; Isbister, G. Long-term Effects of Snake Envenoming. Toxins 2019, 11, 193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organisation. Guidelines for the Prevention and Clinical Management of Snakebite in Africa; World Health Organization, Regional Office for Africa: Brazzaville, Republic of the Congo, 2010. [Google Scholar]
- WHO/Regional Office for South-East Asia. Guidelines for the Management of Snakebites, 2nd ed.; Regional Office for South-East Asia, World Health Organisation: New Delhi, India, 2016. [Google Scholar]
- Kini, R.M. Evolution of Three-Finger Toxins—A Versatile Mini Protein Scaffold. Acta Chim. Slov. 2011, 58, 693–701. [Google Scholar]
- Kini, R.M. Molecular moulds with multiple missions: Functional sites in three-finger toxins. Clin. Exp. Pharmacol. Physiol. 2002, 29, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Dubovskii, P.; Konshina, A.; Efremov, R. Cobra cardiotoxins: Membrane interactions and pharmacological potential. Curr. Med. Chem. 2014, 31, 270–287. [Google Scholar] [CrossRef] [PubMed]
- Konshina, A.G.; Krylov, N.A.; Efremov, R.G. Cardiotoxins: Functional Role of Local Conformational Changes. J. Chem. Inf. Model. 2017, 57, 2799–2810. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Ownby, C.L. Skeletal muscle degeneration induced by venom phospholipases A2: Insights into the mechanisms of local and systemic myotoxicity. Toxicon 2003, 42, 915–931. [Google Scholar] [CrossRef]
- Kularatne, S.A.M.; Budagoda, B.D.S.S.; Gawarammana, I.B.; Kularatne, W.K.S. Epidemiology, clinical profile and management issues of cobra (Naja naja) bites in Sri Lanka: First authenticated case series. Trans. R. Soc. Trop. Med. Hyg. 2009, 103, 924–930. [Google Scholar] [CrossRef]
- Alirol, E.; Sharma, S.K.; Bawaskar, H.S.; Kuch, U.; Chappuis, F. Snake bite in South Asia: A review. PLoS Negl. Trop. Dis. 2010, 4, e603. [Google Scholar] [CrossRef] [Green Version]
- Bawaskar, H.; Bawaskar, P.; Punde, D.; Inamdar, M.; Dongare, R.; Bhoite, R. Profile of Snakebite Envenoming in Rural Maharashtra, India. J. Assoc. Physicians India 2008, 56, 88–95. [Google Scholar] [PubMed]
- Sintiprungrat, K.; Watcharatanyatip, K.; Senevirathne, W.D.S.T.; Chaisuriya, P.; Chokchaichamnankit, D.; Srisomsap, C.; Ratanabanangkoon, K. A comparative study of venomics of Naja naja from India and Sri Lanka, clinical manifestations and antivenomics of an Indian polyspecific antivenom. J. Proteom. 2016, 132, 131–143. [Google Scholar] [CrossRef]
- Silva, A.; Isbister, G.K. Current research into snake antivenoms, their mechanisms of action and applications. Biochem. Soc. Trans. 2020, 48, 537–546. [Google Scholar] [CrossRef] [Green Version]
- Mao, Y.C.; Liu, P.Y.; Chiang, L.C.; Lai, C.S.; Lai, K.L.; Ho, C.H.; Wang, T.H.; Yang, C.C. Naja atra snakebite in Taiwan. Clin. Toxicol. 2018, 56, 273–280. [Google Scholar] [CrossRef]
- Wong, O.F.; Lam, T.S.K.; Fung, H.T.; Choy, C.H. Five-year experience with Chinese cobra (Naja atra)-related injuries in two acute hospitals in Hong Kong. Hong Kong Med. J. 2010, 16, 36–43. [Google Scholar]
- Maduwage, K.; Silva, A.; O’Leary, M.A.; Hodgson, W.C.; Isbister, G.K. Efficacy of Indian polyvalent snake antivenoms against Sri Lankan snake venoms: Lethality studies or clinically focussed in vitro studies. Sci. Rep. 2016, 6, 26778. [Google Scholar] [CrossRef] [Green Version]
- Leong, P.K.; Tan, N.H.; Fung, S.Y.; Sim, S.M. Cross neutralisation of Southeast Asian cobra and krait venoms by Indian polyvalent antivenoms. Trans. R. Soc. Trop. Med. Hyg. 2012, 106, 731–737. [Google Scholar] [CrossRef]
- Theakston, R.D.G.; Phillips, R.E.; Warrell, D.A.; Galagedera, Y.; Abeysekera, D.T.D.J.; Dissanayaka, P.; de Silva, A.; Aloysius, D.J.D.; Galagedara, Y.; Abeysekara, D.; et al. Envenoming by the common krait (Bungarus caeruleus) and Sri Lankan cobra (Naja naja naja): Efficacy and complications of therapy with Haffkine antivenom. Trans. R. Soc. Trop. Med. Hyg. 1990, 84, 301–308. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; León, G.; Lomonte, B. Pharmacokinetic-pharmacodynamic relationships of immunoglobulin therapy for envenomation. Clin. Pharmacokinet. 2003, 42, 721–741. [Google Scholar] [CrossRef]
- Chippaux, J.-P.; Goyffon, M. Venoms, Antivenoms and Immunotherapy. Toxicon 1998, 36, 823–846. [Google Scholar] [CrossRef]
- World Health Organisation. Guidelines for the Production, Control and Regulation of Snake Antivenom Immunoglobulins (Replacement of Annex 2 of WHO Technical Report Series, No. 964); World Health Organisation: Geneva, Switzerland, 2017. [Google Scholar]
- Gutiérrez, J.M.; Solano, G.; Pla, D.; Herrera, M.; Segura, Á.; Vargas, M.; Villalta, M.; Sánchez, A.; Sanz, L.; Lomonte, B.; et al. Preclinical evaluation of the efficacy of antivenoms for snakebite envenoming: State-of-the-art and challenges ahead. Toxins 2017, 9, 163. [Google Scholar] [CrossRef] [Green Version]
- Hart, A.J.; Hodgson, W.C.; O’leary, M.; Isbister, G.K. Pharmacokinetics and pharmacodynamics of the myotoxic venom of Pseudechis australis (mulga snake) in the anesthetised rat. Clin. Toxicol. 2014, 52, 604–610. [Google Scholar] [CrossRef] [PubMed]
- Madhushani, U.; Isbister, G.K.; Tasoulis, T.; Hodgson, W.C.; Silva, A. In-Vitro Neutralization of the Neurotoxicity of Coastal Taipan Venom by Australian Polyvalent Antivenom: The Window of Opportunity. Toxins 2020, 12, 690. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Madhushani, U.; Thakshila, P.; Hodgson, W.C.; Isbister, G.K.; Silva, A. Effect of Indian Polyvalent Antivenom in the Prevention and Reversal of Local Myotoxicity Induced by Common Cobra (Naja naja) Venom from Sri Lanka In Vitro. Toxins 2021, 13, 308. https://doi.org/10.3390/toxins13050308
Madhushani U, Thakshila P, Hodgson WC, Isbister GK, Silva A. Effect of Indian Polyvalent Antivenom in the Prevention and Reversal of Local Myotoxicity Induced by Common Cobra (Naja naja) Venom from Sri Lanka In Vitro. Toxins. 2021; 13(5):308. https://doi.org/10.3390/toxins13050308
Chicago/Turabian StyleMadhushani, Umesha, Prabhani Thakshila, Wayne C. Hodgson, Geoffrey K. Isbister, and Anjana Silva. 2021. "Effect of Indian Polyvalent Antivenom in the Prevention and Reversal of Local Myotoxicity Induced by Common Cobra (Naja naja) Venom from Sri Lanka In Vitro" Toxins 13, no. 5: 308. https://doi.org/10.3390/toxins13050308
APA StyleMadhushani, U., Thakshila, P., Hodgson, W. C., Isbister, G. K., & Silva, A. (2021). Effect of Indian Polyvalent Antivenom in the Prevention and Reversal of Local Myotoxicity Induced by Common Cobra (Naja naja) Venom from Sri Lanka In Vitro. Toxins, 13(5), 308. https://doi.org/10.3390/toxins13050308