Ageratina adenophora Disrupts the Intestinal Structure and Immune Barrier Integrity in Rats

 , ,
, ,
Abstract
:1. Introduction
2. Results
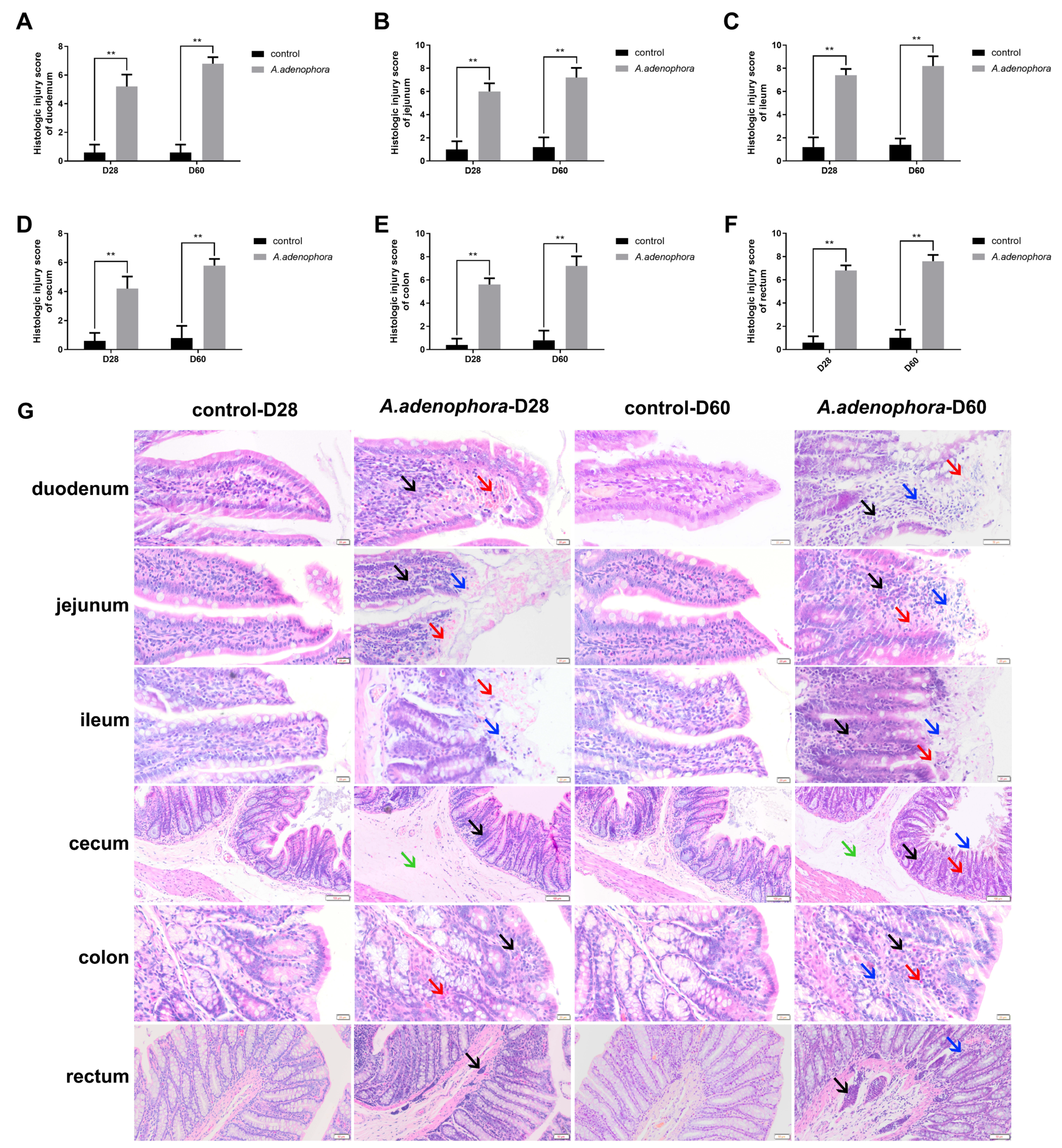
2.1. A. adenophora Administration Increased Intestinal Pathological Injury in Rat
2.1.1. Effects of A. adenophora on Histopathological Changes in the Small Intestinal Sections of Rats
2.1.2. Effects of A. adenophora on Histopathology of Large Intestinal Sections in Rats
2.1.3. Effect of A. adenophora on Villi Height, Crypt Depth, and the Villi Height Ratio/Crypt Depth (v/c) of the Small Intestine Sections in Rats
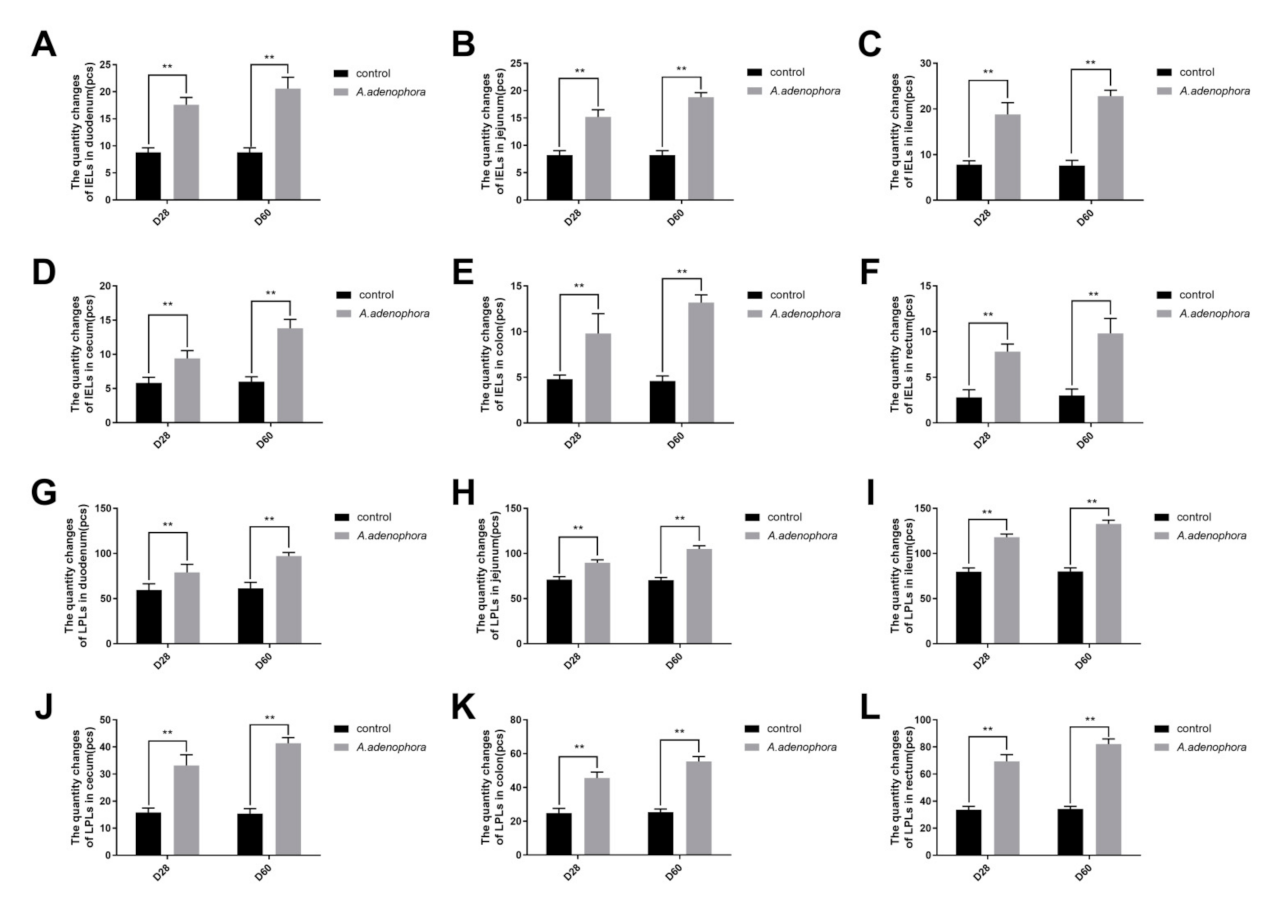
2.1.4. Effects of A. adenophora on the IELS and LPLS Count of Intestinal Sections in Rats
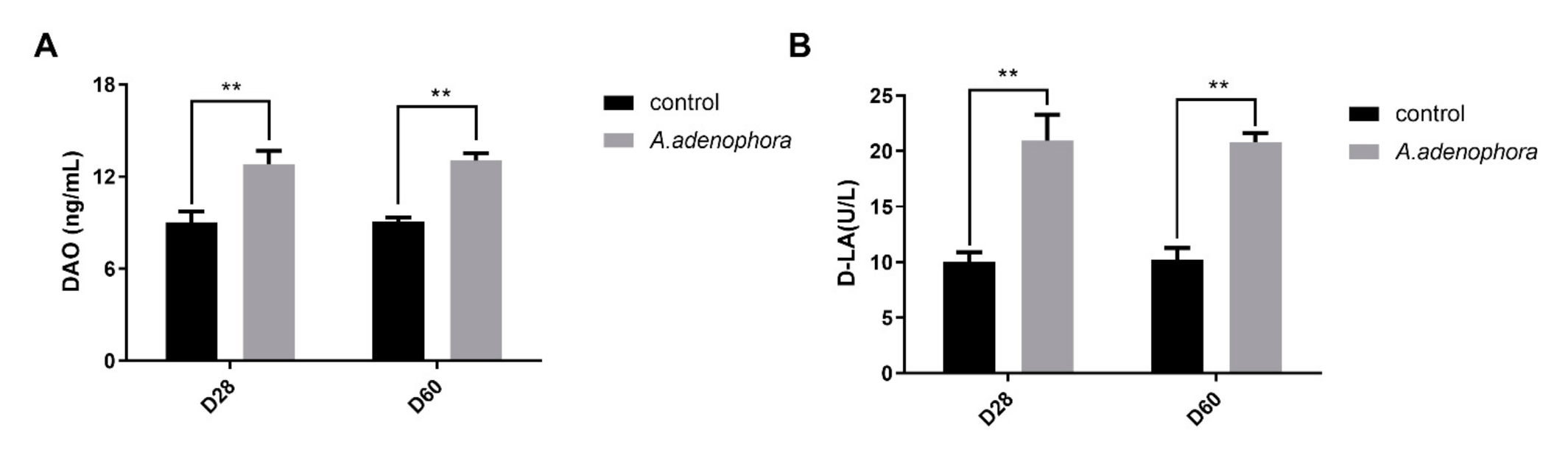
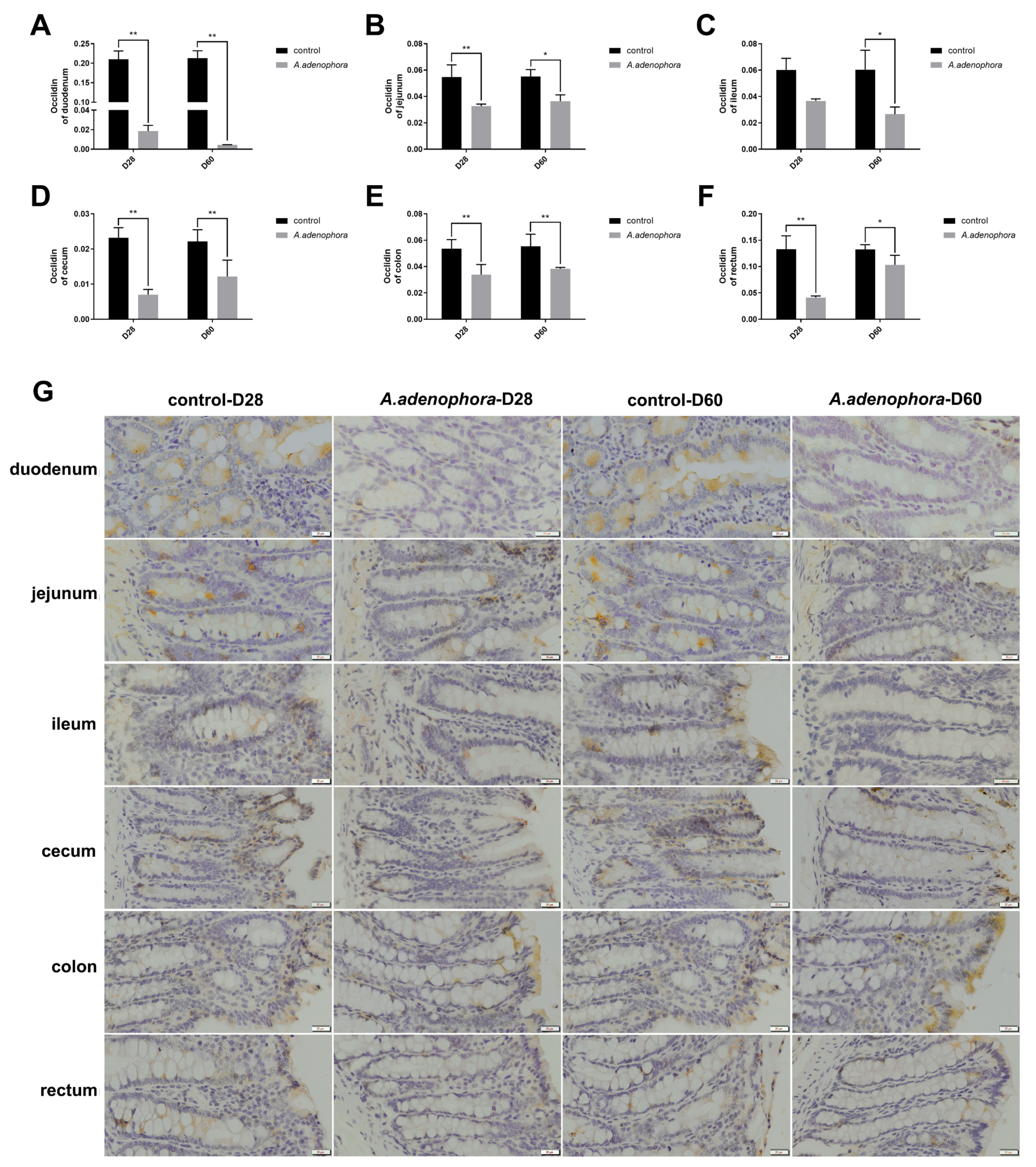
2.2. A. adenophora Disrupts Intestinal Barrier Integrity
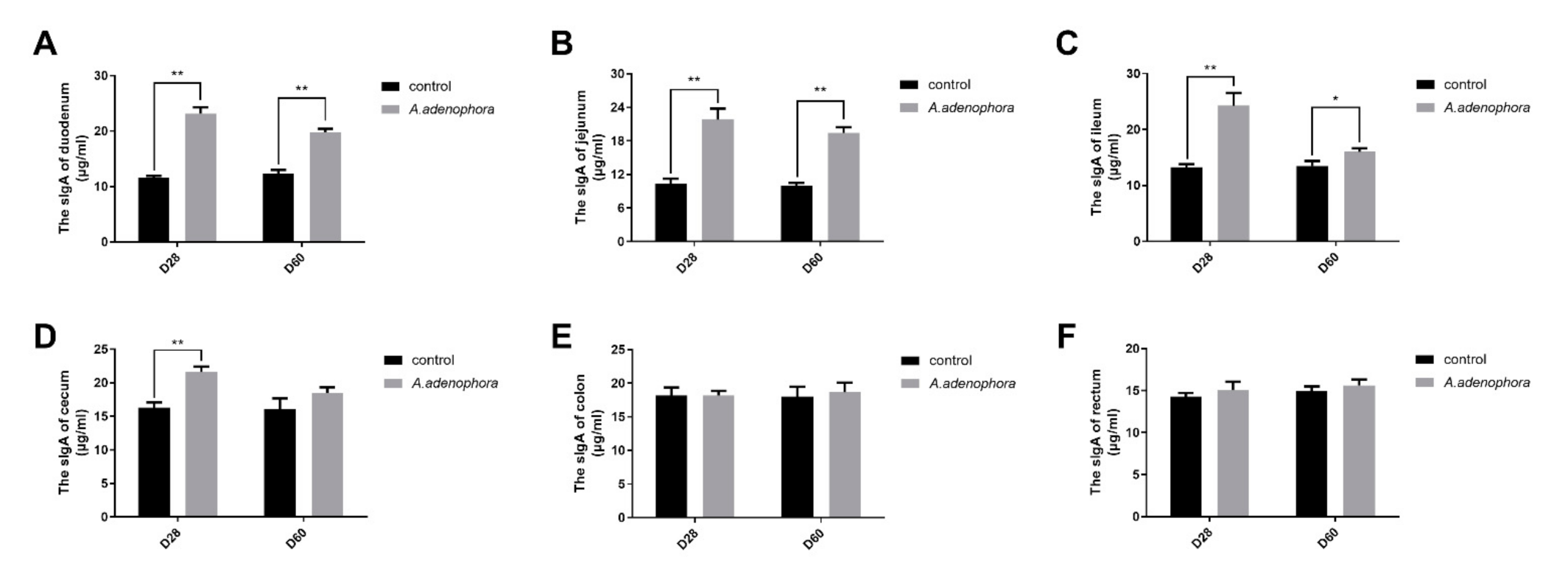
2.3. Effects of A. adenophora on the Concentration of Intestinal sIgA of Rats
2.4. Expression Levels of Intestinal Tissue Pro- and Anti-Inflammation-Related Cytokines Levels
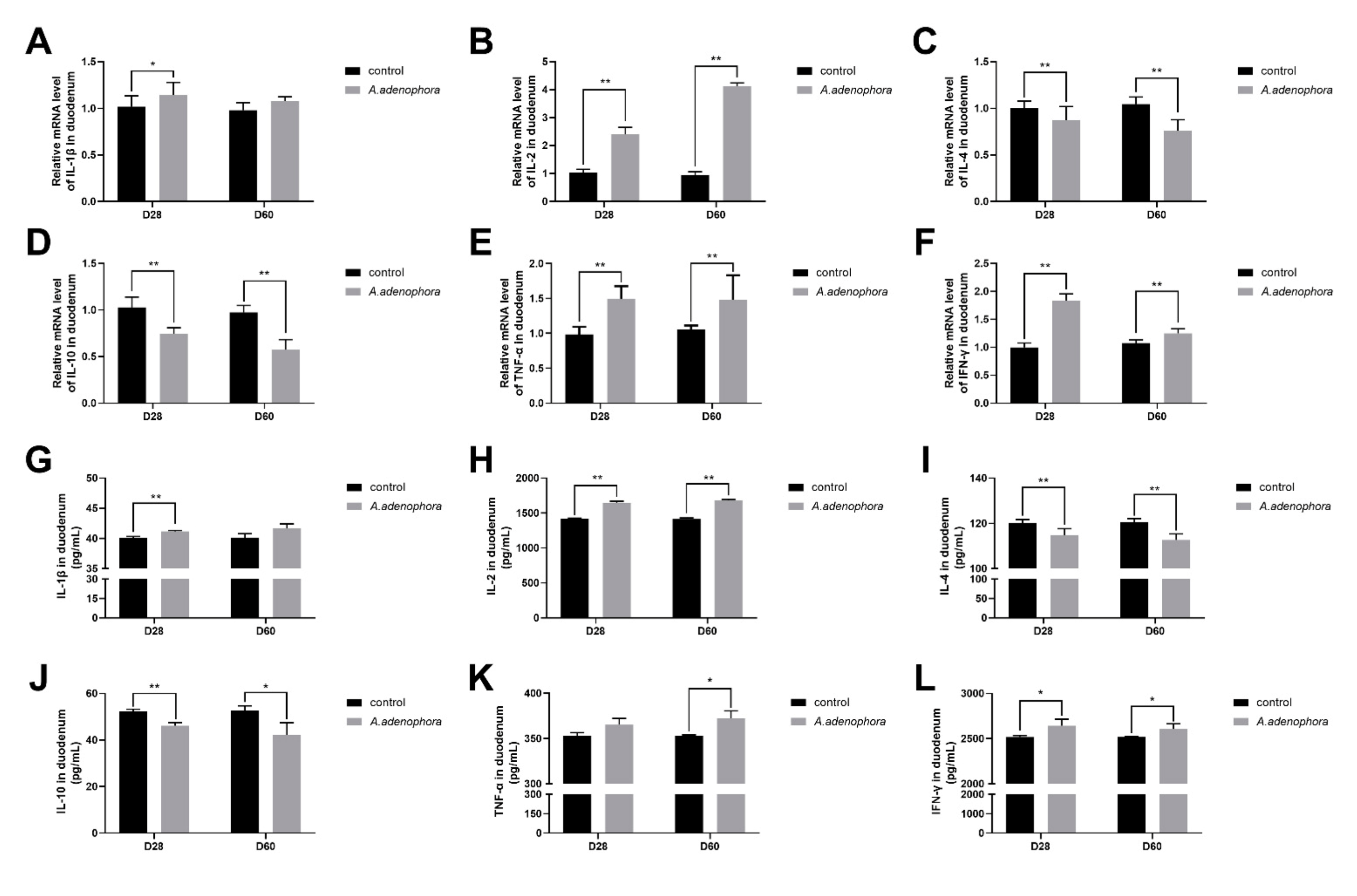
2.4.1. mRNA and Protein (ELISA) Expressions of Pro and Anti-Inflammation-Related Cytokines in Duodenum of A. adenophora-Fed Rats
2.4.2. mRNA and Protein (ELISA) Expression Levels of Pro- and Anti-Inflammation-Related Cytokines in the Jejunum of A. adenophora-Fed Rats
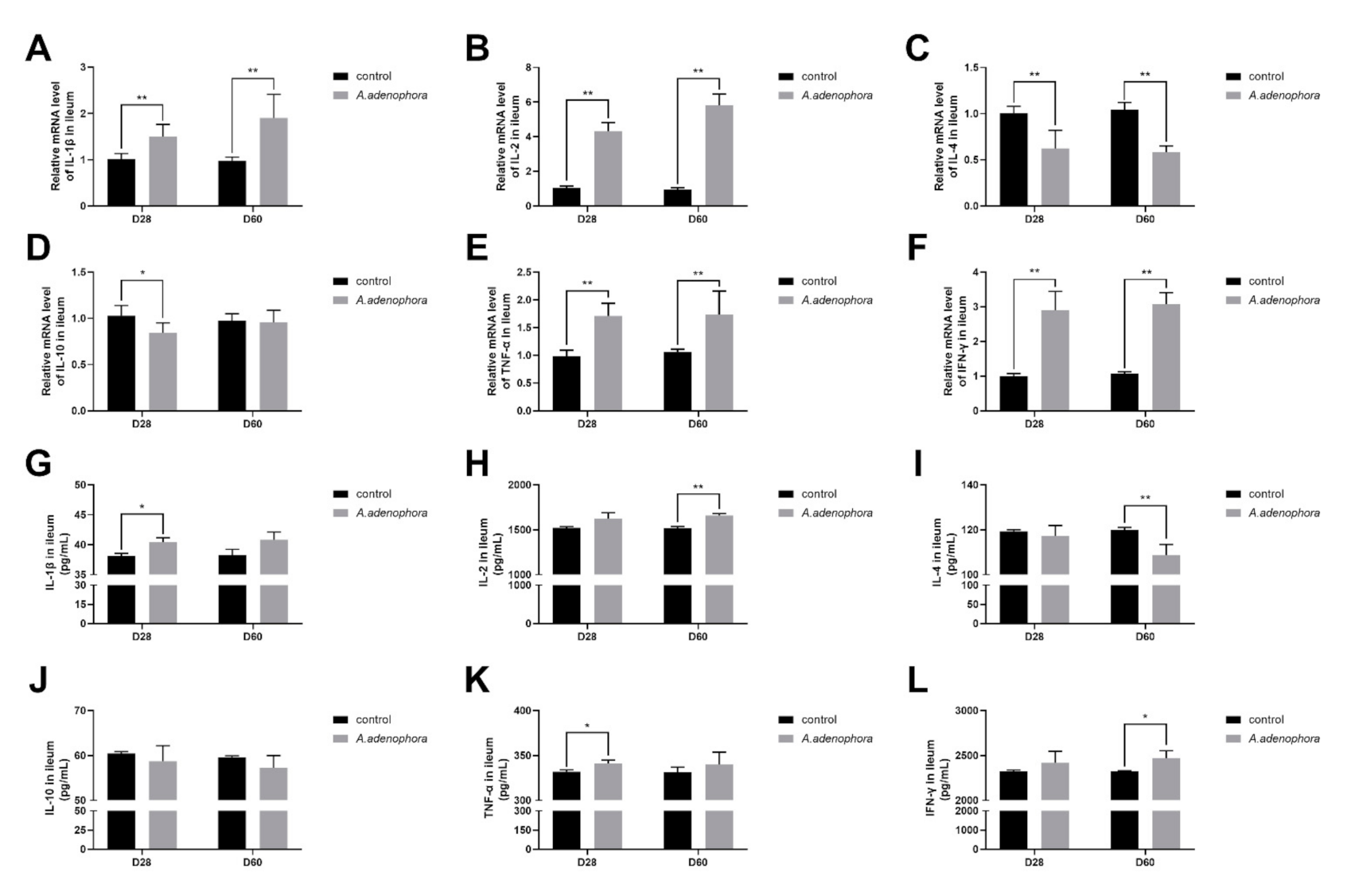
2.4.3. mRNA and Protein (ELISA) Expressions Levels of Pro- and Anti-Inflammation-Related Cytokines in the Ileum of A. adenophora-Fed Rats
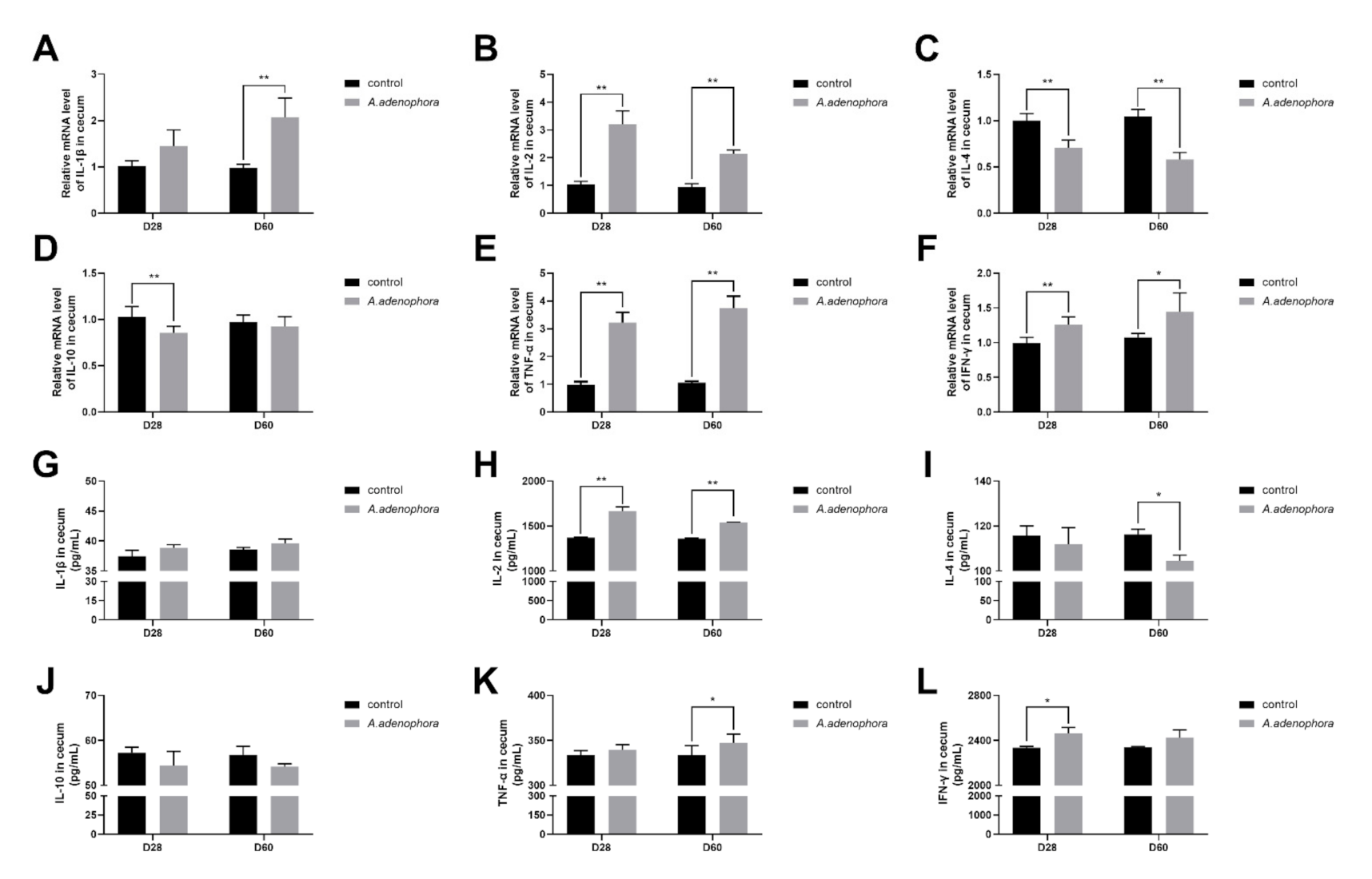
2.4.4. mRNA and Protein (ELISA) Expressions of Pro- and Anti-Inflammation-Related Cytokines in the Cecum of A. adenophora-Fed Rats
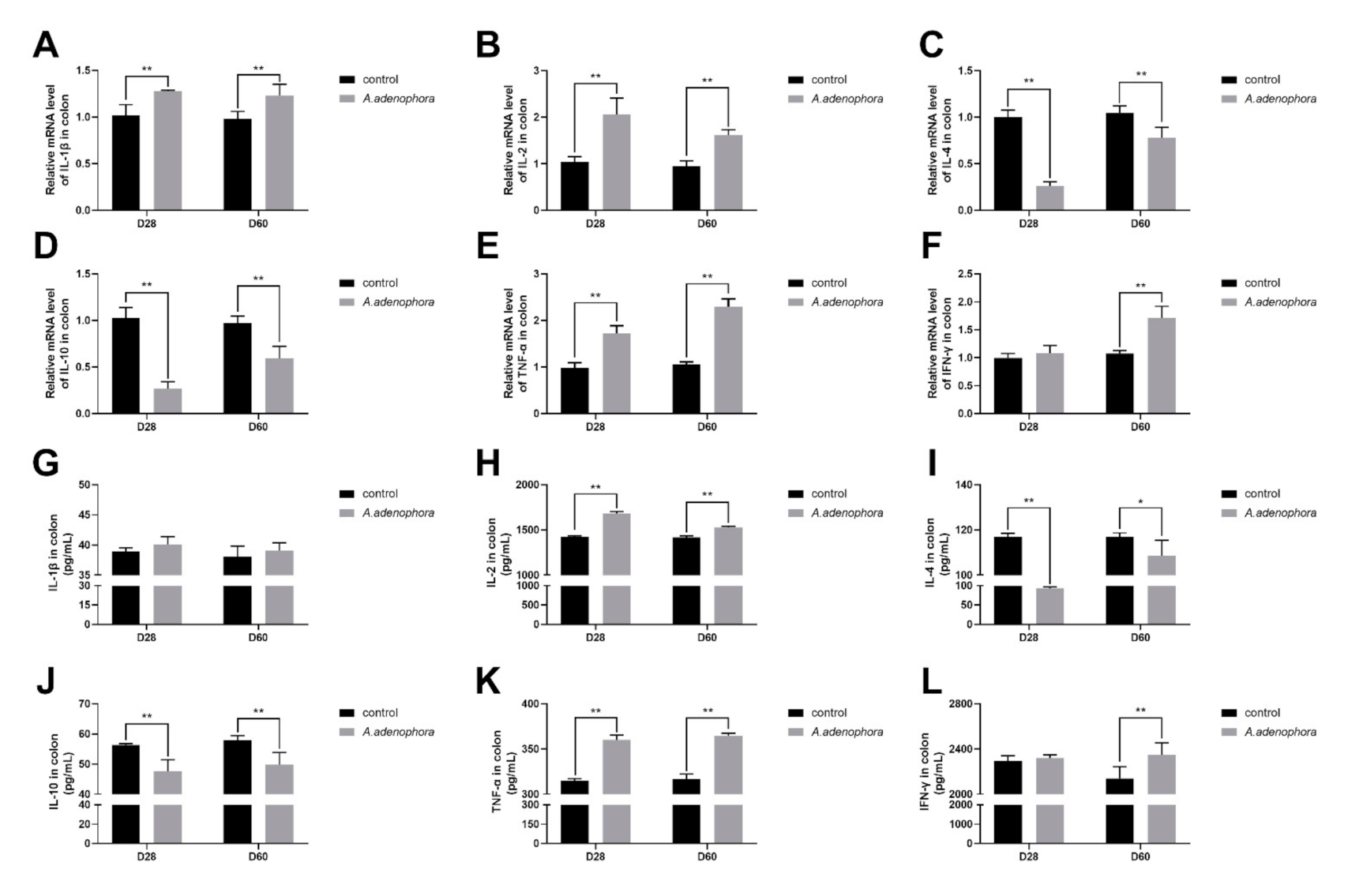
2.4.5. mRNA and Protein (ELISA) Expression Levels of Pro- and Anti-Inflammation-Related Cytokines in the Colon of A. adenophora-Fed Rats
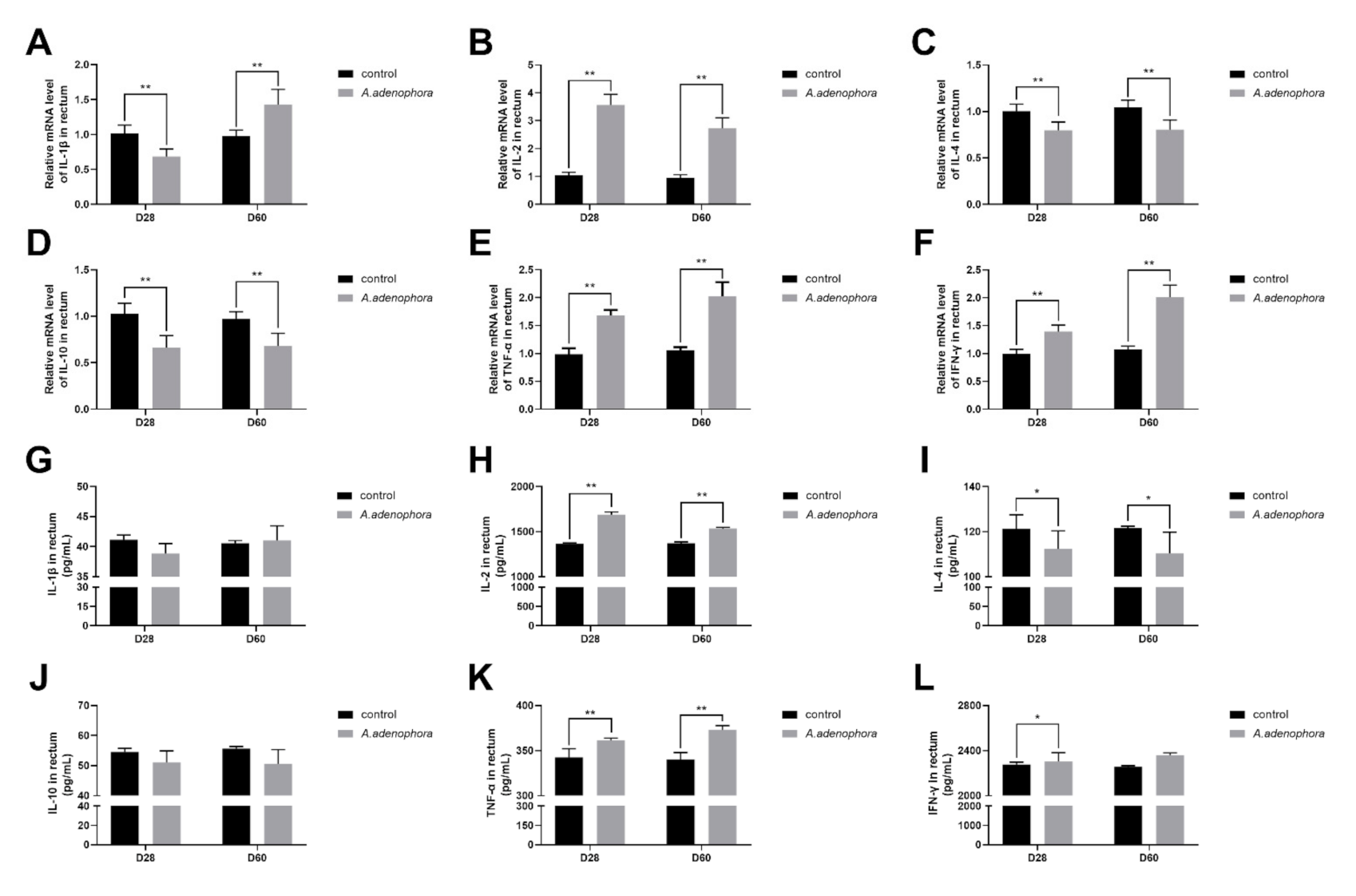
2.4.6. mRNA and Protein (ELISA) Expressions Levels of Pro- and Anti-Inflammation-Related Cytokines in the Rectum of A. adenophora-Fed Rats
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Material
5.2. Animals Experiment and Plant Materials
5.3. Ethics Statement
5.4. Histopathological Evaluation
5.5. Enzyme-Linked Immunosorbent Assay
5.6. Immunohistochemistry Assay
5.7. RNA Extraction and Quantitative Real-Time PCR Analysis
5.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Laboratory Animal Welfare and Ethics guideline
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Qiang, S. The history and status of the study on Crofton weed (Eupatorium adenophorum Spreng), a worldwide weed. J. Wuhan Botan. Res. 1998, 16, 366–372. [Google Scholar]
- Sang, W.; Li, Z.; Axmacher, J. Invasion pattern of Eupatorium adenophorum Spreng in southern China. Biol. Invasions 2010, 12, 1721–1730. [Google Scholar] [CrossRef]
- O’Sullivan, B. Crofton weed (Eupatorium adenophorum) toxicity in horses. Aust. Veter.-J. 1979, 55, 19–21. [Google Scholar] [CrossRef]
- Verma, A.; Yadava, B.; Sampath, K. Possible use of Spreng (Eupatorium adenophorum) in animal feeding. Indian J. Anim. Nutr. 1987, 4, 189–192. [Google Scholar]
- Tripathi, Y.; Saini, N.; Anjum, N.; Verma, P. A review of ethnomedicinal, phytochemical, pharmacological and toxicological aspects of Eupatorium adenophorum spreng. Asian J. Biomed. Pharm. Sci. 2018, 8, 25–35. [Google Scholar]
- Sun, W.; Zeng, C.; Yue, D.; Liu, S.; Ren, Z.; Zuo, Z.; Deng, J.; Peng, G.; Hu, Y. Ageratina adenophora causes spleen toxicity by inducing oxidative stress and pyroptosis in mice. R. Soc. Open Sci. 2019, 6, 190127. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.; Zeng, C.; Yue, D.; Hu, Y. Involvement of mitochondrial dysfunction in hepatotoxicity induced by Ageratina adenophora in mice. J. Zhejiang Univ. Sci. B 2019, 20, 693–698. [Google Scholar] [CrossRef]
- He, Y.; Chen, W.; Hu, Y.; Luo, B.; Wu, L.; Qiao, Y.; Mo, Q.; Xu, R.; Zhou, Y.; Ren, Z.; et al. E. adenophorum induces cell cycle and apoptosis of renal cells through mitochondrial pathway and caspase activation in Saanen goat. PLoS ONE 2015, 10, e0138504. [Google Scholar] [CrossRef] [PubMed]
- Nie, X.; Lv, S.; Zhang, Y.; Du, X.; Wang, L.; Biradar, S.; Tan, X.; Wan, F.; Weining, S. Complete chloroplast genome sequence of a major invasive species, crofton weed (Ageratina adenophora). PLoS ONE 2012, 7, e36869. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Yang, J.; Cao, A.; Liu, Y.; An, Y.; Shi, J. A new sequiterpenoid from Eupatorium adenophorum Spreng. Chin. J. Chem. 2006, 24, 1375–1377. [Google Scholar] [CrossRef]
- Ouyang, C.; Liu, X.; Liu, Q.; Bai, J.; Li, H.; Li, Y.; Wang, Q.; Yan, D.; Mao, L.; Cao, A.; et al. Toxicity assessment of cadinene sesquiterpenes from Eupatorium adenophorum in mice. Nat. Prod. Bioprospect. 2014, 5, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Cao, L.; Zhang, L.; Yuan, X.; Zhao, B. Hepatotoxicity of Eupatorium adenophorum extracts and identification of major hepatotoxic components. Nat. Prod. Res. 2017, 31, 2788–2792. [Google Scholar] [CrossRef]
- Oelrichs, P.B.; Calanasan, C.A.; Macleod, J.K.; Seawright, A.A.; Ng, J.C. The isolation of a compound from Eupatorium adenophorum (Spreng) (Ageratina adenophora) causing hepatotoxicity in mice. Nat. Toxins 1995, 3, 350–354. [Google Scholar] [CrossRef]
- Bhardwaj, R.; Singh, A.; Sharma, O.; Dawra, R.; Kurade, N.; Mahato, S. Hepatotoxicity and cholestasis in rats induced by the sesquiterpene 9-oxo-10,11-dehydroageraphorone isolated from Eupatorium adenophorum. J. Biochem. Mol. Toxicol. 2001, 15, 279–286. [Google Scholar] [CrossRef]
- Okyere, S.; Mo, Q.; Pei, G.; Ren, Z.; Deng, J.; Hu, Y. Euptox A induces G0/G1 arrest and apoptosis of hepatocyte via ROS, mitochondrial dysfunction and caspases-dependent path- ways in vivo. J. Toxicol. Sci. 2020, 45, 661–671. [Google Scholar] [CrossRef]
- Mo, Q.; Hu, L.; Weng, J.; Zhang, Y.; Zhou, Y.; Xu, R.; Zuo, Z.; Deng, J.; Ren, Z.; Zhong, Z.; et al. Euptox A induces G1 arrest and autophagy via p38 MAPK- and PI3K/Akt/mTOR-mediated pathways in mouse splenocytes. J. Histochem. Cytochem. 2017, 65, 543–558. [Google Scholar] [CrossRef] [Green Version]
- Chelakkot, C.; Ghim, J.; Ryu, S. Mechanisms regulating intestinal barrier integrity and its pathological implications. Exp. Mol. Med. 2018, 50, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odenwald, M.; Turner, J. The intestinal epithelial barrier: A therapeutic target? Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 9–21. [Google Scholar] [CrossRef]
- De Mey, J.; Freund, J. Understanding epithelial homeostasis in the intestine: An old battlefield of ideas, recent breakthroughs and remaining controversies. Tissue Barriers 2013, 1, e24965. [Google Scholar] [CrossRef] [Green Version]
- Catalioto, R.; Maggi, C.; Giuliani, S. Intestinal epithelial barrier dysfunction in disease and possible therapeutical interventions. Curr. Med. Chem. 2011, 18, 398–426. [Google Scholar] [CrossRef]
- Kaushal, V.; Dawra, R.; Sharma, O.; Kurade, N. Biochemical Alterations in the Blood Plasma of Rats Associated with Hepatotoxicity Induced by Eupatorium adenophorum. Vet. Res. Commun. 2001, 25, 601–608. [Google Scholar] [CrossRef]
- Ouyang, C.; Liu, X.; Yan, D.; Li, Y.; Wang, Q.; Cao, A.; Guo, M. Immunotoxicity assessment of cadinene sesquiterpenes from Eupatorium adenophorum in mice. J. Integr. Agric. 2016, 15, 2319–2325. [Google Scholar] [CrossRef] [Green Version]
- Feng, Z.; Li, T.; Wu, C.; Tao, L.; Blachier, F.; Yin, Y. Monosodium L-glutamate and dietary fat exert opposite effects on the proximal and distal intestinal health in growing pigs. Appl. Physiol. Nutr. Metab. 2016, 40, 353–363. [Google Scholar] [CrossRef]
- Fuller, M. The Encyclopedia of Farm Animal Nutrition; CAB International: Wallingford, UK, 2004; pp. 244–246. [Google Scholar]
- Chwen, L.; Foo, H.; Thanh, N.; Choe, D. Growth performance, plasma Fatty acids, villous height and crypt depth of preweaning piglets fed with medium chain triacylglycerol. Asian-Australas. J. Anim. Sci. 2013, 26, 700–704. [Google Scholar] [CrossRef]
- Xue, G.; Barekatain, R.; Wu, S.; Choct, M.; Swick, R. Dietary L-glutamine supplementation improves growth performance, gut morphology, and serum biochemical indices of broiler chickens during necrotic enteritis challenge. Poult. Sci. 2018, 97, 1334–1341. [Google Scholar] [CrossRef]
- Saeid, J.; Mohamed, A.; Al-Baddy, M. Effect of garlic powder (Allium sativum) and black seed (Nigella sativa) on broiler growth performance and intestinal morphology. Iran. J. Appl. Anim. Sci. 2013, 3, 185–188. [Google Scholar]
- Talcott, P. Black Locust (Robinia pseudoacacia) Chapter 21-Toxicologic problems. In Equine Internal Medicine, 4th ed.; Elsevier Health Sciences: New York, NY, USA, 2018; pp. 1460–1512. [Google Scholar]
- Cheroutre, H. Starting at the beginning: New perspectives on the biology of mucosal T cells. Annu. Rev. Immunol. 2004, 22, 217–246. [Google Scholar] [CrossRef]
- Tamura, A.; Soga, H.; Yaguchi, K.; Yamagishi, M.; Toyota, T.; Sato, J.; Oka, Y.; Itoh, T. Distribution of two types of lymphocytes (intraepithelial and lamina-propria-associated) in the murine small intestine. Cell Tissue Res. 2003, 313, 47–53. [Google Scholar] [CrossRef]
- Chang, F.; Mahadeva, U.; Deere, H. Pathological and clinical significance of increased intraepithelial lymphocytes (IELs) in small bowel mucosa. Review article. APMIS 2005, 113, 385–399. [Google Scholar] [CrossRef]
- Xin, J.; Zeng, D.; Wang, H.; Sun, N.; Zhao, Y.; Dan, Y.; Pan, K.; Jing, B.; Ni, X. Probiotic Lactobacillus johnsonii BS15 Promotes Growth Performance, Intestinal Immunity, and Gut Microbiota in Piglets. Probiotics Antimicrob. Proteins 2020, 12, 184–193. [Google Scholar] [CrossRef]
- Hagmeyer, R. Heal Your Gut by Balancing Your SIgA Levels. Available online: https://www.drhagmeyer.com/heal-your-gut-by-balancing-your-siga-levels/ (accessed on 17 March 2021).
- Lin, M.; Du, L.; Brandtzaeg, P.; Pan-Hammarström, Q. IgA subclass switch recombination in human mucosal and systemic immune compartments. Mucosal Immunol. 2014, 7, 511–520. [Google Scholar] [CrossRef] [Green Version]
- Kanerud, L.; Engstrom, G.; Tarkowski, A. Evidence for differential effects of sulphasalazine on systemic and mucosal immunity in rheumatoid arthritis. Ann. Rheum. Dis. 1995, 54, 256–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.-D.; He, Y.; Yu, H.-Y.; Liu, Y.-L.; Ge, Y.-X.; Li, X.-T.; Wang, Y.; Guo, M.-R.; Qu, Y.-L.; Qin, X.-F.; et al. Unconjugated bilirubin alleviates experimental ulcerative colitis by regulating intestinal barrier function and immune inflammation. World J. Gastroenterol. 2019, 25, 1865–1878. [Google Scholar] [CrossRef] [PubMed]
- Antoni, L.; Nuding, S.; Wehkamp, J.; Stange, E. Intestinal barrier in inflammatory bowel disease. World J. Gastroenterol. 2014, 20, 1165–1179. [Google Scholar] [CrossRef]
- Honzawa, Y.; Nakase, H.; Matsuura, M.; Chiba, T. Clinical significance of serum diamine oxidase activity in inflammatory bowel disease: Importance of evaluation of small intestinal permeability. Inflamm. Bowel Dis. 2011, 17, E23–E25. [Google Scholar] [CrossRef]
- Tian, J.; Ke, X.; Yuan, Y.; Yang, W.; Tang, X.; Qu, J.; Qu, W.; Fu, S.; Zheng, Y.; Fan, J.; et al. Two generation reproduction toxicity study of GmDREB3 gene modified wheat in Wistar rats. Food Chem. Toxicol. 2021, 153, 112310. [Google Scholar] [CrossRef]
- Ruan, P.; Gong, Z.J.; Zhang, Q.R. Changes of plasma D(-)-lactate, diamine oxidase and endotoxin in patients with liver cirrhosis. Hepatobiliary Pancreat. Dis. Int. 2004, 3, 58–61. [Google Scholar] [PubMed]
- Chen, J.L.; Zheng, P.; Zhang, C.; Yu, B.; He, J.; Yu, J.; Luo, J.Q.; Mao, X.B.; Huang, Z.Q.; Chen, D.W. Benzoic acid beneficially affects growth performance of weaned pigs which was associated with changes in gut bacterial populations, morphology indices and growth factor gene expression. J. Anim. Physiol. Anim. Nutr. 2016, 101, 1137–1146. [Google Scholar] [CrossRef]
- Chen, F.; Chen, J.; Chen, Q.; Yang, L.; Yin, J.; Li, Y.; Huang, X. Lactobacillus delbrueckii Protected Intestinal Integrity, Alleviated Intestinal Oxidative Damage, and Activated Toll-Like Receptor–Bruton’s Tyrosine Kinase–Nuclear Factor Erythroid 2-Related Factor 2 Pathway in Weaned Piglets Challenged with Lipopolysaccharide. Antioxidants 2021, 10, 468. [Google Scholar]
- Yang, L.; Liu, G.; Lian, K.; Qiao, Y.; Zhang, B.; Zhu, X.; Luo, Y.; Shang, Y.; Gu, X. Dietary leonurine hydrochloride supplementation attenuates lipopolysaccharide challenge-induced intestinal inflammation and barrier dysfunction by inhibiting the NF-κB/MAPK signaling pathway in broilers. J. Anim. Sci. 2019, 97, 1679–1692. [Google Scholar] [CrossRef]
- Kumar, A.; Chatterjee, I.; Anbazhagan, A.N.; Jayawardena, D.; Priyamvada, S.; Alrefai, W.A.; Sun, J.; Borthakur, A.; Dudeja, P.K. Cryptosporidium parvum disrupts intestinal epithelial barrier function via altering expression of key tight junction and adherens junction proteins. Cell Microbiol. 2018, 20, e12830. [Google Scholar] [CrossRef]
- Marchiando, A.; Graham, W.; Turner, J. Epithelial barriers in homeostasis and disease. Annu. Rev. Pathol. 2010, 5, 119–144. [Google Scholar] [CrossRef] [PubMed]
- Jeong, C.H.; Kwon, H.C.; Kim, D.H.; Cheng, W.N.; Kang, S.; Shin, D.-M.; Yune, J.H.; Yoon, J.E.; Chang, Y.H.; Sohn, H.; et al. Effects of Aluminum on the Integrity of the Intestinal Epithelium: An in Vitro and in Vivo Study. Environ. Health Perspect. 2020, 128, 17013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S. Intestinal permeability regulation by tight junction: Implication on inflammatory bowel diseases. Intest. Res. 2015, 13, 11–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, G.; Weber, C. Molecular aspects of tight junction barrier function. Curr. Opin. Pharmacol. 2014, 19, 84–89. [Google Scholar] [CrossRef] [Green Version]
- Van Kaer, L.; Olivares-Villagomez, D. Development, Homeostasis, and Functions of Intestinal Intraepithelial Lymphocytes. J. Immunol. 2018, 200, 2235–2244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Sadi, R.; Khatib, K.; Guo, S.; Ye, D.; Youssef, M.; Ma, T. Occludin regulates macromolecule flux across the intestinal epithelial tight junction barrier. Am. J. Physiol. Liver Physiol. 2011, 300, G1054–G1064. [Google Scholar] [CrossRef] [Green Version]
- Buschmann, M.M.; Shen, L.; Rajapakse, H.; Raleigh, D.R.; Wang, Y.; Wang, Y.; Lingaraju, A.; Zha, J.; Abbott, E.; McAuley, E.M.; et al. Occludin OCEL-domain interactions are required for maintenance and regulation of the tight junction barrier to macromolecular flux. Mol. Biol. Cell 2013, 24, 3056–3068. [Google Scholar] [CrossRef]
- Romero, A.; Ares, I.; Ramos, E.; Castellano, V.; Martínez, M.; Martínez-Larrañaga, M.R.; Anadón, A.; Martínez, M.A. Mycotoxins modify the barrier function of Caco-2 cells through differential gene expression of specific claudin isoforms: Protective effect of illite mineral clay. Toxicology 2016, 353–354, 21–33. [Google Scholar] [CrossRef]
- Jiang, N.; Li, Y.; Shu, T.; Wang, J. Cytokines and inflammation in adipogenesis: An updated review. Front. Med. 2019, 13, 314–329. [Google Scholar] [CrossRef]
- Zhang, K.; Shi, S.; Han, W. Research progress in cytokines with chemokine-like function. Cell Mol Immunol. 2018, 15, 660–662. [Google Scholar] [CrossRef]
- Farré, R.; Fiorani, M.; Abdu Rahiman, S.; Matteoli, G. Intestinal Permeability, Inflammation and the Role of Nutrients. Nutrients 2020, 12, 1185. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xue, Y.; Li, Y.; Liu, F.; Yan, Y.; Zhang, H.; Jin, Q. Effects of Isatis root polysaccharide in mice infected with H3N2 swine influenza virus. Res. Vet. Sci. 2018, 119, 91–98. [Google Scholar] [CrossRef]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef]
- Nathan, C. Points of control in inflammation. Nature 2002, 420, 846–852. [Google Scholar] [CrossRef]
- Chen, W. Effects of Different Doses of Eupatorium adenophorum on Clinical Pathology of Saanen Dairy Goats. Master’s Thesis, Sichuan Agricultural University, Sichuan, China, 2015. [Google Scholar]
- Van Itallie, C.; Anderson, J. Claudins and epithelial paracellular transport. Annu Rev. Physiol. 2006, 68, 403–429. [Google Scholar] [CrossRef]
- Bai, J.; Cao, A.; Guo, M.; Liu, X.; Liu, X.; Liang, H.; Zhou, B. Identification of 9-oxo-10,11- dehydroagerophorone in Eupatorium Adenophorum by HPLC. Chin. Bull. Bot. 2011, 46, 470–475. [Google Scholar]
- Bai, J. Foundational Studies of Toxicity Substances in Exotic Invasive Plant Eupatorium adenophorum Spreng. Ph.D. Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2012. [Google Scholar]
- Fu, J.; Hu, L.; Shi, Z.; Sun, W.; Yue, D.; Wang, Y.; Ma, X.; Ren, Z.; Zuo, Z.; Peng, G.; et al. Two metabolites isolated from endophytic fungus Coniochaeta sp. F-8 in Ageratina adenophora exhibit antioxidative activity and cytotoxicity. Nat. Prod. Res. 2019, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Zeng, C.; Liu, S.; Fu, J.; Hu, L.; Shi, Z.; Yue, D.; Ren, Z.; Zhong, Z.; Zuo, Z.; et al. Ageratina adenophora induces mice hepatotoxicity via ROS-NLRP3-mediated pyroptosis. Sci. Rep. 2018, 8, 16032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, Z.; Gao, P.; Okyere, S.K.; Cui, Y.; Wen, J.; Jing, B.; Deng, J.; Hu, Y. Ageratina adenophora inhibits spleen immune function in rats via the loss of the FRC network and Th1-Th2 cell ratio elevation. Toxins 2021, 13, 309. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Intestinal Segment | Index | Treatments | D28 | D60 |
|---|---|---|---|---|
| duodenum | Villus height | Control | 488.63 ± 35.29 A | 485.42 ± 25.87 A |
| A. adenophora | 390.11 ± 28.23 B | 384.76 ± 26.39 B | ||
| crypt depth | Control | 78.88 ± 15.50 B | 74.91 ± 14.37 B | |
| A. adenophora | 107.51 ± 6.72 A | 135.81 ± 35.53 A | ||
| Villus height/crypt depth ratio | Control | 6.32 ± 0.54 A | 6.41 ± 0.49 A | |
| A. adenophora | 3.51 ± 0.40 B | 3.14 ± 0.87 B | ||
| jejunum | Villus height | Control | 400.55 ± 10.96 A | 418.26 ± 12.75 A |
| A. adenophora | 275.63 ± 40.78 B | 258.43 ± 28.20 B | ||
| crypt depth | Control | 89.72 ± 11.73 B | 89.34 ± 15.15 B | |
| A. adenophora | 100.18 ± 10.32 A | 193.43 ± 25.19 A | ||
| Villus height/crypt depth ratio | Control | 4.49 ± 0.57 A | 4.69 ± 0.48 A | |
| A. adenophora | 2.82 ± 0.43 B | 1.36 ± 0.23 B | ||
| ileum | Villus height | Control | 367.09 ± 25.05 A | 369.49 ± 27.86 A |
| A. adenophora | 205.11 ± 36.04 B | 267.88 ± 24.93 B | ||
| crypt depth | Control | 71.32 ± 11.05 B | 80.82 ± 12.32 B | |
| A. adenophora | 107.79 ± 6.80 A | 163.53 ± 13.79 A | ||
| Villus height/crypt depth ratio | Control | 5.03 ± 0.39 A | 4.97 ± 0.75 A | |
| A. adenophora | 2.33 ± 0.34 B | 1.66 ± 0.24 C |
| Ingredients | Content % |
|---|---|
| Water | 10 |
| Crude protein | 18 |
| Crude fat | 4 |
| Crude fiber | 5 |
| Crude ash | 8 |
| Calcium | 1.5 |
| Phosphorus | 1 |
| Lysine | 0.82 |
| Methionine + Cystine | 0.53 |
| Appearance | Score |
|---|---|
| Edema | 0 or 1 (absent or present) |
| Angiogenesis | 0 or 1 (absent or present) |
| Muscle thickening | 0, 1, 2, or 3 (absent, mild→severe) |
| Cellular infiltration | 0, 1, 2, or 3 (absent, mild→severe) |
| Loss of mucosal architecture | 0, 1, 2, or 3 (absent, mild→severe) |
| Gene Name | Primer | Sequence (5′ and 3′) | Product Length (bp) | Annealing Temperature (°C) | Sequence Number |
|---|---|---|---|---|---|
| IL-1β | Forward | GCAGCTTTCGACAGTGAGGA | 86 | 55.0 | NM_031512.2 |
| Reverse | CCCAAGTCAAGGGCTTGGAA | ||||
| IL-2 | Forward | TCTGCAGCGTGTGTTGGATT | 143 | 52.38 | NM_053836.1 |
| Reverse | CTGGCTCATCATCGAATTGGC | ||||
| IL-4 | Forward | TGTAGAGGTGTCAGCGGTCT | 70 | 60.25 | NM_201270.1 |
| Reverse | TCAGTGTTGTGAGCGTGGAC | ||||
| IL-10 | Forward | CTGGCTCAGCACTGCTATGT | 86 | 60.11 | NM_012854.2 |
| Reverse | GCAGTTATTGTCACCCCGGA | ||||
| TNF-α | Forward | CTCAAGCCCTGGTATGAGCC | 90 | 60.18 | NM_012675.3 |
| Reverse | CTTGGGCAGGTTGACCTCAG | ||||
| IFN-γ | Forward | CATCGCCAAGTTCGAGGTGA | 87 | 60.00 | NM_138880.3 |
| Reverse | TCTGGTGACAGCTGGTGAATC | ||||
| β-actin | Forward | ACGGTCAGGTCATCACTATCG | 112 | 65.20 | NM_031144.3 |
| Reverse | GGCATAGAGGTCTTTACGGATG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, Y.; Okyere, S.K.; Gao, P.; Wen, J.; Cao, S.; Wang, Y.; Deng, J.; Hu, Y. Ageratina adenophora Disrupts the Intestinal Structure and Immune Barrier Integrity in Rats. Toxins 2021, 13, 651. https://doi.org/10.3390/toxins13090651
Cui Y, Okyere SK, Gao P, Wen J, Cao S, Wang Y, Deng J, Hu Y. Ageratina adenophora Disrupts the Intestinal Structure and Immune Barrier Integrity in Rats. Toxins. 2021; 13(9):651. https://doi.org/10.3390/toxins13090651
Chicago/Turabian StyleCui, Yujing, Samuel Kumi Okyere, Pei Gao, Juan Wen, Suizhong Cao, Ya Wang, Junliang Deng, and Yanchun Hu. 2021. "Ageratina adenophora Disrupts the Intestinal Structure and Immune Barrier Integrity in Rats" Toxins 13, no. 9: 651. https://doi.org/10.3390/toxins13090651
APA StyleCui, Y., Okyere, S. K., Gao, P., Wen, J., Cao, S., Wang, Y., Deng, J., & Hu, Y. (2021). Ageratina adenophora Disrupts the Intestinal Structure and Immune Barrier Integrity in Rats. Toxins, 13(9), 651. https://doi.org/10.3390/toxins13090651