Evaluation of Signaling Pathways Profiling in Human Dermal Endothelial Cells Treated by Snake Venom Cysteine-Rich Secretory Proteins (svCRiSPs) from North American Snakes Using Reverse Phase Protein Array (RPPA)

 ,
,  ,
,
Abstract
:1. Introduction
2. Results and Discussion
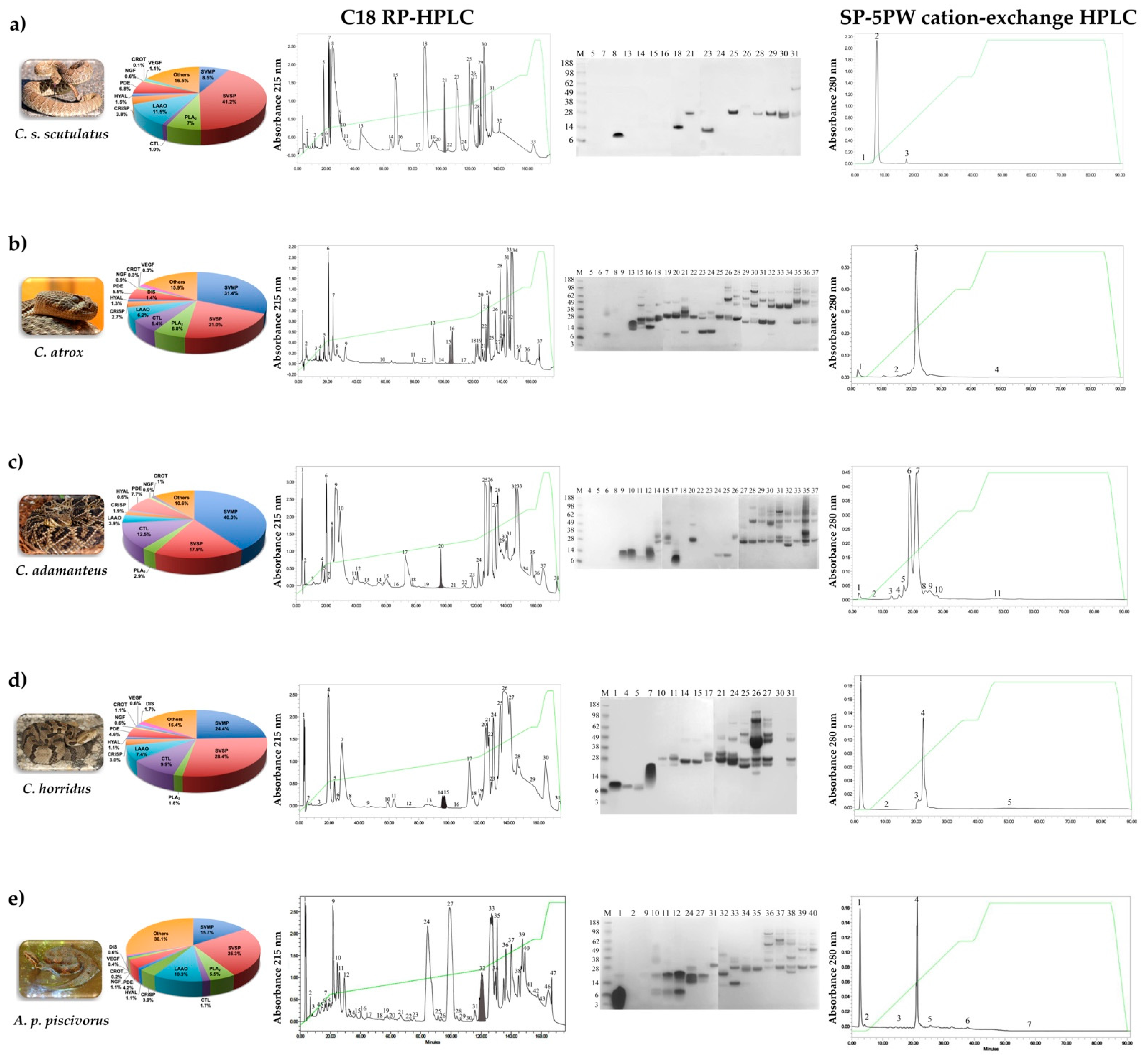
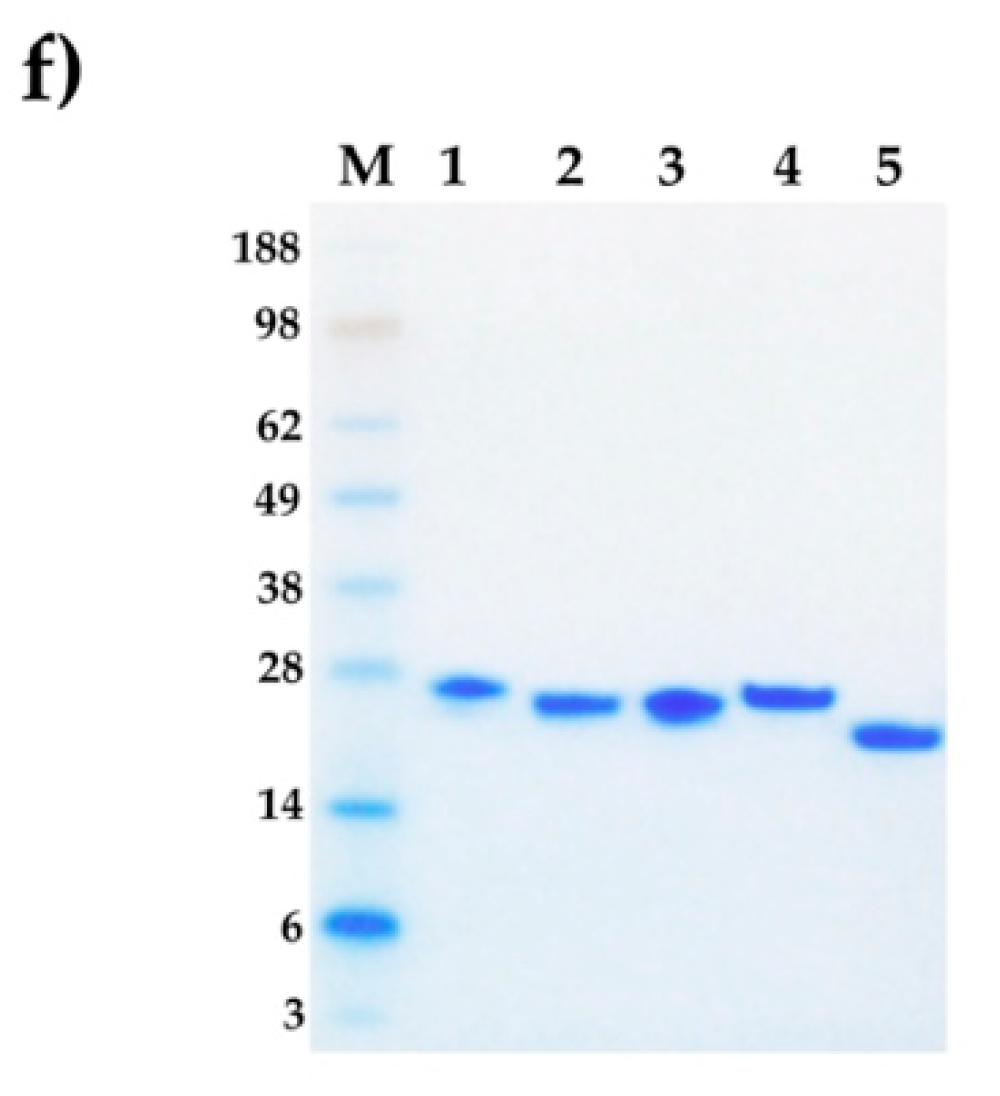
2.1. Venomic Analysis and Purification of svCRiSPs
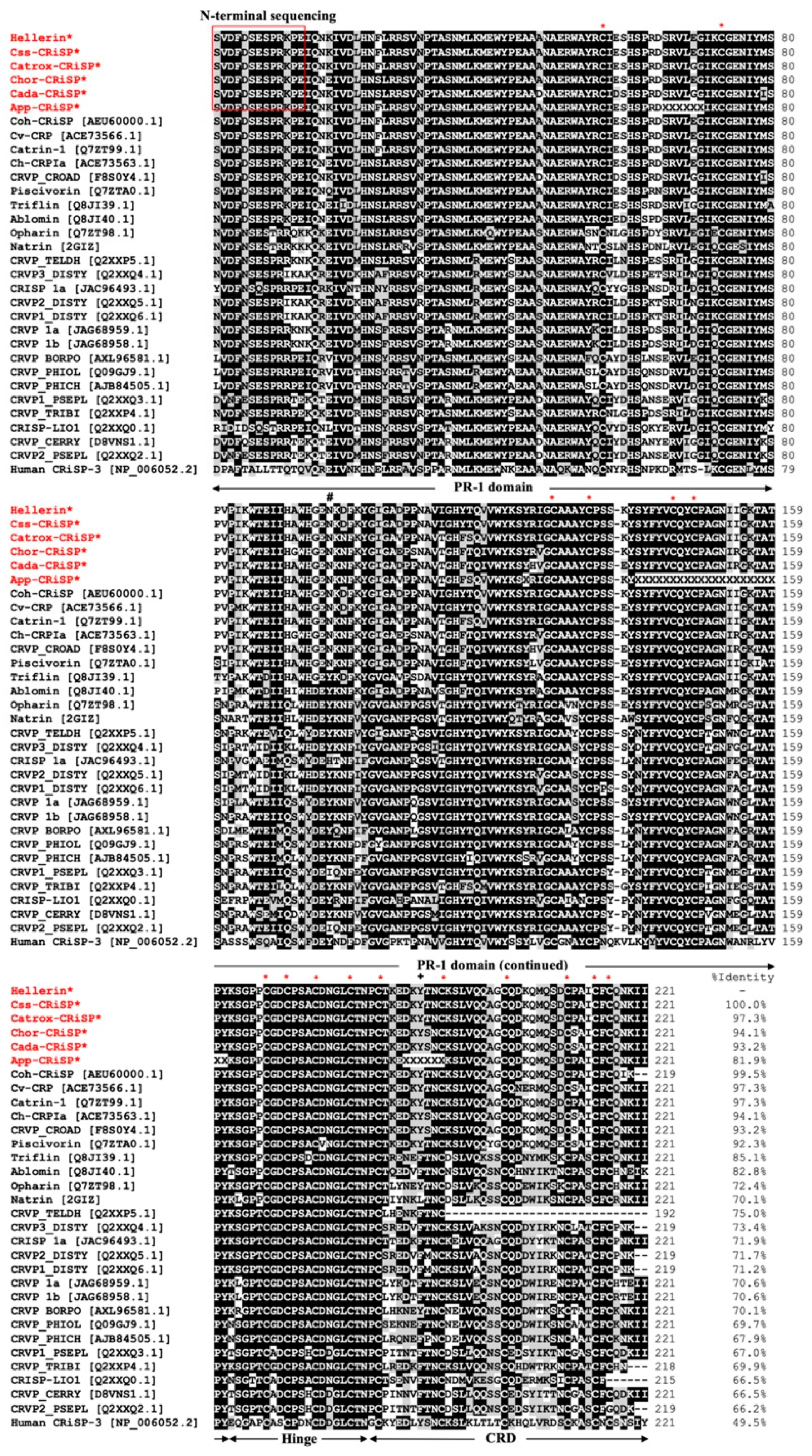
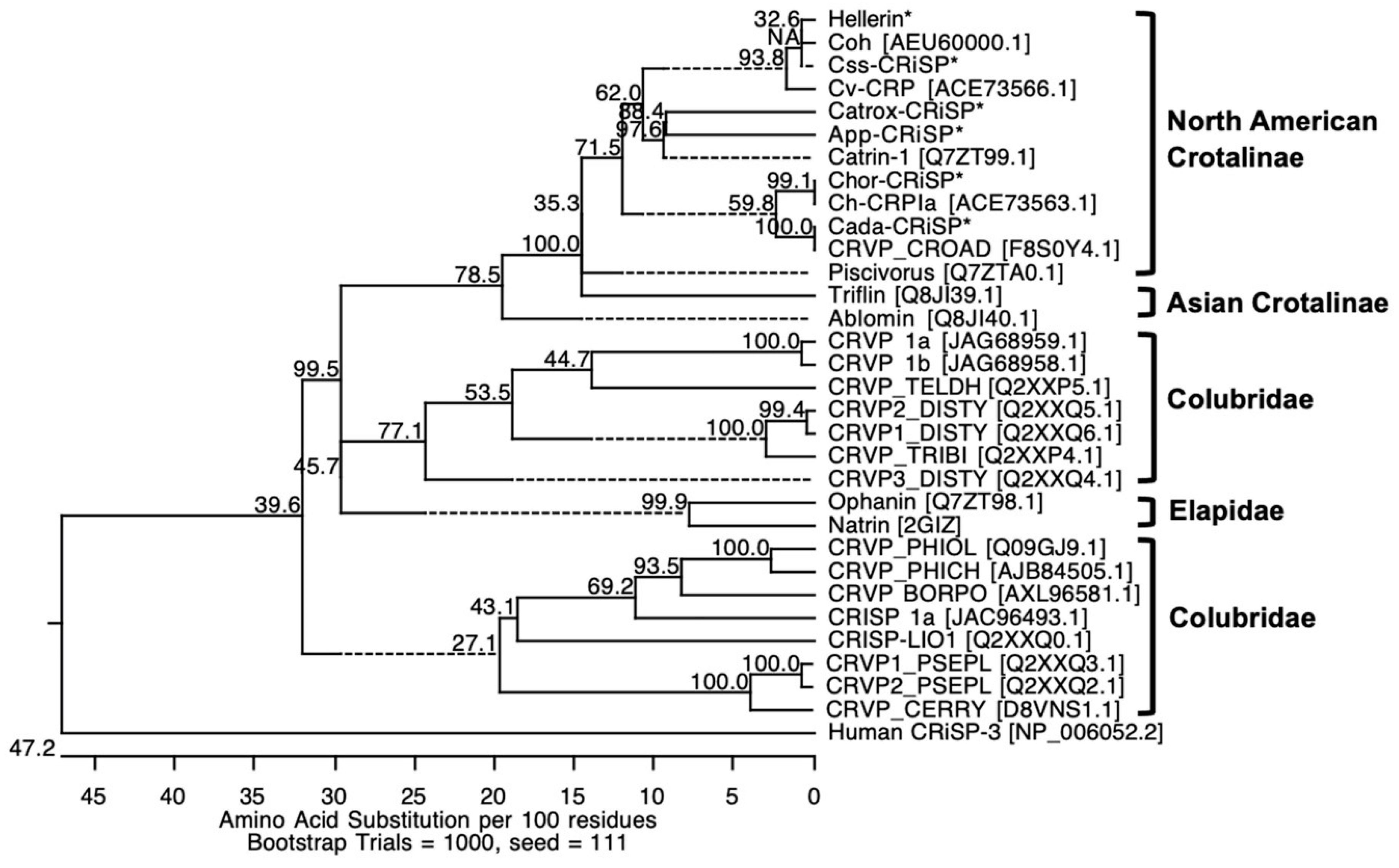
2.2. Full Amino Acid Sequences of svCRiSPs
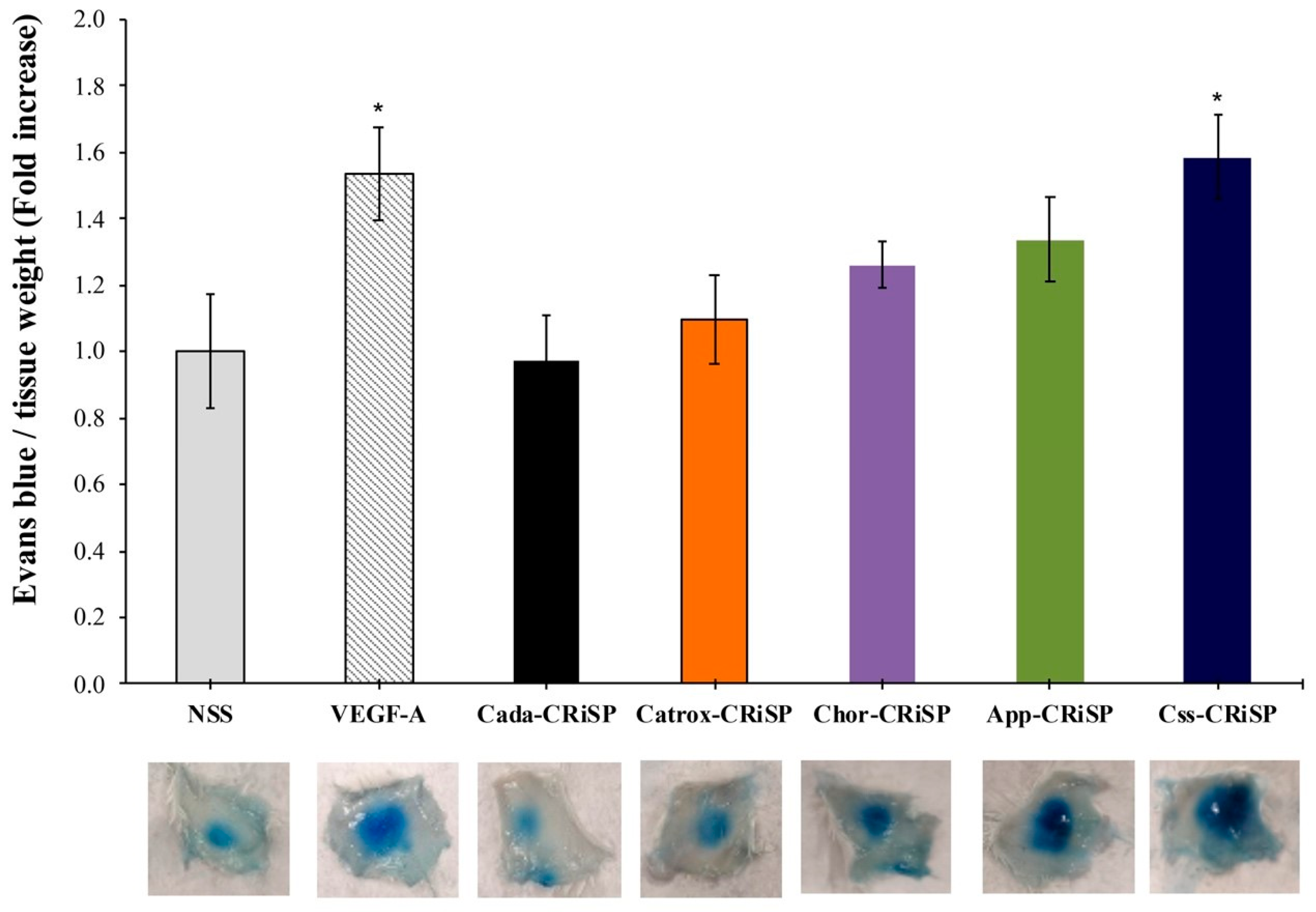
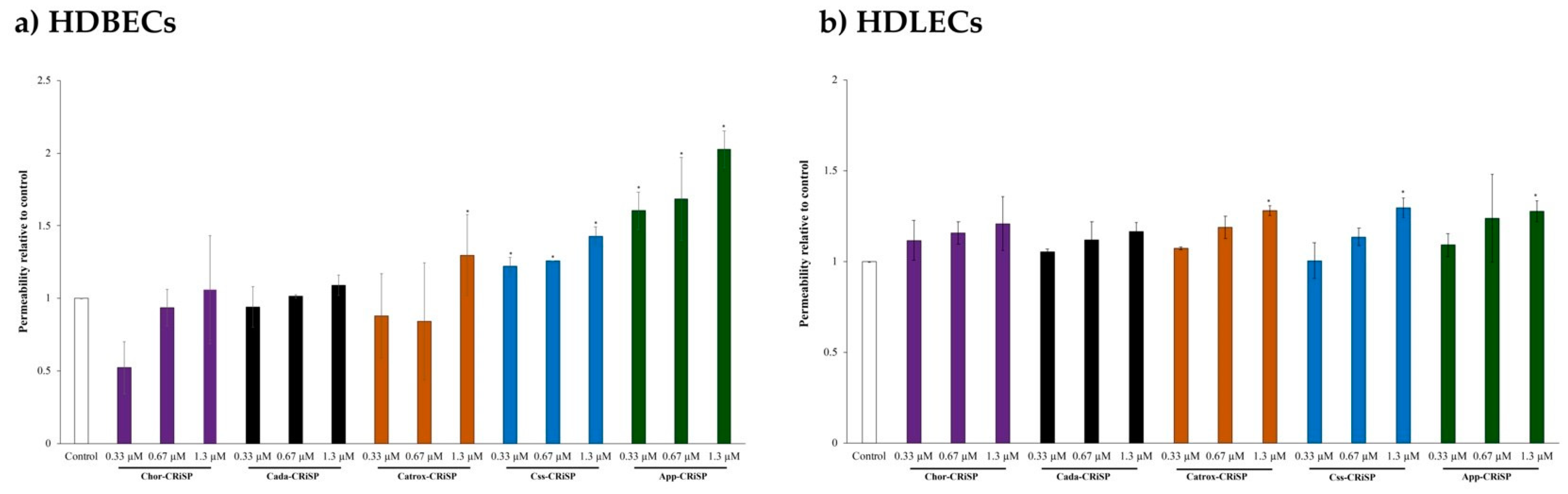
2.3. Measurement of Vascular Permeability and Endothelial Permeability
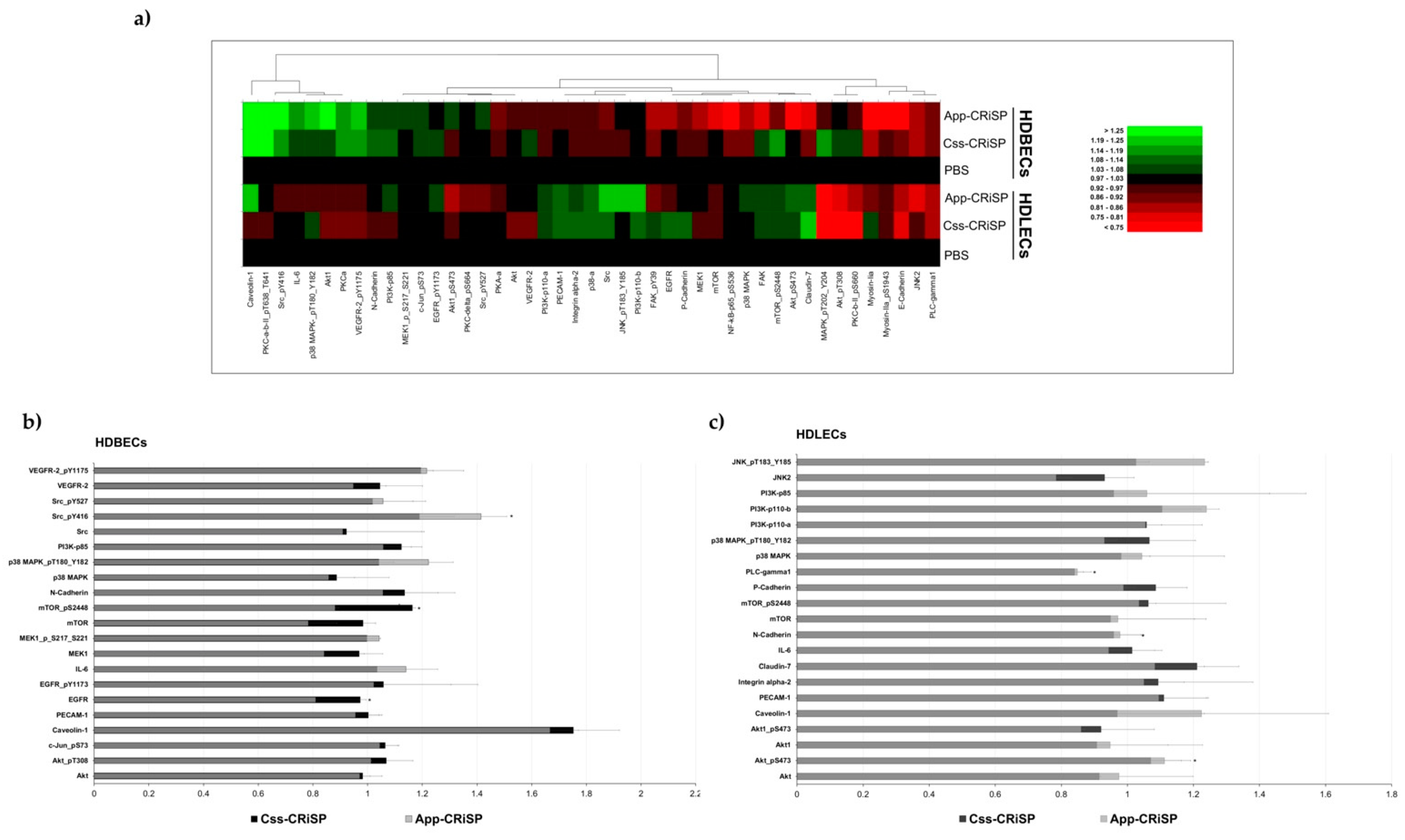
2.4. Reverse Phase Protein Array (RPPA)
3. Conclusions
4. Materials and Methods
4.1. Snake Venoms
4.2. Isolation of svCRiSPs
4.3. SDS-PAGE and N-Terminal Sequencing
4.4. Protein Identification of svCRiSPs by LC-MS/MS Analysis and Data Processing
4.5. Multiple Sequence Alignment
4.6. Vascular Permeability In Vivo (Miles Assay)
4.7. Cell Cultures
4.8. Endothelial Permeability Assay
4.9. Reverse Phase Protein Array (RPPA)
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gutierrez, J.M.; Calvete, J.J.; Habib, A.G.; Harrison, R.A.; Williams, D.J.; Warrell, D.A. Snakebite envenoming. Nat. Rev. Dis. Primers. 2017, 3, 17063. [Google Scholar] [CrossRef]
- Gibbs, G.M.; O’Bryan, M.K. Cysteine rich secretory proteins in reproduction and venom. Soc. Reprod. Fertil. Suppl. 2007, 65, 261–267. [Google Scholar]
- Sunagar, K.; Johnson, W.E.; O’Brien, S.J.; Vasconcelos, V.; Antunes, A. Evolution of CRISPs associated with toxicoferan-reptilian venom and mammalian reproduction. Mol. Biol. Evol. 2012, 29, 1807–1822. [Google Scholar] [CrossRef] [Green Version]
- Tadokoro, T.; Modahl, C.M.; Maenaka, K.; Aoki-Shioi, N. Cysteine-Rich Secretory Proteins (CRISPs) From Venomous Snakes: An Overview of the Functional Diversity in a Large and Underappreciated Superfamily. Toxins 2020, 12, 175. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, N.; Yamazaki, Y.; Brown, R.L.; Fujimoto, Z.; Morita, T.; Mizuno, H. Structures of pseudechetoxin and pseudecin, two snake-venom cysteine-rich secretory proteins that target cyclic nucleotide-gated ion channels: Implications for movement of the C-terminal cysteine-rich domain. Acta Crystallogr. D Biol. Crystallogr. 2008, 64, 1034–1042. [Google Scholar] [CrossRef]
- Wang, F.; Li, H.; Liu, M.N.; Song, H.; Han, H.M.; Wang, Q.L.; Yin, C.C.; Zhou, Y.C.; Qi, Z.; Shu, Y.Y.; et al. Structural and functional analysis of natrin, a venom protein that targets various ion channels. Biochem. Biophys. Res. Commun. 2006, 351, 443–448. [Google Scholar] [CrossRef]
- Wang, Y.L.; Kuo, J.H.; Lee, S.C.; Liu, J.S.; Hsieh, Y.C.; Shih, Y.T.; Chen, C.J.; Chiu, J.J.; Wu, W.G. Cobra CRISP functions as an inflammatory modulator via a novel Zn2+- and heparan sulfate-dependent transcriptional regulation of endothelial cell adhesion molecules. J. Biol. Chem. 2010, 285, 37872–37883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lecht, S.; Chiaverelli, R.A.; Gerstenhaber, J.; Calvete, J.J.; Lazarovici, P.; Casewell, N.R.; Harrison, R.; Lelkes, P.I.; Marcinkiewicz, C. Anti-angiogenic activities of snake venom CRISP isolated from Echis carinatus sochureki. Biochim. Biophys. Acta 2015, 1850, 1169–1179. [Google Scholar] [CrossRef]
- Adade, C.M.; Carvalho, A.L.; Tomaz, M.A.; Costa, T.F.; Godinho, J.L.; Melo, P.A.; Lima, A.P.; Rodrigues, J.C.; Zingali, R.B.; Souto-Padron, T. Crovirin, a snake venom cysteine-rich secretory protein (CRISP) with promising activity against Trypanosomes and Leishmania. PLoS Negl. Trop. Dis. 2014, 8, e3252. [Google Scholar] [CrossRef]
- Suntravat, M.; Cromer, W.E.; Marquez, J.; Galan, J.A.; Zawieja, D.C.; Davies, P.; Salazar, E.; Sánchez, E.E. The isolation and characterization of a new snake venom cysteine-rich secretory protein (svCRiSP) from the venom of the Southern Pacific rattlesnake and its effect on vascular permeability. Toxicon 2019, 165, 22–30. [Google Scholar] [CrossRef]
- Lomonte, B.; Tsai, W.C.; Ureña-Diaz, J.M.; Sanz, L.; Mora-Obando, D.; Sánchez, E.E.; Fry, B.G.; Gutiérrez, J.M.; Gibbs, H.L.; Sovic, M.G.; et al. Venomics of New World pit vipers: Genus-wide comparisons of venom proteomes across Agkistrodon. J. Proteom. 2014, 96, 103–116. [Google Scholar] [CrossRef] [Green Version]
- Calvete, J.J.; Fasoli, E.; Sanz, L.; Boschetti, E.; Righetti, P.G. Exploring the venom proteome of the western diamondback rattlesnake, Crotalus atrox, via snake venomics and combinatorial peptide ligand library approaches. J. Proteome Res. 2009, 8, 3055–3067. [Google Scholar] [CrossRef] [Green Version]
- Rokyta, D.R.; Lemmon, A.R.; Margres, M.J.; Aronow, K. The venom-gland transcriptome of the eastern diamondback rattlesnake (Crotalus adamanteus). BMC Genom. 2012, 13, 312. [Google Scholar] [CrossRef] [Green Version]
- Rokyta, D.R.; Wray, K.P.; McGivern, J.J.; Margres, M.J. The transcriptomic and proteomic basis for the evolution of a novel venom phenotype within the Timber Rattlesnake (Crotalus horridus). Toxicon 2015, 98, 34–48. [Google Scholar] [CrossRef]
- Matsunaga, Y.; Yamazaki, Y.; Hyodo, F.; Sugiyama, Y.; Nozaki, M.; Morita, T. Structural divergence of cysteine-rich secretory proteins in snake venoms. J. Biochem. 2009, 145, 365–375. [Google Scholar] [CrossRef]
- Creighton, C.J.; Huang, S. Reverse phase protein arrays in signaling pathways: A data integration perspective. Drug Des. Dev. Ther. 2015, 9, 3519–3527. [Google Scholar]
- Mehta, D.; Malik, A.B. Signaling mechanisms regulating endothelial permeability. Physiol. Rev. 2006, 86, 279–367. [Google Scholar] [CrossRef]
- Komarova, Y.; Kruse, K.J.; Mehta, D.; Malik, A.B. Response by Komarova et al. to Letter Regarding Article, “Protein Interactions at Endothelial Junctions and Signaling Mechanisms Regulating Endothelial Permeability”. Circ. Res. 2017, 120, e28. [Google Scholar] [CrossRef] [Green Version]
- Balda, M.S.; Matter, K. Tight junctions as regulators of tissue remodelling. Curr. Opin. Cell Biol. 2016, 42, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, N. Defenders and Challengers of Endothelial Barrier Function. Front. Immunol. 2017, 8, 1847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radeva, M.Y.; Waschke, J. Mind the gap: Mechanisms regulating the endothelial barrier. Acta Physiol. 2018, 222, e12860. [Google Scholar] [CrossRef]
- Predescu, S.A.; Predescu, D.N.; Palade, G.E. Plasmalemmal vesicles function as transcytotic carriers for small proteins in the continuous endothelium. Am. J. Physiol. 1997, 272, H937–H949. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.G.; Kamen, B.A.; Rothberg, K.G.; Lacey, S.W. Potocytosis: Sequestration and transport of small molecules by caveolae. Science 1992, 255, 410–411. [Google Scholar] [CrossRef] [PubMed]
- Frank, P.G.; Woodman, S.E.; Park, D.S.; Lisanti, M.P. Caveolin, caveolae, and endothelial cell function. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 1161–1168. [Google Scholar] [CrossRef]
- Navarro, A.; Anand-Apte, B.; Parat, M.O. A role for caveolae in cell migration. FASEB J. 2004, 18, 1801–1811. [Google Scholar] [CrossRef]
- Wang, N.; Zhang, D.; Sun, G.; Zhang, H.; You, Q.; Shao, M.; Yue, Y. Lipopolysaccharide-induced caveolin-1 phosphorylation-dependent increase in transcellular permeability precedes the increase in paracellular permeability. Drug Des. Dev. Ther. 2015, 9, 4965–4977. [Google Scholar]
- Jiang, R.; Cai, J.; Zhu, Z.; Chen, D.; Wang, J.; Wang, Q.; Teng, Y.; Huang, Y.; Tao, M.; Xia, A.; et al. Hypoxic trophoblast HMGB1 induces endothelial cell hyperpermeability via the TRL-4/caveolin-1 pathway. J. Immunol. (Baltim. Md. 1950) 2014, 193, 5000–5012. [Google Scholar] [CrossRef]
- Cavallaro, U.; Liebner, S.; Dejana, E. Endothelial cadherins and tumor angiogenesis. Exp. Cell Res. 2006, 312, 659–667. [Google Scholar] [CrossRef]
- Turley, R.S.; Tokuhisa, Y.; Toshimitsu, H.; Lidsky, M.E.; Padussis, J.C.; Fontanella, A.; Deng, W.; Augustine, C.K.; Beasley, G.M.; Davies, M.A.; et al. Targeting N-cadherin increases vascular permeability and differentially activates AKT in melanoma. Ann. Surg. 2015, 261, 368–377. [Google Scholar] [CrossRef] [Green Version]
- Kruse, K.; Lee, Q.S.; Sun, Y.; Klomp, J.; Yang, X.; Huang, F.; Sun, M.Y.; Zhao, S.; Hong, Z.; Vogel, S.M.; et al. N-cadherin signaling via Trio assembles adherens junctions to restrict endothelial permeability. J. Cell Biol. 2019, 218, 299–316. [Google Scholar] [CrossRef] [PubMed]
- Fasano, A.; Fiorentini, C.; Donelli, G.; Uzzau, S.; Kaper, J.B.; Margaretten, K.; Ding, X.; Guandalini, S.; Comstock, L.; Goldblum, S.E. Zonula occludens toxin modulates tight junctions through protein kinase C-dependent actin reorganization, in vitro. J. Clin. Investig. 1995, 96, 710–720. [Google Scholar] [CrossRef]
- Seth, A.; Yan, F.; Polk, D.B.; Rao, R.K. Probiotics ameliorate the hydrogen peroxide-induced epithelial barrier disruption by a PKC- and MAP kinase-dependent mechanism. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G1060–G1069. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Yoshinaga, N.; Tanabe, S. Interleukin-6 (IL-6) regulates Claudin-2 expression and tight junction permeability in intestinal epithelium. J. Biol. Chem. 2011, 286, 31263–31271. [Google Scholar] [CrossRef] [Green Version]
- Marchiando, A.M.; Shen, L.; Graham, W.V.; Weber, C.R.; Schwarz, B.T.; Austin, J.R., 2nd; Raleigh, D.R.; Guan, Y.; Watson, A.J.; Montrose, M.H.; et al. Caveolin-1-dependent occludin endocytosis is required for TNF-induced tight junction regulation in vivo. J. Cell Biol. 2010, 189, 111–126. [Google Scholar] [CrossRef] [Green Version]
- Al-Sadi, R.; Guo, S.; Ye, D.; Dokladny, K.; Alhmoud, T.; Ereifej, L.; Said, H.M.; Ma, T.Y. Mechanism of IL-1β modulation of intestinal epithelial barrier involves p38 kinase and activating transcription factor-2 activation. J. Immunol. 2013, 190, 6596–6606. [Google Scholar] [CrossRef] [Green Version]
- Catalioto, R.M.; Maggi, C.A.; Giuliani, S. Intestinal epithelial barrier dysfunction in disease and possible therapeutical interventions. Curr. Med. Chem. 2011, 18, 398–426. [Google Scholar] [CrossRef] [PubMed]
- Weis, S.M.; Cheresh, D.A. Pathophysiological consequences of VEGF-induced vascular permeability. Nature 2005, 437, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Olsson, A.K.; Dimberg, A.; Kreuger, J.; Claesson-Welsh, L. VEGF receptor signalling—In control of vascular function. Nat. Rev. Mol. Cell Biol. 2006, 7, 359–371. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.M.; Yuan, Y.; Zawieja, D.C.; Tinsley, J.; Granger, H.J. Role of phospholipase C, protein kinase C, and calcium in VEGF-induced venular hyperpermeability. Am. J. Physiol. 1999, 276, H535–H542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, P.D.; Klein, R.R.; Troutman, M.D.; Desai, S.; Thakker, D.R. Phospholipase C-gamma modulates epithelial tight junction permeability through hyperphosphorylation of tight junction proteins. J. Biol. Chem. 2002, 277, 35760–35765. [Google Scholar] [CrossRef] [Green Version]
- Eliceiri, B.P.; Paul, R.; Schwartzberg, P.L.; Hood, J.D.; Leng, J.; Cheresh, D.A. Selective requirement for Src kinases during VEGF-induced angiogenesis and vascular permeability. Mol. Cell 1999, 4, 915–924. [Google Scholar] [CrossRef]
- Pedram, A.; Razandi, M.; Levin, E.R. Deciphering vascular endothelial cell growth factor/vascular permeability factor signaling to vascular permeability. Inhibition by atrial natriuretic peptide. J. Biol. Chem. 2002, 277, 44385–44398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trani, M.; Dejana, E. New insights in the control of vascular permeability: Vascular endothelial-cadherin and other players. Curr. Opin. Hematol. 2015, 22, 267–272. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CRiSPs | Positions | Theo. MH+ [Da] | Sequenced Fragment |
|---|---|---|---|
| Css-CRiSP | 30–36 | 856.48869 | (R)KPEIQNK(I) |
| 37–45 | 1126.63675 | (K)IVDLHNFLR(R) | |
| 46–57 | 1333.68926 | (R)RSVNPTASNMLK(M) | |
| 58–74 | 2129.94978 | (K)MEWYPEAAANAERWAYR(C) | |
| 75–85 | 1343.61207 | (R)CIESHSPRDSR(V) | |
| 86–91 | 658.4134 | (R)VLEGIK(C) | |
| 92–104 | 1523.72326 | (K)CGENIYMSPVPIK(W) | |
| 105–117 | 1620.79175 | (K)WTEIIHAWHGENK(D) | |
| 121–142 | 2448.23466 | (K)YGIGADPPNAVIGHYTQVVWYK(S) | |
| 146–157 | 1284.57112 | (R)IGCAAAYCPSSK(Y) | |
| 158–175 | 2202.99871 | (K)YSYFYVCQYCPAGNIIGK(T) | |
| 176–181 | 680.36137 | (K)TATPYK(S) | |
| 182–212 | 3563.40587 | (K)SGPPCGDCPSACDNGLCTNPCTKEDKYTNCK(S) | |
| 213–238 | 3117.33151 | (K)SLVQQAGCQDKQMQSDCPAICFCQNK(I) | |
| 224–240 | 2128.92826 | (K)QMQSDCPAICFCQNKII(-) | |
| Catrox-CRiSP | 30–45 | 1964.10761 | (R)KPEIQNKIVDLHNFL(R) |
| 46–57 | 1333.68926 | (R)RSVNPTASNMLK(M) | |
| 58–74 | 2129.94978 | (K)MEWYPEAAANAERWAYR(C) | |
| 75–82 | 985.45199 | (R)CIESHSPR(D) | |
| 86–104 | 2091.09769 | (R)VLGGIKCGENIYMSPVPIK(W) | |
| 105–120 | 2009.99805 | (K)WTEIIHAWHGENKNFK(Y) | |
| 121–145 | 2796.42564 | (K)YGIGAVPPNAVTGHFSQVVWYKSYR(I) | |
| 146–175 | 3468.55199 | (R)IGCAAAYCPSSKYSYFYVCQYCPAGNIIGK(T) | |
| 176–204 | 3186.30535 | (K)TATPYKSGPPCGDCPSACDNGLCTNPCTK(E) | |
| 205–223 | 2272.03326 | (K)EDKYTNCKSLVQQAGCQDK(Q) | |
| 224–240 | 2128.92826 | (K)QMQSDCPAICFCQNKII(-) | |
| Cada-CRiSP | 30–46 | 2060.17233 | (R)KPEIQNKIVDLHNSLRR(S) |
| 47–57 | 1177.58815 | (R)SVNPTASNMLK(M) | |
| 58–74 | 2173.93961 | (K)MEWYPEAADNAERWAYR(C) | |
| 75–85 | 1329.59642 | (R)CIDSHSPRDSR(V) | |
| 83–104 | 2415.30644 | (R)DSRVLGGIKCGENIYISPVPIK(W) | |
| 105–117 | 1620.79175 | (K)WTEIIHAWHGENK(N) | |
| 121–142 | 2434.21901 | (K)YGIGADPPNAVTGHFTQIVWYK(S) | |
| 143–173 | 3700.53886 | (K)SYHVGCAAAYCPSSEYSYFYVCQYCPAGNIR(G) | |
| 174–181 | 865.47779 | (R)GKTATPYK(S) | |
| 182–212 | 3549.39022 | (K)SGPPCGDCPSACDNGLCTNPCTKEDKYSNCK(S) | |
| 213–240 | 3333.4789 | (K)SLVQQAGCQDKQMQSDCSAICFCQNKII(-) | |
| Chor-CRiSP | 1–10 | 1180.51167 | SVDFDSESPR(K) |
| 11–27 | 2061.11996 | (R)KPEIQNEIVDLHNSLRR(S) | |
| 28–38 | 1177.58815 | (R)SVNPTASNMLK(M) | |
| 39–55 | 2129.94978 | (K)MEWYPEAAANAERWAYR(C) | |
| 56–66 | 1343.61207 | (R)CIESHSPRDSR(V) | |
| 67–85 | 2163.11882 | (R)VLEGIKCGENIYMSPVPIK(W) | |
| 86–98 | 1606.7761 | (K)WTEIIHGWHGENK(N) | |
| 102–123 | 2438.21392 | (K)YGIGAEPSNAVTGHFTQIVWYK(S) | |
| 124–138 | 1676.75194 | (K)SYRVGCAAAYCPSSK(Y) | |
| 139–156 | 2246.01576 | (K)YSYFYVCQYCPAGNIRGK(T) | |
| 157–185 | 3186.30535 | (K)TATPYKSGPPCGDCPSACDNGLCTNPCTK(E) | |
| 186–204 | 2258.01761 | (K)EDKYSNCKSLVQQAGCQDK(Q) | |
| 205–221 | 2118.90753 | (K)QMQSDCSAICFCQNKII(-) | |
| App-CRiSP | 30–36 | 856.48869 | (R)KPEIQNK(I) |
| 37–45 | 1126.63675 | (K)IVDLHNFL(R) | |
| 46–57 | 1333.68926 | (R)RSVNPTASNMLK(M) | |
| 58–70 | 1553.66892 | (K)MEWYPEAAANAER(W) | |
| 75–82 | 985.45199 | (R)CIESHSPR(D) | |
| 92–104 | 1523.72326 | (K)CGENIYMSPVPIK(W) | |
| 105–117 | 1620.79175 | (K)WTEIIHAWHGENK(N) | |
| 121–142 | 2390.22918 | (K)YGIGAVPPNAVTGHFSQVVWYK(S) | |
| 146–157 | 1284.57112 | (R)IGCAAAYCPSSK(Y) | |
| 182–204 | 2524.96182 | (K)SGPPCGDCPSACDNGLCTNPCTK(E) | |
| 213–223 | 1233.58921 | (K)SLVQQAGCQDK(Q) | |
| 224–238 | 1902.76014 | (K)QMQSDCPAICFCQNK(I) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suntravat, M.; Sanchez, O.; Reyes, A.; Cirilo, A.; Ocheltree, J.S.; Galan, J.A.; Salazar, E.; Davies, P.; Sanchez, E.E. Evaluation of Signaling Pathways Profiling in Human Dermal Endothelial Cells Treated by Snake Venom Cysteine-Rich Secretory Proteins (svCRiSPs) from North American Snakes Using Reverse Phase Protein Array (RPPA). Toxins 2021, 13, 613. https://doi.org/10.3390/toxins13090613
Suntravat M, Sanchez O, Reyes A, Cirilo A, Ocheltree JS, Galan JA, Salazar E, Davies P, Sanchez EE. Evaluation of Signaling Pathways Profiling in Human Dermal Endothelial Cells Treated by Snake Venom Cysteine-Rich Secretory Proteins (svCRiSPs) from North American Snakes Using Reverse Phase Protein Array (RPPA). Toxins. 2021; 13(9):613. https://doi.org/10.3390/toxins13090613
Chicago/Turabian StyleSuntravat, Montamas, Oscar Sanchez, Armando Reyes, Abcde Cirilo, Jack S. Ocheltree, Jacob A. Galan, Emelyn Salazar, Peter Davies, and Elda E. Sanchez. 2021. "Evaluation of Signaling Pathways Profiling in Human Dermal Endothelial Cells Treated by Snake Venom Cysteine-Rich Secretory Proteins (svCRiSPs) from North American Snakes Using Reverse Phase Protein Array (RPPA)" Toxins 13, no. 9: 613. https://doi.org/10.3390/toxins13090613
APA StyleSuntravat, M., Sanchez, O., Reyes, A., Cirilo, A., Ocheltree, J. S., Galan, J. A., Salazar, E., Davies, P., & Sanchez, E. E. (2021). Evaluation of Signaling Pathways Profiling in Human Dermal Endothelial Cells Treated by Snake Venom Cysteine-Rich Secretory Proteins (svCRiSPs) from North American Snakes Using Reverse Phase Protein Array (RPPA). Toxins, 13(9), 613. https://doi.org/10.3390/toxins13090613