Corepressors SsnF and RcoA Regulate Development and Aflatoxin B1 Biosynthesis in Aspergillus flavus NRRL 3357



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
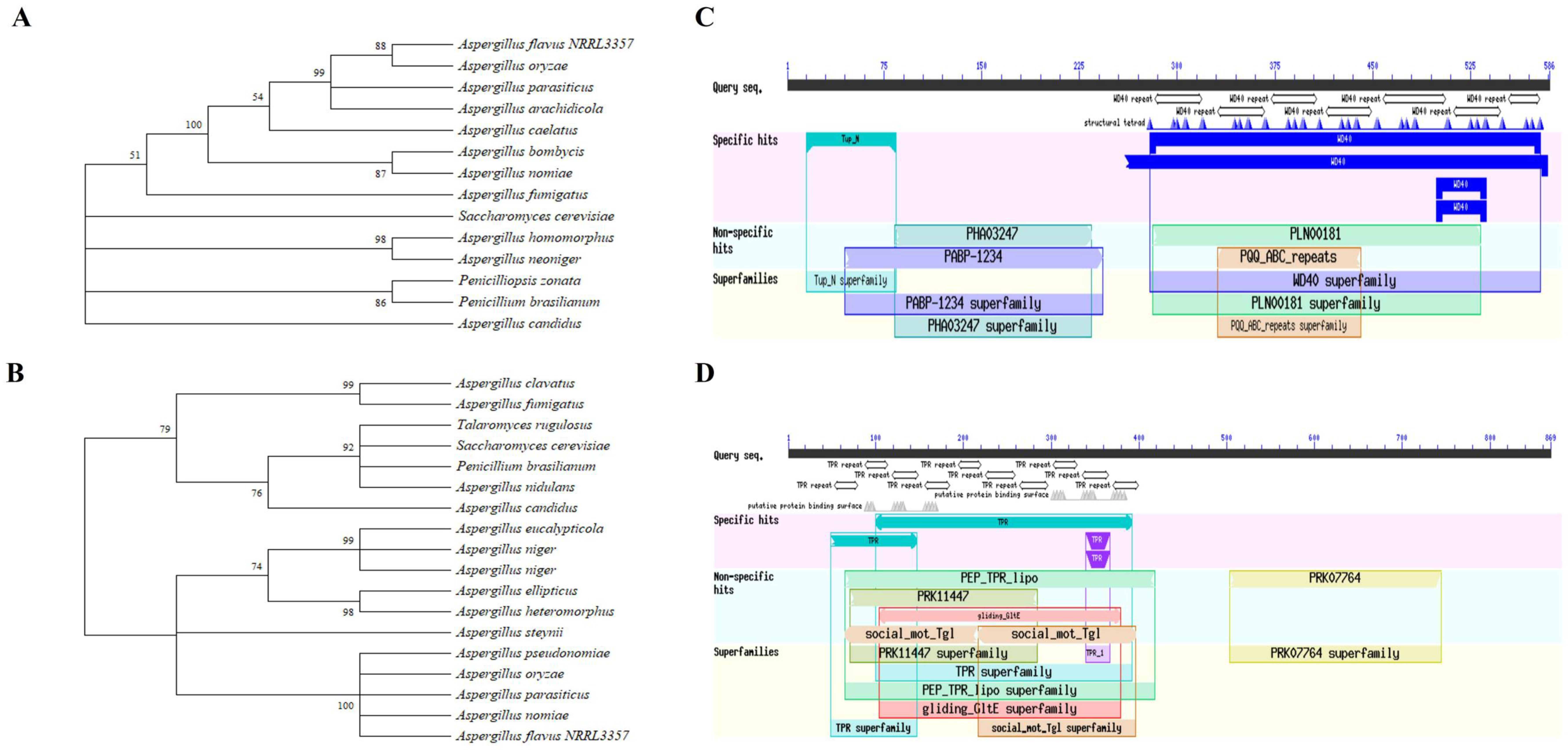
2.1. Bioinformatic Analyses and Deletion Mutant Constructions of SsnF and RcoA in A. flavus
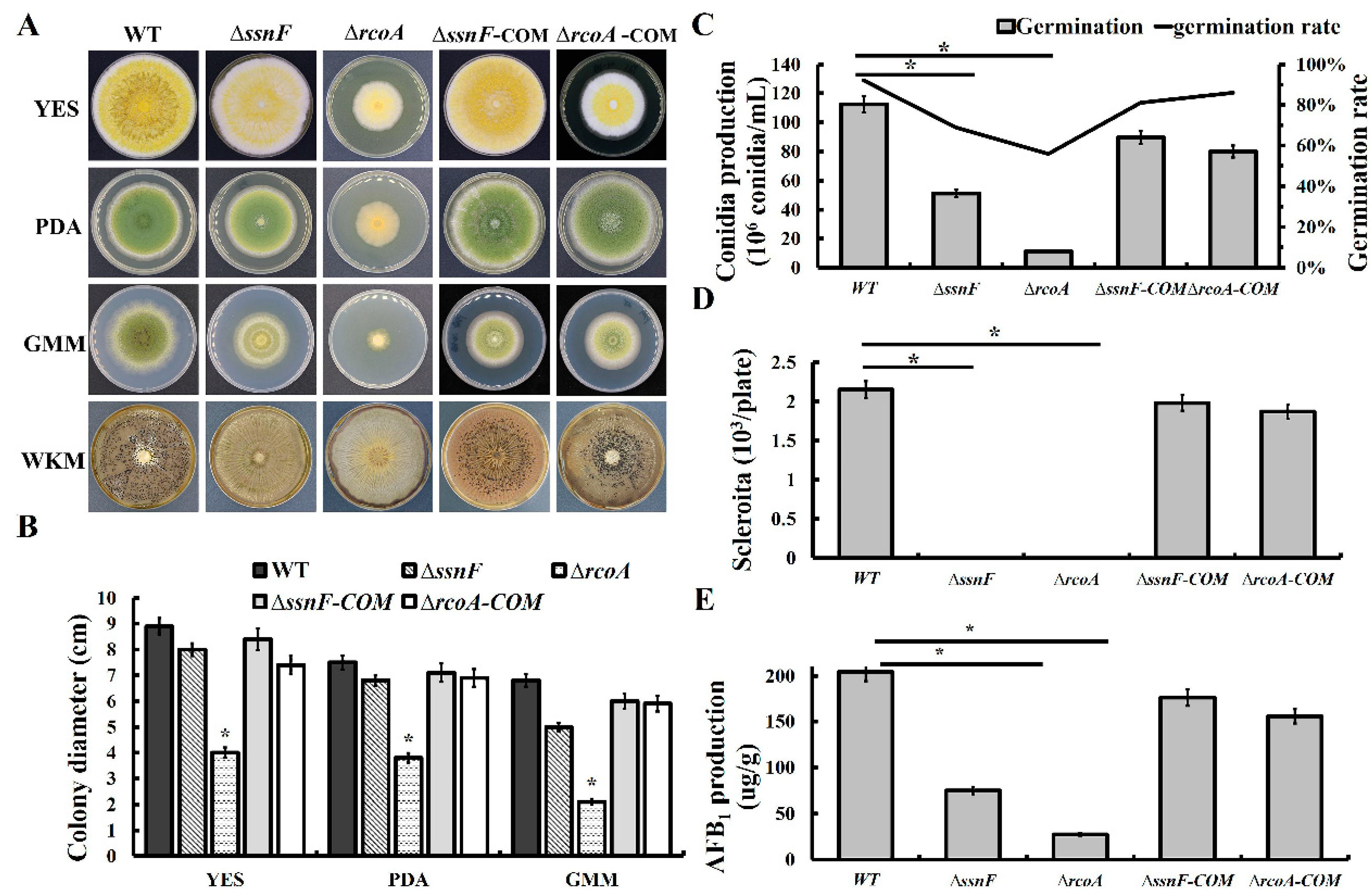
2.2. SsnF and RcoA Are Necessary for Vegetative Growth and Fungal Development in A. flavus
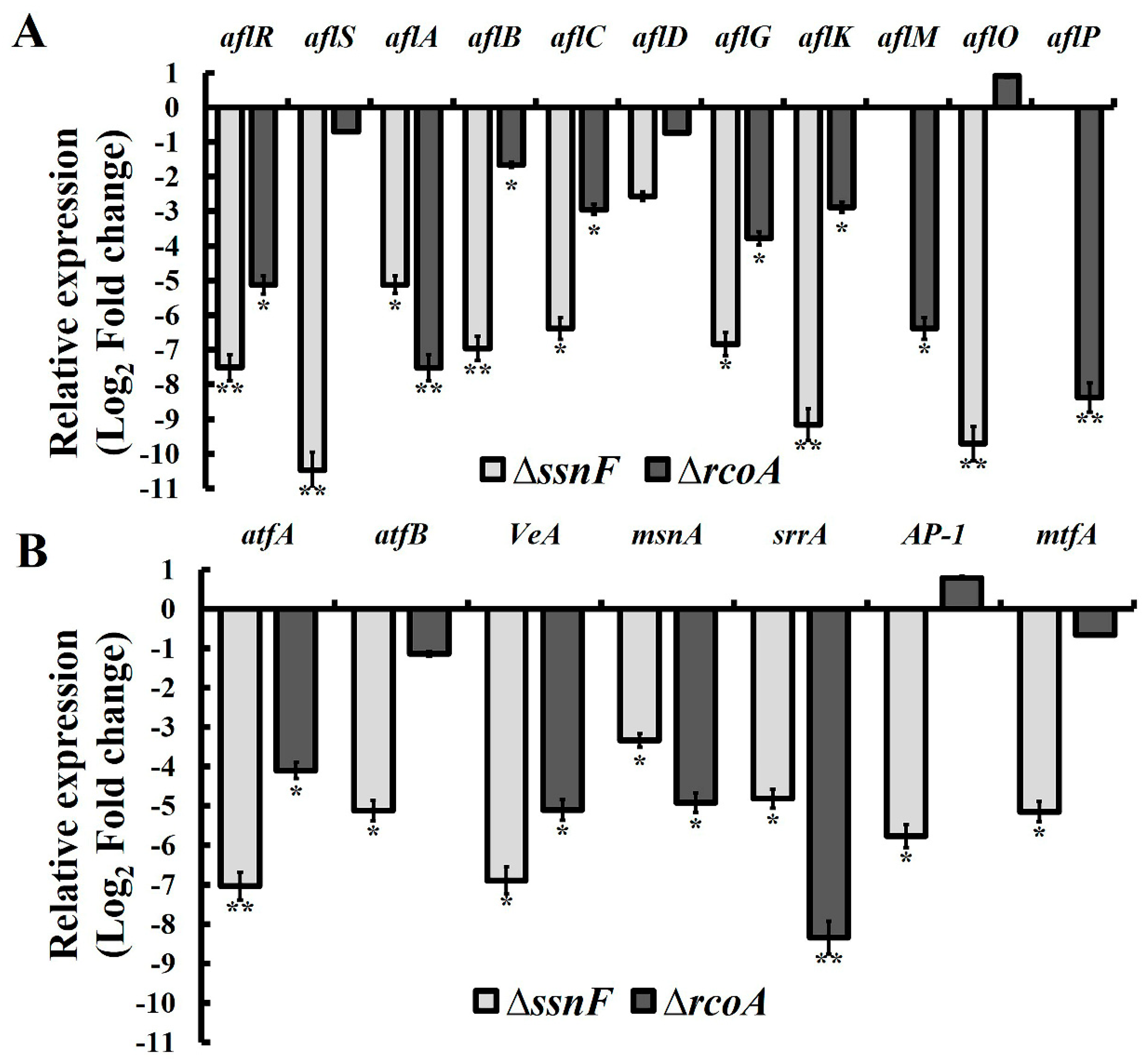
2.3. SsnF and RcoA Regulate AFB1 Biosynthesis by Modulating the Transcriptional Expressions of AF Cluster Genes and AF Related Regulators
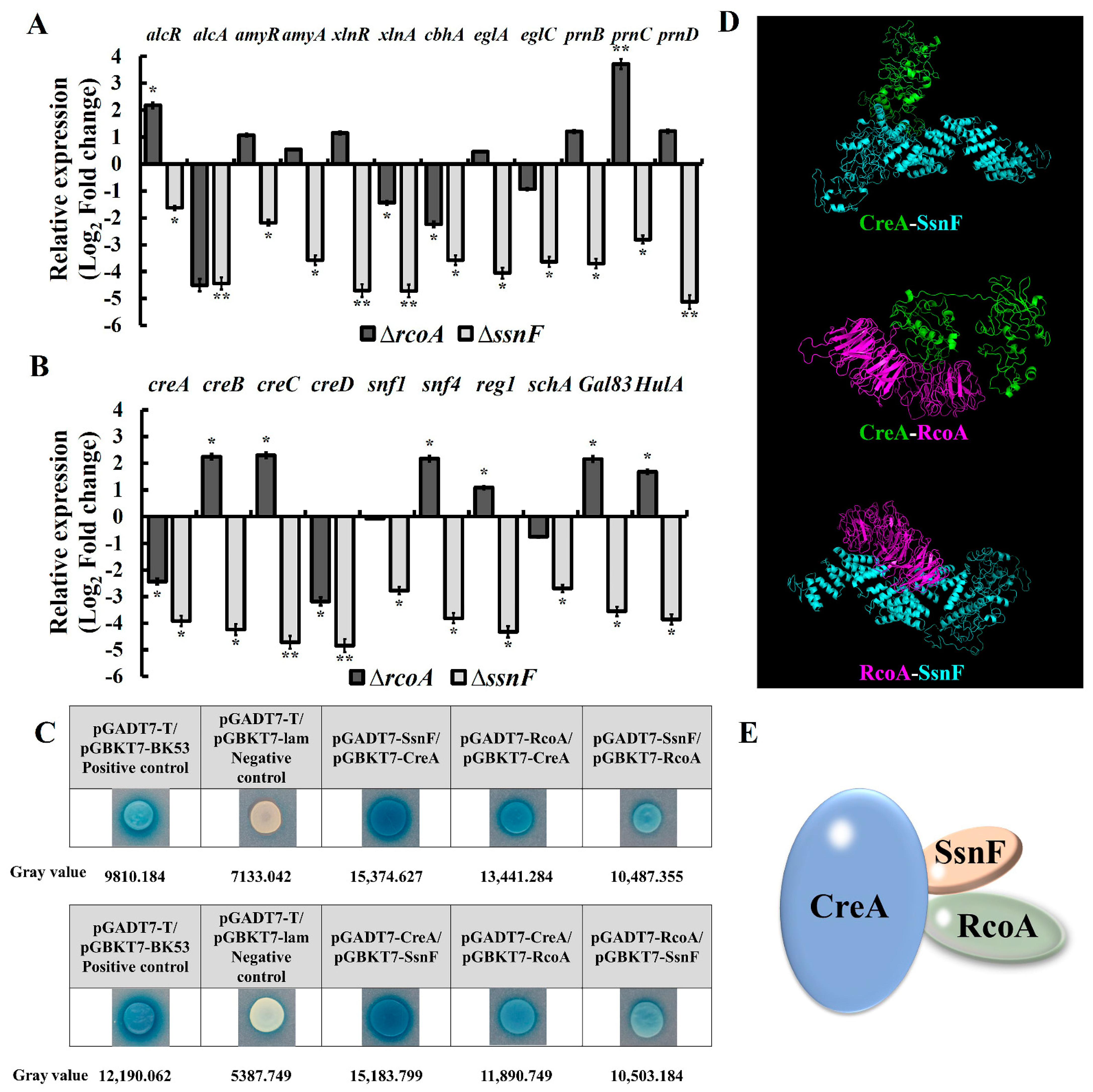
2.4. SsnF and RcoA Affect the Expressions of Hydrolase Genes by Regulating CCR Pathway Factors
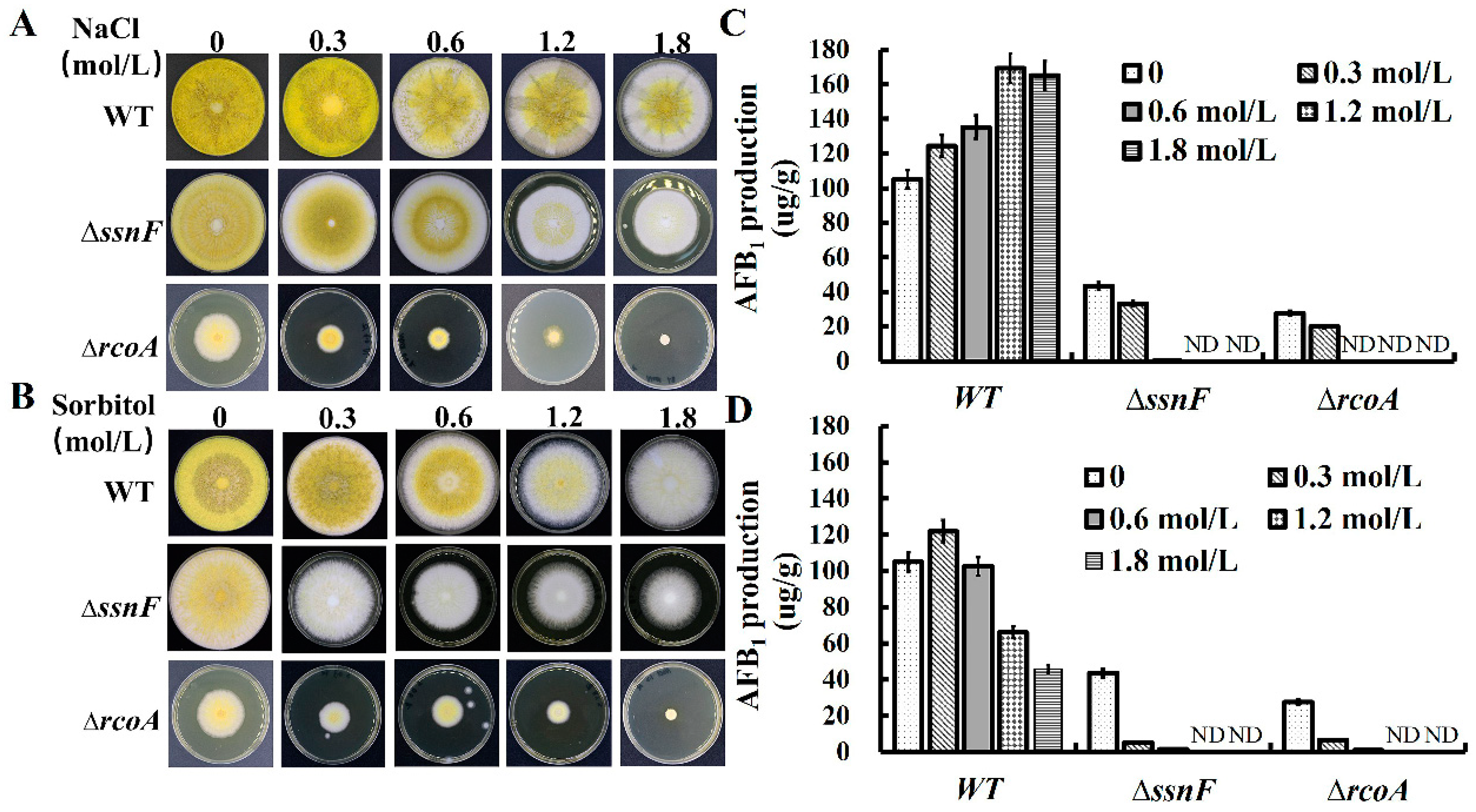
2.5. SsnF and RcoA Are Associated with the Response to Osmotic Stress in A. flavus
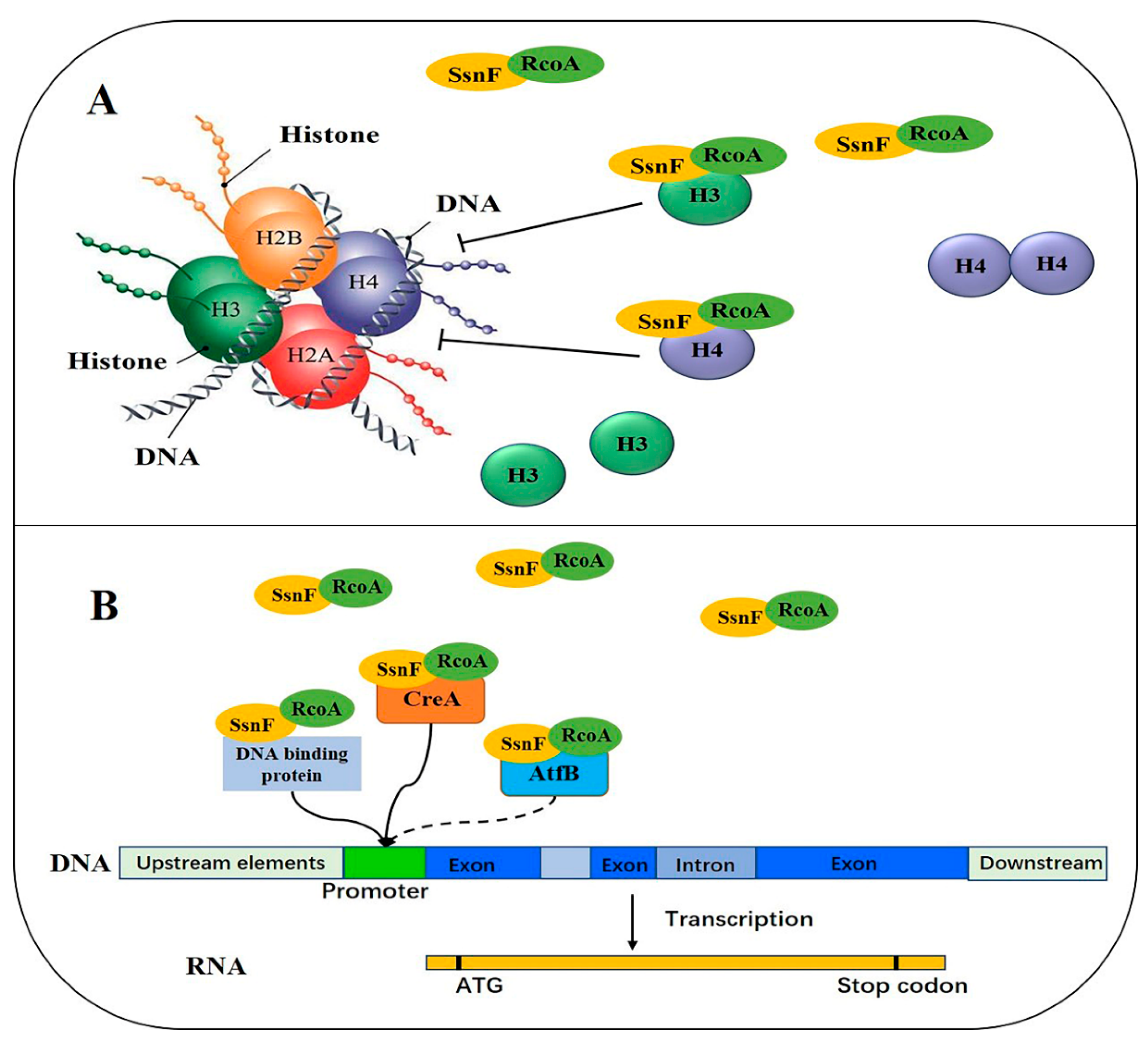
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Strains and Culture Conditions
5.2. Sequence Analyses and Phylogenetic Tree Construction
5.3. Construction of the Deletion Cassette and Complementary Strains
5.4. The Extraction and Detection of AFB1 Production
5.5. RNA Extraction and RT-qPCR Analysis
5.6. Yeast Two-Hybrid Assay
5.7. Analysis of Osmotic Stress on Different Mutants
5.8. Protein Docking Analysis
5.9. Statistical Analysis
5.10. Experimental Design
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Klich, M.A. Biogeography of Aspergillus species in soil and litter. Mycologia 2002, 94, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Benkerroum, N. Aflatoxins: Producing-molds, structure, health issues and incidence in Southeast Asian and Sub-Saharan African countries. Int. J. Environ. Res. Public Health 2020, 17, 1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amaike, S.; Keller, N.P. Aspergillus flavus. Annu. Rev. Phytopathol. 2011, 49, 107–133. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Felicia, W. Global burden of aflatoxin-induced hepatocellular carcinoma: A risk assessment. Environ. Health Perspect. 2010, 118, 818–824. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Qiu, Y.; Wang, X.; Gu, Y.; Zhao, Y.; Wang, Y.; Yue, T.; Yuan, Y. Inhibitory effects of eurotium cristatum on growth and aflatoxin B1 biosynthesis in Aspergillus flavus. Front. Microbiol. 2020, 11, 921. [Google Scholar] [CrossRef]
- Caceres, I.; Khoury, A.A.; Khoury, R.E.; Lorber, S.; Oswald, I.P.; Khoury, A.E.; Atoui, A.; Puel, O.; Bailly, J.D. Aflatoxin biosynthesis and genetic regulation: A review. Toxins 2020, 12, 150. [Google Scholar] [CrossRef] [Green Version]
- Amare, M.G.; Keller, N.P. Molecular mechanisms of Aspergillus flavus secondary metabolism and development. Fungal Genet. Biol. 2014, 66, 11–18. [Google Scholar] [CrossRef]
- Gallo, A.; Solfrizzo, M.; Epifani, F.; Panzarini, G.; Perrone, G. Effect of temperature and water activity on gene expression and aflatoxin biosynthesis in Aspergillus flavus on almond medium. Int. J. Food Microbiol. 2016, 217, 162–169. [Google Scholar] [CrossRef]
- Szilágyi, M.; Miskei, M.; Karányi, Z.; Lenkey, B.; Pócsi, I.; Emri, T. Transcriptome changes initiated by carbon starvation in Aspergillus nidulans. Microbiology 2013, 159, 176–190. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.P.; Carlson, M. Regulation of Snf1 protein kinase in response to environmental stress. J. Biol. Chem. 2007, 282, 16838–16845. [Google Scholar] [CrossRef] [Green Version]
- Ries, L.N.A.; Beattie, S.R.; Espeso, E.A.; Cramer, R.A.; Goldman, G.H. Diverse Regulation of the CreA carbon catabolite repressor in Aspergillus nidulans. Genetics 2016, 203, 335–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adnan, M.; Zheng, W.; Islam, W.; Arif, M.; Abubakar, Y.S.; Wang, Z.; Lu, G. Carbon catabolite repression in Filamentous Fungi. Int. J. Mol. Sci. 2017, 19, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubicek, C.P.; Mikus, M.; Schuster, A.; Schmoll, M.; Seiboth, B. Metabolic engineering strategies for the improvement of cellulase production by Hypocrea jecorina. Biotechnol. Biofuels 2009, 2, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, X.; Yu, A.; Zhang, C.; Pi, L.; Bai, X.; Xiao, D. Functional analysis of the global repressor Tup1 for maltose metabolism in Saccharomyces cerevisiae: Different roles of the functional domains. Microb. Cell. Factories 2017, 16, 194. [Google Scholar] [CrossRef] [Green Version]
- Papamichos-Chronakis, M.; Gligoris, T.; Tzamarias, D. The Snf1 kinase controls glucose repression in yeast by modulating interactions between the Mig1 repressor and the Cyc8-Tup1 co-repressor. EMBO Rep. 2004, 5, 368–372. [Google Scholar] [CrossRef] [Green Version]
- García, I.; Mathieu, M.; Nikolaev, I.; Felenbok, B.; Scazzocchio, C. Roles of the Aspergillus nidulans homologues of Tup1 and Ssn6 in chromatin structure and cell viability. FEMS Microbiol. Lett. 2008, 289, 146–154. [Google Scholar] [CrossRef] [Green Version]
- Fasoyin, O.E.; Wang, B.; Qiu, M.; Han, X.; Chung, K.R.; Wang, S. Carbon catabolite repression gene creA regulates morphology, aflatoxin biosynthesis and virulence in Aspergillus flavus. Fungal Genet. Biol. 2018, 115, 41–51. [Google Scholar] [CrossRef]
- Hwang, C.S.; Oh, J.H.; Huh, W.K.; Yim, H.S.; Kang, S.O. Ssn6, an important factor of morphological conversion and virulence in Candida albicans. Mol. Microbiol. 2003, 47, 1029–1043. [Google Scholar] [CrossRef]
- Tanaka, M.; Gomi, K. Induction and repression of hydrolase genes in Aspergillus oryzae. Front. Microbiol. 2021, 12, 677603. [Google Scholar] [CrossRef]
- Márquez, J.A.; Pascual-Ahuir, A.; Proft, M.; Serrano, R. The Ssn6-Tup1 repressor complex of Saccharomyces cerevisiae is involved in the osmotic induction of HOG-dependent and -independent genes. EMBO J. 1998, 17, 2543–2553. [Google Scholar] [CrossRef] [Green Version]
- Treitel, M.A.; Carlson, M. Repression by SSN6-TUP1 is directed by MIG1, a repressor/activator protein. Proc. Natl. Acad. Sci. USA 1995, 92, 3132–3136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Zhang, W.; Cao, Y.; Zheng, F.; Zhao, G.; Lv, X.; Meng, X.; Liu, W. Interdependent recruitment of CYC8/TUP1 and the transcriptional activator XYR1 at target promoters is required for induced cellulase gene expression in Trichoderma reesei. PLoS Genet. 2021, 17, e1009351. [Google Scholar] [CrossRef] [PubMed]
- Lettow, J.; Aref, R.; Schüller, H.J. Transcriptional repressor Gal80 recruits corepressor complex Cyc8-Tup1 to structural genes of the Saccharomyces cerevisiae GAL regulon. Curr. Genet. 2021, 68, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Watson, A.D.; Fau, E.D.; Bone, J.R.; Mukai, Y.; Yu, Y.; Du, W.; Stillman, D.J.; Roth, S.Y. Ssn6-Tup1 interacts with class I histone deacetylases required for repression. Genes Dev. 2000, 14, 2737–2744. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.L.; Johnson, A.D. Turning genes off by Ssn6-Tup1: A conserved system of transcriptional repression in eukaryotes. Trends Biochem. Sci. 2000, 25, 325–330. [Google Scholar] [CrossRef]
- Gong, J.; Huang, Q.; Liang, W.; Wei, Y.; Huang, G. The general transcriptional repressor Tup1 governs filamentous development in Candida tropicalis. Acta Biochim. Biophys. Sin. 2019, 51, 463–470. [Google Scholar] [CrossRef]
- Hicks, J.; Lockington, R.A.; Strauss, J.; Dieringer, D.; Kubicek, C.P.; Kelly, J.; Keller, N. RcoA has pleiotropic effects on Aspergillus nidulans cellular development. Mol. Microbiol. 2001, 39, 1482–1493. [Google Scholar] [CrossRef]
- Hong, S.Y.; Roze, L.V.; Linz, J.E. Oxidative stress-related transcription factors in the regulation of secondary metabolism. Toxins 2013, 5, 683–702. [Google Scholar] [CrossRef] [Green Version]
- Roze, L.V.; Chanda, A.; Wee, J.; Awad, D.; Linz, J.E. Stress-related transcription factor AtfB integrates secondary metabolism with oxidative stress response in Aspergilli. J. Biol. Chem. 2011, 286, 35137–35148. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Zhi, Q.Q.; Li, J.Y.; Keller, N.P.; He, Z.M. The antioxidant gallic acid inhibits aflatoxin formation in Aspergillus flavus by modulating transcription factors FarB and CreA. Toxins 2018, 10, 270. [Google Scholar] [CrossRef] [Green Version]
- Varanasi, U.S.; Klis, M.; Mikesell, P.B.; Trumbly, R.J. The Cyc8 (Ssn6)-Tup1 corepressor complex is composed of one Cyc8 and four Tup1 subunits. Mol. Cell. Biol. 1996, 16, 6707–6714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tartas, A.; Zarkadas, C.; Palaiomylitou, M.; Gounalaki, N.; Tzamarias, D.; Vlassi, M.A.O. Ssn6-Tup1 global transcriptional co-repressor: Role of the N-terminal glutamine-rich region of Ssn6. PLoS ONE 2017, 12, e0186363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, P.; Scharfenstein, L.L.; Li, R.W.; Arroyo-Manzanares, N.; De Saeger, S.; Diana Di Mavungu, J. Aspergillus flavus aswA, a gene homolog of Aspergillus nidulans oefC, regulates sclerotial development and biosynthesis of sclerotium-associated secondary metabolites. Fungal Genet. Biol. 2017, 104, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA 5: Molecular evolutionary genetics analysis using maximumlikelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, P.; Scharfenstein, L.L.; Wei, Q.; Bhatnagar, D. Development and refinement of a high-efficiency gene-targeting system for Aspergillus flavus. J. Microbiol. Methods 2010, 81, 240–246. [Google Scholar] [CrossRef]
- Liang, D.; Xing, F.; Selvaraj, J.N.L.; Liu, X.; Wang, L.; Hua, H.; Zhou, L.; Zhao, Y.; Wang, Y.; Liu, Y. Inhibitory effect of cinnamaldehyde, citral, and eugenol on aflatoxin biosynthetic gene expression and aflatoxin B1 biosynthesis in Aspergillus flavus. J. Food. Sci. 2015, 80, M2917–M2924. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Olson, A.J. Using AutoDock for ligand-receptor docking. Curr. Protoc. Bioinform. 2008, 24, 8–14. [Google Scholar] [CrossRef]
- El-Hachem, N.; Haibe-Kains, B.; Khalil, A.; Kobeissy, F.H.; Nemer, G. AutoDock and AutoDockTools for protein-ligand docking: Beta-site amyloid precursor protein cleaving enzyme 1(BACE1) as a case study. Methods Mol. Biol. 2017, 1598, 391–403. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, X.; Jiang, Y.; Ma, L.; Luo, S.; Du, H.; Li, X.; Xing, F. Corepressors SsnF and RcoA Regulate Development and Aflatoxin B1 Biosynthesis in Aspergillus flavus NRRL 3357. Toxins 2022, 14, 174. https://doi.org/10.3390/toxins14030174
Ma X, Jiang Y, Ma L, Luo S, Du H, Li X, Xing F. Corepressors SsnF and RcoA Regulate Development and Aflatoxin B1 Biosynthesis in Aspergillus flavus NRRL 3357. Toxins. 2022; 14(3):174. https://doi.org/10.3390/toxins14030174
Chicago/Turabian StyleMa, Xiaoyun, Yiran Jiang, Longxue Ma, Shujuan Luo, Haolan Du, Xu Li, and Fuguo Xing. 2022. "Corepressors SsnF and RcoA Regulate Development and Aflatoxin B1 Biosynthesis in Aspergillus flavus NRRL 3357" Toxins 14, no. 3: 174. https://doi.org/10.3390/toxins14030174
APA StyleMa, X., Jiang, Y., Ma, L., Luo, S., Du, H., Li, X., & Xing, F. (2022). Corepressors SsnF and RcoA Regulate Development and Aflatoxin B1 Biosynthesis in Aspergillus flavus NRRL 3357. Toxins, 14(3), 174. https://doi.org/10.3390/toxins14030174