Mechanisms of Action of the Peptide Toxins Targeting Human and Rodent Acid-Sensing Ion Channels and Relevance to Their In Vivo Analgesic Effects

 , , , , and
, , , , and
Abstract
:1. Molecular and Functional Properties of ASICs
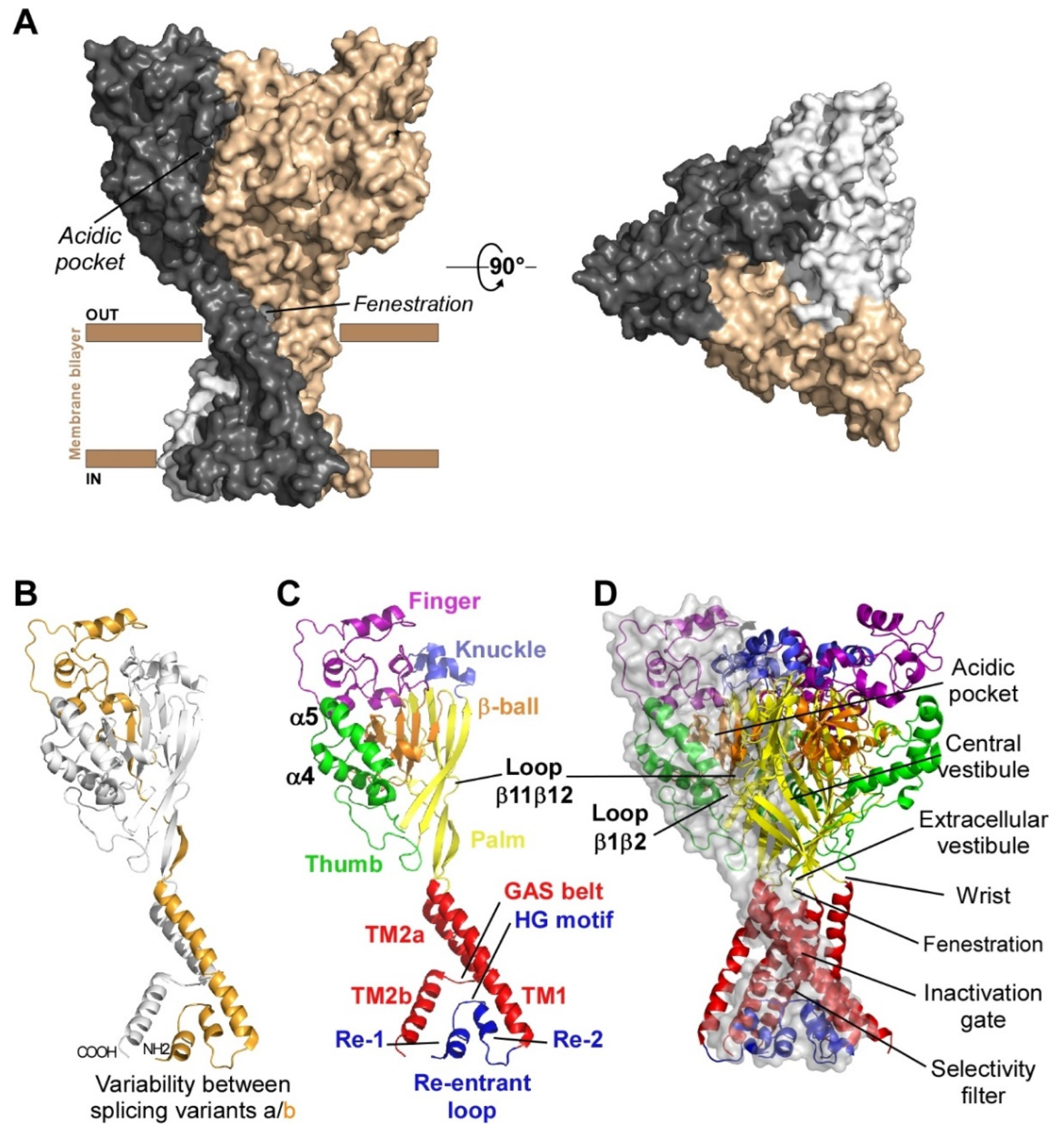
1.1. Subunits Diversity and Structure
1.2. pH-Dependency
1.3. pH-Dependent Gating
1.4. Pathophysiological Roles in Pain Sensing
2. Dual Effects of Animal Toxins Targeting ASICs
2.1. Preliminary Remarks on the Models and Data Interpretation
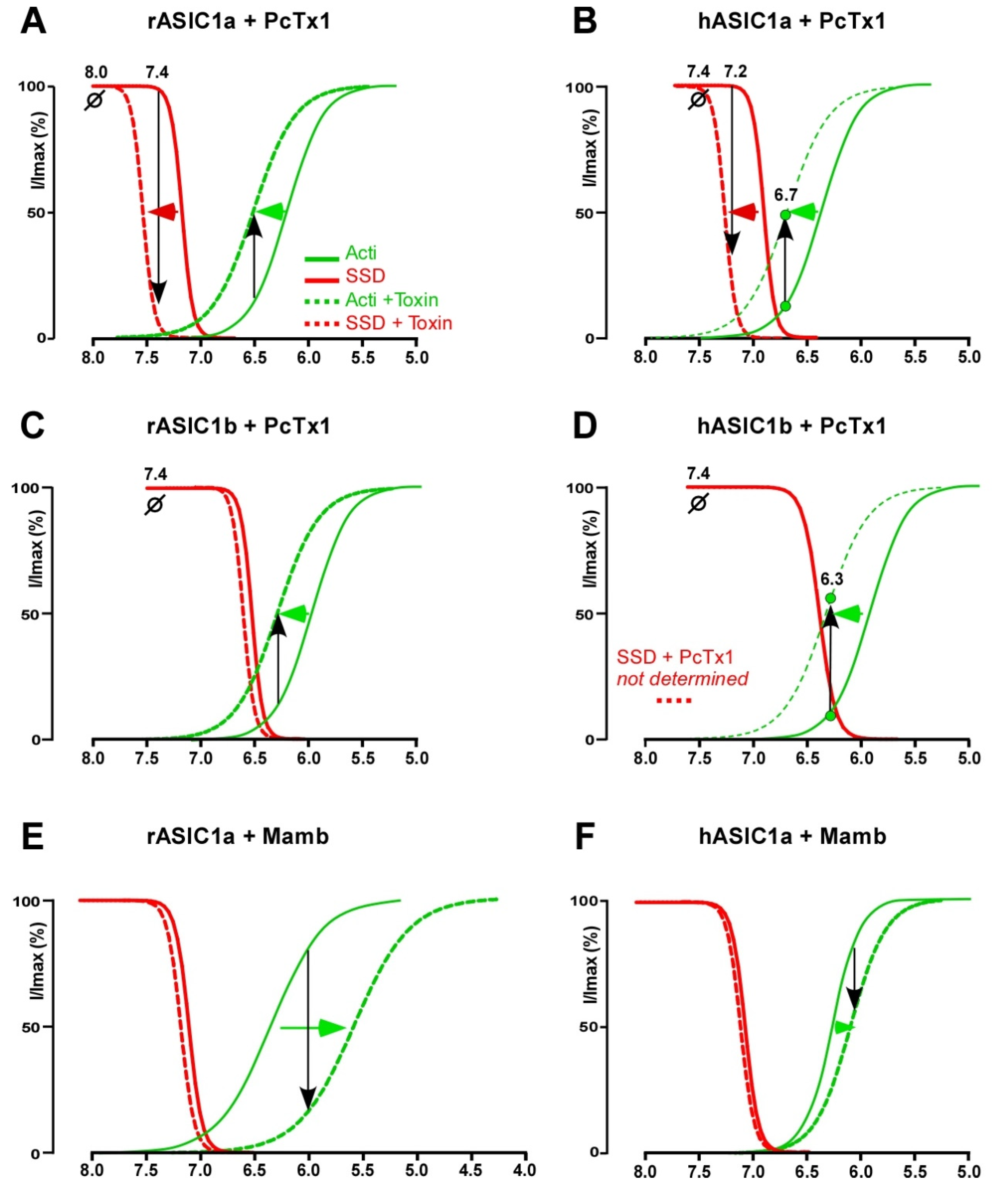
2.2. PcTx1 and Related Toxins
2.2.1. Pharmacological Profile
2.2.2. PcTx1, a Gating Modifier Stabilizing Open and Desensitized States
Biophysical Mechanisms
Structural Mechanisms
PcTx1-Related Compounds
2.3. MitTx, a Painful Toxin
2.3.1. Pharmacological Profile
2.3.2. MitTx, a Gating Modifier Stabilizing the Open State
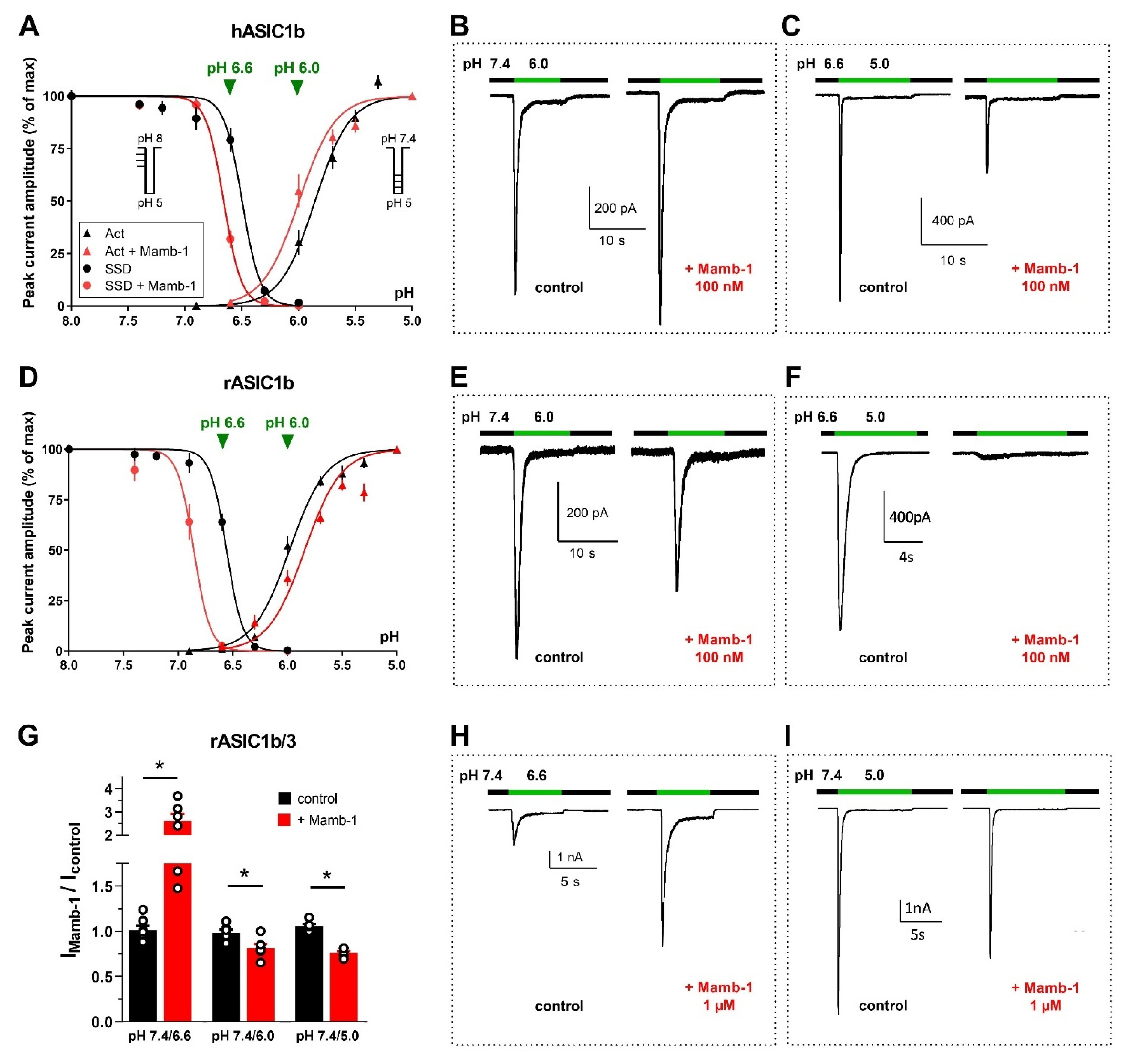
2.4. Mambalgin
2.4.1. Pharmacological Profile
2.4.2. Mambalgin, a Gating Modifier Stabilizing the Closed State
Biophysical Mechanisms and Relevance to In Vivo Analgesic Effects
Structural Mechanisms
2.5. APETx2 and APETx-like Peptides
2.5.1. Pharmacological Profile
2.5.2. APETx2, a Pore Blocker?
2.6. Other Animal Toxins Targeting ASICs
3. Expression of ASICs in the Nervous System and Peptide Toxin Effects on Native Currents
3.1. Expression of ASICs in Neurons
3.1.1. Expression in Peripheral Sensory Neurons
3.1.2. Expression in Central Neurons
3.1.3. Expression in Glial Cells
3.2. Effects of ASIC-Targeting Peptide Toxins on Native Currents
4. Pathophysiological Relevance of ASICs and in Vivo Effects of ASIC-Targeting Peptide Toxins
4.1. Relevance in Pain
4.2. Relevance in Other Pathological Situations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Krishtal, O.A.; Pidoplichko, V.I. A receptor for protons in the nerve cell membrane. Neuroscience 1980, 5, 2325–2327. [Google Scholar] [CrossRef]
- Waldmann, R.; Bassilana, F.; de Weille, J.; Champigny, G.; Heurteaux, C.; Lazdunski, M. Molecular cloning of a non-inactivating proton-gated Na+ channel specific for sensory neurons. J. Biol. Chem. 1997, 272, 20975–20978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waldmann, R.; Champigny, G.; Bassilana, F.; Heurteaux, C.; Lazdunski, M. A proton-gated cation channel involved in acid-sensing. Nature 1997, 386, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Kellenberger, S.; Schild, L.; Ohlstein, E.H. International Union of Basic and Clinical Pharmacology. XCI. Structure, Function, and Pharmacology of Acid-Sensing Ion Channels and the Epithelial Na+ Channel. Pharmacol. Rev. 2015, 67, 1–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenbaum, T.; Morales-Lazaro, S.L.; Islas, L.D. TRP channels: A journey towards a molecular understanding of pain. Nat. Rev. Neurosci. 2022, 23, 596–610. [Google Scholar] [CrossRef]
- Comes, N.; Gasull, X.; Callejo, G. Proton Sensing on the Ocular Surface: Implications in Eye Pain. Front. Pharm. 2021, 12, 773871. [Google Scholar] [CrossRef]
- Aneiros, E.; Cao, L.; Papakosta, M.; Stevens, E.B.; Phillips, S.; Grimm, C. The biophysical and molecular basis of TRPV1 proton gating. EMBO J. 2011, 30, 994–1002. [Google Scholar] [CrossRef]
- Feliciangeli, S.; Chatelain, F.C.; Bichet, D.; Lesage, F. The family of K2P channels: Salient structural and functional properties. J. Physiol. 2015, 593, 2587–2603. [Google Scholar] [CrossRef] [Green Version]
- Napolitano, L.M.R.; Torre, V.; Marchesi, A. CNG channel structure, function, and gating: A tale of conformational flexibility. Pflug. Arch. 2021, 473, 1423–1435. [Google Scholar] [CrossRef]
- Marchesi, A.; Arcangeletti, M.; Mazzolini, M.; Torre, V. Proton transfer unlocks inactivation in cyclic nucleotide-gated A1 channels. J. Physiol. 2015, 593, 857–870. [Google Scholar] [CrossRef]
- Zha, X.M.; Xiong, Z.G.; Simon, R.P. pH and proton-sensitive receptors in brain ischemia. J. Cereb. Blood Flow Metab. 2022, 42, 1349–1363. [Google Scholar] [CrossRef] [PubMed]
- Jasti, J.; Furukawa, H.; Gonzales, E.B.; Gouaux, E. Structure of acid-sensing ion channel 1 at 1.9 A resolution and low pH. Nature 2007, 449, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Lingueglia, E.; de Weille, J.R.; Bassilana, F.; Heurteaux, C.; Sakai, H.; Waldmann, R.; Lazdunski, M. A modulatory subunit of acid sensing ion channels in brain and dorsal root ganglion cells. J. Biol. Chem. 1997, 272, 29778–29783. [Google Scholar] [CrossRef] [Green Version]
- Hesselager, M.; Timmermann, D.B.; Ahring, P.K. pH Dependency and Desensitization Kinetics of Heterologously Expressed Combinations of Acid-sensing Ion Channel Subunits. J. Biol. Chem. 2004, 279, 11006–11015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzales, E.B.; Kawate, T.; Gouaux, E. Pore architecture and ion sites in acid-sensing ion channels and P2X receptors. Nature 2009, 460, 599–604. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Chen, Z.; Sigworth, F.J.; Canessa, C.M. Structure and analysis of nanobody binding to the human ASIC1a ion channel. eLife 2021, 10, e67115. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Bassler, E.L.; Ngo-Anh, T.J.; Geisler, H.S.; Ruppersberg, J.P.; Grunder, S. Molecular and functional characterization of acid-sensing ion channel (ASIC) 1b. J. Biol. Chem. 2001, 276, 33782–33787. [Google Scholar] [CrossRef] [Green Version]
- Hoagland, E.N.; Sherwood, T.W.; Lee, K.G.; Walker, C.J.; Askwith, C.C. Identification of a Calcium Permeable Human Acid-sensing Ion Channel 1 Transcript Variant. J. Biol. Chem. 2010, 285, 41852–41862. [Google Scholar] [CrossRef] [Green Version]
- Jacquot, F.; Khoury, S.; Labrum, B.; Delanoe, K.; Pidoux, L.; Barbier, J.; Delay, L.; Bayle, A.; Aissouni, Y.; Barriere, D.A.; et al. Lysophosphatidylcholine 16: 0 mediates chronic joint pain associated to rheumatic diseases through acid-sensing ion channel 3. Pain 2022, 163, 1999–2013. [Google Scholar] [CrossRef]
- Marra, S.; Ferru-Clément, R.; Breuil, V.; Delaunay, A.; Christin, M.; Friend, V.; Sebille, S.; Cognard, C.; Ferreira, T.; Roux, C.; et al. Non-acidic activation of pain-related Acid-Sensing Ion Channel 3 by lipids. EMBO J. 2016, 35, 414–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delaunay, A.; Gasull, X.; Salinas, M.; Noël, J.; Friend, V.; Lingueglia, E.; Deval, E. Human ASIC3 channel dynamically adapts its activity to sense the extracellular pH in both acidic and alkaline directions. Proc. Natl. Acad. Sci. USA 2012, 109, 13124–13129. [Google Scholar] [CrossRef] [Green Version]
- Deval, E.; Salinas, M.; Baron, A.; Lingueglia, E.; Lazdunski, M. ASIC2b-dependent Regulation of ASIC3, an Essential Acid-sensing Ion Channel Subunit in Sensory Neurons via the Partner Protein PICK-1. J. Biol. Chem. 2004, 279, 19531–19539. [Google Scholar] [CrossRef] [Green Version]
- Sivils, A.; Yang, F.; Wang, J.Q.; Chu, X.-P. Acid-Sensing Ion Channel 2: Function and Modulation. Membranes 2022, 12, 113. [Google Scholar] [CrossRef] [PubMed]
- Salinas, M.; Lazdunski, M.; Lingueglia, E. Structural Elements for the Generation of Sustained Currents by the Acid Pain Sensor ASIC3. J. Biol. Chem. 2009, 284, 31851–31859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartoi, T.; Augustinowski, K.; Polleichtner, G.; Gründer, S.; Ulbrich, M.H. Acid-sensing ion channel (ASIC) 1a/2a heteromers have a flexible 2:1/1:2 stoichiometry. Proc. Natl. Acad. Sci. USA 2014, 111, 8281–8286. [Google Scholar] [CrossRef] [Green Version]
- de Weille, J.R.; Bassilana, F.; Lazdunski, M.; Waldmann, R. Identification, functional expression and chromosomal localisation of a sustained human proton-gated cation channel. FEBS Lett. 1998, 433, 257–260. [Google Scholar] [CrossRef] [Green Version]
- Askwith, C.C.; Wemmie, J.A.; Price, M.P.; Rokhlina, T.; Welsh, M.J. Acid-sensing Ion Channel 2 (ASIC2) Modulates ASIC1 H+-activated Currents in Hippocampal Neurons. J. Biol. Chem. 2004, 279, 18296–18305. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Kalbacher, H.; Grunder, S. The tarantula toxin psalmotoxin 1 inhibits acid-sensing ion channel (ASIC) 1a by increasing its apparent H+ affinity. J. Gen. Physiol. 2005, 126, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Sherwood, T.W.; Askwith, C.C. Endogenous Arginine-Phenylalanine-Amide-related Peptides Alter Steady-state Desensitization of ASIC1a. J. Biol. Chem. 2008, 283, 1818–1830. [Google Scholar] [CrossRef]
- Cristofori-Armstrong, B.; Saez, N.J.; Chassagnon, I.R.; King, G.F.; Rash, L.D. The modulation of acid-sensing ion channel 1 by PcTx1 is pH-, subtype- and species-dependent: Importance of interactions at the channel subunit interface and potential for engineering selective analogues. Biochem. Pharmacol. 2019, 163, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Kalbacher, H.; Grunder, S. Interaction of acid-sensing ion channel (ASIC) 1 with the tarantula toxin psalmotoxin 1 is state dependent. J. Gen. Physiol. 2006, 127, 267–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diochot, S.; Baron, A.; Salinas, M.; Douguet, D.; Scarzello, S.; Dabert-Gay, A.-S.; Debayle, D.; Friend, V.; Alloui, A.; Lazdunski, M.; et al. Black mamba venom peptides target acid-sensing ion channels to abolish pain. Nature 2012, 490, 552–555. [Google Scholar] [CrossRef]
- Cristofori-Armstrong, B.; Budusan, E.; Rash, L.D. Mambalgin-3 potentiates human acid-sensing ion channel 1b under mild to moderate acidosis: Implications as an analgesic lead. Proc. Natl. Acad. Sci. USA 2021, 118, e2021581118. [Google Scholar] [CrossRef] [PubMed]
- Vaithia, A.; Vullo, S.; Peng, Z.; Alijevic, O.; Kellenberger, S. Accelerated Current Decay Kinetics of a Rare Human Acid-Sensing ion Channel 1a Variant That Is Used in Many Studies as Wild Type. Front. Mol. Neurosci. 2019, 12, 133. [Google Scholar] [CrossRef]
- Rook, M.L.; Miaro, M.; Couch, T.; Kneisley, D.L.; Musgaard, M.; MacLean, D.M. Mutation of a conserved glutamine residue does not abolish desensitization of acid-sensing ion channel 1. J. Gen. Physiol. 2021, 153, e202012855. [Google Scholar] [CrossRef]
- Baron, A.; Voilley, N.; Lazdunski, M.; Lingueglia, E. Acid sensing ion channels in dorsal spinal cord neurons. J. Neurosci. 2008, 28, 1498–1508. [Google Scholar] [CrossRef] [Green Version]
- Baron, A.; Schaefer, L.; Lingueglia, E.; Champigny, G.; Lazdunski, M. Zn2+ and H+ are coactivators of acid-sensing ion channels. J. Biol. Chem. 2001, 276, 35361–35367. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Kuenze, G.; Meiler, J.; Canessa, C.M. An arginine residue in the outer segment of hASIC1a TM1 affects both proton affinity and channel desensitization. J. Gen. Physiol. 2021, 153, e202012802. [Google Scholar] [CrossRef]
- Sherwood, T.W.; Lee, K.G.; Gormley, M.G.; Askwith, C.C. Heteromeric Acid-Sensing Ion Channels (ASICs) Composed of ASIC2b and ASIC1a Display Novel Channel Properties and Contribute to Acidosis-Induced Neuronal Death. J. Neurosci. 2011, 31, 9723–9734. [Google Scholar] [CrossRef]
- Bassilana, F.; Champigny, G.; Waldmann, R.; de Weille, J.R.; Heurteaux, C.; Lazdunski, M. The acid-sensitive ionic channel subunit ASIC and the mammalian degenerin MDEG form a heteromultimeric H+-gated Na+ channel with novel properties. J. Biol. Chem. 1997, 272, 28819–28822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hattori, T.; Chen, J.; Harding, A.M.S.; Price, M.P.; Lu, Y.; Abboud, F.M.; Benson, C.J. ASIC2a and ASIC3 Heteromultimerize to Form pH-Sensitive Channels in Mouse Cardiac Dorsal Root Ganglia Neurons. Circ. Res. 2009, 105, 279–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osmakov, D.I.; Koshelev, S.G.; Andreev, Y.A.; Kozlov, S.A. Endogenous Isoquinoline Alkaloids Agonists of Acid-Sensing Ion Channel Type 3. Front. Mol. Neurosci. 2017, 10, 282. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Paukert, M.; Kadurin, I.; Pusch, M.; Gründer, S. Strong modulation by RFamide neuropeptides of the ASIC1b/3 heteromer in competition with extracellular calcium. Neuropharmacology 2006, 50, 964–974. [Google Scholar] [CrossRef]
- Malysz, J.; Scott, V.E.; Faltynek, C.; Gopalakrishnan, M. Characterization of human ASIC2a homomeric channels stably expressed in murine Ltk− cells. Life Sci. 2008, 82, 30–40. [Google Scholar] [CrossRef]
- Cho, J.-H.; Askwith, C.C. Potentiation of acid-sensing ion channels by sulfhydryl compounds. Am. J. Physiol.-Cell. Physiol. 2007, 292, C2161–C2174. [Google Scholar] [CrossRef]
- Salinas, M.; Rash, L.D.; Baron, A.; Lambeau, G.; Escoubas, P.; Lazdunski, M. The receptor site of the spider toxin PcTx1 on the proton-gated cation channel ASIC1a. J. Physiol. 2006, 570, 339–354. [Google Scholar] [CrossRef]
- Sutherland, S.P.; Benson, C.J.; Adelman, J.P.; McCleskey, E.W. Acid-sensing ion channel 3 matches the acid-gated current in cardiac ischemia-sensing neurons. Proc. Natl. Acad. Sci. USA 2001, 98, 711–716. [Google Scholar] [CrossRef]
- Salinas, M.; Besson, T.; Delettre, Q.; Diochot, S.; Boulakirba, S.; Douguet, D.; Lingueglia, E. Binding Site and Inhibitory Mechanism of the Mambalgin-2 Pain-relieving Peptide on Acid-sensing Ion Channel 1a. J. Biol. Chem. 2014, 289, 13363–13373. [Google Scholar] [CrossRef] [Green Version]
- Yoder, N.; Yoshioka, C.; Gouaux, E. Gating mechanisms of acid-sensing ion channels. Nature 2018, 555, 397–401. [Google Scholar] [CrossRef]
- Baconguis, I.; Bohlen, C.J.; Goehring, A.; Julius, D.; Gouaux, E. X-Ray Structure of Acid-Sensing Ion Channel 1–Snake Toxin Complex Reveals Open State of a Na+-Selective Channel. Cell 2014, 156, 717–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, D.; Liu, S.; Li, S.; Zhang, M.; Yang, F.; Wen, M.; Shi, P.; Wang, T.; Pan, M.; Chang, S.; et al. Structural insights into human acid-sensing ion channel 1a inhibition by snake toxin mambalgin1. eLife 2020, 9, e57096. [Google Scholar] [CrossRef] [PubMed]
- Yoder, N.; Gouaux, E. Divalent cation and chloride ion sites of chicken acid sensing ion channel 1a elucidated by x-ray crystallography. PLoS ONE 2018, 13, e0202134. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Yang, Y.; Canessa, C.M. Interaction of the Aromatics Tyr-72/Trp-288 in the Interface of the Extracellular and Transmembrane Domains Is Essential for Proton Gating of Acid-sensing Ion Channels. J. Biol. Chem. 2009, 284, 4689–4694. [Google Scholar] [CrossRef] [Green Version]
- Yoder, N.; Gouaux, E. The His-Gly motif of acid-sensing ion channels resides in a reentrant ‘loop’ implicated in gating and ion selectivity. eLife 2020, 9, e56527. [Google Scholar] [CrossRef]
- Bargeton, B.; Kellenberger, S. The Contact Region between Three Domains of the Extracellular Loop of ASIC1a Is Critical for Channel Function. J. Biol. Chem. 2010, 285, 13816–13826. [Google Scholar] [CrossRef] [Green Version]
- Paukert, M.; Babini, E.; Pusch, M.; Gründer, S. Identification of the Ca2+ Blocking Site of Acid-sensing Ion Channel (ASIC) 1. J. Gen. Physiol. 2004, 124, 383–394. [Google Scholar] [CrossRef] [Green Version]
- Paukert, M.; Chen, X.; Polleichtner, G.; Schindelin, H.; Gründer, S. Candidate Amino Acids Involved in H+ Gating of Acid-sensing Ion Channel 1a. J. Biol. Chem. 2008, 283, 572–581. [Google Scholar] [CrossRef] [Green Version]
- Klipp, R.C.; Bankston, J.R. Structural determinants of acid-sensing ion channel potentiation by single chain lipids. J. Gen. Physiol. 2022, 154, e202213156. [Google Scholar] [CrossRef]
- Baconguis, I.; Gouaux, E. Structural plasticity and dynamic selectivity of acid-sensing ion channel–spider toxin complexes. Nature 2012, 489, 400–405. [Google Scholar] [CrossRef]
- Wu, Y.; Chen, Z.; Canessa, C.M. A valve-like mechanism controls desensitization of functional mammalian isoforms of acid-sensing ion channels. eLife 2019, 8, e45851. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Yang, Y.; Canessa, C.M. Asn415 in the β11-β12 Linker Decreases Proton-dependent Desensitization of ASIC1. J. Biol. Chem. 2010, 285, 31285–31291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rook, M.L.; Williamson, A.; Lueck, J.D.; Musgaard, M.; Maclean, D.M. β11-12 linker isomerization governs acid-sensing ion channel desensitization and recovery. eLife 2020, 9, e51111. [Google Scholar] [CrossRef] [PubMed]
- Rook, M.L.; Ananchenko, A.; Musgaard, M.; MacLean, D.M. Molecular Investigation of Chicken Acid-Sensing Ion Channel 1 β11-12 Linker Isomerization and Channel Kinetics. Front. Cell. Neurosci. 2021, 15, 761813. [Google Scholar] [CrossRef] [PubMed]
- Rook, M.L.; Musgaard, M.; MacLean, D.M. Coupling structure with function in acid-sensing ion channels: Challenges in pursuit of proton sensors. J. Physiol. 2020, 599, 417–430. [Google Scholar] [CrossRef] [PubMed]
- Cushman, K.A.; Marsh-Haffner, J.; Adelman, J.P.; McCleskey, E.W. A Conformation Change in the Extracellular Domain that Accompanies Desensitization of Acid-sensing Ion Channel (ASIC) 3. J. Gen. Physiol. 2007, 129, 345–350. [Google Scholar] [CrossRef] [Green Version]
- Della Vecchia, M.C.; Rued, A.C.; Carattino, M.D. Gating Transitions in the Palm Domain of ASIC1a*. J. Biol. Chem. 2013, 288, 5487–5495. [Google Scholar] [CrossRef] [Green Version]
- Krauson, A.J.; Carattino, M.D. The Thumb Domain Mediates Acid-sensing Ion Channel Desensitization. J. Biol. Chem. 2016, 291, 11407–11419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusama, N.; Gautam, M.; Harding, A.M.S.; Snyder, P.M.; Benson, C.J. Acid-sensing ion channels (ASICs) are differentially modulated by anions dependent on their subunit composition. Am. J. Physiol.-Cell. Physiol. 2013, 304, C89–C101. [Google Scholar] [CrossRef]
- Kusama, N.; Harding, A.M.S.; Benson, C.J. Extracellular Chloride Modulates the Desensitization Kinetics of Acid-sensing Ion Channel 1a (ASIC1a). J. Biol. Chem. 2010, 285, 17425–17431. [Google Scholar] [CrossRef]
- Ruan, Z.; Osei-Owusu, J.; Du, J.; Qiu, Z.; Lu, W. Structures and pH-sensing mechanism of the proton-activated chloride channel. Nature 2020, 588, 350–354. [Google Scholar] [CrossRef]
- Osei-Owusu, J.; Kots, E.; Ruan, Z.; Mihaljevic, L.; Chen, K.H.; Tamhaney, A.; Ye, X.; Lu, W.; Weinstein, H.; Qiu, Z. Molecular determinants of pH sensing in the proton-activated chloride channel. Proc. Natl. Acad. Sci. USA 2022, 119, e2200727119. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Li, P.; Wang, C.; Feng, X.; Geng, Q.; Chen, W.; Marthi, M.; Zhang, W.; Gao, C.; Reid, W.; et al. Parkinson’s disease-risk protein TMEM175 is a proton-activated proton channel in lysosomes. Cell 2022, 185, 2292–2308.e2220. [Google Scholar] [CrossRef] [PubMed]
- Karsan, N.; Gonzales, E.B.; Dussor, G. Targeted Acid-Sensing Ion Channel Therapies for Migraine. Neurotherapeutics 2018, 15, 402–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.H.; Chen, C.C. Roles of ASICs in Nociception and Proprioception. Adv. Exp. Med. Biol. 2018, 1099, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Deval, E.; Lingueglia, E. Acid-Sensing Ion Channels and nociception in the peripheral and central nervous systems. Neuropharmacology 2015, 94, 49–57. [Google Scholar] [CrossRef]
- Storozhuk, M.; Cherninskyi, A.; Maximyuk, O.; Isaev, D.; Krishtal, O. Acid-Sensing Ion Channels: Focus on Physiological and Some Pathological Roles in the Brain. Curr. Neuropharmacol. 2021, 19, 1570–1589. [Google Scholar] [CrossRef]
- Heusser, S.A.; Pless, S.A. Acid-sensing ion channels as potential therapeutic targets. Trends Pharmacol. Sci. 2021, 42, 1035–1050. [Google Scholar] [CrossRef]
- Lin, S.-H.; Sun, W.-H.; Chen, C.-C. Genetic exploration of the role of acid-sensing ion channels. Neuropharmacology 2015, 94, 99–118. [Google Scholar] [CrossRef]
- Lin, J.-H.; Hung, C.-H.; Han, D.-S.; Chen, S.-T.; Lee, C.-H.; Sun, W.-Z.; Chen, C.-C. Sensing acidosis: Nociception or sngception? J. Biomed. Sci. 2018, 25, 85. [Google Scholar] [CrossRef]
- Dulai, J.S.; Smith, E.S.J.; Rahman, T. Acid-sensing ion channel 3: An analgesic target. Channels 2021, 15, 94–127. [Google Scholar] [CrossRef] [PubMed]
- Ritzel, R.M.; He, J.; Li, Y.; Cao, T.; Khan, N.; Shim, B.; Sabirzhanov, B.; Aubrecht, T.; Stoica, B.A.; Faden, A.I.; et al. Proton extrusion during oxidative burst in microglia exacerbates pathological acidosis following traumatic brain injury. Glia 2020, 69, 746–764. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.-H.; Jones, S.; Talman, W.T. Cellular Localization of Acid-Sensing Ion Channel 1 in Rat Nucleus Tractus Solitarii. Cell. Mol. Neurobiol. 2017, 38, 219–232. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.N.; Stein, C.; Gaveriaux-Ruff, C. Opioids and Pain. In The Oxford Handbook of the Neurobiology of Pain; Oxford University Press: Oxford, UK, 2020; pp. 727–769. [Google Scholar]
- Chesler, M. Regulation and Modulation of pH in the Brain. Physiol. Rev. 2003, 83, 1183–1221. [Google Scholar] [CrossRef]
- Ugawa, S.; Ueda, T.; Ishida, Y.; Nishigaki, M.; Shibata, Y.; Shimada, S. Amiloride-blockable acid-sensing ion channels are leading acid sensors expressed in human nociceptors. J. Clin. Invest. 2002, 110, 1185–1190. [Google Scholar] [CrossRef] [PubMed]
- Jones, N.G.; Slater, R.; Cadiou, H.; McNaughton, P.; McMahon, S.B. Acid-induced pain and its modulation in humans. J. Neurosci. 2004, 24, 10974–10979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, N.; Tribble, J.; Peterson, A.M.; Jiang, Q.; Wang, J.Q.; Chu, X.-P. Acid-Sensing Ion Channels and Mechanosensation. Int. J. Mol. Sci. 2021, 22, 4810. [Google Scholar] [CrossRef]
- Wang, Y.; O’Bryant, Z.; Wang, H.; Huang, Y. Regulating Factors in Acid-Sensing Ion Channel 1a Function. Neurochem. Res. 2015, 41, 631–645. [Google Scholar] [CrossRef] [PubMed]
- Cullinan, M.M.; Klipp, R.C.; Bankston, J.R. Regulation of acid-sensing ion channels by protein binding partners. Channels 2021, 15, 635–647. [Google Scholar] [CrossRef]
- Deval, E.; Noël, J.; Lay, N.; Alloui, A.; Diochot, S.; Friend, V.; Jodar, M.; Lazdunski, M.; Lingueglia, E. ASIC3, a sensor of acidic and primary inflammatory pain. EMBO J. 2008, 27, 3047–3055. [Google Scholar] [CrossRef]
- Du, J.; Reznikov, L.R.; Price, M.P.; Zha, X.-m.; Lu, Y.; Moninger, T.O.; Wemmie, J.A.; Welsh, M.J. Protons are a neurotransmitter that regulates synaptic plasticity in the lateral amygdala. Proc. Natl. Acad. Sci. USA 2014, 111, 8961–8966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Jiang, N.; Li, J.; Ji, Y.-H.; Xiong, Z.-G.; Zha, X.-m. Two aspects of ASIC function: Synaptic plasticity and neuronal injury. Neuropharmacology 2015, 94, 42–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baron, A.; Lingueglia, E. Pharmacology of acid-sensing ion channels—Physiological and therapeutical perspectives. Neuropharmacology 2015, 94, 19–35. [Google Scholar] [CrossRef]
- Rash, L.D. Acid-Sensing Ion Channel Pharmacology, Past, Present, and Future. Adv. Pharm. 2017, 79, 35–66. [Google Scholar] [CrossRef]
- Escoubas, P.; De Weille, J.R.; Lecoq, A.; Diochot, S.; Waldmann, R.; Champigny, G.; Moinier, D.; Menez, A.; Lazdunski, M. Isolation of a tarantula toxin specific for a class of proton-gated Na+ channels. J. Biol. Chem. 2000, 275, 25116–25121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diochot, S.; Baron, A.; Rash, L.D.; Deval, E.; Escoubas, P.; Scarzello, S.; Salinas, M.; Lazdunski, M. A new sea anemone peptide, APETx2, inhibits ASIC3, a major acid-sensitive channel in sensory neurons. EMBO J. 2004, 23, 1516–1525. [Google Scholar] [CrossRef] [PubMed]
- Baron, A.; Diochot, S.; Salinas, M.; Deval, E.; Noël, J.; Lingueglia, E. Venom toxins in the exploration of molecular, physiological and pathophysiological functions of acid-sensing ion channels. Toxicon 2013, 75, 187–204. [Google Scholar] [CrossRef]
- Bohlen, C.J.; Chesler, A.T.; Sharif-Naeini, R.; Medzihradszky, K.F.; Zhou, S.; King, D.; Sánchez, E.E.; Burlingame, A.L.; Basbaum, A.I.; Julius, D. A heteromeric Texas coral snake toxin targets acid-sensing ion channels to produce pain. Nature 2011, 479, 410–414. [Google Scholar] [CrossRef] [Green Version]
- Saez, N.J.; Mobli, M.; Bieri, M.; Chassagnon, I.R.; Malde, A.K.; Gamsjaeger, R.; Mark, A.E.; Gooley, P.R.; Rash, L.D.; King, G.F. A Dynamic Pharmacophore Drives the Interaction between Psalmotoxin-1 and the Putative Drug Target Acid-Sensing Ion Channel 1a. Mol. Pharmacol. 2011, 80, 796–808. [Google Scholar] [CrossRef] [Green Version]
- Saez, N.J.; Deplazes, E.; Cristofori-Armstrong, B.; Chassagnon, I.R.; Lin, X.; Mobli, M.; Mark, A.E.; Rash, L.D.; King, G.F. Molecular dynamics and functional studies define a hot spot of crystal contacts essential for PcTx1 inhibition of acid-sensing ion channel 1a. Br. J. Pharmacol. 2015, 172, 4985–4995. [Google Scholar] [CrossRef]
- Dawson, R.J.P.; Benz, J.; Stohler, P.; Tetaz, T.; Joseph, C.; Huber, S.; Schmid, G.; Hügin, D.; Pflimlin, P.; Trube, G.; et al. Structure of the Acid-sensing ion channel 1 in complex with the gating modifier Psalmotoxin 1. Nat. Commun. 2012, 3, 936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borg, C.B.; Heusser, S.A.; Colding, J.M.; Pless, S.A. Conformational decoupling in acid-sensing ion channels uncovers mechanism and stoichiometry of PcTx1-mediated inhibition. eLife 2022, 11, e73384. [Google Scholar] [CrossRef]
- Bohlen, C.J.; Julius, D. Receptor-targeting mechanisms of pain-causing toxins: How ow? Toxicon 2012, 60, 254–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mourier, G.; Salinas, M.; Kessler, P.; Stura, E.A.; Leblanc, M.; Tepshi, L.; Besson, T.; Diochot, S.; Baron, A.; Douguet, D.; et al. Mambalgin-1 Pain-relieving Peptide, Stepwise Solid-phase Synthesis, Crystal Structure, and Functional Domain for Acid-sensing Ion Channel 1a Inhibition. J. Biol. Chem. 2016, 291, 2616–2629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salinas, M.; Kessler, P.; Douguet, D.; Sarraf, D.; Tonali, N.; Thai, R.; Servent, D.; Lingueglia, E. Mambalgin-1 pain-relieving peptide locks the hinge between α4 and α5 helices to inhibit rat acid-sensing ion channel 1a. Neuropharmacology 2021, 185, 108453. [Google Scholar] [CrossRef]
- Schroeder, C.I.; Rash, L.D.; Vila-Farrés, X.; Rosengren, K.J.; Mobli, M.; King, G.F.; Alewood, P.F.; Craik, D.J.; Durek, T. Chemical Synthesis, 3D Structure, and ASIC Binding Site of the Toxin Mambalgin-2. Angew. Chem. Int. Ed. 2014, 53, 1017–1020. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.; Yu, Y.; Xue, X.; Pan, M.; Wen, M.; Li, S.; Qu, Q.; Li, X.; Zhang, L.; Li, X.; et al. Cryo-EM structure of the ASIC1a–mambalgin-1 complex reveals that the peptide toxin mambalgin-1 inhibits acid-sensing ion channels through an unusual allosteric effect. Cell Discov. 2018, 4, 27. [Google Scholar] [CrossRef] [Green Version]
- Jensen, J.E.; Durek, T.; Alewood, P.F.; Adams, D.J.; King, G.F.; Rash, L.D. Chemical synthesis and folding of APETx2, a potent and selective inhibitor of acid sensing ion channel 3. Toxicon 2009, 54, 56–61. [Google Scholar] [CrossRef]
- Anangi, R.; Chen, C.-C.; Lin, Y.-W.; Cheng, Y.-R.; Cheng, C.-H.; Chen, Y.-C.; Chu, Y.-P.; Chuang, W.-J. Expression in Pichia pastoris and characterization of APETx2, a specific inhibitor of acid sensing ion channel 3. Toxicon 2010, 56, 1388–1397. [Google Scholar] [CrossRef]
- Qadri, Y.J.; Berdiev, B.K.; Song, Y.; Lippton, H.L.; Fuller, C.M.; Benos, D.J. Psalmotoxin-1 Docking to Human Acid-sensing Ion Channel-1. J. Biol. Chem. 2009, 284, 17625–17633. [Google Scholar] [CrossRef]
- Samways, D.S.K.; Harkins, A.B.; Egan, T.M. Native and recombinant ASIC1a receptors conduct negligible Ca2+ entry. Cell Calcium 2009, 45, 319–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, C.-x.; Wu, F.-r.; Wang, X.-y.; Tang, J.; Gao, W.-f.; Ge, J.-f.; Chen, F.-h. Inhibition of ASICs reduces rat hepatic stellate cells activity and liver fibrosis: An in vitro and in vivo study. Cell Biol. Int. 2014, 38, 1003–1012. [Google Scholar] [CrossRef] [PubMed]
- Joeres, N.; Augustinowski, K.; Neuhof, A.; Assmann, M.; Gründer, S. Functional and pharmacological characterization of two different ASIC1a/2a heteromers reveals their sensitivity to the spider toxin PcTx1. Sci. Rep. 2016, 6, 27647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Hagan, R.; Schoellerman, J. Dual actions of Psalmotoxin at ASIC1a and ASIC2a heteromeric channels (ASIC1a/2a). Sci. Rep. 2018, 8, 7179. [Google Scholar] [CrossRef] [Green Version]
- Gregory, N.S.; Gautam, M.; Benson, C.J.; Sluka, K.A. Acid Sensing Ion Channel 1a (ASIC1a) Mediates Activity-induced Pain by Modulation of Heteromeric ASIC Channel Kinetics. Neuroscience 2018, 386, 166–174. [Google Scholar] [CrossRef]
- Lee, J.Y.P.; Saez, N.J.; Cristofori-Armstrong, B.; Anangi, R.; King, G.F.; Smith, M.T.; Rash, L.D. Inhibition of acid-sensing ion channels by diminazene and APETx2 evoke partial and highly variable antihyperalgesia in a rat model of inflammatory pain. Br. J. Pharmacol. 2018, 175, 2204–2218. [Google Scholar] [CrossRef]
- Andreev, Y.; Osmakov, D.; Koshelev, S.; Maleeva, E.; Logashina, Y.; Palikov, V.; Palikova, Y.; Dyachenko, I.; Kozlov, S. Analgesic Activity of Acid-Sensing Ion Channel 3 (ASIC3) Inhibitors: Sea Anemones Peptides Ugr9-1 and APETx2 versus Low Molecular Weight Compounds. Mar. Drugs 2018, 16, 500. [Google Scholar] [CrossRef] [Green Version]
- Escoubas, P.; Bernard, C.; Lambeau, G.; Lazdunski, M.; Darbon, H. Recombinant production and solution structure of PcTx1, the specific peptide inhibitor of ASIC1a proton-gated cation channels. Protein Sci. 2003, 12, 1332–1343. [Google Scholar] [CrossRef] [Green Version]
- Verkest, C.; Diochot, S.; Lingueglia, E.; Baron, A. C-Jun N-Terminal Kinase Post-Translational Regulation of Pain-Related Acid-Sensing Ion Channels 1b and 3. J. Neurosci. 2021, 41, 8673–8685. [Google Scholar] [CrossRef]
- Baron, A.; Waldmann, R.; Lazdunski, M. ASIC-like, proton-activated currents in rat hippocampal neurons. J. Physiol. 2002, 539, 485–494. [Google Scholar] [CrossRef]
- Duan, B.; Wu, L.J.; Yu, Y.Q.; Ding, Y.; Jing, L.; Xu, L.; Chen, J.; Xu, T.L. Upregulation of acid-sensing ion channel ASIC1a in spinal dorsal horn neurons contributes to inflammatory pain hypersensitivity. J. Neurosci. 2007, 27, 11139–11148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzuca, M.; Heurteaux, C.; Alloui, A.; Diochot, S.; Baron, A.; Voilley, N.; Blondeau, N.; Escoubas, P.; Gelot, A.; Cupo, A.; et al. A tarantula peptide against pain via ASIC1a channels and opioid mechanisms. Nat. Neurosci. 2007, 10, 943–945. [Google Scholar] [CrossRef] [PubMed]
- Er, S.Y.; Cristofori-Armstrong, B.; Escoubas, P.; Rash, L.D. Discovery and molecular interaction studies of a highly stable, tarantula peptide modulator of acid-sensing ion channel 1. Neuropharmacology 2017, 127, 185–195. [Google Scholar] [CrossRef] [Green Version]
- Chassagnon, I.R.; McCarthy, C.A.; Chin, Y.K.Y.; Pineda, S.S.; Keramidas, A.; Mobli, M.; Pham, V.; De Silva, T.M.; Lynch, J.W.; Widdop, R.E.; et al. Potent neuroprotection after stroke afforded by a double-knot spider-venom peptide that inhibits acid-sensing ion channel 1a. Proc. Natl. Acad. Sci. USA 2017, 114, 3750–3755. [Google Scholar] [CrossRef] [Green Version]
- Duggan, N.M.; Saez, N.J.; Clayton, D.; Budusan, E.; Watson, E.E.; Tucker, I.J.; Rash, L.D.; King, G.F.; Payne, R.J. Total Synthesis of the Spider-Venom Peptide Hi1a. Org. Lett. 2021, 23, 8375–8379. [Google Scholar] [CrossRef] [PubMed]
- Redd, M.A.; Scheuer, S.E.; Saez, N.J.; Yoshikawa, Y.; Chiu, H.S.; Gao, L.; Hicks, M.; Villanueva, J.E.; Joshi, Y.; Chow, C.Y.; et al. Therapeutic Inhibition of Acid-Sensing Ion Channel 1a Recovers Heart Function After Ischemia–Reperfusion Injury. Circulation 2021, 144, 947–960. [Google Scholar] [CrossRef]
- Cristofori-Armstrong, B. Discovery and modulation of acid-sensing ion channel modulating venom peptides. Ph.D. Thesis, The University of Queensland, Brisbane, Australia, 29 March 2019. [Google Scholar]
- Rahman, T.; Smith, E.S.J. In silico assessment of interaction of sea anemone toxin APETx2 and acid sensing ion channel 3. Biochem. Biophys. Res. Commun. 2014, 450, 384–389. [Google Scholar] [CrossRef] [Green Version]
- Gründer, S.; Augustinowski, K. Toxin binding reveals two open state structures for one acid-sensing ion channel. Channels 2014, 6, 409–413. [Google Scholar] [CrossRef] [Green Version]
- Besson, T.; Lingueglia, E.; Salinas, M. Pharmacological modulation of Acid-Sensing Ion Channels 1a and 3 by amiloride and 2-guanidine-4-methylquinazoline (GMQ). Neuropharmacology 2017, 125, 429–440. [Google Scholar] [CrossRef] [PubMed]
- Buta, A.; Maximyuk, O.; Kovalskyy, D.; Sukach, V.; Vovk, M.; Ievglevskyi, O.; Isaeva, E.; Isaev, D.; Savotchenko, A.; Krishtal, O. Novel Potent Orthosteric Antagonist of ASIC1a Prevents NMDAR-Dependent LTP Induction. J. Med. Chem. 2015, 58, 4449–4461. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Lu, J.-F.; Huang, Z.-Y.; Liu, Y.-J.; Cai, L.-B.; Wen, X.-L.; Song, X.-L.; Xiong, J.; Sun, P.-Y.; Zhang, H.; et al. Pharmacological Validation of ASIC1a as a Druggable Target for Neuroprotection in Cerebral Ischemia Using an Intravenously Available Small Molecule Inhibitor. Front. Pharmacol. 2022, 13, 849498. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Boiteux, C.; Alijevic, O.; Liang, C.; Bernèche, S.; Kellenberger, S. Molecular determinants of desensitization in an ENaC/degenerin channel. FASEB J. 2013, 27, 5034–5045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Springauf, A.; Bresenitz, P.; Gründer, S. The Interaction between Two Extracellular Linker Regions Controls Sustained Opening of Acid-sensing Ion Channel 1. J. Biol. Chem. 2011, 286, 24374–24384. [Google Scholar] [CrossRef] [Green Version]
- Aldrich, R.W.; Yang, H.; Yu, Y.; Li, W.-G.; Yu, F.; Cao, H.; Xu, T.-L.; Jiang, H. Inherent Dynamics of the Acid-Sensing Ion Channel 1 Correlates with the Gating Mechanism. PLoS Biol. 2009, 7, e1000151. [Google Scholar] [CrossRef] [Green Version]
- Swain, S.M.; Bera, A.K. Coupling of Proton Binding in Extracellular Domain to Channel Gating in Acid-Sensing Ion Channel. J. Mol. Neurosci. 2013, 51, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Yang, Y.; Canessa, C.M. Leu85 in the β1-β2 Linker of ASIC1 Slows Activation and Decreases the Apparent Proton Affinity by Stabilizing a Closed Conformation. J. Biol. Chem. 2010, 285, 22706–22712. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Yang, Y.; Canessa, C.M. Two residues in the extracellular domain convert a nonfunctional ASIC1 into a proton-activated channel. Am. J. Physiol.-Cell. Physiol. 2010, 299, C66–C73. [Google Scholar] [CrossRef]
- Salinas Castellanos, L.C.; Uchitel, O.D.; Weissmann, C. Signaling Pathways in Proton and Non-proton ASIC1a Activation. Front. Cell. Neurosci. 2021, 15, 735414. [Google Scholar] [CrossRef]
- Baron, A.; Deval, E.; Salinas, M.; Lingueglia, E.; Voilley, N.; Lazdunski, M. Protein kinase C stimulates the acid-sensing ion channel ASIC2a via the PDZ domain-containing protein PICK1. J. Biol. Chem. 2002, 277, 50463–50468. [Google Scholar] [CrossRef] [Green Version]
- Diochot, S.; Alloui, A.; Rodrigues, P.; Dauvois, M.; Friend, V.; Aissouni, Y.; Eschalier, A.; Lingueglia, E.; Baron, A. Analgesic effects of mambalgin peptide inhibitors of acid-sensing ion channels in inflammatory and neuropathic pain. Pain 2016, 157, 552–559. [Google Scholar] [CrossRef]
- Chang, C.-T.; Fong, S.W.; Lee, C.-H.; Chuang, Y.-C.; Lin, S.-H.; Chen, C.-C. Involvement of Acid-Sensing Ion Channel 1b in the Development of Acid-Induced Chronic Muscle Pain. Front. Neurosci. 2019, 13, 1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, G.T.; Gutteridge, A.; Fox, H.D.E.; Wilbrey, A.L.; Cao, L.; Cho, L.T.; Brown, A.R.; Benn, C.L.; Kammonen, L.R.; Friedman, J.H.; et al. Characterizing Human Stem Cell–derived Sensory Neurons at the Single-cell Level Reveals Their Ion Channel Expression and Utility in Pain Research. Mol. Ther. 2014, 22, 1530–1543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, Y.C.; Park, S.S.; Subieta, A.R.; Brennan, T.J. Changes in tissue pH and temperature after incision indicate acidosis may contribute to postoperative pain. Anesthesiology 2004, 101, 468–475. [Google Scholar] [CrossRef]
- Verkest, C.; Piquet, E.; Diochot, S.; Dauvois, M.; Lanteri-Minet, M.; Lingueglia, E.; Baron, A. Effects of systemic inhibitors of acid-sensing ion channels 1 (ASIC1) against acute and chronic mechanical allodynia in a rodent model of migraine. Br. J. Pharmacol. 2018, 175, 4154–4166. [Google Scholar] [CrossRef] [Green Version]
- Chagot, B.; Diochot, S.; Pimentel, C.; Lazdunski, M.; Darbon, H. Solution structure of APETx1 from the sea anemone Anthopleura elegantissima: A new fold for an HERG toxin. Proteins 2005, 59, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Deval, E.; Noel, J.; Gasull, X.; Delaunay, A.; Alloui, A.; Friend, V.; Eschalier, A.; Lazdunski, M.; Lingueglia, E. Acid-Sensing Ion Channels in Postoperative Pain. J. Neurosci. 2011, 31, 6059–6066. [Google Scholar] [CrossRef] [Green Version]
- Karczewski, J.; Spencer, R.H.; Garsky, V.M.; Liang, A.; Leitl, M.D.; Cato, M.J.; Cook, S.P.; Kane, S.; Urban, M.O. Reversal of acid-induced and inflammatory pain by the selective ASIC3 inhibitor, APETx2. Br. J. Pharmacol. 2010, 161, 950–960. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, H.T.J.; Hodson, N.; Baird, P.; Richardson, S.M.; Hoyland, J.A. Acidic pH promotes intervertebral disc degeneration: Acid-sensing ion channel -3 as a potential therapeutic target. Sci. Rep. 2016, 6, 37360. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.Y.; Yan, W.W.; Zhang, X.L.; Liu, H.; Zhang, L.C. ASIC3 in the cerebrospinal fluid-contacting nucleus of brain parenchyma contributes to inflammatory pain in rats. Neurol. Res. 2013, 36, 270–275. [Google Scholar] [CrossRef]
- Martínez-Rojas, V.A.; Barragán-Iglesias, P.; Rocha-González, H.I.; Murbartián, J.; Granados-Soto, V. Role of TRPV1 and ASIC3 in formalin-induced secondary allodynia and hyperalgesia. Pharmacol. Rep. 2014, 66, 964–971. [Google Scholar] [CrossRef]
- Yan, J.; Wei, X.; Bischoff, C.; Edelmayer, R.M.; Dussor, G. pH-Evoked Dural Afferent Signaling Is Mediated by ASIC3 and Is Sensitized by Mast Cell Mediators. Headache J. Head Face Pain 2013, 53, 1250–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izumi, M.; Ikeuchi, M.; Ji, Q.; Tani, T. Local ASIC3 modulates pain and disease progression in a rat model of osteoarthritis. J. Biomed. Sci. 2012, 19, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugimura, N.; Ikeuchi, M.; Izumi, M.; Kawano, T.; Aso, K.; Kato, T.; Ushida, T.; Yokoyama, M.; Tani, T. Repeated intra-articular injections of acidic saline produce long-lasting joint pain and widespread hyperalgesia. Eur. J. Pain 2015, 19, 629–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callejo, G.; Castellanos, A.; Castany, M.; Gual, A.; Luna, C.; Acosta, M.C.; Gallar, J.; Giblin, J.P.; Gasull, X. Acid-sensing ion channels detect moderate acidifications to induce ocular pain. Pain 2015, 156, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.-N.; Lee, C.-H.; Lin, S.-H.; Wong, C.-W.; Sun, W.-H.; Wood, J.N.; Chen, C.-C. Roles of ASIC3, TRPV1, and NaV1.8 in the Transition from Acute to Chronic Pain in a Mouse Model of Fibromyalgia. Mol. Pain 2014, 10, 40. [Google Scholar] [CrossRef] [PubMed]
- Gregory, J.; McGowan, L. An examination of the prevalence of acute pain for hospitalised adult patients: A systematic review. J. Clin. Nurs. 2016, 25, 583–598. [Google Scholar] [CrossRef]
- Hiasa, M.; Okui, T.; Allette, Y.M.; Ripsch, M.S.; Sun-Wada, G.-H.; Wakabayashi, H.; Roodman, G.D.; White, F.A.; Yoneda, T. Bone Pain Induced by Multiple Myeloma Is Reduced by Targeting V-ATPase and ASIC3. Cancer Res. 2017, 77, 1283–1295. [Google Scholar] [CrossRef] [Green Version]
- Hanaka, M.; Iba, K.; Dohke, T.; Kanaya, K.; Okazaki, S.; Yamashita, T. Antagonists to TRPV1, ASICs and P2X have a potential role to prevent the triggering of regional bone metabolic disorder and pain-like behavior in tail-suspended mice. Bone 2018, 110, 284–294. [Google Scholar] [CrossRef]
- Kanaya, K.; Iba, K.; Abe, Y.; Dohke, T.; Okazaki, S.; Matsumura, T.; Yamashita, T. Acid-sensing ion channel 3 or P2X2/3 is involved in the pain-like behavior under a high bone turnover state in ovariectomized mice. J. Orthop. Res. 2016, 34, 566–573. [Google Scholar] [CrossRef]
- Blanchard, M.G.; Rash, L.D.; Kellenberger, S. Inhibition of voltage-gated Na+ currents in sensory neurones by the sea anemone toxin APETx2. Br. J. Pharmacol. 2012, 165, 2167–2177. [Google Scholar] [CrossRef]
- Peigneur, S.; Béress, L.; Möller, C.; Marí, F.; Forssmann, W.G.; Tytgat, J. A natural point mutation changes both target selectivity and mechanism of action of sea anemone toxins. FASEB J. 2012, 26, 5141–5151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, J.E.; Cristofori-Armstrong, B.; Anangi, R.; Rosengren, K.J.; Lau, C.H.Y.; Mobli, M.; Brust, A.; Alewood, P.F.; King, G.F.; Rash, L.D. Understanding the Molecular Basis of Toxin Promiscuity: The Analgesic Sea Anemone Peptide APETx2 Interacts with Acid-Sensing Ion Channel 3 and hERG Channels via Overlapping Pharmacophores. J. Med. Chem. 2014, 57, 9195–9203. [Google Scholar] [CrossRef]
- Kalina, R.; Gladkikh, I.; Dmitrenok, P.; Chernikov, O.; Koshelev, S.; Kvetkina, A.; Kozlov, S.; Kozlovskaya, E.; Monastyrnaya, M. New APETx-like peptides from sea anemone Heteractis crispa modulate ASIC1a channels. Peptides 2018, 104, 41–49. [Google Scholar] [CrossRef]
- Kalina, R.S.; Koshelev, S.G.; Zelepuga, E.A.; Kim, N.Y.; Kozlov, S.A.; Kozlovskaya, E.P.; Monastyrnaya, M.M.; Gladkikh, I.N. APETx-Like Peptides from the Sea Anemone Heteractis crispa, Diverse in Their Effect on ASIC1a and ASIC3 Ion Channels. Toxins 2020, 12, 266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinheiro-Junior, E.L.; Kalina, R.; Gladkikh, I.; Leychenko, E.; Tytgat, J.; Peigneur, S. A Tale of Toxin Promiscuity: The Versatile Pharmacological Effects of Hcr 1b-2 Sea Anemone Peptide on Voltage-Gated Ion Channels. Mar. Drugs 2022, 20, 147. [Google Scholar] [CrossRef]
- Osmakov, D.I.; Kozlov, S.A.; Andreev, Y.A.; Koshelev, S.G.; Sanamyan, N.P.; Sanamyan, K.E.; Dyachenko, I.A.; Bondarenko, D.A.; Murashev, A.N.; Mineev, K.S.; et al. Sea Anemone Peptide with Uncommon β-Hairpin Structure Inhibits Acid-sensing Ion Channel 3 (ASIC3) and Reveals Analgesic Activity. J. Biol. Chem. 2013, 288, 23116–23127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez, A.A.; Salceda, E.; Garateix, A.G.; Zaharenko, A.J.; Peigneur, S.; López, O.; Pons, T.; Richardson, M.; Díaz, M.; Hernández, Y.; et al. A novel sea anemone peptide that inhibits acid-sensing ion channels. Peptides 2014, 53, 3–12. [Google Scholar] [CrossRef]
- Reimers, C.; Lee, C.-H.; Kalbacher, H.; Tian, Y.; Hung, C.-H.; Schmidt, A.; Prokop, L.; Kauferstein, S.; Mebs, D.; Chen, C.-C.; et al. Identification of a cono-RFamide from the venom of Conus textile that targets ASIC3 and enhances muscle pain. Proc. Natl. Acad. Sci. USA 2017, 114, E3507–E3515. [Google Scholar] [CrossRef] [Green Version]
- Báez, A.; Salceda, E.; Fló, M.; Graña, M.; Fernández, C.; Vega, R.; Soto, E. α-Dendrotoxin inhibits the ASIC current in dorsal root ganglion neurons from rat. Neurosci. Lett. 2015, 606, 42–47. [Google Scholar] [CrossRef]
- Skarżyński, T. Crystal structure of α-dendrotoxin from the green mamba venom and its comparison with the structure of bovine pancreatic trypsin inhibitor. J. Mol. Biol. 1992, 224, 671–683. [Google Scholar] [CrossRef]
- Cristofori-Armstrong, B.; Rash, L.D. Acid-sensing ion channel (ASIC) structure and function: Insights from spider, snake and sea anemone venoms. Neuropharmacology 2017, 127, 173–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández, C.; Konno, K.; Salceda, E.; Vega, R.; Zaharenko, A.J.; Soto, E. Sa12b Peptide from Solitary Wasp Inhibits ASIC Currents in Rat Dorsal Root Ganglion Neurons. Toxins 2019, 11, 585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, P.; Torck, A.; Quigley, L.; Wangzhou, A.; Neiman, M.; Rao, C.; Lam, T.; Kim, J.-Y.; Kim, T.H.; Zhang, M.Q.; et al. Comparative transcriptome profiling of the human and mouse dorsal root ganglia: An RNA-seq–based resource for pain and sensory neuroscience research. Pain 2018, 159, 1325–1345. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.C.; Zimmer, A.; Sun, W.H.; Hall, J.; Brownstein, M.J. A role for ASIC3 in the modulation of high-intensity pain stimuli. Proc. Natl. Acad. Sci. USA 2002, 99, 8992–8997. [Google Scholar] [CrossRef] [Green Version]
- Price, M.P.; McIlwrath, S.L.; Xie, J.; Cheng, C.; Qiao, J.; Tarr, D.E.; Sluka, K.A.; Brennan, T.J.; Lewin, G.R.; Welsh, M.J. The DRASIC cation channel contributes to the detection of cutaneous touch and acid stimuli in mice. Neuron 2001, 32, 1071–1083. [Google Scholar] [CrossRef] [Green Version]
- Fu, H.; Fang, P.; Zhou, H.-Y.; Zhou, J.; Yu, X.-W.; Ni, M.; Zheng, J.-Y.; Jin, Y.; Chen, J.-G.; Wang, F.; et al. Acid-sensing ion channels in trigeminal ganglion neurons innervating the orofacial region contribute to orofacial inflammatory pain. Clin. Exp. Pharmacol. Physiol. 2016, 43, 193–202. [Google Scholar] [CrossRef]
- Nakamura, M.; Jang, I.-S. Characterization of proton-induced currents in rat trigeminal mesencephalic nucleus neurons. Brain Res. 2014, 1583, 12–22. [Google Scholar] [CrossRef]
- Lu, Y.; Ma, X.; Sabharwal, R.; Snitsarev, V.; Morgan, D.; Rahmouni, K.; Drummond, H.A.; Whiteis, C.A.; Costa, V.; Price, M.; et al. The Ion Channel ASIC2 Is Required for Baroreceptor and Autonomic Control of the Circulation. Neuron 2009, 64, 885–897. [Google Scholar] [CrossRef] [Green Version]
- Dusenkova, S.; Ru, F.; Surdenikova, L.; Nassenstein, C.; Hatok, J.; Dusenka, R.; Banovcin, P.; Kliment, J.; Tatar, M.; Kollarik, M. The expression profile of acid-sensing ion channel (ASIC) subunits ASIC1a, ASIC1b, ASIC2a, ASIC2b, and ASIC3 in the esophageal vagal afferent nerve subtypes. Am. J. Physiol. -Gastrointest. Liver Physiol. 2014, 307, G922–G930. [Google Scholar] [CrossRef]
- Kupari, J.; Häring, M.; Agirre, E.; Castelo-Branco, G.; Ernfors, P. An Atlas of Vagal Sensory Neurons and Their Molecular Specialization. Cell Rep. 2019, 27, 2508–2523.e2504. [Google Scholar] [CrossRef]
- Wu, W.L.; Lin, Y.W.; Min, M.Y.; Chen, C.C. Mice lacking Asic3 show reduced anxiety-like behavior on the elevated plus maze and reduced aggression. Genes Brain Behav. 2010, 9, 603–614. [Google Scholar] [CrossRef] [PubMed]
- Zeisel, A.; Hochgerner, H.; Lönnerberg, P.; Johnsson, A.; Memic, F.; van der Zwan, J.; Häring, M.; Braun, E.; Borm, L.E.; La Manno, G.; et al. Molecular Architecture of the Mouse Nervous System. Cell 2018, 174, 999–1014.e1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Usoskin, D.; Furlan, A.; Islam, S.; Abdo, H.; Lönnerberg, P.; Lou, D.; Hjerling-Leffler, J.; Haeggström, J.; Kharchenko, O.; Kharchenko, P.V.; et al. Unbiased classification of sensory neuron types by large-scale single-cell RNA sequencing. Nat. Neurosci. 2014, 18, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Molliver, D.C.; Immke, D.C.; Fierro, L.; Pare, M.; Rice, F.L.; McCleskey, E.W. ASIC3, an acid-sensing ion channel, is expressed in metaboreceptive sensory neurons. Mol. Pain 2005, 1, 35. [Google Scholar] [CrossRef]
- Nguyen, M.Q.; Le Pichon, C.E.; Ryba, N. Stereotyped transcriptomic transformation of somatosensory neurons in response to injury. eLife 2019, 8, e49679. [Google Scholar] [CrossRef]
- Obukhov, A.G.; Nguyen, M.Q.; Wu, Y.; Bonilla, L.S.; von Buchholtz, L.J.; Ryba, N.J.P. Diversity amongst trigeminal neurons revealed by high throughput single cell sequencing. PLoS ONE 2017, 12, e0185543. [Google Scholar] [CrossRef] [Green Version]
- Papalampropoulou-Tsiridou, M.; Labrecque, S.; Godin, A.G.; De Koninck, Y.; Wang, F. Differential Expression of Acid—Sensing Ion Channels in Mouse Primary Afferents in Naïve and Injured Conditions. Front. Cell. Neurosci. 2020, 14, 103. [Google Scholar] [CrossRef]
- Páez, O.; Segura-Chama, P.; Almanza, A.; Pellicer, F.; Mercado, F. Properties and Differential Expression of H+ Receptors in Dorsal Root Ganglia: Is a Labeled-Line Coding for Acid Nociception Possible? Front. Physiol. 2021, 12, 733267. [Google Scholar] [CrossRef]
- Duzhyy, D.E.; Voitenko, N.V.; Belan, P.V. Peripheral Inflammation Results in Increased Excitability of Capsaicin-Insensitive Nociceptive DRG Neurons Mediated by Upregulation of ASICs and Voltage-Gated Ion Channels. Front. Cell. Neurosci. 2021, 15, 723295. [Google Scholar] [CrossRef]
- Mamet, J.; Baron, A.; Lazdunski, M.; Voilley, N. Proinflammatory mediators, stimulators of sensory neuron excitability via the expression of acid-sensing ion channels. J. Neurosci. 2002, 22, 10662–10670. [Google Scholar] [CrossRef]
- Voilley, N.; de Weille, J.; Mamet, J.; Lazdunski, M. Nonsteroid anti-inflammatory drugs inhibit both the activity and the inflammation-induced expression of acid-sensing ion channels in nociceptors. J. Neurosci. 2001, 21, 8026–8033. [Google Scholar] [CrossRef] [PubMed]
- Tavares-Ferreira, D.; Shiers, S.; Ray, P.R.; Wangzhou, A.; Jeevakumar, V.; Sankaranarayanan, I.; Cervantes, A.M.; Reese, J.C.; Chamessian, A.; Copits, B.A.; et al. Spatial transcriptomics of dorsal root ganglia identifies molecular signatures of human nociceptors. Sci. Transl. Med. 2022, 14, eabj8186. [Google Scholar] [CrossRef] [PubMed]
- McKemy, D.D.; Flegel, C.; Schöbel, N.; Altmüller, J.; Becker, C.; Tannapfel, A.; Hatt, H.; Gisselmann, G. RNA-Seq Analysis of Human Trigeminal and Dorsal Root Ganglia with a Focus on Chemoreceptors. PLoS ONE 2015, 10, e0128951. [Google Scholar] [CrossRef] [Green Version]
- LaPaglia, D.M.; Sapio, M.R.; Burbelo, P.D.; Thierry-Mieg, J.; Thierry-Mieg, D.; Raithel, S.J.; Ramsden, C.E.; Iadarola, M.J.; Mannes, A.J. RNA-Seq investigations of human post-mortem trigeminal ganglia. Cephalalgia 2017, 38, 912–932. [Google Scholar] [CrossRef]
- Prato, V.; Taberner, F.J.; Hockley, J.R.F.; Callejo, G.; Arcourt, A.; Tazir, B.; Hammer, L.; Schad, P.; Heppenstall, P.A.; Smith, E.S.; et al. Functional and Molecular Characterization of Mechanoinsensitive “Silent” Nociceptors. Cell Rep. 2017, 21, 3102–3115. [Google Scholar] [CrossRef] [Green Version]
- Bakken, T.E.; Jorstad, N.L.; Hu, Q.; Lake, B.B.; Tian, W.; Kalmbach, B.E.; Crow, M.; Hodge, R.D.; Krienen, F.M.; Sorensen, S.A.; et al. Comparative cellular analysis of motor cortex in human, marmoset and mouse. Nature 2021, 598, 111–119. [Google Scholar] [CrossRef]
- Häring, M.; Zeisel, A.; Hochgerner, H.; Rinwa, P.; Jakobsson, J.E.T.; Lönnerberg, P.; La Manno, G.; Sharma, N.; Borgius, L.; Kiehn, O.; et al. Neuronal atlas of the dorsal horn defines its architecture and links sensory input to transcriptional cell types. Nat. Neurosci. 2018, 21, 869–880. [Google Scholar] [CrossRef]
- Sathyamurthy, A.; Johnson, K.R.; Matson, K.J.E.; Dobrott, C.I.; Li, L.; Ryba, A.R.; Bergman, T.B.; Kelly, M.C.; Kelley, M.W.; Levine, A.J. Massively Parallel Single Nucleus Transcriptional Profiling Defines Spinal Cord Neurons and Their Activity during Behavior. Cell Rep. 2018, 22, 2216–2225. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.J.; Duan, B.; Mei, Y.D.; Gao, J.; Chen, J.G.; Zhuo, M.; Xu, L.; Wu, M.; Xu, T.L. Characterization of acid-sensing ion channels in dorsal horn neurons of rat spinal cord. J. Biol. Chem. 2004, 279, 43716–43724. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Fu, X.; Huang, L.; Wang, X.; Lu, Z.; Zhu, F.; Xiao, Z. Increased Asics Expression via the Camkii-CREB Pathway in a Novel Mouse Model of Trigeminal Pain. Cell. Physiol. Biochem. 2018, 46, 568–578. [Google Scholar] [CrossRef]
- Li, X.; Xu, L.S.; Xu, Y.F.; Yang, Q.; Fang, Z.X.; Yao, M.; Chen, W.Y. The gene regulatory network in different brain regions of neuropathic pain mouse models. Eur. Rev. Med. Pharm. Sci. 2020, 24, 5053–5061. [Google Scholar] [CrossRef]
- Huang, C.; Hu, Z.-l.; Wu, W.-N.; Yu, D.-F.; Xiong, Q.-J.; Song, J.-R.; Shu, Q.; Fu, H.; Wang, F.; Chen, J.-G. Existence and distinction of acid-evoked currents in rat astrocytes. Glia 2010, 58, 1415–1424. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.-W.; Hu, Z.-L.; Ni, M.; Fang, P.; Zhang, P.-W.; Shu, Q.; Fan, H.; Zhou, H.-Y.; Ni, L.; Zhu, L.-Q.; et al. Acid-sensing ion channels promote the inflammation and migration of cultured rat microglia. Glia 2015, 63, 483–496. [Google Scholar] [CrossRef] [PubMed]
- Cegielski, V.; Chakrabarty, R.; Ding, S.; Wacker, M.J.; Monaghan-Nichols, P.; Chu, X.-P. Acid-Sensing Ion Channels in Glial Cells. Membranes 2022, 12, 119. [Google Scholar] [CrossRef]
- Yang, F.; Sun, X.; Ding, Y.; Ma, H.; Yang, T.O.; Ma, Y.; Wei, D.; Li, W.; Xu, T.; Jiang, W. Astrocytic Acid-Sensing Ion Channel 1a Contributes to the Development of Chronic Epileptogenesis. Sci. Rep. 2016, 6, 31581. [Google Scholar] [CrossRef] [Green Version]
- Feldman, D.H.; Horiuchi, M.; Keachie, K.; McCauley, E.; Bannerman, P.; Itoh, A.; Itoh, T.; Pleasure, D. Characterization of acid-sensing ion channel expression in oligodendrocyte-lineage cells. Glia 2008, 56, 1238–1249. [Google Scholar] [CrossRef] [Green Version]
- Baumann, T.K.; Burchiel, K.J.; Ingram, S.L.; Martenson, M.E. Responses of adult human dorsal root ganglion neurons in culture to capsaicin and low pH. Pain 1996, 65, 31–38. [Google Scholar] [CrossRef]
- Baumann, T.K.; Chaudhary, P.; Martenson, M.E. Background potassium channel block and TRPV1 activation contribute to proton depolarization of sensory neurons from humans with neuropathic pain. Eur. J. Neurosci. 2004, 19, 1343–1351. [Google Scholar] [CrossRef]
- North, R.Y.; Li, Y.; Ray, P.; Rhines, L.D.; Tatsui, C.E.; Rao, G.; Johansson, C.A.; Zhang, H.; Kim, Y.H.; Zhang, B.; et al. Electrophysiological and transcriptomic correlates of neuropathic pain in human dorsal root ganglion neurons. Brain 2019, 142, 1215–1226. [Google Scholar] [CrossRef]
- Li, M.; Inoue, K.; Branigan, D.; Kratzer, E.; Hansen, J.C.; Chen, J.W.; Simon, R.P.; Xiong, Z.-G. Acid-Sensing Ion Channels in Acidosis-Induced Injury of Human Brain Neurons. J. Cereb. Blood Flow Metab. 2010, 30, 1247–1260. [Google Scholar] [CrossRef]
- Xu, Y.; Jiang, Y.Q.; Li, C.; He, M.; Rusyniak, W.G.; Annamdevula, N.; Ochoa, J.; Leavesley, S.J.; Xu, J.; Rich, T.C.; et al. Human ASIC1a mediates stronger acid-induced responses as compared with mouse ASICIa. FASEB J. 2018, 32, 3832–3843. [Google Scholar] [CrossRef] [Green Version]
- Neuhof, A.; Tian, Y.; Reska, A.; Falkenburger, B.H.; Gründer, S. Large Acid-Evoked Currents, Mediated by ASIC1a, Accompany Differentiation in Human Dopaminergic Neurons. Front. Cell. Neurosci. 2021, 15, 668008. [Google Scholar] [CrossRef] [PubMed]
- Kalinovskii, A.P.; Osmakov, D.I.; Koshelev, S.G.; Lubova, K.I.; Korolkova, Y.V.; Kozlov, S.A.; Andreev, Y.A. Retinoic Acid-Differentiated Neuroblastoma SH-SY5Y Is an Accessible In Vitro Model to Study Native Human Acid-Sensing Ion Channels 1a (ASIC1a). Biology 2022, 11, 167. [Google Scholar] [CrossRef] [PubMed]
- Steen, K.H.; Steen, A.E.; Reeh, P.W. A dominant role of acid pH in inflammatory excitation and sensitization of nociceptors in rat skin, in vitro. J. Neurosci. 1995, 15, 3982–3989. [Google Scholar] [CrossRef] [Green Version]
- McMahon, S.B.; Jones, N.G. Plasticity of pain signaling: Role of neurotrophic factors exemplified by acid-induced pain. J. Neurobiol. 2004, 61, 72–87. [Google Scholar] [CrossRef]
- Heber, S.; Ciotu, C.I.; Hartner, G.; Gold-Binder, M.; Ninidze, N.; Gleiss, A.; Kress, H.-G.; Fischer, M.J.M. TRPV1 antagonist BCTC inhibits pH 6.0-induced pain in human skin. Pain 2020, 161, 1532–1541. [Google Scholar] [CrossRef]
- Morgan, D.L.; Borys, D.J.; Stanford, R.; Kjar, D.; Tobleman, W. Texas Coral Snake (Micrurus tener) Bites. South. Med. J. 2007, 100, 152–156. [Google Scholar] [CrossRef]
- Lopes, D.M.; Denk, F.; McMahon, S.B. The Molecular Fingerprint of Dorsal Root and Trigeminal Ganglion Neurons. Front. Mol. Neurosci. 2017, 10, 304. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Edelmayer, R.M.; Wei, X.; De Felice, M.; Porreca, F.; Dussor, G. Dural afferents express acid-sensing ion channels: A role for decreased meningeal pH in migraine headache. Pain 2011, 152, 106–113. [Google Scholar] [CrossRef] [Green Version]
- Holton, C.M.; Strother, L.C.; Dripps, I.; Pradhan, A.A.; Goadsby, P.J.; Holland, P.R. Acid-sensing ion channel 3 blockade inhibits durovascular and nitric oxide-mediated trigeminal pain. Br. J. Pharmacol. 2020, 177, 2478–2486. [Google Scholar] [CrossRef]
- Dussor, G. ASICs as therapeutic targets for migraine. Neuropharmacology 2015, 94, 64–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holland, P.R.; Akerman, S.; Andreou, A.P.; Karsan, N.; Wemmie, J.A.; Goadsby, P.J. Acid-sensing ion channel 1: A novel therapeutic target for migraine with aura. Ann. Neurol. 2012, 72, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Ding, J.; Wu, J.; Liu, T.; Liang, J.; Tang, Q.; Jiao, M. Resveratrol attenuates bone cancer pain through regulating the expression levels of ASIC3 and activating cell autophagy. Acta Biochim. Et Biophys. Sin. 2017, 49, 1008–1014. [Google Scholar] [CrossRef] [Green Version]
- Morgan, M.; Thai, J.; Trinh, P.; Habib, M.; Effendi, K.N.; Ivanusic, J.J. ASIC3 inhibition modulates inflammation-induced changes in the activity and sensitivity of Aδ and C fiber sensory neurons that innervate bone. Mol. Pain 2020, 16, 1744806920975950. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Kolker, S.J.; Sluka, K.A. Acid-Sensing Ion Channel 3 Expression in Mouse Knee Joint Afferents and Effects of Carrageenan-Induced Arthritis. J. Pain 2009, 10, 336–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurczak, A.; Delay, L.; Barbier, J.; Simon, N.; Krock, E.; Sandor, K.; Agalave, N.M.; Rudjito, R.; Wigerblad, G.; Rogóż, K.; et al. Antibody-induced pain-like behavior and bone erosion: Links to subclinical inflammation, osteoclast activity, and acid-sensing ion channel 3–dependent sensitization. Pain 2022, 163, 1542–1559. [Google Scholar] [CrossRef]
- Pidoux, L.; Delanoe, K.; Barbier, J.; Marchand, F.; Lingueglia, E.; Deval, E. Single Subcutaneous Injection of Lysophosphatidyl-Choline Evokes ASIC3-Dependent Increases of Spinal Dorsal Horn Neuron Activity. Front. Mol. Neurosci. 2022, 15, 880651. [Google Scholar] [CrossRef]
- Holzer, P. Acid-sensing ion channels in gastrointestinal function. Neuropharmacology 2015, 94, 72–79. [Google Scholar] [CrossRef] [Green Version]
- Yiangou, Y.; Facer, P.; Smith, J.A.; Sangameswaran, L.; Eglen, R.; Birch, R.; Knowles, C.; Williams, N.; Anand, P. Increased acid-sensing ion channel ASIC-3 in inflamed human intestine. Eur. J. Gastroenterol. Hepatol. 2001, 13, 891–896. [Google Scholar] [CrossRef]
- Han, X.; Zhang, Y.; Lee, A.; Li, Z.; Gao, J.; Wu, X.; Zhao, J.; Wang, H.; Chen, D.; Zou, D.; et al. Upregulation of acid sensing ion channels is associated with esophageal hypersensitivity in GERD. FASEB J. 2021, 36, e22083. [Google Scholar] [CrossRef]
- Olesen, A.E.; Nielsen, L.M.; Larsen, I.M.; Drewes, A.M. Randomized clinical trial: Efficacy and safety of PPC-5650 on experimental esophageal pain and hyperalgesia in healthy volunteers. Scand. J. Gastroenterol. 2014, 50, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Sluka, K.A.; Gregory, N.S. The dichotomized role for acid sensing ion channels in musculoskeletal pain and inflammation. Neuropharmacology 2015, 94, 58–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walder, R.Y.; Rasmussen, L.A.; Rainier, J.D.; Light, A.R.; Wemmie, J.A.; Sluka, K.A. ASIC1 and ASIC3 Play Different Roles in the Development of Hyperalgesia After Inflammatory Muscle Injury. J. Pain 2010, 11, 210–218. [Google Scholar] [CrossRef] [Green Version]
- Hung, C.-H.; Lee, C.-H.; Tsai, M.-H.; Chen, C.-H.; Lin, H.-F.; Hsu, C.-Y.; Lai, C.-L.; Chen, C.-C. Activation of acid-sensing ion channel 3 by lysophosphatidylcholine 16:0 mediates psychological stress-induced fibromyalgia-like pain. Ann. Rheum. Dis. 2020, 79, 1644–1656. [Google Scholar] [CrossRef]
- Taguchi, T.; Katanosaka, K.; Yasui, M.; Hayashi, K.; Yamashita, M.; Wakatsuki, K.; Kiyama, H.; Yamanaka, A.; Mizumura, K. Peripheral and spinal mechanisms of nociception in a rat reserpine-induced pain model. Pain 2015, 156, 415–427. [Google Scholar] [CrossRef]
- Yen, L.-T.; Hsieh, C.-L.; Hsu, H.-C.; Lin, Y.-W. Preventing the induction of acid saline-induced fibromyalgia pain in mice by electroacupuncture or APETx2 injection. Acupunct. Med. 2020, 38, 188–193. [Google Scholar] [CrossRef]
- Gao, M.; Yan, X.; Lu, Y.; Ren, L.; Zhang, S.; Zhang, X.; Kuang, Q.; Liu, L.; Zhou, J.; Wang, Y.; et al. Retrograde nerve growth factor signaling modulates tooth mechanical hyperalgesia induced by orthodontic tooth movement via acid-sensing ion channel 3. Int. J. Oral Sci. 2021, 13, 18. [Google Scholar] [CrossRef]
- Long, H.; Wang, Y.; Jian, F.; Liao, L.-N.; Yang, X.; Lai, W.-L. Current advances in orthodontic pain. Int. J. Oral Sci. 2016, 8, 67–75. [Google Scholar] [CrossRef]
- Yang, H.; Shan, D.; Jin, Y.; Liang, H.; Liu, L.; Guan, Y.; Chen, C.; Luo, Q.; Yang, Y.; Lai, W.; et al. The Role of Acid-sensing Ion Channel 3 in the Modulation of Tooth Mechanical Hyperalgesia Induced by Orthodontic Tooth Movement. Neuroscience 2020, 442, 274–285. [Google Scholar] [CrossRef]
- Osada, A.; Hitomi, S.; Nakajima, A.; Hayashi, Y.; Shibuta, I.; Tsuboi, Y.; Motoyoshi, M.; Iwata, K.; Shinoda, M. Periodontal acidification contributes to tooth pain hypersensitivity during orthodontic tooth movement. Neurosci. Res. 2022, 177, 103–110. [Google Scholar] [CrossRef]
- Zhang, S.; Divaris, K.; Moss, K.; Yu, N.; Barros, S.; Marchesan, J.; Morelli, T.; Agler, C.; Kim, S.J.; Wu, D.; et al. The Novel ASIC2 Locus Is Associated with Severe Gingival Inflammation. JDR Clin. Transl. Res. 2016, 1, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Matriconl, J.; Gelotl, A.; Etiennel, M.; Lazdunskil, M.; Mullerl, E.; Ardidl, D. Spinal cord plasticity and acid-sensing ion channels involvement in a rodent model of irritable bowel syndrome. Eur. J. Pain 2011, 15, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Duan, B.; Wang, Y.Z.; Yang, T.; Chu, X.P.; Yu, Y.; Huang, Y.; Cao, H.; Hansen, J.; Simon, R.P.; Zhu, M.X.; et al. Extracellular Spermine Exacerbates Ischemic Neuronal Injury through Sensitization of ASIC1a Channels to Extracellular Acidosis. J. Neurosci. 2011, 31, 2101–2112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Z.-G.; Zhu, X.-M.; Chu, X.-P.; Minami, M.; Hey, J.; Wei, W.-L.; MacDonald, J.F.; Wemmie, J.A.; Price, M.P.; Welsh, M.J.; et al. Neuroprotection in Ischemia. Cell 2004, 118, 687–698. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.B.; Jin, K.; Minami, M.; Chen, D.; Simon, R.P. Global Ischemia Induces Expression of Acid-Sensing Ion Channel 2a in Rat Brain. J. Cereb. Blood Flow Metab. 2016, 21, 734–740. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Duan, B.; Wang, D.-G.; Deng, X.-H.; Zhang, G.-Y.; Xu, L.; Xu, T.-L. Coupling between NMDA Receptor and Acid-Sensing Ion Channel Contributes to Ischemic Neuronal Death. Neuron 2005, 48, 635–646. [Google Scholar] [CrossRef] [Green Version]
- Pignataro, G.; Simon, R.P.; Xiong, Z.G. Prolonged activation of ASIC1a and the time window for neuroprotection in cerebral ischaemia. Brain 2006, 130, 151–158. [Google Scholar] [CrossRef]
- Ren, Y.; Li, C.; Chang, J.; Wang, R.; Wang, Y.; Chu, X.-P. Hi1a as a Novel Neuroprotective Agent for Ischemic Stroke by Inhibition of Acid-Sensing Ion Channel 1a. Transl. Stroke Res. 2017, 9, 96–98. [Google Scholar] [CrossRef]
- Hu, R.; Duan, B.; Wang, D.; Yu, Y.; Li, W.; Luo, H.; Lu, P.; Lin, J.; Zhu, G.; Wan, Q.; et al. Role of Acid-Sensing Ion Channel 1a in the Secondary Damage of Traumatic Spinal Cord Injury. Ann. Surg. 2011, 254, 353–362. [Google Scholar] [CrossRef]
- Vergo, S.; Craner, M.J.; Etzensperger, R.; Attfield, K.; Friese, M.A.; Newcombe, J.; Esiri, M.; Fugger, L. Acid-sensing ion channel 1 is involved in both axonal injury and demyelination in multiple sclerosis and its animal model. Brain 2011, 134, 571–584. [Google Scholar] [CrossRef] [Green Version]
- Friese, M.A.; Craner, M.J.; Etzensperger, R.; Vergo, S.; Wemmie, J.A.; Welsh, M.J.; Vincent, A.; Fugger, L. Acid-sensing ion channel-1 contributes to axonal degeneration in autoimmune inflammation of the central nervous system. Nat. Med. 2007, 13, 1483–1489. [Google Scholar] [CrossRef] [PubMed]
- Arun, T.; Tomassini, V.; Sbardella, E.; de Ruiter, M.B.; Matthews, L.; Leite, M.I.; Gelineau-Morel, R.; Cavey, A.; Vergo, S.; Craner, M.; et al. Targeting ASIC1 in primary progressive multiple sclerosis: Evidence of neuroprotection with amiloride. Brain 2013, 136, 106–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mango, D.; Nisticò, R. Neurodegenerative Disease: What Potential Therapeutic Role of Acid-Sensing Ion Channels? Front. Cell. Neurosci. 2021, 15, 730641. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Ramírez, A.; Vega, R.; Soto, E. Acid-Sensing Ion Channels as Potential Therapeutic Targets in Neurodegeneration and Neuroinflammation. Mediat. Inflamm. 2017, 2017, 3728096. [Google Scholar] [CrossRef] [Green Version]
- Arias, R.L.; Sung, M.-L.A.; Vasylyev, D.; Zhang, M.-Y.; Albinson, K.; Kubek, K.; Kagan, N.; Beyer, C.; Lin, Q.; Dwyer, J.M.; et al. Amiloride is neuroprotective in an MPTP model of Parkinson’s disease. Neurobiol. Dis. 2008, 31, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Cao, Y.-B.; Hu, L.-F.; Yang, Y.-P.; Li, J.; Wang, F.; Liu, C.-F. ASICs mediate the modulatory effect by paeoniflorin on alpha-synuclein autophagic degradation. Brain Res. 2011, 1396, 77–87. [Google Scholar] [CrossRef]
- Gu, X.-S.; Wang, F.; Zhang, C.-Y.; Mao, C.-J.; Yang, J.; Yang, Y.-P.; Liu, S.; Hu, L.-F.; Liu, C.-F. Neuroprotective Effects of Paeoniflorin on 6-OHDA-Lesioned Rat Model of Parkinson’s Disease. Neurochem. Res. 2016, 41, 2923–2936. [Google Scholar] [CrossRef]
- Lai, K.; Song, X.-L.; Shi, H.-S.; Qi, X.; Li, C.-Y.; Fang, J.; Wang, F.; Maximyuk, O.; Krishtal, O.; Xu, T.-L.; et al. Bilirubin enhances the activity of ASIC channels to exacerbate neurotoxicity in neonatal hyperbilirubinemia in mice. Sci. Transl. Med. 2020, 12, eaax1337. [Google Scholar] [CrossRef]
- Cuesta, A.; del Valle, M.E.; García-Suárez, O.; Viña, E.; Cabo, R.; Vázquez, G.; Cobo, J.L.; Murcia, A.; Alvarez-Vega, M.; García-Cosamalón, J.; et al. Acid-sensing ion channels in healthy and degenerated human intervertebral disc. Connect. Tissue Res. 2014, 55, 197–204. [Google Scholar] [CrossRef]
- Ding, J.; Zhang, R.; Li, H.; Ji, Q.; Cheng, X.; Thorne, R.F.; Hondermarck, H.; Liu, X.; Shen, C. ASIC1 and ASIC3 mediate cellular senescence of human nucleus pulposus mesenchymal stem cells during intervertebral disc degeneration. Aging 2021, 13, 10703–10723. [Google Scholar] [CrossRef]
- Li, X.; Wu, F.R.; Xu, R.S.; Hu, W.; Jiang, D.L.; Ji, C.; Chen, F.H.; Yuan, F.L. Acid-sensing ion channel 1a-mediated calcium influx regulates apoptosis of endplate chondrocytes in intervertebral discs. Expert Opin. Ther. Targets 2013, 18, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Ding, J.; Chen, Y.; Ma, G.; Wei, X.; Zhou, R.; Hu, W. Blockade of ASIC1a inhibits acid-induced rat articular chondrocyte senescence through regulation of autophagy. Hum. Cell 2022, 35, 665–677. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xu, R.-S.; Jiang, D.-L.; He, X.-L.; Jin, C.; Lu, W.-G.; Su, Q.; Yuan, F.-L. Acid-sensing ion channel 1a is involved in acid-induced osteoclastogenesis by regulating activation of the transcription factor NFATc1. FEBS Lett. 2013, 587, 3236–3242. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Han, L.; Chen, H.; Zhang, S.; Zhang, S.; Zhang, H.; Li, Y.; Tao, H.; Li, J. Sa12b Improves Biological Activity of Human Degenerative Nucleus Pulposus Mesenchymal Stem Cells in a Severe Acid Environment by Inhibiting Acid-Sensitive Ion Channels. Front. Bioeng. Biotechnol. 2022, 10, 816362. [Google Scholar] [CrossRef]
- Han, L.; Wang, Z.; Chen, H.; Li, J.; Zhang, S.; Zhang, S.; Shao, S.; Zhang, Y.; Shen, C.; Tao, H. Sa12b-Modified Functional Self-Assembling Peptide Hydrogel Enhances the Biological Activity of Nucleus Pulposus Mesenchymal Stem Cells by Inhibiting Acid-Sensing Ion Channels. Front. Cell Dev. Biol. 2022, 10, 822501. [Google Scholar] [CrossRef]
- Abboud, F.M.; Benson, C.J. ASICs and cardiovascular homeostasis. Neuropharmacology 2015, 94, 87–98. [Google Scholar] [CrossRef] [Green Version]
- Fromy, B.; Lingueglia, E.; Sigaudo-Roussel, D.; Saumet, J.L.; Lazdunski, M. Asic3 is a neuronal mechanosensor for pressure-induced vasodilation that protects against pressure ulcers. Nat. Med. 2012, 18, 1205–1207. [Google Scholar] [CrossRef]
- Weavil, J.C.; Kwon, O.S.; Hughen, R.W.; Zhang, J.; Light, A.R.; Amann, M. Gene and protein expression of dorsal root ganglion sensory receptors in normotensive and hypertensive male rats. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2022, 323, R221–R226. [Google Scholar] [CrossRef] [PubMed]
- Faraci, F.M.; Taugher, R.J.; Lynch, C.; Fan, R.; Gupta, S.; Wemmie, J.A. Acid-Sensing Ion Channels. Circ. Res. 2019, 125, 907–920. [Google Scholar] [CrossRef]
- Gupta, S.C.; Singh, R.; Asters, M.; Liu, J.; Zhang, X.; Pabbidi, M.R.; Watabe, K.; Mo, Y.Y. Regulation of breast tumorigenesis through acid sensors. Oncogene 2015, 35, 4102–4111. [Google Scholar] [CrossRef]
- Yang, C.; Zhu, Z.; Ouyang, X.; Yu, R.; Wang, J.; Ding, G.; Jiang, F. Overexpression of acid-sensing ion channel 1a (ASIC1a) promotes breast cancer cell proliferation, migration and invasion. Transl. Cancer Res. 2020, 9, 7519–7530. [Google Scholar] [CrossRef] [PubMed]
- Bychkov, M.; Shulepko, M.; Osmakov, D.; Andreev, Y.; Sudarikova, A.; Vasileva, V.; Pavlyukov, M.S.; Latyshev, Y.A.; Potapov, A.A.; Kirpichnikov, M.; et al. Mambalgin-2 Induces Cell Cycle Arrest and Apoptosis in Glioma Cells via Interaction with ASIC1a. Cancers 2020, 12, 1837. [Google Scholar] [CrossRef]
- Bychkov, M.L.; Kirichenko, A.V.; Shulepko, M.A.; Mikhaylova, I.N.; Kirpichnikov, M.P.; Lyukmanova, E.N. Mambalgin-2 Inhibits Growth, Migration, and Invasion of Metastatic Melanoma Cells by Targeting the Channels Containing an ASIC1a Subunit Whose Up-Regulation Correlates with Poor Survival Prognosis. Biomedicines 2021, 9, 1324. [Google Scholar] [CrossRef] [PubMed]
- Bychkov, M.L.; Shulepko, M.A.; Vasileva, V.Y.; Sudarikova, A.V.; Kirpichnikov, M.P.; Lyukmanova, E.N. ASIC1a Inhibitor mambalgin-2 Suppresses the Growth of Leukemia Cells by Cell Cycle Arrest. Acta Nat. 2020, 12, 101–116. [Google Scholar] [CrossRef]
- Smoller, J.W.; Gallagher, P.J.; Duncan, L.E.; McGrath, L.M.; Haddad, S.A.; Holmes, A.J.; Wolf, A.B.; Hilker, S.; Block, S.R.; Weill, S.; et al. The Human Ortholog of Acid-Sensing Ion Channel Gene ASIC1a Is Associated With Panic Disorder and Amygdala Structure and Function. Biol. Psychiatry 2014, 76, 902–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gugliandolo, A.; Gangemi, C.; Caccamo, D.; Currò, M.; Pandolfo, G.; Quattrone, D.; Crucitti, M.; Zoccali, R.A.; Bruno, A.; Muscatello, M.R.A. The RS685012 Polymorphism of ACCN2, the Human Ortholog of Murine Acid-Sensing Ion Channel (ASIC1) Gene, is Highly Represented in Patients with Panic Disorder. NeuroMolecular Med. 2015, 18, 91–98. [Google Scholar] [CrossRef]
- Coryell, M.W.; Wunsch, A.M.; Haenfler, J.M.; Allen, J.E.; Schnizler, M.; Ziemann, A.E.; Cook, M.N.; Dunning, J.P.; Price, M.P.; Rainier, J.D.; et al. Acid-Sensing Ion Channel-1a in the Amygdala, a Novel Therapeutic Target in Depression-Related Behavior. J. Neurosci. 2009, 29, 5381–5388. [Google Scholar] [CrossRef] [Green Version]
- Taugher, R.J.; Lu, Y.; Fan, R.; Ghobbeh, A.; Kreple, C.J.; Faraci, F.M.; Wemmie, J.A. ASIC1A in neurons is critical for fear-related behaviors. Genes Brain Behav. 2017, 16, 745–755. [Google Scholar] [CrossRef] [Green Version]
- Pidoplichko, V.I.; Aroniadou-Anderjaska, V.; Prager, E.M.; Figueiredo, T.H.; Almeida-Suhett, C.P.; Miller, S.L.; Braga, M.F.M. ASIC1a Activation Enhances Inhibition in the Basolateral Amygdala and Reduces Anxiety. J. Neurosci. 2014, 34, 3130–3141. [Google Scholar] [CrossRef] [Green Version]
- Ziemann, A.E.; Schnizler, M.K.; Albert, G.W.; Severson, M.A.; Howard, M.A., 3rd; Welsh, M.J.; Wemmie, J.A. Seizure termination by acidosis depends on ASIC1a. Nat. Neurosci. 2008, 11, 816–822. [Google Scholar] [CrossRef]
- Lv, R.-j.; He, J.-s.; Fu, Y.-h.; Zhang, Y.-q.; Shao, X.-q.; Wu, L.-w.; Lu, Q.; Jin, L.-r.; Liu, H. ASIC1a polymorphism is associated with temporal lobe epilepsy. Epilepsy Res. 2011, 96, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Chen, X.; He, J.-J.; Wei, Y.-J.; Zang, Z.-L.; Liu, S.-Y.; Yang, H.; Zhang, C.-Q. Down-Regulated Expression of Acid-Sensing Ion Channel 1a in Cortical Lesions of Patients with Focal Cortical Dysplasia. J. Mol. Neurosci. 2014, 53, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Wang, W.; Gu, J.; Jiang, G.; Wang, K.; Xu, Z.; Li, J.; Chen, G.; Wang, X. Elevated Expression of Acid-Sensing Ion Channel 3 Inhibits Epilepsy via Activation of Interneurons. Mol. Neurobiol. 2014, 53, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Stein, C.; Kopf, A. Pain therapy—Are there new options on the horizon? Best Pract. Res. Clin. Rheumatol. 2019, 33, 101420. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isoform | Species | % Identity | Size (aa) | Name in Genbank | Sequence ID |
|---|---|---|---|---|---|
| ASIC1a | Rattus norvegicus | 98.11% | 526 | ASIC1 | NP_077068.1 |
| Homo sapiens | 528 | ASIC1 isoform b | NP_001086.2 | ||
| ASIC1b | Rattus norvegicus | 93.24% | 559 | ASIC1 isoform X5 | XP_006257440.1 |
| Homo sapiens | 562 | ASIC1 isoform c | NP_001243759.1 | ||
| ASIC2a | Rattus norvegicus | 99.02% | 512 | ASIC2 isoform MDEG1 | NP_001029186.1 |
| Homo sapiens | 512 | ASIC2 isoform MDEG1 | NP_001085.2 | ||
| ASIC2b | Rattus norvegicus | 98.83% | 563 | ASIC2 isoform MDEG2 | NP_037024.2 |
| Homo sapiens | 563 | ASIC2 isoform MDEG2 | NP_899233.1 | ||
| ASIC3 | Rattus norvegicus | 83.68% | 533 | ASIC3 | NP_775158.1 |
| Homo sapiens | 531 | ASIC3 isoform a | NP_004760.1 | ||
| ASIC4 | Rattus norvegicus | 97.22% | 539 | ASIC4 | NP_071570.2 |
| Homo sapiens | 539 | ASIC4 isoform 1 | NP_061144.4 |
| Cloned Channel | ACTIVATION | SSD | ||
|---|---|---|---|---|
| Test pH Threshold/max | pH0.5 | Conditioning pH Threshold/max | pH0.5 | |
| rASIC1a | 7.0/5.5 | 6.4–5.8 chimnqtwxyz | 7.4/6.8 | 7.3–7.1 cehimtyz |
| rASIC1b | 6.4/5.6 | 6.3–5.7 fitwxy# | 7.3/6.6 | 7.0–6.5 fit# |
| m/rASIC2a | 6.0/3.0 | 5.0–3.8 bnqwxz | 7.0/4.5 | 6.3–5.6 mz |
| m/rASIC3 | 7.2/5.5 | 6.8–6.3 otwy | 7.4/6.8 | 7.2–7.0 sty |
| rASIC1a/2a | 6.3/4.5 | 5.6–4.8 nqrw | ||
| m/rASIC1a/2b | 6.8/6.0 | 6.4–6.2 pw | 7.4/7.1 | 7.3 p |
| rASIC1a/1b | 6.3–5.8 w | |||
| rASIC1a/3 | 7.0/5.5 | 6.7–6.3 rtw | 7.0/6.8 | 7.1 t |
| rASIC1b/3 | 6.6/5.9 | 6.7–6.2 tw | 6.9/6.6 | 6.8 t |
| rASIC1b/2a | 4.9 w | |||
| rASIC2a/3 | 7.2/4.5 | 6.1–5.6 rw | ||
| m/rASIC2a/2b | 4.8 bw | |||
| rASIC2b/3 | 6.5 w | |||
| m/rASIC1a/2a/3 | 6.4–5.1 rw | |||
| rASIC1a/2b/3 | 6.3 w | |||
| rASIC1b/2a/3 | 4.9 w | |||
| hASIC1a | 6.8/6.0 | 6.6–6.3 dgikov | 7.0/6.7 | 7.2–6.9 degiko |
| hASIC1b | 6.5/5.5 | 5.9–5.7 gi | 6.7/6.4 | 6.5–6.1 gi |
| hASIC2a | 6.8/3.5 | 5.7 u | 6.0/4.7 | 5.5 u |
| hASIC3a | 7.0/5.5 | 6.6–6.2 aj | 7.0/7.9 | 7.7–7.5 as |
| cASIC1 | 6.8/6.3 | 6.6 l | 7.4/7.1 | 7.3 l |
| Channel | Mamb (1, 2 or 3) IC50/EC50 | PcTx1 IC50/EC50 | APETx2 IC50/EC50 | MitTx EC50 |
|---|---|---|---|---|
| rASIC1a | INH🔾◆ 3–55 nM a,p,q,r | INH🔾◆ 0.3–3.7 nM b,c,o,u | NO🔾◆ at 10 µM y,e | ACT🔾 9 nM i |
| rASIC1b | INH🔾◆ 22–192 nM a,p,q,r | POT🔾 25–100 nM d, u | NO◆ at 3 µM e POT🔾 at 3–10 µM y,# | ACT🔾 23 nM i |
| rASIC2a | NO🔾◆ at 3 µM a,p | NO🔾 at 100 nM b | NO🔾◆ at 3 µM e,# POT🔾 at 10 µM y | POT🔾 at 75 nM i |
| rASIC3 | NO🔾◆ at 3 µM a,p | NO🔾 at 100 nM b | INH🔾◆ 37–63 nM, test pH 6 e,f,g | ACT🔾 830 nM i |
| rASIC1a/2a | INH🔾◆ 152–252 nM a,p,r | NO🔾◆ at 50 nM b,v,w | ACT🔾 at 75 nM i | |
| rASIC1a/2b | INH◆ 61 nM a | INH🔾 3 nM h NO◆ at 300 nM # | ||
| rASIC1a/1b | INH◆ 72 nM a | |||
| rASIC1a/3 | NO◆ at 2 µM a,p | NO◆ at 10 nM b INH◆ at 100 nM, test pH 6 x | INH🔾◆ 2 µM e,# | NO🔾 at 75 nM i |
| rASIC1b/3 | INH◆ at 1 µM # POT◆ at 1 µM, test pH 6.6 # | INH◆ 900 nM, test pH 6 e POT🔾 at 3 µM, test pH 6 # | ||
| rASIC1b/2a | INH◆at 1 µM # | |||
| rASIC1b/2b | INH◆at 100 nM # | |||
| rASIC2a/3 | NO◆ at 3 µM e POT🔾 at 3 µM, test pH 6 # | NO🔾 at 75 nM i | ||
| rASIC2b/3 | INH🔾◆ 117 nM e,# | |||
| mASIC1a/2a/3 | NO◆ at 100 nM x | |||
| hASIC1a | INH🔾◆ 24–203 nM a,q,s,t | NO🔾◆ at 25–60 nM m,n,j INH🔾 3.3 nM, test pH 6 u POT🔾 at 60 nM, test pH 6.7 n | ||
| hASIC1b | INH🔾 at 1 µM POT🔾◆ 60 nM, test pH6 q,# | POT🔾 at 60 nM n | ||
| hASIC2a | NO◆ at 850 nM a | NO◆ at 100 nM j | ||
| hASIC3a | INH🔾◆ 175–344 nM e,z | |||
| hASIC1a/2a | INH◆ 220 nM a | NO◆ at 100 nM j POT🔾 at 200 nM h | ||
| hASIC1a/1b | INH🔾 256 nM q | |||
| hASIC1a/3 | INH🔾 462 nM q | |||
| hASIC1b/3 | INH🔾 at 1 µM q POT🔾 at 1 µM, test pH 6 q | |||
| cASIC1 | INH◆ 124 nM, test pH 6 t | ACT🔾◆ 189 nM l,k,p POT🔾◆ at 20 nM k,m |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Verkest, C.; Salinas, M.; Diochot, S.; Deval, E.; Lingueglia, E.; Baron, A. Mechanisms of Action of the Peptide Toxins Targeting Human and Rodent Acid-Sensing Ion Channels and Relevance to Their In Vivo Analgesic Effects. Toxins 2022, 14, 709. https://doi.org/10.3390/toxins14100709
Verkest C, Salinas M, Diochot S, Deval E, Lingueglia E, Baron A. Mechanisms of Action of the Peptide Toxins Targeting Human and Rodent Acid-Sensing Ion Channels and Relevance to Their In Vivo Analgesic Effects. Toxins. 2022; 14(10):709. https://doi.org/10.3390/toxins14100709
Chicago/Turabian StyleVerkest, Clément, Miguel Salinas, Sylvie Diochot, Emmanuel Deval, Eric Lingueglia, and Anne Baron. 2022. "Mechanisms of Action of the Peptide Toxins Targeting Human and Rodent Acid-Sensing Ion Channels and Relevance to Their In Vivo Analgesic Effects" Toxins 14, no. 10: 709. https://doi.org/10.3390/toxins14100709
APA StyleVerkest, C., Salinas, M., Diochot, S., Deval, E., Lingueglia, E., & Baron, A. (2022). Mechanisms of Action of the Peptide Toxins Targeting Human and Rodent Acid-Sensing Ion Channels and Relevance to Their In Vivo Analgesic Effects. Toxins, 14(10), 709. https://doi.org/10.3390/toxins14100709