The Effect of Australian and Asian Commercial Antivenoms in Reversing the Post-Synaptic Neurotoxicity of O. hannah, N. naja and N. kaouthia Venoms In Vitro



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
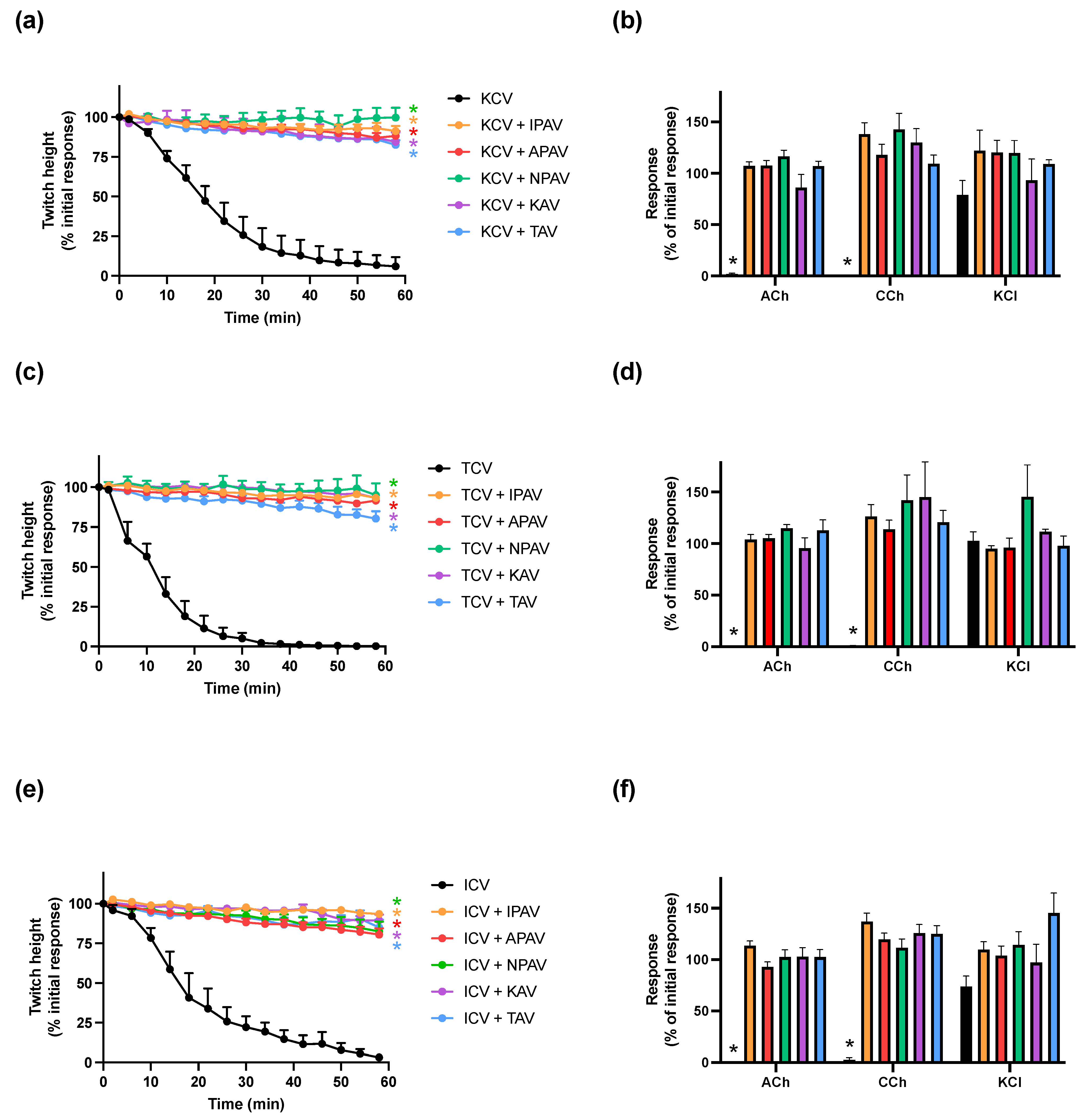
2.1. Prevention of In Vitro Neurotoxicity due to O. hannah, N. kaouthia and N. naja Venoms with Antivenom
2.2. Reversal of In Vitro Neurotoxicity of Venoms by Different Antivenoms
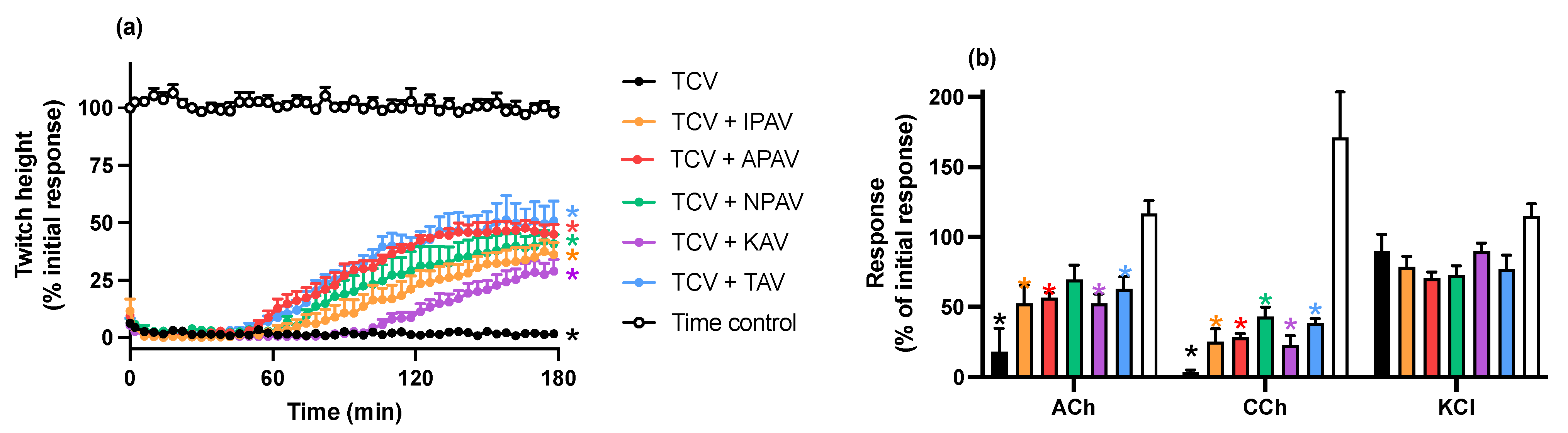
2.2.1. Reversal of In Vitro Neurotoxicity by O. hannah Venom by Antivenoms
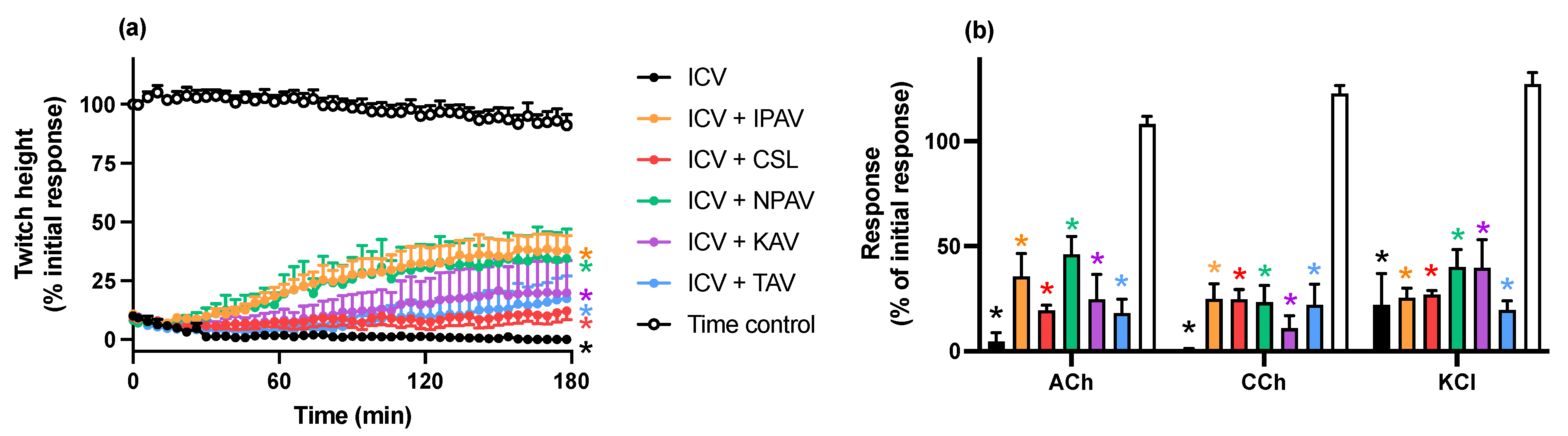
2.2.2. Reversal of In Vitro Neurotoxicity by N. kaouthia Venom by Antivenoms
2.2.3. Reversal of In Vitro Neurotoxicity by N. naja Venom by Antivenoms
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Venoms and Antivenoms
5.2. Chemical and Reagents
5.3. Chick Biventer Cervicis Nerve-Muscle Preparation
5.3.1. Antivenom Prevention Studies
5.3.2. Antivenom Reversal Studies
5.4. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Warrell, D.A. Snake bite. Lancet 2010, 375, 77–88. [Google Scholar] [CrossRef]
- Silva, A.; Hodgson, W.C.; Isbister, G.K. Antivenom for neuromuscular paralysis resulting from snake envenoming. Toxins 2017, 9, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barber, C.M.; Isbister, G.K.; Hodgson, W.C. Alpha neurotoxins. Toxicon 2013, 66, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.; Cristofori-Armstrong, B.; Rash, L.D.; Hodgson, W.C.; Isbister, G.K. Defining the role of post-synaptic α-neurotoxins in paralysis due to snake envenoming in humans. Cell. Mol. Life Sci. 2018, 75, 4465–4478. [Google Scholar] [CrossRef]
- Silva, A.; Isbister, G.K. Current research into snake antivenoms, their mechanisms of action and applications. Biochem. Soc. Trans. 2020, 48, 537–546. [Google Scholar] [CrossRef] [Green Version]
- Huynh, T.M.; Silva, A.; Isbister, G.K.; Hodgson, W.C. Isolation and pharmacological characterization of α-elapitoxin-Oh3a, a long-chain post-synaptic neurotoxin from King Cobra (Ophiophagus hannah) venom. Front. Pharmacol. 2022, 13, 815069. [Google Scholar] [CrossRef]
- Madhushani, U.; Isbister, G.K.; Tasoulis, T.; Hodgson, W.C.; Silva, A. In-vitro neutralization of the neurotoxicity of coastal taipan venom by Australian polyvalent antivenom: The window of opportunity. Toxins 2020, 12, 690. [Google Scholar] [CrossRef]
- Pochanugool, C.; Limthongkul, S.; Wilde, H. Management of thai cobra bites with a single bolus of antivenin. Wilderness Environ. Med. 1997, 8, 20–23. [Google Scholar] [CrossRef]
- Petras, D.; Heiss, P.; Sussmuth, R.D.; Calvete, J.J. Venom proteomics of Indonesian King Cobra, Ophiophagus hannah: Integrating top-Down and bottom-up approaches. J. Proteome Res. 2015, 14, 2539–2556. [Google Scholar] [CrossRef]
- Tan, C.H.; Tan, K.Y.; Fung, S.Y.; Tan, N.H. Venom-gland transcriptome and venom proteome of the Malaysian king cobra (Ophiophagus hannah). BMC Genom. 2015, 16, 687. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.Y.; Tan, C.H.; Fung, S.Y.; Tan, N.H. Venomics, lethality and neutralization of Naja kaouthia (monocled cobra) venoms from three different geographical regions of Southeast Asia. J. Proteom. 2015, 120, 105–125. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, M.; McCleary, R.J.R.; Kesherwani, M.; Kini, R.M.; Velmurugan, D. Comparison of proteomic profiles of the venoms of two of the ‘Big Four’ snakes of India, the Indian cobra (Naja naja) and the common krait (Bungarus caeruleus), and analyses of their toxins. Toxicon 2017, 135, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Chanda, A.; Kalita, B.; Islam, T.; Patra, A.; Mukherjee, A.K. Proteomic analysis to unravel the complex venom proteome of eastern India Naja naja: Correlation of venom composition with its biochemical and pharmacological properties. J. Proteom. 2017, 156, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.Y.; Tan, C.H.; Tan, K.Y.; Quraishi, N.H.; Tan, N.H. Elucidating the biogeographical variation of the venom of Naja naja (spectacled cobra) from Pakistan through a venom-decomplexing proteomic study. J. Proteom. 2018, 175, 156–173. [Google Scholar] [CrossRef]
- Liang, Q.; Huynh, T.M.; Konstantakopoulos, N.; Isbister, G.K.; Hodgson, W.C. An examination of the neutralization of in vitro toxicity of Chinese cobra (Naja atra) venom by different antivenoms. Biomedicines 2020, 8, 377. [Google Scholar] [CrossRef]
- Tin, M.; Mra, R.; Chit, M.; Pe, T.; Warrell, D.A. Bites by the king cobra (Ophiophagus hannah) in Myanmar: Successful treatment of severe neurotoxic envenoming. Q. J. Med. 1991, 80, 751–762. [Google Scholar] [CrossRef]
- Tun, P.; Myint, A.A.; Warrell, D.A.; Myint, T. King cobra (Ophiophagus hannah) bites in Myanmar: Venom antigen levels and development of venom antibodies. Toxicon 1995, 33, 379–382. [Google Scholar] [CrossRef]
- Kularatne, S.A.M.; Budagoda, B.D.S.S.; Gawarammana, I.B.; Kularatne, W.K.S. Epidemiology, clinical profile and management issues of cobra (Naja naja) bites in Sri Lanka: First authenticated case series. Trans. R. Soc. Trop. Med. Hyg. 2009, 103, 924–930. [Google Scholar] [CrossRef]
- Mohapatra, B.; Warrell, D.A.; Suraweera, W.; Bhatia, P.; Dhingra, N.; Jotkar, R.M.; Rodriguez, P.S.; Mishra, K.; Whitaker, R.; Jha, P. Snakebite mortality in India: A nationally representative mortality survey. PLoS Negl. Trop. Dis. 2011, 5, e1018. [Google Scholar] [CrossRef]
- Tasoulis, T.; Isbister, G.K. A review and database of snake venom proteomes. Toxins 2017, 9, 290. [Google Scholar] [CrossRef] [Green Version]
- Kornhauser, R.; Isbister, G.K.; O’Leary, M.A.; Mirtschin, P.; Dunstan, N.; Hodgson, W.C. Cross-neutralisation of the neurotoxic effects of Egyptian cobra venom with commercial tiger snake antivenom. Basic Clin. Pharmacol. Toxicol. 2012, 112, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Leong, P.K.; Fung, S.Y.; Tan, C.H.; Sim, S.M.; Tan, N.H. Immunological cross-reactivity and neutralization of the principal toxins of Naja sumatrana and related cobra venoms by a Thai polyvalent antivenom (Neuro Polyvalent Snake Antivenom). Acta Trop. 2015, 149, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.; Hodgson, W.C.; Isbister, G.K. Cross-neutralisation of in vitro neurotoxicity of Asian and Australian snake neurotoxins and venoms by different antivenoms. Toxins 2016, 8, 302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senji Laxme, R.R.; Attarde, S.; Khochare, S.; Suranse, V.; Martin, G.; Casewell, N.R.; Whitaker, R.; Sunagar, K. Biogeographical venom variation in the Indian spectacled cobra (Naja naja) underscores the pressing need for pan-India efficacious snakebite therapy. PLoS Negl. Trop. Dis. 2021, 15, e0009150. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.Y.; Tan, K.Y.; Tan, N.H.; Gnanathasan, C.A.; Tan, C.H. Elucidating the venom diversity in Sri Lankan Spectacled Cobra (Naja naja) through de novo venom gland transcriptomics, venom proteomics and toxicity neutralization. Toxins 2021, 13, 558. [Google Scholar] [CrossRef]
- Boulain, J.; Ménez, A. Neurotoxin-specific immunoglobulins accelerate dissociation of the neurotoxin-acetylcholine receptor complex. Science 1982, 217, 732–733. [Google Scholar] [CrossRef]
- Boulain, J.C.; Fromageot, P.; Menez, A. Further evidence showing that neurotoxin-acetylcholine receptor dissociation is accelerated by monoclonal neurotoxin-specific immunoglobulin. Mol. Immunol. 1985, 22, 553–556. [Google Scholar] [CrossRef]
- Gatineau, E.; Lee, C.Y.; Fromageot, P.; Menez, A. Reversal of snake neurotoxin binding to mammalian acetylcholine receptor by specific antiserum. Eur. J. Biochem. 1988, 171, 535–539. [Google Scholar] [CrossRef]
- Silva, A.; Hodgson, W.C.; Tasoulis, T.; Isbister, G.K. Rodent lethality models are problematic for evaluating antivenoms for human envenoming. Front. Pharmacol. 2022, 13, 830384. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huynh, T.M.; Hodgson, W.C.; Isbister, G.K.; Silva, A. The Effect of Australian and Asian Commercial Antivenoms in Reversing the Post-Synaptic Neurotoxicity of O. hannah, N. naja and N. kaouthia Venoms In Vitro. Toxins 2022, 14, 277. https://doi.org/10.3390/toxins14040277
Huynh TM, Hodgson WC, Isbister GK, Silva A. The Effect of Australian and Asian Commercial Antivenoms in Reversing the Post-Synaptic Neurotoxicity of O. hannah, N. naja and N. kaouthia Venoms In Vitro. Toxins. 2022; 14(4):277. https://doi.org/10.3390/toxins14040277
Chicago/Turabian StyleHuynh, Tam M., Wayne C. Hodgson, Geoffrey K. Isbister, and Anjana Silva. 2022. "The Effect of Australian and Asian Commercial Antivenoms in Reversing the Post-Synaptic Neurotoxicity of O. hannah, N. naja and N. kaouthia Venoms In Vitro" Toxins 14, no. 4: 277. https://doi.org/10.3390/toxins14040277
APA StyleHuynh, T. M., Hodgson, W. C., Isbister, G. K., & Silva, A. (2022). The Effect of Australian and Asian Commercial Antivenoms in Reversing the Post-Synaptic Neurotoxicity of O. hannah, N. naja and N. kaouthia Venoms In Vitro. Toxins, 14(4), 277. https://doi.org/10.3390/toxins14040277