The Effects of Mixed Fusarium Mycotoxins at EU-Permitted Feed Levels on Weaned Piglets’ Tissue Lipids

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Growth Performance, Absolute and Relative Organ Weights
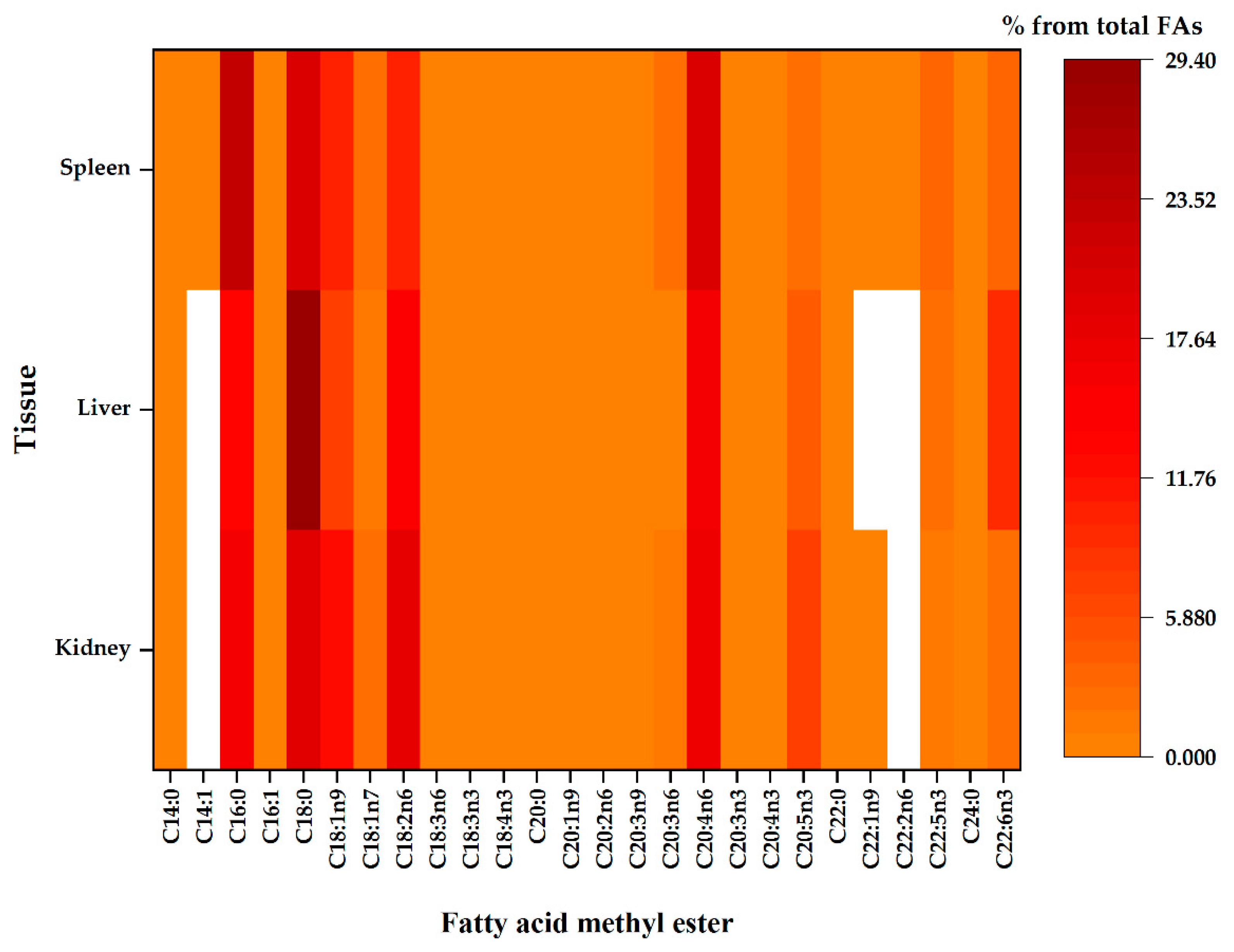
2.2. Kidney Total Phospholipid Fatty Acid Profile
2.3. The Liver Total Phospholipid Fatty Acid Profile
2.4. The Spleen Total Phospholipid Fatty Acid Profile
2.5. Plasma Nitrogenous Compounds
2.6. Plasma Lipid Metabolites and Glucose
2.7. Plasma Enzyme Activities
2.8. The Plasma Ion Concentrations
2.9. Antioxidants and Lipid Peroxidation
3. Discussion
3.1. Growth Performance, Absolute and Relative Organ Weights
3.2. The Kidney Total Phospholipid Fatty Acid Profile
3.3. The liver Total Phospholipid Fatty Acid Profile
3.4. The Spleen Total Phospholipid Fatty Acid Profile
3.5. Plasma Nitrogenous Compounds
3.6. Plasma Lipid Metabolites and Glucose
3.7. Plasma Enzyme Activities
3.8. Plasma Ion Concentrations
3.9. Antioxidant Enzymes and Lipid Peroxidation End Product
4. Conclusions
5. Materials and Methods
5.1. Experimental Animals and Diets
5.2. Mycotoxin Production
5.3. Sample Collections
5.4. Lipid Analysis
5.5. Serum Clinical Chemistry Analysis
5.6. Assessment of Antioxidant Capacity
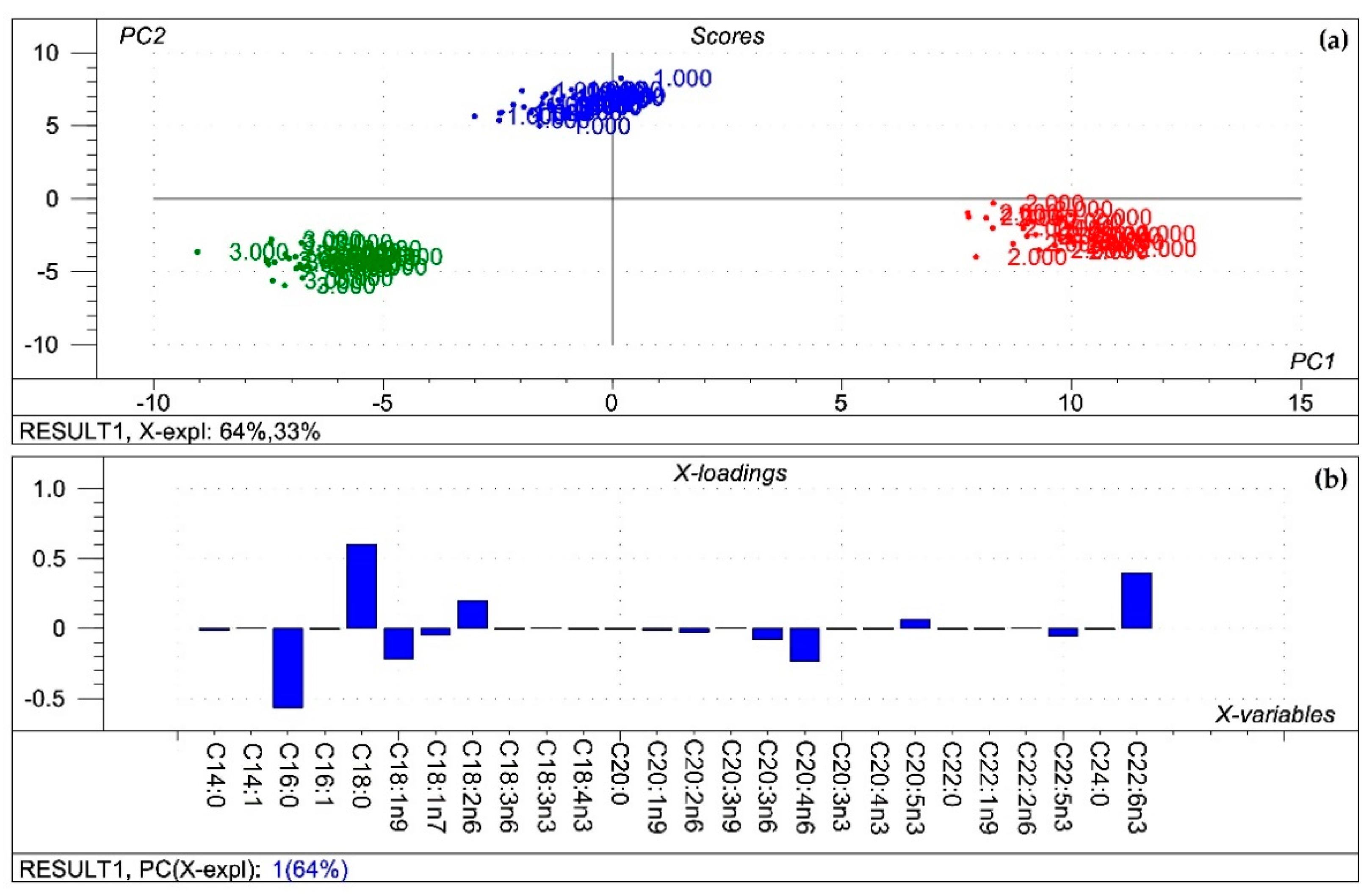
5.7. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marroquín-Cardona, A.G.; Johnson, N.M.; Phillips, T.D.; Hayes, A.W. Mycotoxins in a changing global environment—A review. Food Chem. Toxicol. 2014, 69, 220–230. [Google Scholar] [CrossRef]
- Christiane Gruber-Dorninger, T.; Jenkins, G.S. Global Mycotoxin Occurrence in Feed: A Ten-Year Survey. Toxins 2019, 11, 1–25. [Google Scholar]
- Wu, F.; Groopman, J.D.; Pestka, J.J. Public Health Impacts of Foodborne Mycotoxins. Annu. Rev. Food Sci. Technol. 2014, 5, 351–372. [Google Scholar] [CrossRef] [Green Version]
- Pestka, J.J. Deoxynivalenol: Mechanisms of action, human exposure, and toxicological relevance. Arch. Toxicol. 2010, 84, 663–679. [Google Scholar] [CrossRef]
- Pierron, A.; Alassane-Kpembi, I.; Oswald, I.P. Impact of two mycotoxins deoxynivalenol and fumonisin on pig intestinal health. Porc. Heal. Manag. 2016, 2, 1–8. [Google Scholar] [CrossRef]
- Zhang, G.-L.; Feng, Y.-L.; Song, J.-L.; Zhou, X.-S. Zearalenone: A Mycotoxin With Different Toxic Effect in Domestic and Laboratory Animals’ Granulosa Cells. Front. Genet. 2018, 9, 667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Zhu, D.; Guo, T.; Zhang, Y.; Shi, B.; Shan, A.; Chen, Z. Toxicity of zearalenone on the intestines of pregnant sows and their offspring and alleviation with modified halloysite nanotubes. J. Sci. Food Agric. 2018, 98, 698–706. [Google Scholar] [CrossRef] [PubMed]
- Marin, D.E.; Pistol, G.C.; Neagoe, I.V.; Calin, L.; Taranu, I. Effects of zearalenone on oxidative stress and inflammation in weanling piglets. Food Chem. Toxicol. 2013, 58, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Zinedine, A.; Soriano, J.M.; Moltó, J.C.; Mañes, J. Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: An oestrogenic mycotoxin. Food Chem. Toxicol. 2007, 45, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Voss, K.A.; Riley, R.T. Fumonisin Toxicity and Mechanism of Action: Overview and current perspectives. Food Saf. 2013, 1, 2013006. [Google Scholar] [CrossRef] [Green Version]
- Haschek, W.M.; Gumprecht, L.A.; Smith, G.; Tumbleson, M.E.; Constable, P.D. Fumonisin toxicosis in swine: An overview of porcine pulmonary edema and current perspectives. Environ. Health Perspect. 2001, 109, 251–257. [Google Scholar] [PubMed]
- European Commission. Commission Recommendation of 17 August 2006 on the presence of deoxynivalenol, zearalenone, ochratoxin A, T-2 and HT-2 and fumonisins in products intended for animal feeding. Off. J. Eur. Union 2006, 299, 7–9. [Google Scholar]
- Battilani, P.; Palumbo, R.; Giorni, P.; Dall’Asta, C.; Dellafiora, L.; Gkrillas, A.; Toscano, P.; Crisci, A.; Brera, C.; De Santis, B.; et al. Mycotoxin mixtures in food and feed: Holistic, innovative, flexible risk assessment modelling approach: EFSA Support. Publ. 2020, 17. [Google Scholar] [CrossRef] [Green Version]
- Kócsó, D.J.; Ali, O.; Kovács, M.; Mézes, M.; Balogh, K.; Kachlek, M.L.; Bóta, B.; Zeebone, Y.Y.; Szabó, A. A preliminary study on changes in heat shock protein 70 levels induced by Fusarium mycotoxins in rats: In vivo study. Mycotoxin Res. 2021, 37, 141–148. [Google Scholar] [CrossRef]
- Szabó, A.; Szabó-Fodor, J.; Fébel, H.; Mézes, M.; Balogh, K.; Bázár, G.; Kocsó, D.; Ali, O.; Kovács, M. Individual and combined effects of fumonisin B1, deoxynivalenol and zearalenone on the hepatic and renal membrane lipid integrity of rats. Toxins 2018, 10, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabó-Fodor, J.; Szabó, A.; Kócsó, D.; Marosi, K.; Bóta, B.; Kachlek, M.; Mézes, M.; Balogh, K.; Kövér, G.; Nagy, I.; et al. Interaction between the three frequently co-occurring Fusarium mycotoxins in rats. J. Anim. Physiol. Anim. Nutr. 2018, 103, 370–382. [Google Scholar] [CrossRef] [Green Version]
- Alassane-Kpembi, I.; Schatzmayr, G.; Taranu, I.; Marin, D.; Puel, O.; Oswald, I.P. Mycotoxins co-contamination: Methodological aspects and biological relevance of combined toxicity studies. Crit. Rev. Food Sci. Nutr. 2017, 57, 3489–3507. [Google Scholar] [CrossRef]
- Smith, M.C.; Madec, S.; Coton, E.; Hymery, N. Natural Co-occurrence of mycotoxins in foods and feeds and their in vitro combined toxicological effects. Toxins 2016, 8, 94. [Google Scholar] [CrossRef] [PubMed]
- Escrivá, L.; Font, G.; Manyes, L. In vivo toxicity studies of Fusarium mycotoxins in the last decade: A review. Food Chem. Toxicol. 2015, 78, 185–206. [Google Scholar] [CrossRef]
- Fodor, J.S. Individual and combined effects of subchronic exposure of three Fusarium toxins (fumonisin B, deoxynivalenol and zearalenone) in rabbit bucks. J. Clin. Toxicol. 2015, 5. [Google Scholar] [CrossRef] [Green Version]
- Grenier, B.; Oswald, I.P. Mycotoxin co-contamination of food and feed: Meta-analysis of publications describing toxicological interactions. World Mycotoxin J. 2011, 4, 285–313. [Google Scholar] [CrossRef]
- Qiang, Z.; Truong, M.; Meynen, K.; Murphy, P.A.; Hendrich, S. Efficacy of a mycotoxin binder against dietary fumonisin, deoxynivalenol, and zearalenone in rats. J. Agric. Food Chem. 2011, 59, 7527–7533. [Google Scholar] [CrossRef] [Green Version]
- Burger, H.M.; Abel, S.; Gelderblom, W.C.A. Modulation of key lipid raft constituents in primary rat hepatocytes by fumonisin B1—Implications for cancer promotion in the liver. Food Chem. Toxicol. 2018, 115, 34–41. [Google Scholar] [CrossRef]
- Lee, C.H.; Olson, P.; Evans, R.M. Minireview: Lipid metabolism, metabolic diseases, and peroxisome proliferator-activated receptors. Endocrinology 2003, 144, 2201–2207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabó, A.; Fébel, H.; Ali, O.; Kovács, M. Fumonisin B1 induced compositional modifications of the renal and hepatic membrane lipids in rats–Dose and exposure time dependence. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2019, 36, 1722–1739. [Google Scholar] [CrossRef]
- Riedel, S.; Abel, S.; Burger, H.M.; van der Westhuizen, L.; Swanevelder, S.; Gelderblom, W.C.A. Differential modulation of the lipid metabolism as a model for cellular resistance to fumonisin B 1 -induced cytotoxic effects in vitro. Prostaglandins Leukot. Essent. Fat. Acids 2016, 109, 39–51. [Google Scholar] [CrossRef]
- Szabó, A.; Szabó-Fodor, J.; Fébel, H.; Mézes, M.; Repa, I.; Kovács, M. Acute hepatic effects of low-dose fumonisin B 1 in rats. Acta Vet. Hung. 2016, 64, 436–448. [Google Scholar] [CrossRef] [Green Version]
- Riedel, S.; Abel, S.; Swanevelder, S.; Gelderblom, W.C.A. Induction of an altered lipid phenotype by two cancer promoting treatments in rat liver. Food Chem. Toxicol. 2015, 78, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Loiseau, N.; Polizzi, A.; Dupuy, A.; Therville, N.; Rakotonirainy, M.; Loy, J.; Viadere, J.L.; Cossalter, A.M.; Bailly, J.D.; Puel, O.; et al. New insights into the organ-specific adverse effects of fumonisin B1: Comparison between lung and liver. Arch. Toxicol. 2015, 89, 1619–1629. [Google Scholar] [CrossRef]
- Burger, H.M.; Abel, S.; Snijman, P.W.; Swanevelder, S.; Gelderblom, W.C.A. Altered lipid parameters in hepatic subcellular membrane fractions induced by fumonisin B1. Lipids 2007, 42, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Gelderblom, W.C.A.; Moritz, W.; Swanevelder, S.; Smuts, C.M.; Abel, S. Lipids and Δ6-desaturase activity alterations in rat liver microsomal membranes induced by fumonisin B1. Lipids 2002, 37, 869–877. [Google Scholar] [CrossRef]
- Gelderblom, W.C.A.; Smuts, C.M.; Abel, S.; Snyman, S.D.; Van Der Westhuizen, L.; Huber, W.W.; Swanevelder, S. Effect of fumonisin B1 on the levels and fatty acid composition of selected lipids in rat liver in vivo. Food Chem. Toxicol. 1997, 35, 647–656. [Google Scholar] [CrossRef]
- Szabó, A.; Ali, O.; Lóki, K.; Balogh, K.; Mézes, M.; Bartók, T.; Horváth, L.; Kovács, M. Orally administered fumonisins affect porcine red cell membrane sodium pump activity and lipid profile without apparent oxidative damage. Toxins 2020, 12, 318. [Google Scholar] [CrossRef] [PubMed]
- Ali, O.; Szabó-Fodor, J.; Fébel, H.; Mézes, M.; Balogh, K.; Glávits, R.; Kovács, M.; Zantomasi, A.; Szabó, A. Porcine hepatic response to fumonisin b1 in a short exposure period: Fatty acid profile and clinical investigations. Toxins 2019, 11, 655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lonergan, S.M.; Topel, D.G.; Marple, D.N. Growth curves and growth patterns. In The Science of Animal Growth and Meat Technology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 71–109. [Google Scholar]
- Lugert, V.; Thaller, G.; Tetens, J.; Schulz, C.; Krieter, J. A review on fish growth calculation: Multiple functions in fish production and their specific application. Rev. Aquacult. 2016, 8, 30–42. [Google Scholar] [CrossRef]
- Pierron, A.; Alassane-Kpembi, I.; Oswald, I.P. Impact of mycotoxin on immune response and consequences for pig health. Anim. Nutr. 2016, 2, 63–68. [Google Scholar] [CrossRef]
- Kanora, A.; Maes, D. The role of mycotoxins in pig reproduction: A review. Vet. Med. 2009, 54, 565–576. [Google Scholar] [CrossRef] [Green Version]
- Jia, R.; Liu, W.; Zhao, L.; Cao, L.; Shen, Z. Low doses of individual and combined deoxynivalenol and zearalenone in naturally moldy diets impair intestinal functions via inducing inflammation and disrupting epithelial barrier in the intestine of piglets. Toxicol. Lett. 2020, 333, 159–169. [Google Scholar] [CrossRef]
- Bonnet, M.S.; Roux, J.; Mounien, L.; Dallaporta, M.; Troadec, J.D. Advances in deoxynivalenol toxicity mechanisms: The brain as a target. Toxins 2012, 4, 1120–1138. [Google Scholar] [CrossRef] [Green Version]
- Mayer, E.; Novak, B.; Springler, A.; Schwartz-Zimmermann, H.E.; Nagl, V.; Reisinger, N.; Hessenberger, S.; Schatzmayr, G. Effects of deoxynivalenol (DON) and its microbial biotransformation product deepoxy-deoxynivalenol (DOM-1) on a trout, pig, mouse, and human cell line. Mycotoxin Res. 2017, 33, 297–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gajęcka, M.; Tarasiuk, M.; Zielonka, Ł.; Dąbrowski, M.; Gajęcki, M. Risk assessment for changes in metabolic profile and body weight of pre-pubertal gilts during long-term monotonic exposure to low doses of zearalenone (ZEN). Res. Vet. Sci. 2016, 109, 169–180. [Google Scholar] [CrossRef]
- Gajęcka, M.; Zielonka, Ł.; Gajęcki, M. Activity of Zearalenone in the Porcine Intestinal Tract. Molecules 2017, 22, 18. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.Y.; Wang, H.L.; Cheng, X.L.; Wei, F.; Bai, X.; Lin, R.C.; Vaziri, N.D. Metabolomics analysis reveals the association between lipid abnormalities and oxidative stress, inflammation, fibrosis, and Nrf2 dysfunction in aristolochic acid-induced nephropathy. Sci. Rep. 2015, 5, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Terciolo, C.; Bracarense, A.P.; Souto, P.C.M.C.; Cossalter, A.M.; Dopavogui, L.; Loiseau, N.; Oliveira, C.A.F.; Pinton, P.; Oswald, I.P. Fumonisins at doses below EU regulatory limits induce histological alterations in piglets. Toxins 2019, 11, 548. [Google Scholar] [CrossRef] [Green Version]
- Dobrzyń, A.; Górski, J. Ceramides and sphingomyelins in skeletal muscles of the rat: Content and composition. Effect of prolonged exercise. Am. J. Physiol. Endocrinol. Metab. 2002, 282, 277–285. [Google Scholar] [CrossRef] [Green Version]
- Bondy, G.; Barker, M.; Mueller, R.; Fernie, S.; Miller, J.D.; Armstrong, C.; Hierlihy, S.L.; Rowsell, P.; Suzuki, C. Fumonisin B1 Toxicity in Male Sprague-Dawley Rats. In Fumonisins in Food; Jackson, L.S., DeVries, J.W., Bullerman, L.B., Eds.; Springer US: Boston, MA, USA, 1996; pp. 251–264. [Google Scholar]
- Soni, N.K.; Ross, A.B.; Scheers, N.; Savolainen, O.I.; Nookaew, I.; Gabrielsson, B.G.; Sandberg, A.S. Splenic immune response is down-regulated in C57BL/6J mice fed eicosapentaenoic acid and docosahexaenoic acid enriched high fat diet. Nutrients 2017, 9, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Portois, L.; Sener, A.; Carpentier, Y.A.; Malaisse, W.J. Fatty acid content and pattern of spleen phospholipids and triglycerides in normal and either type-1 or type-2 diabetic rats. Int. J. Mol. Med. 2007, 19, 523–528. [Google Scholar] [CrossRef] [Green Version]
- Fink-Gremmels, J.; Malekinejad, H. Clinical effects and biochemical mechanisms associated with exposure to the mycoestrogen zearalenone. Anim. Feed Sci. Technol. 2007, 137, 326–341. [Google Scholar] [CrossRef]
- Atroshi, F.; Rizzo, A.; Biese, I.; Veijalainen, P.; Salonemi, H.; Sankari, S.; Andersson, K. Fumonisin B1-Induced DNA damage in Rat Liver and Spleen: Effects of Pretreatment With Coenzyme Q10, -Carnitine, A-Tocopherol and Selenium. Pharmacol. Res. 1999, 40, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Marin, D.E.; Taranu, I.; Pascale, F.; Lionide, A.; Burlacu, R.; Bailly, J.-D.; Oswald, I.P. Sex-related differences in the immune response of weanling piglets exposed to low doses of fumonisin extract. Br. J. Nutr. 2006, 95, 1185–1192. [Google Scholar] [CrossRef] [PubMed]
- Szabó, A.; Szabó-Fodor, J.; Kachlek, M.; Mézes, M.; Balogh, K.; Glávits, R.; Ali, O.; Zeebone, Y.; Kovács, M. Dose and Exposure Time-Dependent Renal and Hepatic Effects of Intraperitoneally Administered Fumonisin B1 in Rats. Toxins 2018, 10, 465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kouadio, J.; Moukha, S.; Brou, K.; Gnakri, D. Lipid metabolism disorders, lymphocytes cells death, and renal toxicity induced by very low levels of deoxynivalenol and fumonisin B 1 alone or in combination following 7 days oral administration to mice. Toxicol. Int. 2013, 20, 218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Régnier, M.; Polizzi, A.; Lukowicz, C.; Smati, S.; Lasserre, F.; Lippi, Y.; Naylies, C.; Laffitte, J.; Bétoulières, C.; Montagner, A.; et al. The protective role of liver X receptor (LXR) during fumonisin B1-induced hepatotoxicity. Arch. Toxicol. 2019, 93, 505–517. [Google Scholar] [CrossRef]
- Dilkin, P.; Direito, G.; Simas, M.M.S.; Mallmann, C.A.; Corrêa, B. Chemico-Biological Interactions Toxicokinetics and toxicological effects of single oral dose of fumonisin B1 containing Fusarium verticillioides culture material in weaned piglets. Chem. Biol. Interact. 2010, 185, 157–162. [Google Scholar] [CrossRef]
- Gbore, F.A.; Egbunike, G.N. Toxicological evaluation of dietary fumonisin B1 on serum biochemistry of growing pigs. J. Cent. Eur. Agric. 2009, 10, 255–262. [Google Scholar]
- Gumprecht, L.A.; Beasley, V.R.; Weigel, R.M.; Parker, H.M.; Tumbleson, M.E.; Bacon, C.W.; Meredith, F.I.; Haschek, W.M. Development of fumonisin-induced hepatotoxicity and pulmonary edema in orally dosed swine: Morphological and biochemical alterations. Toxicol. Pathol. 1998, 26, 777–788. [Google Scholar] [CrossRef]
- Rotter, B.A.; Thompson, B.K.; Prelusky, D.B.; Trenholm, H.L.; Stewart, B.; Miller, J.D.; Savard, M.E. Response of growing swine to dietary exposure to pure fumonisin B1 during an eight-week period: Growth and clinical parameters. Nat. Toxins 1996, 4, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Nemesánszky, E.; Lott, J.A. Gamma-glutamyltransferase and its isoenzymes: Progress and problems. Clin. Chem. 1985, 31, 797–803. [Google Scholar] [CrossRef]
- Forman, H.; Lui, R.M.; Tian, L. Glutathione cycling in oxidative stress. Lung Biol. Health Dis 1997, 105, 99–121. [Google Scholar]
- Bohn, A.A. Laboratory evaluation of electrolytes. In Veterinary Hematology and Clinical Chemistry, 2nd ed; Thrall, M.A., Weiser, G., Allison, R.W., Campbell, T.W., Eds.; Wiley-Blackwell: Ames, IA, USA, 2012; pp. 387–392. ISBN 978-0-8138-1027-0. [Google Scholar]
- Taylor, J.G.; Bushinsky, D.A. Calcium and phosphorus homeostasis. Blood Purif. 2009, 27, 387–394. [Google Scholar] [CrossRef]
- El Golli-Bennour, E.; Bacha, H. Hsp70 expression as biomarkers of oxidative stress: Mycotoxins’ exploration. Toxicology 2011, 287, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wu, Q.; Wan, D.; Liu, Q.; Chen, D.; Liu, Z.; Martinez-Larrañaga, M.R.; Martínez, M.A.; Anadón, A.; Yuan, Z. Fumonisins: Oxidative stress-mediated toxicity and metabolism in vivo and in vitro. Arch. Toxicol. 2015, 90, 81–101. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Wang, F.; Yu, D.F.; Wu, P.F.; Chen, J.G. The cytotoxic mechanism of malondialdehyde and protective effect of carnosine via protein cross-linking/mitochondrial dysfunction/reactive oxygen species/MAPK pathway in neurons. Eur. J. Pharmacol. 2011, 650, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Poersch, A.B.; Trombetta, F.; Braga, A.C.M.; Boeira, S.P.; Oliveira, M.S.; Dilkin, P.; Mallmann, C.A.; Fighera, M.R.; Royes, L.F.F.; Oliveira, M.S.; et al. Involvement of oxidative stress in subacute toxicity induced by fumonisin B1 in broiler chicks. Vet. Microbiol. 2014, 174, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Lubos, E.; Loscalzo, J.; Handy, D.E. Glutathione peroxidase-1 in health and disease: From molecular mechanisms to therapeutic opportunities. Antioxidants Redox Signal. 2011, 15, 1957–1997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, T.; Yamamoto, M. Molecular basis of the Keap1-Nrf2 system. Free Radic. Biol. Med. 2015, 88, 93–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labunskyy, V.M.; Hatfield, D.L.; Gladyshev, V.N. Selenoproteins: Molecular pathways and physiological roles. Physiol. Rev. 2014, 94, 739–777. [Google Scholar] [CrossRef] [Green Version]
- Szabó, A.; Nagy, S.; Ali, O.; Gerencsér, Z.; Mézes, M.; Balogh, K.M.; Bartók, T.; Horváth, L.; Mou-hanna, A.; Kovács, M. A 65-Day Fumonisin B Exposure at High Die-tary Levels Has Negligible Effects on the Testicular and Spermatological Parameters of Adult Rabbit Bucks. Toxins 2021, 13, 237. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Leray, C.; Andriamampandry, M.; Gutbier, G.; Cavadenti, J.; Klein-Soyer, C.; Gachet, C.; Cazenave, J.P. Quantitative analysis of vitamin E, cholesterol and phospholipid fatty acids in a single aliquot of human platelets and cultured endothelial cells. J. Chromatogr. B Biomed. Appl. 1997, 696, 33–42. [Google Scholar] [CrossRef]
- Christie, W.W. A simple procedure for rapid transmethylation of glycerolipids and cholesteryl esters. J. Lipid Res. 1982, 23, 1072–1075. [Google Scholar] [CrossRef]
- Hung, C.; Bronec, C.; Napoli, E.; Graham, J.; Stanhope, K.L.; Marsilio, I.; Giron, M.C.; Havel, P.J.; Giulivi, C. Adipose depot-specific effects of ileal interposition surgery in UCD-T2D rats: Unexpected implications for obesity and diabetes. Biochem. J. 2018, 475, 649–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Placer, Z.A.; Cushman, L.L.; Johnson, B.C. Estimation of product of lipid peroxidation (malonyl dialdehyde) in biochemical systems. Anal. Biochem. 1966, 16, 359–364. [Google Scholar] [CrossRef]
- Sedlak, J.; Lindsay, R.H. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman’s reagent. Anal. Biochem. 1968, 25, 192–205. [Google Scholar] [CrossRef]
- Lawrence, R.A.; Burk, R.F. Species, tissue and subcellular distribution of non Se-dependent glutathione peroxidase activity. J. Nutr. 1978, 108, 211–215. [Google Scholar] [CrossRef]
- Weichselbaum, C.T.E. An Accurate and rapid method for the determination of proteins in small amounts of blood serum and plasma. Am. J. Clin. Pathol. 1946, 16, 40–49. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein mesurement with the phenol reagent. J. Biol. Chem. 1951, 193, 165–167. [Google Scholar] [CrossRef]
- Bliss, C.I. The toxicity of poisons applied jointly. Ann. Appl. Biol. 1939, 26, 585–615. [Google Scholar] [CrossRef]
- Kifer, D.; Jakšić, D.; Šegvić Klarić, M. Assessing the effect of mycotoxin combinations: Which mathematical model is (the Most) appropriate? Toxins 2020, 12, 153. [Google Scholar] [CrossRef] [Green Version]
- Multivariate Analysis Software. CAMO Software AS. Nedre Vollgate 8, N-0158; The Unscrambler: Oslo, Norway, 2006. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Parameter | Control | FB | DZ | FDZ |
|---|---|---|---|---|
| M ± SD | M ± SD | M ± SD | M ± SD | |
| Initial BW | 12.2 ± 0.61 | 12.8 ± 0.54 | 12.7 ± 0.67 | 12.4 ± 0.87 |
| 1-AWG | 2.59 ± 0.34 c | 2.58 ± 0.20 b | 2.67 ± 0.24 b | 2.62 ± 0.21 |
| 2-AWG | 1.98 ± 0.28 b | 1.95 ± 0.19 a | 1.98 ± 0.44 a | 2.22 ± 0.25 |
| 3-AWG | 1.55 ± 0.23 Aa | 2.45 ± 0.41 bB | 2.40 ± 0.61 abB | 2.40 ± 0.92 B |
| Final BW | 18.3 ± 0.69 A | 19.8 ± 0.61 B | 19.7 ± 1.07 B | 19.7 ± 1.32 B |
| Heart (g) | 110 ± 13.0 | 110 ± 8.18 | 113 ± 6.47 | 107 ± 8.61 |
| Liver (g) | 406 ± 45.6 B | 398 ± 14.1 B | 379 ± 19.8 AB | 361 ± 23.3 A |
| Kidney (g) | 89.8 ± 5.97 | 93.4 ± 4.40 | 85.6 ± 8.10 | 92.7 ± 16.9 |
| Lung (g) | 226 ± 55.8 | 223 ± 31.3 | 225 ± 46.4 | 201 ± 44.8 |
| Spleen (g) | 35.0 ± 5.14 | 36.4 ± 2.26 | 36.1 ± 5.91 | 36.1 ± 5.61 |
| Rel. heart (%) | 0.60 ± 0.05 | 0.56 ± 0.04 | 0.57 ± 0.03 | 0.55 ± 0.04 |
| Rel. liver (%) | 2.22 ± 0.24 B | 2.01 ± 0.05 AB | 1.93 ± 0.11 A | 1.85 ± 0.18 A |
| Rel. kidney (%) | 0.49 ± 0.03 | 0.47 ± 0.03 | 0.44 ± 0.05 | 0.47 ± 0.07 |
| Rel. lung (%) | 1.24 ± 0.32 | 1.13 ± 0.15 | 1.15 ± 0.27 | 1.01 ± 0.16 |
| Rel. spleen (%) | 0.19 ± 0.03 | 0.19 ± 0.01 | 0.18 ± 0.03 | 0.19 ± 0.03 |
| Parameter | Period | Control | FB | DZ | FDZ |
|---|---|---|---|---|---|
| M ± SD | M ± SD | M ± SD | M ± SD | ||
| FC (g/week) | overall | 12,201 ± 469 | 12,463 ± 89.4 | 12,492 ± 19.6 | 12,499 ± 1.71 |
| 1-week | 3930 ± 414 | 4063 ± 89.4 a | 4092 ± 19.6 a | 4099 ± 1.71 a | |
| 2-week | 4163 ± 91.3 | 4200 ± 0.00 b | 4200 ± 0.00 b | 4200 ± 0.00 b | |
| 3-week | 4108 ± 225 | 4200 ± 0.00 b | 4200 ± 0.00 b | 4200 ± 0.00 b | |
| FCR (g diet/g BW) | overall | 2.00 ± 0.08 A | 1.79 ± 0.10 B | 1.78 ± 0.14 B | 1.75 ± 0.19 B |
| 1-week | 1.53 ± 0.06 c | 1.58 ± 0.10 b | 1.54 ± 0.14 b | 1.58 ± 0.15 | |
| 2-week | 2.13 ± 0.30 b | 2.17 ± 0.19 a | 2.23 ± 0.65 a | 1.91 ± 0.19 | |
| 3-week | 2.69 ± 0.34 aA | 1.76 ± 0.33 bB | 1.84 ± 0.41 abB | 1.91 ± 0.53 B |
| Fatty Acid | Control | FB | DZ | FDZ |
|---|---|---|---|---|
| M ± SD | M ± SD | M ± SD | M ± SD | |
| C14:0 | 0.14 ± 0.02 a | 0.17 ± 0.02 b | 0.14 ± 0.01 ab | 0.17 ± 0.02 ab |
| C16:0 | 16.2 ± 0.76 a | 17.4 ± 0.59 b | 16.8 ± 0.42 ab | 17. 7 ± 0.48 b |
| C16:1 | 0.27 ± 0.03 a | 0.32 ± 0.04 ab | 0.33 ± 0.03 ab | 0.33 ± 0.03 b |
| C18:0 | 19.1 ± 0.57 | 19.4 ± 0.57 | 19.2 ± 0.21 | 19.5 ± 0.25 |
| C18:1n9 | 12.5 ± 0.40 | 12.1 ± 0.64 | 11.9 ± 0.64 | 12.6 ± 0.46 |
| C18:1n7 | 1.99 ± 0.05 | 2.03 ± 0.20 | 2.15 ± 0.14 | 2.09 ± 0.15 |
| C18:2n6 | 17.9 ± 0.91 | 18.6 ± 0.72 | 18.6 ± 0.41 | 18.3 ± 0.49 |
| C18:3n6 | 0.08 ± 0.01 | 0.09 ± 0.01 | 0.08 ± 0.01 | 0.10 ± 0.02 |
| C18:3n3 | 0.31 ± 0.14 | 0.39 ± 0.03 | 0.38 ± 0.03 | 0.30 ± 0.17 |
| C18:4n3 | 0.11 ± 0.13 | 0.05 ± 0.01 | 0.03 ± 0.02 | 0.10 ± 0.15 |
| C20:0 | 0.12 ± 0.03 b | 0.07 ± 0.01 a | 0.08 ± 0.01 a | 0.09 ± 0.01 a |
| C20:1n9 | 0.25 ± 0.05 | 0.22 ± 0.04 | 0.23 ± 0.03 | 0.22 ± 0.01 |
| C20:2n6 | 0.75 ± 0.06 b | 0.71 ± 0.03 ab | 0.70 ± 0.08 ab | 0.63 ± 0.05 a |
| C20:3n9 | 0.09 ± 0.01 | 0.09 ± 0.01 | 0.09 ± 0.00 | 0.09 ± 0.01 |
| C20:3n6 | 1.11 ± 0.07 | 1.23 ± 0.14 | 1.17 ± 0.09 | 1.27 ± 0.26 |
| C20:4n6 | 17.2 ± 0.85 | 16.7 ± 1.23 | 17.5 ± 0.63 | 16.6 ± 0.89 |
| C20:3n3 | 0.20 ± 0.02 b | 0.18 ± 0.03 ab | 0.19 ± 0.03 ab | 0.16 ± 0.01 a |
| C20:4n3 | 0.09 ± 0.01 | 0.11 ± 0.03 | 0.10 ± 0.01 | 0.11 ± 0.04 |
| C20:5n3 | 7.23 ± 0.52 | 6.34 ± 0.47 | 6.61 ± 0.72 | 6.50 ± 0.59 |
| C22:0 | 0.18 ± 0.07 b | 0.07 ± 0.03 a | 0.10 ± 0.01 a | 0.07 ± 0.01 a |
| C22:1n9 | 0.04 ± 0.01 b | 0.03 ± 0.01 a | 0.04 ± 0.01 ab | 0.03 ± 0.01 a |
| C22:5n3 | 1.45 ± 0.15 b | 1.47 ± 0.22 b | 1.23 ± 0.04 ab | 1.14 ± 0.09 a |
| C24:0 | 0.12 ± 0.02 b | 0.09 ± 0.04 ab | 0.06 ± 0.03 a | 0.07 ± 0.03 ab |
| C22:6n3 | 2.60 ± 0.25 b | 2.18 ± 0.17 a | 2.30 ± 0.15 ab | 2.24 ± 0.30 ab |
| SFA | 35.8 ± 0.31 a | 37.3 ± 0.54 b | 36.4 ± 0.29 a | 37.5 ± 0.66 b |
| UFA | 64.2 ± 0.31 b | 62.7 ± 0.54 a | 63.6 ± 0.29 b | 62.5 ± 0.66 a |
| MUFA | 15.1 ± 0.49 | 14.8 ± 0.91 | 14.8 ± 0.77 | 15.3 ± 0.59 |
| PUFA | 49.2 ± 0.52 b | 48.1 ± 0.82 ab | 49.0 ± 0.78 b | 47.3 ± 0.75 a |
| PUFA/MUFA | 3.26 ± 0.13 | 3.27 ± 0.25 | 3.32 ± 0.21 | 3.10 ± 0.15 |
| n-3 | 12.0 ± 0.43 b | 10.7 ± 0.51 a | 10.8 ± 0.73 a | 10.3 ± 0.85 a |
| n-6 | 37.1 ± 0.55 ab | 37.3 ± 0.89 ab | 38.0 ± 0.40 b | 36.9 ± 0.48 a |
| n-6/n-3 | 3.10 ± 0.14 a | 3.49 ± 0.22 ab | 3.52 ± 0.26 b | 3.59 ± 0.32 b |
| UI | 183 ± 1.95 c | 176 ± 3.09 ab | 179 ± 3.19 bc | 173 ± 1.94 a |
| ACL | 19.0 ± 0.19 b | 18.6 ± 0.29 ab | 18.4 ± 0.18 a | 18.4 ± 0.21 a |
| Fatty Acid | Control | FB | DZ | FDZ |
|---|---|---|---|---|
| M ± SD | M ± SD | M ± SD | M ± SD | |
| C14:0 | 0.07 ± 0.01 a | 0.10 ± 0.01 b | 0.08 ± 0.01 ab | 0.08 ± 0.01 ab |
| C16:0 | 13.0 ± 0.63 | 13.2 ± 0.47 | 13.6 ± 0.55 | 13.3 ± 0.15 |
| C16:1 | 0.32 ± 0.04 a | 0.41 ± 0.04 b | 0.40 ± 0.04 b | 0.37 ± 0.04 ab |
| C18:0 | 29.4 ± 0.80 b | 28.1 ± 0.86 a | 28.1 ± 0.87 ab | 29.2 ± 0.38 ab |
| C18:1n9 | 7.05 ± 0.46 a | 8.28 ± 0.34 b | 7.58 ± 0.45 ab | 7.71 ± 0.55 ab |
| C18:1n7 | 1.31 ± 0.12 | 1.64 ± 0.28 | 1.57 ± 0.29 | 1.37 ± 0.14 |
| C18:2n6 | 14.6 ± 0.92 | 15.5 ± 0.68 | 14.5 ± 0.81 | 14.3 ± 0.44 |
| C18:3n6 | 0.13 ± 0.02 | 0.15 ± 0.04 | 0.15 ± 0.03 | 0.16 ± 0.04 |
| C18:3n3 | 0.21 ± 0.05 | 0.26 ± 0.05 | 0.21 ± 0.03 | 0.24 ± 0.03 |
| C18:4n3 | 0.03 ± 0.02 ab | 0.05 ± 0.03 b | 0.02 ± 0.01 a | 0.02 ± 0.00 ab |
| C20:0 | 0.04 ± 0.01 | 0.05 ± 0.01 | 0.04 ± 0.01 | 0.04 ± 0.00 |
| C20:1n9 | 0.12 ± 0.01 | 0.14 ± 0.01 | 0.12 ± 0.02 | 0.12 ± 0.01 |
| C20:2n6 | 0.42 ± 0.06 | 0.46 ± 0.03 | 0.42 ± 0.09 | 0.41 ± 0.05 |
| C20:3n9 | 0.12 ± 0.08 | 0.13 ± 0.04 | 0.16 ± 0.04 | 0.12 ± 0.02 |
| C20:3n6 | 0.73 ± 0.04 | 0.69 ± 0.13 | 0.73 ± 0.15 | 0.83 ± 0.13 |
| C20:4n6 | 16.4 ± 0.81 | 16.4 ± 0.60 | 16.8 ± 0.82 | 16.2 ± 0.69 |
| C20:3n3 | 0.05 ± 0.01 | 0.06 ± 0.01 | 0.05 ± 0.02 | 0.05 ± 0.01 |
| C20:4n3 | 0.02 ± 0.00 | 0.02 ± 0.00 | 0.02 ± 0.00 | 0.02 ± 0.00 |
| C20:5n3 | 3.94 ± 0.31 | 3.72 ± 0.24 | 3.46 ± 0.48 | 3.66 ± 0.44 |
| C22:0 | 0.05 ± 0.01 b | 0.03 ± 0.01 ab | 0.02 ± 0.00 a | 0.02 ± 0.00 a |
| C22:5n3 | 2.46 ± 0.33 | 2.63 ± 0.37 | 2.62 ± 0.20 | 2.48 ± 0.48 |
| C24:0 | 0.02 ± 0.01 | 0.02 ± 0.01 | 0.01 ± 0.00 | 0.01 ± 0.00 |
| C22:6n3 | 9.64 ± 0.75 b | 8.02 ± 0.61 a | 9.25 ± 0.42 b | 9.32 ± 0.44 b |
| SFA | 42.5 ± 0.48 b | 41.4 ± 0.59 a | 41.9 ± 0.64 ab | 42.6 ± 0.48 b |
| UFA | 57.5 ± 0.48 a | 58.6 ± 0.59 b | 58.1 ± 0.64 ab | 57.4 ± 0.48 a |
| MUFA | 8.80 ± 0.53 a | 10.5 ± 0.35 b | 9.67 ± 0.62 ab | 9.57 ± 0.67 ab |
| PUFA | 48.7 ± 0.66 | 48.1 ± 0.51 | 48.5 ± 0.50 | 47.8 ± 0.49 |
| PUFA/MUFA | 5.55 ± 0.38 b | 4.60 ± 0.17 a | 5.03 ± 0.35 ab | 5.02 ± 0.37 ab |
| n-3 | 16.3 ± 1.04 b | 14.8 ± 1.11 a | 15.6 ± 0.43 ab | 15.8 ± 0.63 ab |
| n-6 | 32.2 ± 1.00 | 33.2 ± 0.98 | 32.7 ± 0.64 | 31.9 ± 0.75 |
| n-6/n-3 | 1.98 ± 0.18 | 2.27 ± 0.25 | 2.09 ± 0.09 | 2.02 ± 0.12 |
| UI | 198 ± 3.86 b | 192 ± 4.06 a | 197 ± 2.08 ab | 195 ± 1.98 ab |
| ACL | 18.7 ± 0.03 b | 18.6 ± 0.03 a | 18.6 ± 0.02 ab | 18.6 ± 0.03 ab |
| Fatty Acid | Control | FB | DZ | FDZ |
|---|---|---|---|---|
| M ± SD | M ± SD | M ± SD | M ± SD | |
| C14:0 | 0.34 ± 0.03 | 0.33 ± 0.08 | 0.31 ± 0.02 | 0.34 ± 0.03 |
| C14:1 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.01 ± 0.01 |
| C16:0 | 22.9 ± 0.72 | 23.0 ± 1.41 | 22.4 ± 0.42 | 22.9 ± 0.56 |
| C16:1 | 0.41 ± 0.02 | 0.40 ± 0.04 | 0.41 ± 0.03 | 0.41 ± 0.04 |
| C18:0 | 19.7 ± 0.54 | 19.6 ± 0.87 | 19.8 ± 0.40 | 19.5 ± 0.74 |
| C18:1n9 | 10.6 ± 0.40 | 10.7 ± 0.61 | 10.4 ± 0.37 | 10.9 ± 0.23 |
| C18:1n7 | 2.15 ± 0.09 | 2.13 ± 0.15 | 2.31 ± 0.11 | 2.17 ± 0.15 |
| C18:2n6 | 10.6 ± 0.78 | 10.5 ± 0.46 | 9.89 ± 0.83 | 10.0 ± 0.64 |
| C18:3n6 | 0.17 ± 0.03 | 0.17 ± 0.02 | 0.18 ± 0.04 | 0.19 ± 0.01 |
| C18:3n3 | 0.10 ± 0.02 | 0.12 ± 0.01 | 0.11 ± 0.01 | 0.12 ± 0.02 |
| C18:4n3 | 0.08 ± 0.01 a | 0.11 ± 0.05 ab | 0.13 ± 0.04 ab | 0.18 ± 0.11 b |
| C20:0 | 0.13 ± 0.01 ab | 0.12 ± 0.01 a | 0.14 ± 0.20 b | 0.28 ± 0.02 ab |
| C20:1n9 | 0.30 ± 0.04 | 0.28 ± 0.02 | 0.30 ± 0.02 | 0.28 ± 0.02 |
| C20:2n6 | 0.81 ± 0.10 | 0.84 ± 0.06 | 0.83 ± 0.10 | 0.84 ± 0.06 |
| C20:3n9 | 0.10 ± 0.03 | 0.09 ± 0.02 | 0.11 ± 0.03 | 0.10 ± 0.01 |
| C20:3n6 | 2.08 ± 0.12 | 2.08 ± 0.13 | 2.05 ± 0.11 | 2.13 ± 0.19 |
| C20:4n6 | 20.1 ± 1.23 | 20.6 ± 0.61 | 21.3 ± 0.90 | 20.7 ± 0.60 |
| C20:3n3 | 0.08 ± 0.01 | 0.09 ± 0.01 | 0.08 ± 0.01 | 0.10 ± 0.01 |
| C20:4n3 | 0.03 ± 0.01 | 0.04 ± 0.00 | 0.03 ± 0.00 | 0.04 ± 0.01 |
| C20:5n3 | 2.01 ± 0.13 | 1.91 ± 0.15 | 1.77 ± 0.21 | 1.94 ± 0.25 |
| C22:0 | 0.13 ± 0.01 bc | 0.10 ± 0.02 ab | 0.14 ± 0.02 c | 0.09 ±0.02 a |
| C22:1n9 | 0.08 ± 0.02 | 0.07 ± 0.01 | 0.08 ± 0.01 | 0.08 ± 0.01 |
| C22:2n6 | 0.04 ± 0.01 | 0.04 ± 0.00 | 0.04 ± 0.01 | 0.04 ± 0.00 |
| C22:5n3 | 3.83 ± 0.06 | 3.64 ± 0.20 | 3.87 ± 0.37 | 3.64 ± 0.11 |
| C24:0 | 0.05 ± 0.01 ab | 0.05 ± 0.03 ab | 0.06 ± 0.03 b | 0.03 ± 0.01 a |
| C22:6n3 | 3.23 ± 0.16 | 2.98 ± 0.18 | 3.26 ± 0.22 | 3.20 ± 0.24 |
| SFA | 43.2 ± 0.36 | 43.2 ± 0.62 | 42.7 ± 0.55 | 42.9 ± 1.02 |
| UFA | 56.8 ± 0.36 | 56.8 ± 0.62 | 57.1 ± 0.55 | 57.1 ± 1.02 |
| MUFA | 13.6 ± 0.51 | 13.6 ± 0.62 | 13.5 ± 0.42 | 13.8 ± 0.37 |
| PUFA | 43.2 ± 0.51 | 43.2 ± 0.76 | 43.6 ± 0.40 | 43.2 ± 0.70 |
| PUFA/MUFA | 3.19 ± 0.15 | 3.19 ± 0.18 | 3.22 ± 0.11 | 3.13 ± 0.05 |
| n-3 | 9.37 ± 0.12 b | 8.88 ± 0.27 a | 9.25 ± 0.30 b | 9.21 ± 0.45 ab |
| n-6 | 33.7 ± 0.48 | 34.3 ± 0.80 | 34.2 ± 0.30 | 33.9 ± 0.47 |
| n-6/n-3 | 3.60 ± 0.06 a | 3.86 ± 0.17 b | 3.70 ± 0.13 ab | 3.69 ± 0.18 ab |
| UI | 173 ± 3.34 | 172 ± 2.79 | 176 ± 1.59 | 174 ± 2.17 |
| ACL | 18.3 ± 0.04 ab | 18.3 ± 0.05 a | 18.4 ± 0.01 b | 18.3 ± 0.01 ab |
| Parameter | Period | Control | FB | DZ | FDZ |
|---|---|---|---|---|---|
| M ± SD | M ± SD | M ± SD | M ± SD | ||
| Total Protein (g/L) | 1-week | 53.2 ± 1.33 a | 52.2 ± 2.86 | 50.8 ± 3.87 | 52.8 ± 3.54 a |
| 2-week | 55.0 ± 2.19 ab | 55.5 ± 2.59 | 53.2 ± 3.54 | 57.8 ± 3.60 b | |
| 3-week | 55.8 ± 1.60 b | 56.0 ± 3.52 | 54.4 ± 4.22 | 58.0 ± 2.97 b | |
| Albumin (g/L) | 1-week | 37.3 ± 2.50 | 35.3 ± 1.86 | 35.3 ± 3.93 | 35.8 ± 4.31 |
| 2-week | 39.7 ± 3.20 | 37.5 ± 2.26 | 38.5 ± 3.73 | 38.7 ± 3.44 | |
| 3-week | 41.0 ± 3.46 | 37.8 ± 2.71 | 38.8 ± 3.27 | 39.2 ± 3.54 | |
| Urea (mmol/L) | 1-week | 2.87 ± 0.27 AbB | 3.34 ± 0.21B | 2.83 ± 0.42 A | 2.50 ± 0.18 A |
| 2-week | 3.25 ± 0.63 b | 3.05 ± 0.40 | 3.02 ± 0.50 | 2.62 ± 0.33 | |
| 3-week | 2.05 ± 0.32 aA | 2.88 ± 0.23 B | 2.78 ± 0.20 B | 2.28 ± 0.35 A | |
| Creatinine (μmol/L) | 1-week | 75.6 ± 8.65 aA | 93.5 ± 10.4 B | 80.3 ± 11.1 AB | 77.7 ± 8.16 aAB |
| 2-week | 87.7 ± 9.27 Aab | 107 ± 10.8 B | 94.7 ± 10.5 AB | 92.2 ± 5.45 AbB | |
| 3-week | 88.0 ± 7.10 Ab | 112 ± 17.0 B | 94.0 ± 12.0 AB | 102 ± 10.9 AbB |
| Parameter | Period | Control | FB | DZ | FDZ |
|---|---|---|---|---|---|
| M ± SD | M ± SD | M ± SD | M ± SD | ||
| Glucose | 1-week | 6.02 ± 0.63 b | 6.84 ± 0.61 b | 6.03 ± 0.40 b | 6.52 ± 1.03 b |
| 2-week | 6.05 ± 0.57 b | 7.15 ± 0.74 b | 6.88 ± 0.81 b | 6.65 ± 0.56 b | |
| 3-week | 4.12 ± 0.46 a | 4.22 ± 0.93 a | 4.08 ± 0.60 a | 4.18 ± 0.68 a | |
| TGs | 1-week | 0.43 ± 0.06 a | 0.53 ± 0.10 | 0.47 ± 0.09 | 0.44 ± 0.11 |
| 2-week | 0.55 ± 0.09 b | 0.58 ± 0.11 | 0.49 ± 0.10 | 0.56 ± 0.21 | |
| 3-week | 0.37 ± 0.07 a | 0.47 ± 0.07 | 0.43 ± 0.12 | 0.47 ± 0.10 | |
| tCHO | 1-week | 2.34 ± 0.19 | 2.44 ± 0.25 | 2.16 ± 0.23 | 2.31 ± 0.28 |
| 2-week | 2.37 ± 0.16 AB | 2.65 ± 0.24 B | 2.25 ± 0.16 A | 2.51 ± 0.28 AB | |
| 3-week | 2.38 ± 0.16 | 2.48 ± 0.24 | 2.31 ± 0.09 | 2.53 ± 0.25 | |
| HDL | 1-week | 1.09 ± 0.12 | 1.08 ± 0.16 | 1.03 ± 0.14 | 1.07 ± 0.23 |
| 2-week | 1.12 ± 0.12 | 1.20 ± 0.11 | 1.14 ± 0.12 | 1.20 ± 0.19 | |
| 3-week | 1.14 ± 0.16 | 1.09 ± 0.15 | 1.16 ± 0.11 | 1.21 ± 0.18 | |
| LDL | 1-week | 1.06 ± 0.14 | 1.12 ± 0.10 | 0.91 ± 0.12 | 1.04 ± 0.20 |
| 2-week | 0.99 ± 0.09 AB | 1.18 ± 0.17 B | 0.89 ± 0.12 A | 1.05 ± 0.14 AB | |
| 3-week | 1.09 ± 0.06 | 1.18 ± 0.19 | 0.95 ± 0.11 | 1.10 ± 0.26 |
| Parameter | Period | Control | FB | DZ | FDZ |
|---|---|---|---|---|---|
| M ± SD | M ± SD | M ± SD | M ± SD | ||
| LDH | 1-week | 1093 ± 209 | 1009 ± 132 a | 1101 ± 281 | 1055 ± 208 |
| 2-week | 1093 ± 186 | 1163 ± 157 ab | 972 ± 78.0 | 1007 ± 137 | |
| 3-week | 1059 ± 174 | 1237 ± 147 b | 1263 ± 363 | 1097 ± 118 | |
| AST | 1-week | 96.7 ± 51.1 | 53.8 ± 15.5 | 66.2 ± 42.6 | 62.5 ± 21.8 |
| 2-week | 66.5 ± 29.9 | 68.0 ± 27.3 | 40.8 ± 7.55 | 68.2 ± 38.8 | |
| 3-week | 60.7 ± 25.7 | 72.8 ± 19.7 | 79.0 ± 38.4 | 56.8 ± 10.9 | |
| ALT | 1-week | 70.8 ± 11.0 | 78.7 ± 18.8 | 67.8 ± 17.4 | 71.5 ± 16.0 |
| 2-week | 72.8 ± 13.5 | 89.8 ± 22.9 | 73.0 ± 18.8 | 78.0 ± 20.0 | |
| 3-week | 70.8 ± 16.4 | 81.3 ± 14.9 | 74.0 ± 14.9 | 70.2 ± 15.1 | |
| AST/ALT | 1-week | 1.35 ± 0.63 | 0.70 ± 0.19 | 1.00 ± 0.59 | 0.87 ± 0.23 |
| 2-week | 1.00 ± 0.72 | 0.74 ± 0.13 | 0.57 ± 0.07 | 0.85 ± 0.29 | |
| 3-week | 0.89 ± 0.44 | 0.90 ± 0.18 | 0.99 ± 0.55 | 0.83 ± 0.16 | |
| GGT | 1-week | 32.7 ± 9.93 | 48.0 ± 11.5 | 36.0 ± 15.9 | 45.8 ± 8.98 |
| 2-week | 32.7 ± 12.6 A | 52.8 ± 12.9 AB | 36.0 ± 12.9 A | 57.7 ± 25.1 B | |
| 3-week | 38.3 ± 14.4 A | 70.0 ± 16.6 B | 54.6 ± 20.4 AB | 64.5 ± 14.9 AB | |
| ALP | 1-week | 359 ± 53.9 | 412 ± 84.5 | 339 ± 70.3 | 401 ± 134 |
| 2-week | 327 ± 59.1 | 378 ± 87.9 | 284 ± 44.7 | 386 ± 122 | |
| 3-week | 284 ± 47.6 AB | 334 ± 69.6 AB | 264 ± 64.2 A | 373 ± 102 B | |
| CK | 1-week | 1318 ± 668 | 718 ± 177 | 1064 ± 294 | 1551 ± 599 |
| 2-week | 1093 ± 462 | 976 ± 485 | 987 ± 409 | 1038 ± 492 | |
| 3-week | 1225 ± 412 | 1044 ± 389 | 1479 ± 536 | 1097 ± 533 |
| Parameter | Period | Control | FB | DZ | FDZ |
|---|---|---|---|---|---|
| M ± SD | M ± SD | M ± SD | M ± SD | ||
| Na | 1-week | 147 ± 2.51 | 150 ± 2.42 | 148 ± 1.60 | 148 ± 1.87 |
| 2-week | 149 ± 2.34 | 149 ± 2.50 | 149 ± 1.97 | 149 ± 2.37 | |
| 3-week | 148 ± 1.94 | 149 ± 1.05 | 148 ± 1.67 | 149 ± 1.86 | |
| Ca | 1-week | 3.23 ± 0.11 b | 3.41 ± 0.14 b | 3.20 ± 0.21 b | 3.22 ± 0.27 |
| 2-week | 3.18 ± 0.17 b | 3.11 ± 0.27 b | 3.22 ± 0.12 b | 3.13 ± 0.28 | |
| 3-week | 2.54 ± 0.23 aA | 2.65 ± 0.18 aAB | 2.49 ± 0.33 aA | 2.85 ± 0.27 B | |
| Corr. Ca | 1-week | 2.05 ± 0.08 b | 2.23 ± 0.11 c | 2.16 ± 0.39 b | 2.06 ± 0.19 b |
| 2-week | 1.96 ± 0.17 b | 1.94 ± 0.23 b | 2.02 ± 0.12 b | 1.94 ± 0.19 ab | |
| 3-week | 1.40 ± 0.22 aA | 1.56 ± 0.19 aAB | 1.40 ± 0.27 aA | 1.69 ± 0.23 aB | |
| P | 1-week | 2.85 ± 0.17 B | 2.63 ± 0.11 aAB | 2.51 ± 0.12 aA | 2.56 ± 0.25 aA |
| 2-week | 2.67 ± 0.20 b | 2.66 ± 0.13 a | 2.45 ± 0.08 a | 2.75 ± 0.46 ab | |
| 3-week | 2.62 ± 0.12 A | 3.10 ± 0.27 AbB | 3.15 ± 0.43 AbB | 3.07 ± 0.22 bB |
| Plasma | |||||
|---|---|---|---|---|---|
| Parameter | Period | Control | FB | DZ | FDZ |
| M ± SD | M ± SD | M ± SD | M ± SD | ||
| GSH (micromol/g prot.) | 1-week | 2.88 ± 0.24 | 2.94 ± 0.30 | 2.86 ± 0.20 a | 2.88± 0.26 |
| 2-week | 2.68 ± 0.77 | 2.62 ± 0.49 | 2.31 ± 0.66 ab | 2.87 ± 0.38 | |
| 3-week | 3.15 ± 0.18 | 3.10 ± 0.26 | 3.34 ± 0.13 b | 2.98 ± 0.33 | |
| GPx (U/g prot.) | 1-week | 3.29 ± 0.35 | 3.25 ± 0.32 a | 3.36 ± 0.52 a | 3.47 ± 0.27 |
| 2-week | 3.49 ± 0.29 | 3.28 ± 0.17 a | 3.16 ± 0.17 a | 3.22 ± 0.26 | |
| 3-week | 4.07 ± 0.96 | 3.81 ± 0.34 b | 3.93 ± 0.15 b | 4.07 ± 0.92 | |
| MDA (nmol/g) | 1-week | 7.21 ± 1.10 | 8.13 ± 0.48 | 7.56 ± 1.22 | 7.76 ± 0.95 |
| 2-week | 8.56 ± 1.15 | 8.86 ± 1.09 | 7.65 ± 0.97 | 8.72 ± 1.43 | |
| 3-week | 8.31 ± 0.33 | 9.20 ± 0.80 | 8.22 ± 0.76 | 8.75 ± 0.52 | |
| Kidney | |||||
| GSH (micromol/g prot.) | 3-week | 3.91 ± 0.46 | 3.05 ± 0.57 | 3.84 ± 0.63 | 3.14 ± 0.96 |
| GPx (U/g prot.) | 3-week | 4.28 ± 0.65 B | 3.35 ± 0.51 A | 4.09 ± 0.67 AB | 3.19 ± 0.34 A |
| MDA(nmol/g) | 3-week | 46.9 ± 6.43 B | 35.2 ± 4.54 A | 43.8 ± 8.51 AB | 42.5 ± 6.04 AB |
| Liver | |||||
| GSH (micromol/g prot.) | 3-week | 5.30 ± 0.88 | 4.59 ± 0.50 | 4.13 ± 0.21 | 4.59 ± 1.04 |
| GPx (U/g prot.) | 3-week | 4.63 ± 0.70 | 4.33 ± 0.62 | 4.10 ± 0.34 | 4.27 ± 1.05 |
| MDA(nmol/g) | 3-week | 47.0 ± 7.87 | 47.2 ± 8.14 | 43.1 ± 14.3 | 50.0 ±13.3 |
| Lung | |||||
| GSH (micromol/g prot.) | 3-week | 2.92 ± 0.41 | 2.66 ± 0.66 | 2.57 ± 0.57 | 2.66± 0.40 |
| GPx (U/g prot.) | 3-week | 3.38 ± 0.50 | 3.06 ± 0.29 | 2.97 ± 0.49 | 3.04 ± 0.17 |
| MDA(nmol/g) | 3-week | 28.6 ± 5.18 | 32.2 ± 5.71 | 31.5 ± 4.14 | 32.5 ± 4.38 |
| Composition | Calculated from 100% Dry Matter/kg |
|---|---|
| DE (MJ/kg) | 16.3 |
| ME (MJ/kg) | 15.6 |
| CP (%) | 19.8 |
| Lysine (%) | 1.5 |
| Methionine (%) | 0.6 |
| EE (%) | 5.2 |
| CF (%) | 3.3 |
| Calcium (%) | 0.8 |
| Phosphorus (%) | 0.6 |
| Sodium (%) | 0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, O.; Mézes, M.; Balogh, K.; Kovács, M.; Szabó, A. The Effects of Mixed Fusarium Mycotoxins at EU-Permitted Feed Levels on Weaned Piglets’ Tissue Lipids. Toxins 2021, 13, 444. https://doi.org/10.3390/toxins13070444
Ali O, Mézes M, Balogh K, Kovács M, Szabó A. The Effects of Mixed Fusarium Mycotoxins at EU-Permitted Feed Levels on Weaned Piglets’ Tissue Lipids. Toxins. 2021; 13(7):444. https://doi.org/10.3390/toxins13070444
Chicago/Turabian StyleAli, Omeralfaroug, Miklós Mézes, Krisztián Balogh, Melinda Kovács, and András Szabó. 2021. "The Effects of Mixed Fusarium Mycotoxins at EU-Permitted Feed Levels on Weaned Piglets’ Tissue Lipids" Toxins 13, no. 7: 444. https://doi.org/10.3390/toxins13070444
APA StyleAli, O., Mézes, M., Balogh, K., Kovács, M., & Szabó, A. (2021). The Effects of Mixed Fusarium Mycotoxins at EU-Permitted Feed Levels on Weaned Piglets’ Tissue Lipids. Toxins, 13(7), 444. https://doi.org/10.3390/toxins13070444