Assessment of Zearalenone-Induced Cell Survival and of Global Gene Regulation in Mouse TM4 Sertoli Cells

Abstract
:1. Introduction
2. Results
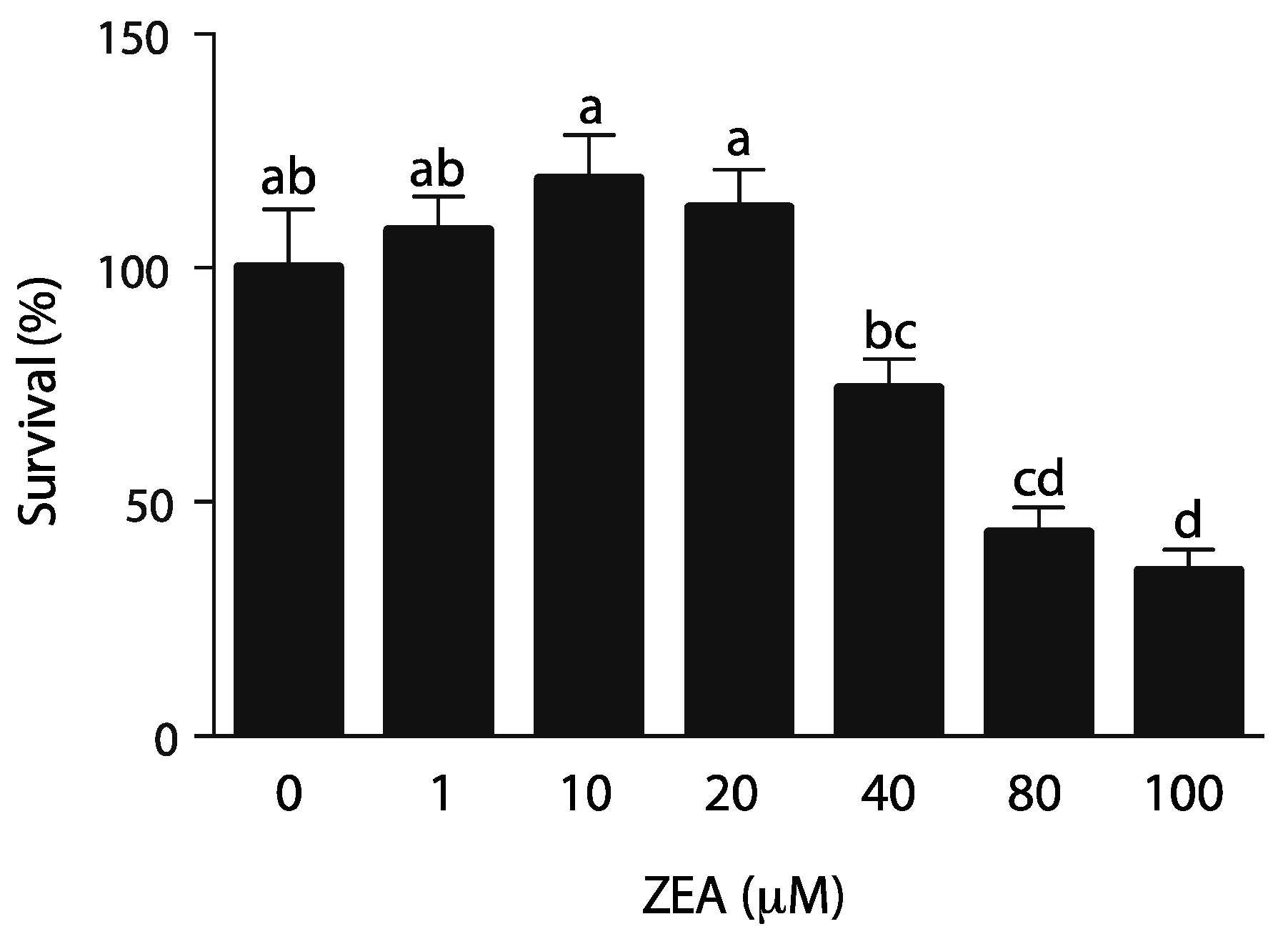
2.1. TM4 Immature Sertoli Cell Line Is Sensitive to ZEA
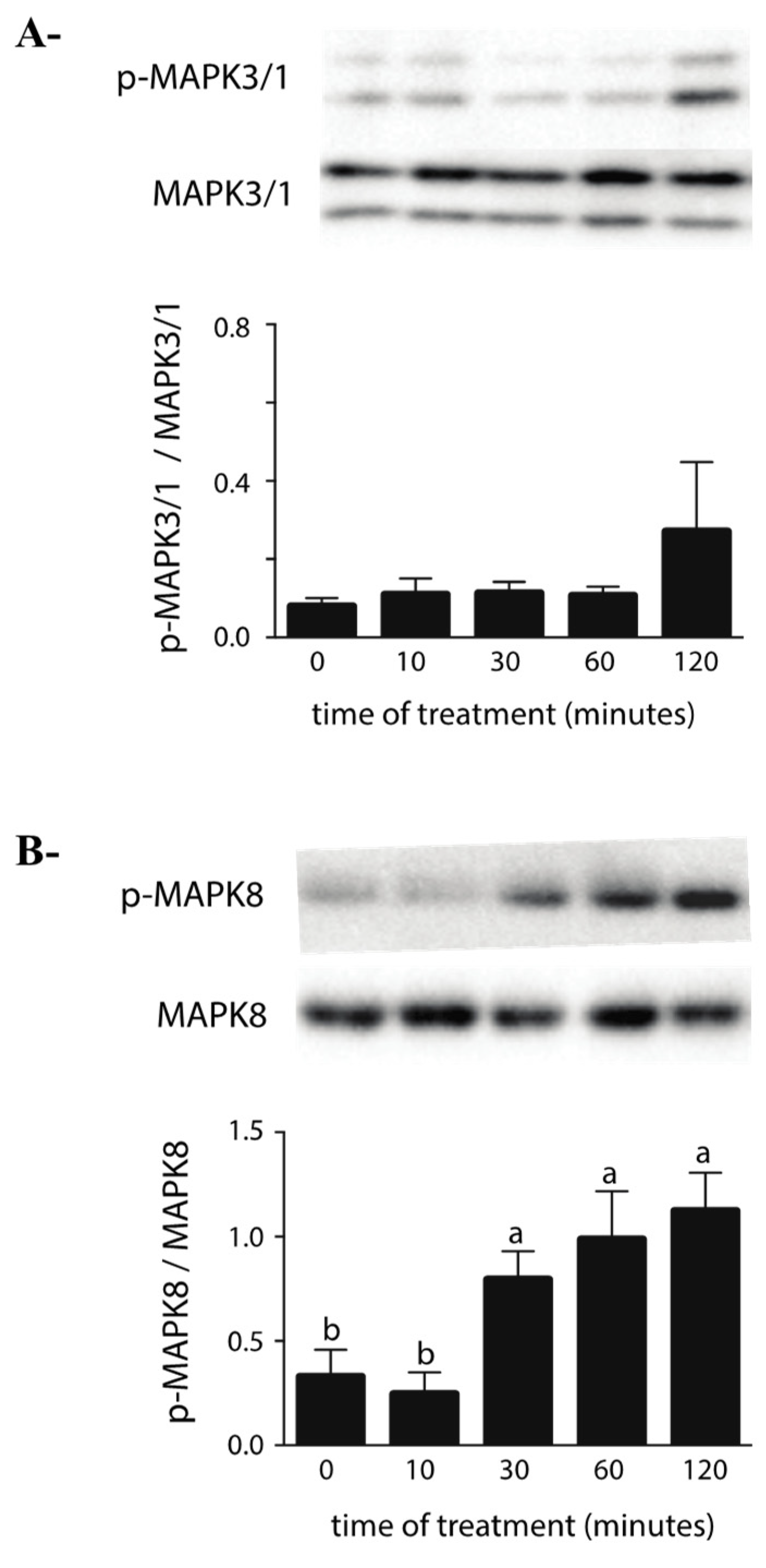
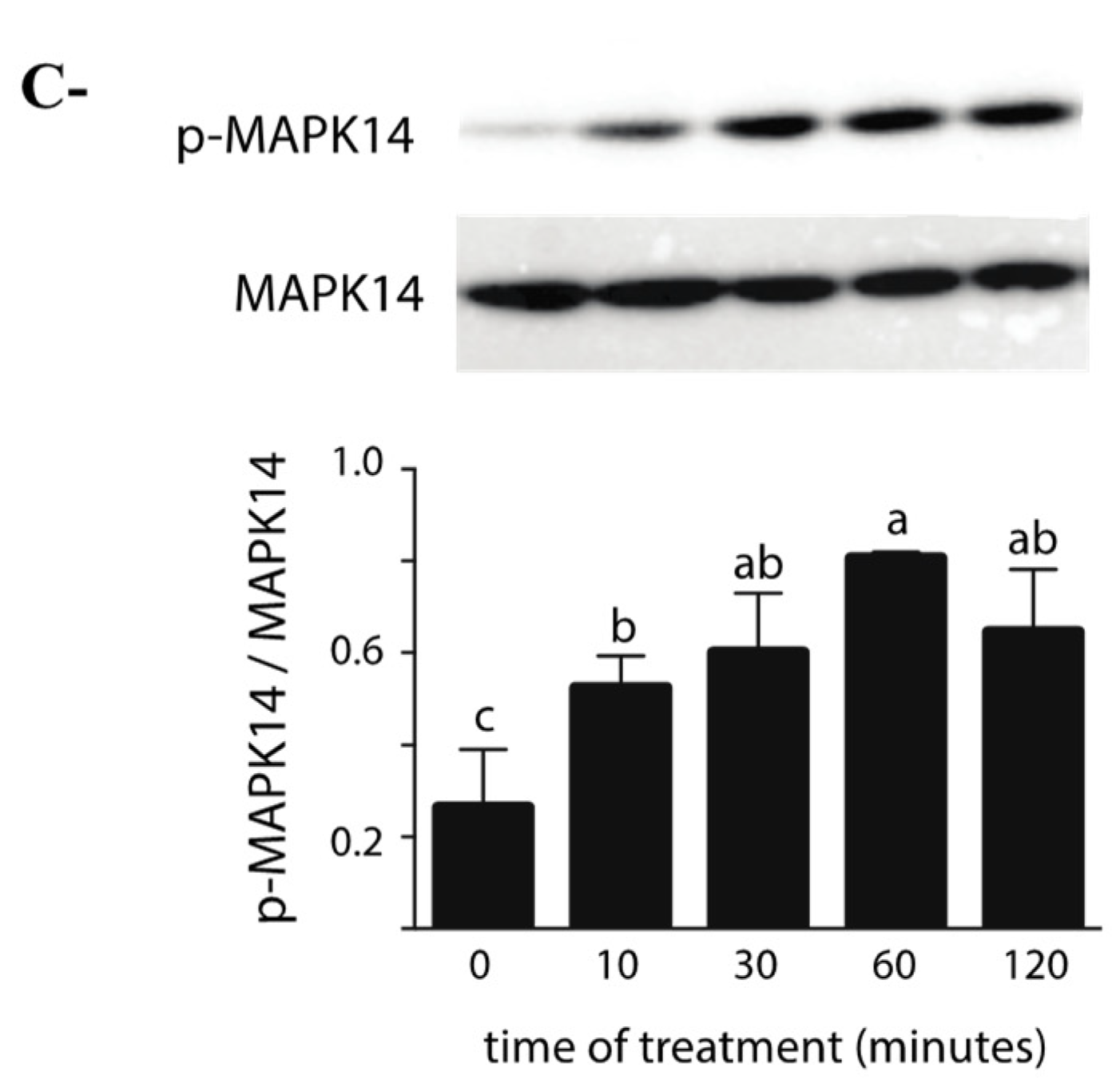
2.2. Effects of ZEA on the MAPK Pathway
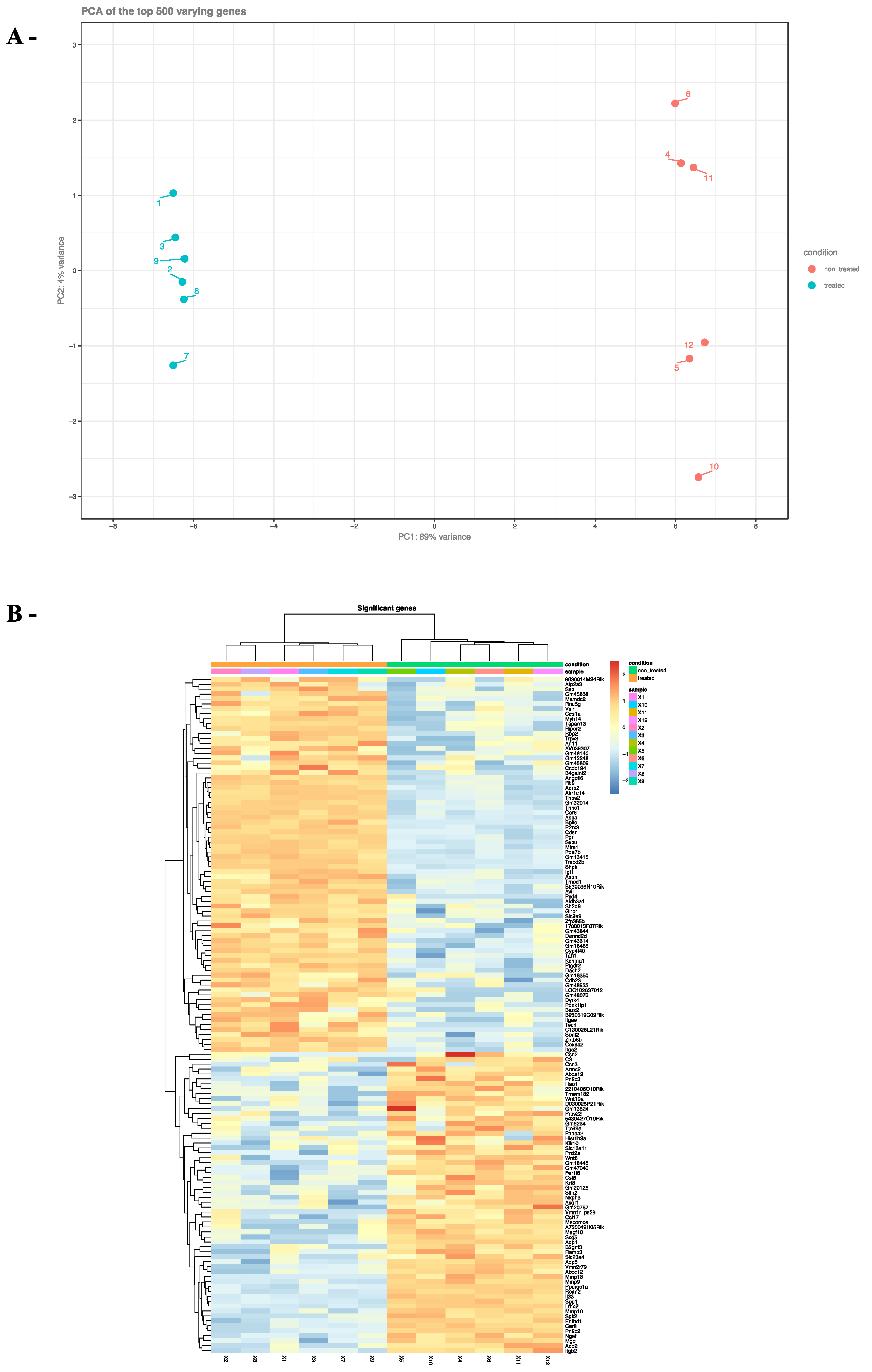
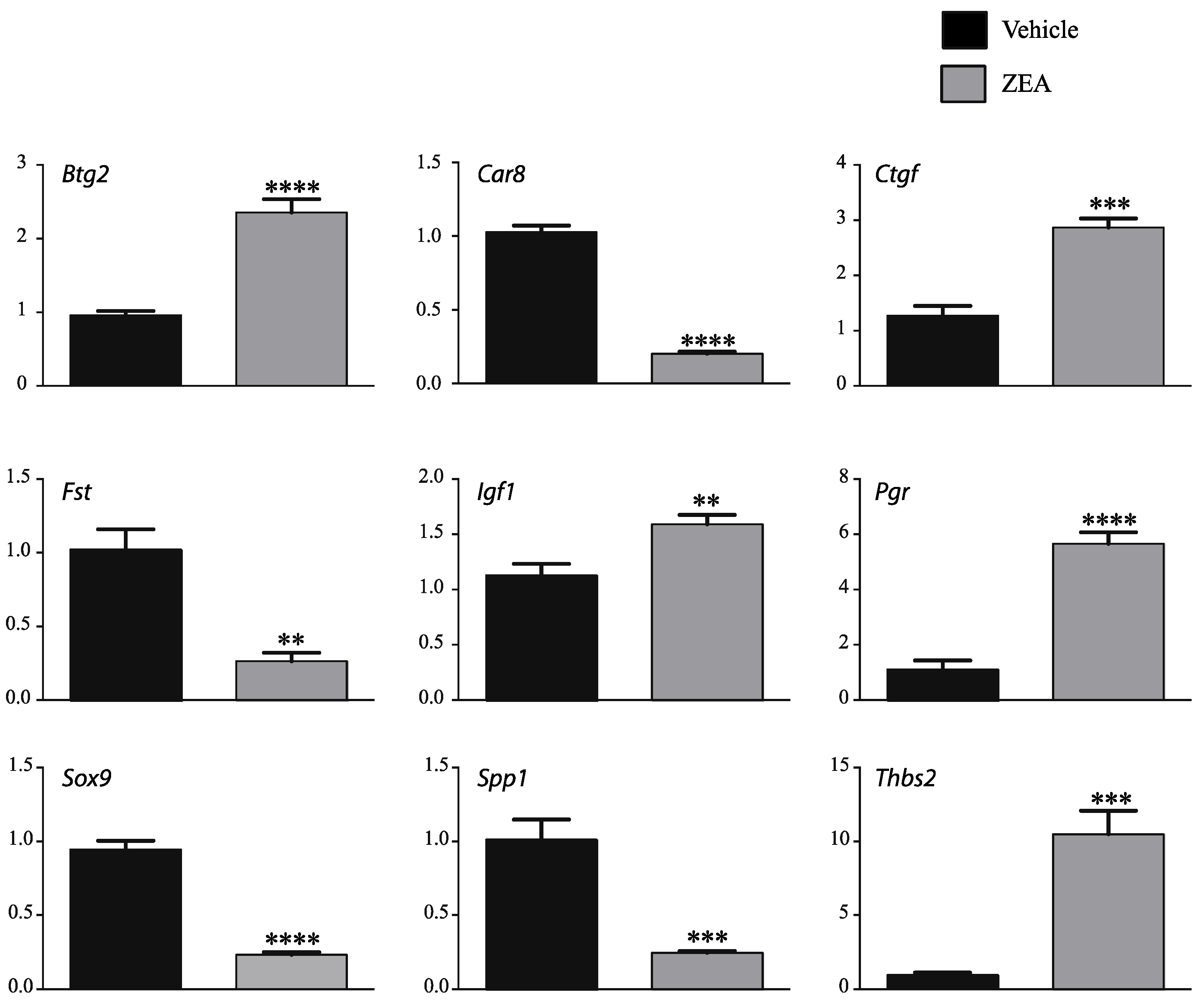
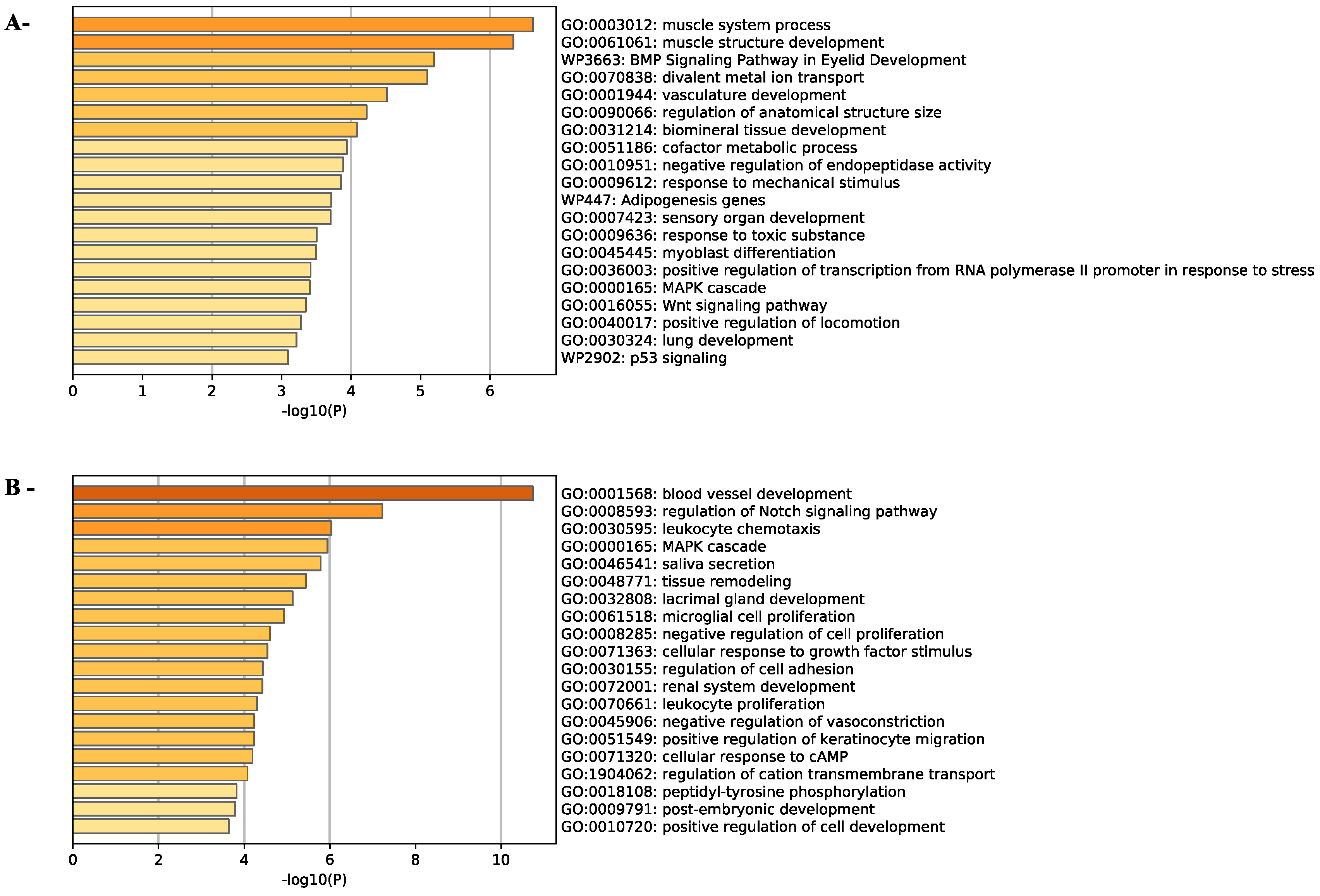
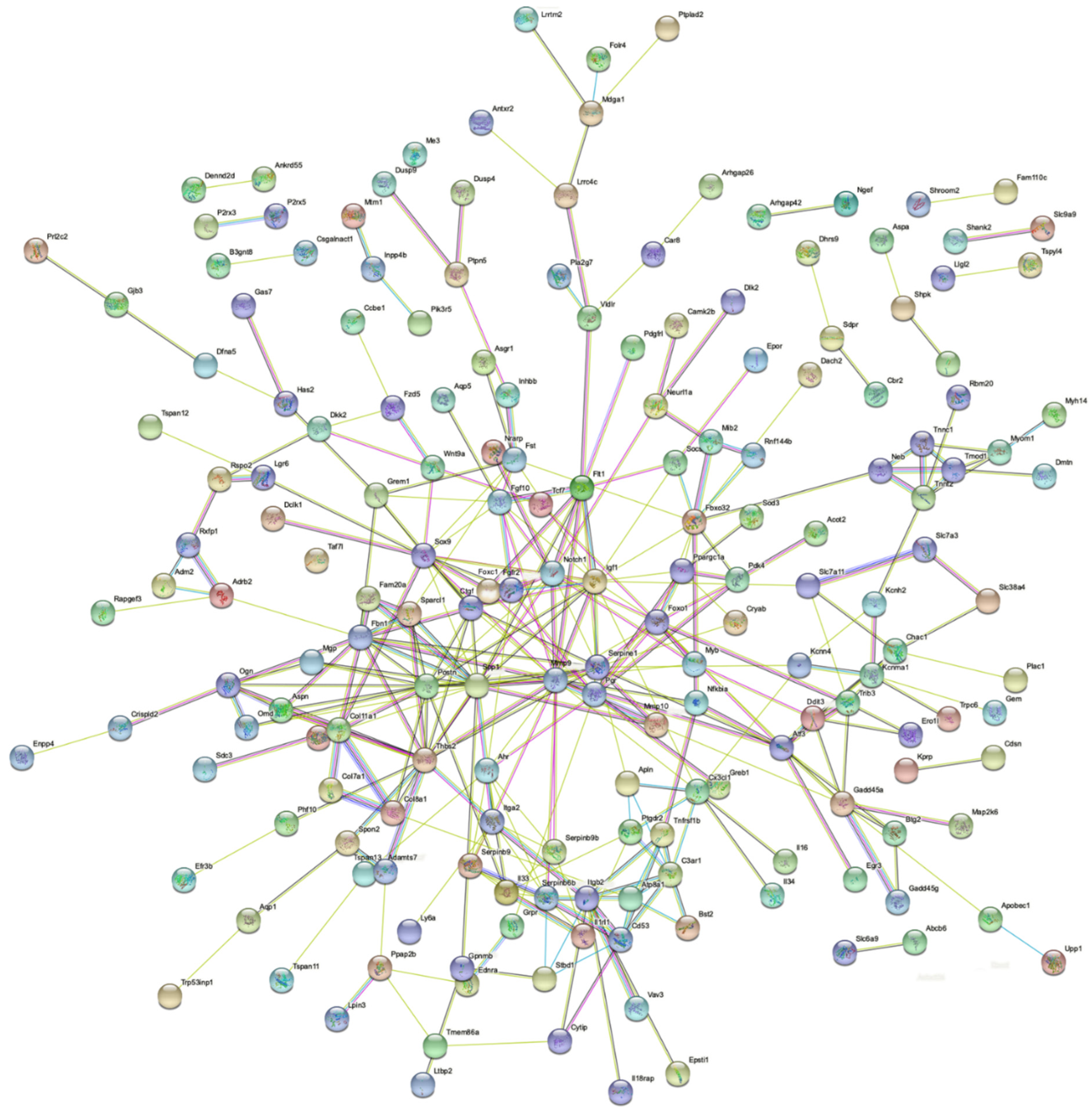
2.3. Effects of ZEA on TM4 Cell Transcriptome
3. Discussion
4. Materials and Methods
4.1. Cells
4.2. Determination of Viable Proliferating Cells
4.3. Western Blot Analysis
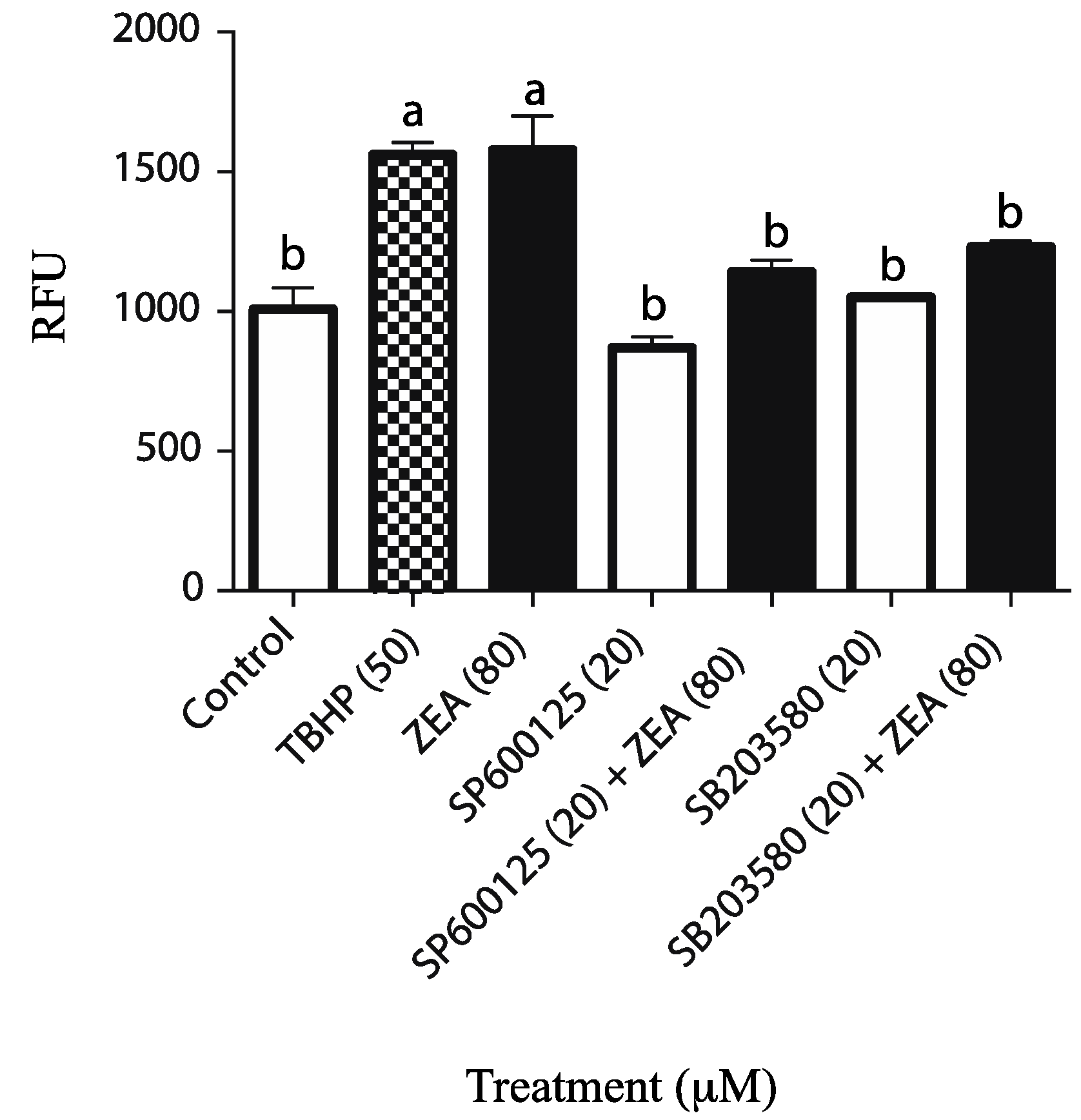
4.4. Detection of Cellular ROS
4.5. RNAseq and RT-qPCR Analyses
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Rotgers, E.; Jørgensen, A.; Yao, H.H. At the Crossroads of Fate-Somatic Cell Lineage Specification in the Fetal Gonad. Endocr. Rev. 2018, 39, 739–759. [Google Scholar] [CrossRef] [PubMed]
- Rebourcet, D.; Darbey, A.; Monteiro, A.; Soffientini, U.; Tsai, Y.T.; Handel, I.; Pitetti, J.L.; Nef, S.; Smith, L.B.; O’Shaughnessy, P.J. Sertoli Cell Number Defines and Predicts Germ and Leydig Cell Population Sizes in the Adult Mouse Testis. Endocrinology 2017, 158, 2955–2969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- França, L.R.; Hess, R.A.; Dufour, J.M.; Hofmann, M.C.; Griswold, M.D. The Sertoli cell: One hundred fifty years of beauty and plasticity. Andrology 2016, 4, 189–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucas, T.F.G.; Lazari, M.F.M.; Porto, C.S. Differential role of the estrogen receptors ESR1 and ESR2 on the regulation of proteins involved with proliferation and differentiation of Sertoli cells from 15-day-old rats. Mol. Cell. Endocrinol. 2014, 382, 84–96. [Google Scholar] [CrossRef] [PubMed]
- Macheroni, C.; Lucas, T.F.G.; Porto, C.S. The role of estrogen receptors in rat Sertoli cells at different stages of development. Heliyon 2020, 6, e05363. [Google Scholar] [CrossRef]
- Sinkevicius, K.W.; Laine, M.; Lotan, T.L.; Woloszyn, K.; Richburg, J.H.; Greene, G.L. Estrogen-dependent and -independent estrogen receptor-alpha signaling separately regulate male fertility. Endocrinology 2009, 150, 2898–2905. [Google Scholar] [CrossRef] [PubMed]
- Royer, C.; Lucas, T.F.; Lazari, M.F.; Porto, C.S. 17Beta-estradiol signaling and regulation of proliferation and apoptosis of rat Sertoli cells. Biol. Reprod. 2012, 86, 108. [Google Scholar] [CrossRef]
- Massart, F.; Saggese, G. Oestrogenic mycotoxin exposures and precocious pubertal development. Int. J. Androl. 2010, 33, 369–376. [Google Scholar] [CrossRef]
- Kuiper, G.G.; Lemmen, J.G.; Carlsson, B.; Corton, J.C.; Safe, S.H.; van der Saag, P.T.; van der Burg, B.; Gustafsson, J.A. Interaction of estrogenic chemicals and phytoestrogens with estrogen receptor beta. Endocrinology 1998, 139, 4252–4263. [Google Scholar] [CrossRef]
- Lo, E.K.K.; Lee, J.C.; Turner, P.C.; El-Nezami, H. Low dose of zearalenone elevated colon cancer cell growth through G protein-coupled estrogenic receptor. Sci. Rep. 2021, 11, 7403. [Google Scholar] [CrossRef]
- Takemura, H.; Shim, J.Y.; Sayama, K.; Tsubura, A.; Zhu, B.T.; Shimoi, K. Characterization of the estrogenic activities of zearalenone and zeranol in vivo and in vitro. J. Steroid Biochem. Mol. Biol. 2007, 103, 170–177. [Google Scholar] [CrossRef]
- Mueller, S.O.; Simon, S.; Chae, K.; Metzler, M.; Korach, K.S. Phytoestrogens and their human metabolites show distinct agonistic and antagonistic properties on estrogen receptor alpha (ERalpha) and ERbeta in human cells. Toxicol. Sci. 2004, 80, 14–25. [Google Scholar] [CrossRef] [Green Version]
- Jee, Y.; Noh, E.M.; Cho, E.S.; Son, H.Y. Involvement of the Fas and Fas ligand in testicular germ cell apoptosis by zearalenone in rat. J. Vet. Sci. 2010, 11, 115–119. [Google Scholar] [CrossRef] [Green Version]
- Kim, I.H.; Son, H.Y.; Cho, S.W.; Ha, C.S.; Kang, B.H. Zearalenone induces male germ cell apoptosis in rats. Toxicol. Lett. 2003, 138, 185–192. [Google Scholar] [CrossRef]
- Yang, J.Y.; Zhang, Y.F.; Wang, Y.Q.; Cui, S. Toxic effects of zearalenone and alpha-zearalenol on the regulation of steroidogenesis and testosterone production in mouse Leydig cells. Toxicol. Vitr. 2007, 21, 558–565. [Google Scholar] [CrossRef]
- Koraichi, F.; Inoubli, L.; Lakhdari, N.; Meunier, L.; Vega, A.; Mauduit, C.; Benahmed, M.; Prouillac, C.; Lecoeur, S. Neonatal exposure to zearalenone induces long term modulation of ABC transporter expression in testis. Toxicology 2013, 310, 29–38. [Google Scholar] [CrossRef]
- Wang, Y.; Zheng, W.; Bian, X.; Yuan, Y.; Gu, J.; Liu, X.; Liu, Z.; Bian, J. Zearalenone induces apoptosis and cytoprotective autophagy in primary Leydig cells. Toxicol. Lett. 2014, 226, 182–191. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, B.; Huang, K.; He, X.; Luo, Y.; Liang, R.; Luo, H.; Shen, X.L.; Xu, W. Mitochondrial proteomic analysis reveals the molecular mechanisms underlying reproductive toxicity of zearalenone in MLTC-1 cells. Toxicology 2014, 324, 55–67. [Google Scholar] [CrossRef]
- Lin, P.; Chen, F.; Sun, J.; Zhou, J.; Wang, X.; Wang, N.; Li, X.; Zhang, Z.; Wang, A.; Jin, Y. Mycotoxin zearalenone induces apoptosis in mouse Leydig cells via an endoplasmic reticulum stress-dependent signalling pathway. Reprod. Toxicol. 2015, 52, 71–77. [Google Scholar] [CrossRef]
- Zatecka, E.; Ded, L.; Elzeinova, F.; Kubatova, A.; Dorosh, A.; Margaryan, H.; Dostalova, P.; Korenkova, V.; Hoskova, K.; Peknicova, J. Effect of zearalenone on reproductive parameters and expression of selected testicular genes in mice. Reprod. Toxicol. 2014, 45, 20–30. [Google Scholar] [CrossRef]
- Zheng, W.; Wang, B.; Si, M.; Zou, H.; Song, R.; Gu, J.; Yuan, Y.; Liu, X.; Zhu, G.; Bai, J.; et al. Zearalenone altered the cytoskeletal structure via ER stress- autophagy- oxidative stress pathway in mouse TM4 Sertoli cells. Sci. Rep. 2018, 8, 3320. [Google Scholar] [CrossRef]
- Zheng, W.L.; Wang, B.J.; Wang, L.; Shan, Y.P.; Zou, H.; Song, R.L.; Wang, T.; Gu, J.H.; Yuan, Y.; Liu, X.Z.; et al. ROS-Mediated Cell Cycle Arrest and Apoptosis Induced by Zearalenone in Mouse Sertoli Cells via ER Stress and the ATP/AMPK Pathway. Toxins 2018, 10, 24. [Google Scholar] [CrossRef] [Green Version]
- Feng, N.; Wang, B.; Cai, P.; Zheng, W.; Zou, H.; Gu, J.; Yuan, Y.; Liu, X.; Liu, Z.; Bian, J. ZEA-induced autophagy in TM4 cells was mediated by the release of Ca(2+) activates CaMKKβ-AMPK signaling pathway in the endoplasmic reticulum. Toxicol. Lett. 2020, 323, 1–9. [Google Scholar] [CrossRef]
- Mather, J.P. Establishment and characterization of two distinct mouse testicular epithelial cell lines. Biol. Reprod. 1980, 23, 243–252. [Google Scholar]
- Long, M.; Yang, S.H.; Shi, W.; Li, P.; Guo, Y.; Guo, J.; He, J.B.; Zhang, Y. Protective effect of proanthocyanidin on mice Sertoli cell apoptosis induced by zearalenone via the Nrf2/ARE signalling pathway. Environ. Sci. Pollut. Res. Int. 2017, 24, 26724–26733. [Google Scholar] [CrossRef]
- Yu, J.Y.; Zheng, Z.H.; Son, Y.O.; Shi, X.; Jang, Y.O.; Lee, J.C. Mycotoxin zearalenone induces AIF- and ROS-mediated cell death through p53- and MAPK-dependent signaling pathways in RAW264.7 macrophages. Toxic. Vitr. 2011, 25, 1654–1663. [Google Scholar] [CrossRef]
- Paruthiyil, S.; Cvoro, A.; Tagliaferri, M.; Cohen, I.; Shtivelman, E.; Leitman, D.C. Estrogen receptor β causes a G2 cell cycle arrest by inhibiting CDK1 activity through the regulation of cyclin B1, GADD45A, and BTG2. Breast Cancer Res. Treat. 2011, 129, 777–784. [Google Scholar] [CrossRef]
- Greathouse, K.L.; Cook, J.D.; Lin, K.; Davis, B.J.; Berry, T.D.; Bredfeldt, T.G.; Walker, C.L. Identification of uterine leiomyoma genes developmentally reprogrammed by neonatal exposure to diethylstilbestrol. Reprod. Sci. 2008, 15, 765–778. [Google Scholar] [CrossRef]
- Greathouse, K.L.; Bredfeldt, T.; Everitt, J.I.; Lin, K.; Berry, T.; Kannan, K.; Mittelstadt, M.L.; Ho, S.M.; Walker, C.L. Environmental estrogens differentially engage the histone methyltransferase EZH2 to increase risk of uterine tumorigenesis. Mol. Cancer Res. 2012, 10, 546–557. [Google Scholar] [CrossRef] [Green Version]
- Ogo, Y.; Taniuchi, S.; Ojima, F.; Hayashi, S.; Murakami, I.; Saito, Y.; Takeuchi, S.; Kudo, T.; Takahashi, S. IGF-1 gene expression is differentially regulated by estrogen receptors α and β in mouse endometrial stromal cells and ovarian granulosa cells. J. Reprod. Dev. 2014, 60, 216–223. [Google Scholar] [CrossRef] [Green Version]
- Umayahara, Y.; Kawamori, R.; Watada, H.; Imano, E.; Iwama, N.; Morishima, T.; Yamasaki, Y.; Kajimoto, Y.; Kamada, T. Estrogen regulation of the insulin-like growth factor I gene transcription involves an AP-1 enhancer. J. Biol. Chem. 1994, 269, 16433–16442. [Google Scholar] [CrossRef]
- Hewitt, S.C.; Li, Y.; Li, L.; Korach, K.S. Estrogen-mediated regulation of Igf1 transcription and uterine growth involves direct binding of estrogen receptor alpha to estrogen-responsive elements. J. Biol. Chem. 2010, 285, 2676–2685. [Google Scholar] [CrossRef] [Green Version]
- Petz, L.N.; Ziegler, Y.S.; Loven, M.A.; Nardulli, A.M. Estrogen receptor alpha and activating protein-1 mediate estrogen responsiveness of the progesterone receptor gene in MCF-7 breast cancer cells. Endocrinology 2002, 143, 4583–4591. [Google Scholar] [CrossRef]
- Petz, L.N.; Ziegler, Y.S.; Schultz, J.R.; Kim, H.; Kemper, J.K.; Nardulli, A.M. Differential regulation of the human progesterone receptor gene through an estrogen response element half site and Sp1 sites. J. Steroid Biochem. Mol. Biol. 2004, 88, 113–122. [Google Scholar] [CrossRef]
- Barske, L.A.; Capel, B. Estrogen represses SOX9 during sex determination in the red-eared slider turtle Trachemys scripta. Dev. Biol. 2010, 341, 305–314. [Google Scholar] [CrossRef]
- Stewart, M.K.; Mattiske, D.M.; Pask, A.J. Estrogen suppresses SOX9 and activates markers of female development in a human testis-derived cell line. BMC Mol. Cell Biol. 2020, 21, 66. [Google Scholar] [CrossRef]
- Stewart, M.K.; Mattiske, D.M.; Pask, A.J. Exogenous Oestrogen Impacts Cell Fate Decision in the Developing Gonads: A Potential Cause of Declining Human Reproductive Health. Int. J. Mol. Sci. 2020, 21, 8377. [Google Scholar] [CrossRef]
- Slavin, S.; Yeh, C.R.; Da, J.; Yu, S.; Miyamoto, H.; Messing, E.M.; Guancial, E.; Yeh, S. Estrogen receptor α in cancer-associated fibroblasts suppresses prostate cancer invasion via modulation of thrombospondin 2 and matrix metalloproteinase 3. Carcinogenesis 2014, 35, 1301–1309. [Google Scholar] [CrossRef] [Green Version]
- Cheng, L.; Yang, Z.; Wang, X.; Jiao, Y.; Xie, X.; Lin, J.; Zhang, H.; Han, J.; Jiang, K.; Ye, Q. Suppression of estrogen receptor transcriptional activity by connective tissue growth factor. PLoS ONE 2011, 6, e20028. [Google Scholar] [CrossRef]
- Barakat, B.; O’Connor, A.E.; Gold, E.; de Kretser, D.M.; Loveland, K.L. Inhibin, activin, follistatin and FSH serum levels and testicular production are highly modulated during the first spermatogenic wave in mice. Reproduction 2008, 136, 345–359. [Google Scholar] [CrossRef] [Green Version]
- Rotgers, E.; Cisneros-Montalvo, S.; Nurmio, M.; Toppari, J. Retinoblastoma protein represses E2F3 to maintain Sertoli cell quiescence in mouse testis. J. Cell Sci. 2019, 132, jcs229849. [Google Scholar] [CrossRef] [Green Version]
- Pitetti, J.L.; Calvel, P.; Zimmermann, C.; Conne, B.; Papaioannou, M.D.; Aubry, F.; Cederroth, C.R.; Urner, F.; Fumel, B.; Crausaz, M.; et al. An essential role for insulin and IGF1 receptors in regulating sertoli cell proliferation, testis size, and FSH action in mice. Mol. Endocrinol. 2013, 27, 814–827. [Google Scholar] [CrossRef]
- Chaboissier, M.C.; Kobayashi, A.; Vidal, V.I.; Lützkendorf, S.; van de Kant, H.J.; Wegner, M.; de Rooij, D.G.; Behringer, R.R.; Schedl, A. Functional analysis of Sox8 and Sox9 during sex determination in the mouse. Development 2004, 131, 1891–1901. [Google Scholar] [CrossRef] [Green Version]
- Barrionuevo, F.; Bagheri-Fam, S.; Klattig, J.; Kist, R.; Taketo, M.M.; Englert, C.; Scherer, G. Homozygous inactivation of Sox9 causes complete XY sex reversal in mice. Biol. Reprod. 2006, 74, 195–201. [Google Scholar] [CrossRef]
- Lue, Y.; Wang, C.; Lydon, J.P.; Leung, A.; Li, J.; Swerdloff, R.S. Functional role of progestin and the progesterone receptor in the suppression of spermatogenesis in rodents. Andrology 2013, 1, 308–317. [Google Scholar] [CrossRef]
- Tomooka, Y.; Yasui, T. Electron microscopic study of the response of the adrenocortical X-zone in mice treated with sex steroids. Cell Tissue Res. 1978, 194, 269–277. [Google Scholar] [CrossRef]
- Yue, J.; López, J.M. Understanding MAPK Signaling Pathways in Apoptosis. Int. J. Mol. Sci. 2020, 21, 2346. [Google Scholar] [CrossRef] [Green Version]
- Cai, G.; Sun, K.; Xia, S.; Feng, Z.; Zou, H.; Gu, J.; Yuan, Y.; Zhu, J.; Liu, Z.; Bian, J. Decrease in immune function and the role of mitogen-activated protein kinase (MAPK) overactivation in apoptosis during T lymphocytes activation induced by zearalenone, deoxynivalenol, and their combinations. Chemosphere 2020, 255, 126999. [Google Scholar] [CrossRef]
- Hu, K.; Gong, X.; Ai, Q.; Lin, L.; Dai, J.; Cai, L.; Jiang, R.; Ge, P.; Zhang, L. Endogenous AMPK acts as a detrimental factor in fulminant hepatitis via potentiating JNK-dependent hepatocyte apoptosis. Cell Death Dis. 2017, 8, e2637. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Miller, E.J.; Ninomiya-Tsuji, J.; Russell, R.R.; Young, L.H., 3rd. AMP-activated protein kinase activates p38 mitogen-activated protein kinase by increasing recruitment of p38 MAPK to TAB1 in the ischemic heart. Circ. Res. 2005, 97, 872–879. [Google Scholar] [CrossRef] [Green Version]
- Lanna, A.; Henson, S.M.; Escors, D.; Akbar, A.N. The kinase p38 activated by the metabolic regulator AMPK and scaffold TAB1 drives the senescence of human T cells. Nat. Immunol. 2014, 15, 965–972. [Google Scholar] [CrossRef] [Green Version]
- Kim, G.T.; Lee, S.H.; Kim, J.I.; Kim, Y.M. Quercetin regulates the sestrin 2-AMPK-p38 MAPK signaling pathway and induces apoptosis by increasing the generation of intracellular ROS in a p53-independent manner. Int. J. Mol. Med. 2014, 33, 863–869. [Google Scholar] [CrossRef] [Green Version]
- Andersson, Y.; Le, H.; Juell, S.; Fodstad, Ø. AMP-activated protein kinase protects against anti-epidermal growth factor receptor-Pseudomonas exotoxin A immunotoxin-induced MA11 breast cancer cell death. Mol. Cancer Ther. 2006, 5, 1050–1059. [Google Scholar] [CrossRef] [Green Version]
- Sui, X.; Kong, N.; Ye, L.; Han, W.; Zhou, J.; Zhang, Q.; He, C.; Pan, H. p38 and JNK MAPK pathways control the balance of apoptosis and autophagy in response to chemotherapeutic agents. Cancer Lett. 2014, 344, 174–179. [Google Scholar] [CrossRef]
- Wu, L.; Dong, H.; Zhao, J.; Wang, Y.; Yang, Q.; Jia, C.; Ma, J. Diosgenin stimulates rat TM4 cell proliferation through activating plasma membrane translocation and transcriptional activity of estrogen receptors. Biol. Reprod. 2015, 92, 24. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Lei, Z. Chromatin Immunoprecipitation with Estrogen Receptor 1 and the Promoter of Greb1 in TM4 Sertoli Cells. Methods Mol. Biol. 2016, 1366, 67–77. [Google Scholar]
- Kim, Y.; Bingham, N.; Sekido, R.; Parker, K.L.; Lovell-Badge, R.; Capel, B. Fibroblast growth factor receptor 2 regulates proliferation and Sertoli differentiation during male sex determination. Proc. Natl. Acad. Sci. USA 2007, 104, 16558–16563. [Google Scholar] [CrossRef] [Green Version]
- Bagheri-Fam, S.; Sim, H.; Bernard, P.; Jayakody, I.; Taketo, M.M.; Scherer, G.; Harley, V.R. Loss of Fgfr2 leads to partial XY sex reversal. Dev. Biol. 2008, 314, 71–83. [Google Scholar] [CrossRef] [Green Version]
- Nef, S.; Verma-Kurvari, S.; Merenmies, J.; Vassalli, J.D.; Efstratiadis, A.; Accili, D.; Parada, L.F. Testis determination requires. Nature 2003, 426, 291–295. [Google Scholar]
- Schmahl, J.; Kim, Y.; Colvin, J.S.; Ornitz, D.M.; Capel, B. Fgf9 induces proliferation and nuclear localization of FGFR2 in Sertoli precursors during male sex determination. Development 2004, 131, 3627–3636. [Google Scholar] [CrossRef] [Green Version]
- Schmahl, J.; Capel, B. Cell proliferation is necessary for the determination of male fate in the gonad. Dev. Biol. 2003, 258, 264–276. [Google Scholar] [CrossRef] [Green Version]
- Garcia, T.X.; Hofmann, M.C. NOTCH signaling in Sertoli cells regulates gonocyte fate. Cell Cycle 2013, 12, 2538–2545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, T.X.; DeFalco, T.; Capel, B.; Hofmann, M.C. Constitutive activation of NOTCH1 signaling in Sertoli cells causes gonocyte exit from quiescence. Dev. Biol. 2013, 377, 188–201. [Google Scholar] [CrossRef] [Green Version]
- Piprek, R.P.; Kolasa, M.; Podkowa, D.; Kloc, M.; Kubiak, J.Z. Transcriptional profiling validates involvement of extracellular matrix and proteinases genes in mouse gonad development. Mech. Dev. 2018, 149, 9–19. [Google Scholar] [CrossRef]
- Matoba, S.; Hiramatsu, R.; Kanai-Azuma, M.; Tsunekawa, N.; Harikae, K.; Kawakami, H.; Kurohmaru, M.; Kanai, Y. Establishment of testis-specific SOX9 activation requires high-glucose metabolism in mouse sex differentiation. Dev. Biol. 2008, 324, 76–87. [Google Scholar] [CrossRef] [Green Version]
- Cool, J.; DeFalco, T.; Capel, B. Testis formation in the fetal mouse: Dynamic and complex de novo tubulogenesis. Wiley Interdiscip. Rev. Dev. Biol. 2012, 1, 847–859. [Google Scholar] [CrossRef]
- Delbes, G.; Levacher, C.; Habert, R. Estrogen effects on fetal and neonatal testicular development. Reproduction 2006, 132, 527–538. [Google Scholar] [CrossRef] [Green Version]
- Skakkebaek, N.E.; Rajpert-De Meyts, E.; Buck Louis, G.M.; Toppari, J.; Andersson, A.M.; Eisenberg, M.L.; Jensen, T.K.; Jørgensen, N.; Swan, S.H.; Sapra, K.J.; et al. Male Reproductive Disorders and Fertility Trends: Influences of Environment and Genetic Susceptibility. Physiol. Rev. 2016, 96, 55–97. [Google Scholar] [CrossRef] [Green Version]
- Shima, Y.; Miyabayashi, K.; Haraguchi, S.; Arakawa, T.; Otake, H.; Baba, T.; Matsuzaki, S.; Shishido, Y.; Akiyama, H.; Tachibana, T.; et al. Contribution of Leydig and Sertoli cells to testosterone production in mouse fetal testes. Mol. Endocrinol. 2013, 27, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Basciani, S.; Mariani, S.; Spera, G.; Gnessi, L. Role of platelet-derived growth factors in the testis. Endocr. Rev. 2010, 31, 916–939. [Google Scholar] [CrossRef] [Green Version]
- Yao, H.H.; Whoriskey, W.; Capel, B. Desert Hedgehog/Patched 1 signaling specifies fetal Leydig cell fate in testis organogenesis. Genes. Dev. 2002, 16, 1433–1440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic. Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Fold-Change | Padj |
|---|---|---|
| tgdr2 | 13.22 | 1.10 × 10−13 |
| Kcnma1 | 13.19 | 4.57 × 10−13 |
| Avil | 12.45 | 2.17 × 10−14 |
| Car6 | 10.74 | 6.92 × 10−35 |
| Slc9a9 | 10.18 | 1.55 × 10−09 |
| Shpk | 10.16 | 5.42 × 10−148 |
| Myh14 | 9.19 | 3.55 × 10−22 |
| Taf7l | 8.40 | 1.56 × 10−15 |
| Thbs2 | 8.03 | 2.73 × 10−72 |
| Gm32014 | 7.98 | 3.04 × 10−63 |
| Ripor2 | 7.78 | 2.20 × 10−27 |
| Gm43314 | 7.47 | 5.40 × 10−11 |
| Akr1c14 | 7.34 | 2.98 × 10−49 |
| Tnnc1 | 7.14 | 4.15 × 10−65 |
| Itga2 | 7.10 | 7.99 × 10−20 |
| P2rx3 | 6.77 | 2.07 × 10−49 |
| Pde7b | 6.58 | 6.72 × 10−32 |
| Cdsn | 6.53 | 1.39 × 10−112 |
| B930036N10Rik | 6.22 | 2.32 × 10−21 |
| Aspn | 5.95 | 3.09 × 10−10 |
| Aspa | 5.93 | 1.16 × 10−132 |
| Gm13415 | 5.75 | 8.28 × 10−26 |
| Zbtb8b | 5.60 | 2.23 × 10−26 |
| Igf1 | 5.58 | 1.90 × 10−33 |
| Trabd2b | 5.57 | 4.33 × 10−146 |
| Dach2 | 5.51 | 1.31 × 10−16 |
| Adrb2 | 5.46 | 9.24 × 10−58 |
| Tspan13 | 5.32 | 2.63 × 10−08 |
| Glrp1 | 4.76 | 4.20 × 10−06 |
| Angptl6 | 4.58 | 3.93 × 10−21 |
| Dennd2d | 4.51 | 1.19 × 10−09 |
| Gm16485 | 4.44 | 1.029 × 10−06 |
| Tmod1 | 4.39 | 2.60 × 10−11 |
| Sybu | 4.38 | 1.80 × 10−65 |
| Pgr | 4.34 | 4.36 × 10−75 |
| Gene | Fold-Change | Padj |
|---|---|---|
| Aqp1 | −26.28 | 9.45 × 10−15 |
| Prl2c2 | −19.87 | 1.87 × 10−69 |
| Fer1l6 | −12.35 | 6.90 × 10−26 |
| Mmp10 | −10.47 | 8.09 × 10−13 |
| Car8 | −6.93 | 6.11 × 10−32 |
| Il33 | −6.72 | 5.82 × 10−168 |
| Ppargc1a | −6.08 | 9.72 × 10−100 |
| Vmn2r79 | −5.94 | 7.08 × 10−23 |
| Scg5 | −5.02 | 1.56 × 10−14 |
| Aqp5 | −4.86 | 7.48 × 10−08 |
| Asgr1 | −4.48 | 5.97 × 10−05 |
| Spp1 | −4.43 | 1.24 × 10−198 |
| Ngef | −4.43 | 5.69 × 10−12 |
| Ltbp2 | −4.42 | 9.61 × 10−125 |
| Itgb2 | −4.38 | 4.24 × 10−11 |
| Enthd1 | −4.31 | 2.07 × 10−07 |
| Gm47040 | −4.30 | 3.48 × 10−08 |
| Mgp | −4.15 | 1.37 × 10−14 |
| Rcan2 | −4.15 | 2.24 × 10−66 |
| Mmp9 | −4.12 | 2.75 × 10−30 |
| Tcim | −3.94 | 3.24 × 10−115 |
| Inpp4b | −3.87 | 5.52 × 10−42 |
| Fst | −3.77 | 3.50 × 10−251 |
| Tmem179 | −3.77 | 3.31 × 10−07 |
| Apln | −3.73 | 1.42 × 10−174 |
| Ahr | −3.72 | 2.16 × 10−15 |
| Tspan11 | −3.60 | 7.52 × 10−117 |
| Neto2 | −3.57 | 9.91 × 10−75 |
| Sox9 | −3.56 | 1.28 × 10−26 |
| Ccdc88c | −3.52 | 1.19 × 10−12 |
| Junos | −3.50 | 5.81 × 10−12 |
| Epsti1 | −3.44 | 1.21 × 10−07 |
| Tmem200a | −3.43 | 7.27 × 10−11 |
| Gas1 | −3.42 | 9.01 × 10−93 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Savard, C.; Gawhary, S.; Boyer, A.; Chorfi, Y. Assessment of Zearalenone-Induced Cell Survival and of Global Gene Regulation in Mouse TM4 Sertoli Cells. Toxins 2022, 14, 98. https://doi.org/10.3390/toxins14020098
Savard C, Gawhary S, Boyer A, Chorfi Y. Assessment of Zearalenone-Induced Cell Survival and of Global Gene Regulation in Mouse TM4 Sertoli Cells. Toxins. 2022; 14(2):98. https://doi.org/10.3390/toxins14020098
Chicago/Turabian StyleSavard, Christian, Sadaf Gawhary, Alexandre Boyer, and Younes Chorfi. 2022. "Assessment of Zearalenone-Induced Cell Survival and of Global Gene Regulation in Mouse TM4 Sertoli Cells" Toxins 14, no. 2: 98. https://doi.org/10.3390/toxins14020098
APA StyleSavard, C., Gawhary, S., Boyer, A., & Chorfi, Y. (2022). Assessment of Zearalenone-Induced Cell Survival and of Global Gene Regulation in Mouse TM4 Sertoli Cells. Toxins, 14(2), 98. https://doi.org/10.3390/toxins14020098