Aflatoxin Contamination, Its Impact and Management Strategies: An Updated Review



Abstract
:1. Introduction
2. Aflatoxin Contamination
3. Impact on Human and Animal Health
3.1. Aspergillosis
3.2. Aflatoxicosis
3.3. Cancer
4. Detection of Aflatoxin-Producing Strains
4.1. Culture-Based Techniques
4.2. Molecular Based Techniques
5. Quantification/Detection of Aflatoxin
5.1. Immunochemical Methods for Detection of Aflatoxin
5.2. Biosensor-Based Techniques
5.3. Optical Immunosensor
5.4. Electrochemical Immunosensors
5.5. Sensor Array Method
5.6. Microfluidic Sensor Method
5.7. Chromatographic Methods
5.8. Spectroscopic Methods
6. Control of Aflatoxin Contamination
6.1. Biological Control
6.1.1. Atoxigenic Aspergillus Strains against Toxigenic Aspergillus Strains
6.1.2. Biological control at the experimental stages
Biological Control with Yeasts
Trichoderma spp.
Penicillium spp.
Biological Control with Bacteria
- (a)
- Bacillus spp.
- (b)
- Pseudomonas spp.
- (c)
- Lactobacillus spp.
- (d)
- Streptomyces spp.
- (e)
- Other Bacterial spp.
6.2. Chemical Control
6.3. Physical Control
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nazhand, A.; Durazzo, A.; Lucarini, M.; Souto, E.B.; Santini, A.J.F. Characteristics, occurrence, detection and detoxification of aflatoxins in foods and feeds. Foods 2020, 9, 644. [Google Scholar] [CrossRef]
- Eskola, M.; Kos, G.; Elliott, C.T.; Hajšlová, J.; Mayar, S.; Krska, R. Worldwide contamination of food-crops with mycotoxins: Validity of the widely cited ‘FAO estimate’ of 25%. Crit. Rev. Food. Sci. Nutr. 2020, 60, 2773–2789. [Google Scholar] [CrossRef] [PubMed]
- Campione, E.; Gaziano, R.; Doldo, E.; Marino, D.; Falconi, M.; Iacovelli, F.; Tagliaferri, D.; Pacello, L.; Bianchi, L.; Lanna, C.; et al. Antifungal effect of all-trans retinoic acid against Aspergillus fumigatus in vitro and in a pulmonary aspergillosis in vivo model. Antimicrob. Agents Chemother. 2020, 65, e01874-20. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.M.; Ahmad, M.; Ali, A.; Hamid, R.; Javed, S.; Abdin, M.Z. Detection of Aspergillus flavus and Aspergillus parasiticus from aflatoxin-contaminated peanuts and their differentiation using PCR-RFLP. Ann. Microbiol. 2014, 64, 1597–1605. [Google Scholar] [CrossRef]
- Luo, S.; Du, H.; Kebede, H.; Liu, Y.; Xing, F. Contamination status of major mycotoxins in agricultural product and food stuff in Europe. Food Control 2021, 127, 108120. [Google Scholar] [CrossRef]
- Kumar, P.; Mahato, D.K.; Kamle, M.; Mohanta, T.K.; Kang, S.G. Aflatoxins: A global concern for food safety, human health and their management. Front. Microbiol. 2017, 7, 2170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marr, K.A.; Platt, A.; Tornheim, J.A.; Zhang, S.X.; Datta, K.; Cardozo, C.; Garcia-Vidal, C. Aspergillosis complicating severe coronavirus disease. Emerg. Infect. Dis. 2021, 27, 18–25. [Google Scholar] [CrossRef]
- FDA (US. Food & Drug Adminitrtaion). CPG Sec. 683.100 Action Levels for Aflatoxins in Animal Feeds. 2019. Available online: https://www.fda.gov/regulatory-information/search-fda-guidance-documents/cpg-sec-683100-action-levels-aflatoxins-animal-feeds (accessed on 19 December 2021).
- Commission Directive 2003/100/EC of 31 October 2003 amending Annex I to Directive 2002/32/EC of the European Parliament and of the Council on undesirable substances in animal feed. OJEU 2003, 285, 33–37. Available online: https://eur-lex.europa.eu/legal content/EN/TXT/PDF/?uri = CELEX:32003L0100&from=EN (accessed on 19 December 2021).
- Kaale, L.; Kimanya, M.; Macha, I.; Mlalila, N. Aflatoxin contamination and recommendations to improve its control: A review. World Mycotoxin J. 2021, 14, 27–40. [Google Scholar] [CrossRef]
- Benkerroum, N. Aflatoxins: Producing-Molds, Structure, Health Issues and Incidence in Southeast Asian and Sub-Saharan African Countries. Int. J. Environ. Res. Public Health 2020, 17, 1215. [Google Scholar] [CrossRef] [Green Version]
- Abdin, M.; Ahmad, M.M.; Javed, S. Advances in molecular detection of Aspergillus: An update. Arch. Microbiol. 2010, 192, 409–425. [Google Scholar] [CrossRef]
- Santini, A.; Ritieni, A. Aflatoxins: Risk, exposure and remediation. In Aflatoxins: Recent Advances and Future Prospects; IntechOpen: London, UK, 2013; pp. 343–376. [Google Scholar]
- Coppock, R.W.; Christian, R.G.; Jacobsen, B.J. Aflatoxins. In Veterinary Toxicology, 3rd ed.; Gupta, R.C., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 983–994. [Google Scholar] [CrossRef]
- Benkerroum, N. Chronic and Acute Toxicities of Aflatoxins: Mechanisms of Action. Int. J. Environ. Res. Public Health 2020, 17, 423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lalah, J.O.; Omwoma, S.; Orony, D.A.O. Aflatoxin B1: Chemistry, environmental and diet sources and potential exposure in human in Kenya. In Aflatoxin B1 Occurrence, Detection and Toxicological Effects; BoD–Books on Demand: Norderstedt, Germany, 2020. [Google Scholar] [CrossRef]
- Marchese, S.; Polo, A.; Ariano, A.; Velotto, S.; Costantini, S.; Severino, L. Aflatoxin B1 and M1: Biological Properties and Their Involvement in Cancer Development. Toxins 2018, 10, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostry, V.; Malir, F.; Toman, J.; Grosse, Y. Mycotoxins as human carcinogens—the IARC Monographs classification. Mycotoxin Res. 2017, 33, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Amaike, S.; Keller, N.P. Aspergillus flavus. Annu. Rev. Phytopathol. 2011, 49, 107–133. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.W.; Afzaal, S. Aflatoxins in Pakistani Foods: A serious threat to food safety. J. Hyg. Eng. Des. 2014, 9, 20–25. Available online: https://keypublishing.org/jhed/wp-content/uploads/2020/07/02.-Full-Paper-Shinawar-Waseem-Ali.pdf (accessed on 19 December 2021).
- Callicott, K.A.; Kachapalula, P.; Edmunds, D.; Singh, P.; Jaime, R.; Islam, M.S.; Shenge, K.; Adhikari, B.N.; Arone-Maxwell, L.; Ching’anda, C.; et al. Brief Protocols for Research on Management of Aflatoxin-Producing Fungi, 2nd ed.; ARS, USDA: Phoenix, AZ, USA, 2018. [Google Scholar]
- Liu, X.; Guan, X.; Xing, F.; Lv, C.; Dai, X.; Liu, Y. Effect of water activity and temperature on the growth of Aspergillus flavus, the expression of aflatoxin biosynthetic genes and aflatoxin production in shelled peanuts. Food Control 2017, 82, 325–332. [Google Scholar] [CrossRef]
- Abdel-Hadi, A.; Schmidt-Heydt, M.; Parra, R.; Geisen, R.; Magan, N. A systems approach to model the relationship between aflatoxin gene cluster expression, environmental factors, growth and toxin production by Aspergillus flavus. J. R. Soc. Interface 2012, 9, 757–767. [Google Scholar] [CrossRef] [Green Version]
- Bernáldez, V.; Cordoba, J.J.; Magan, N.; Peromingo, B.; Rodríguez, A. The influence of ecophysiological factors on growth, aflR gene expression and aflatoxin B1 production by a type strain of Aspergillus flavus. LWT Food Sci. Technol. 2017, 83, 283–291. [Google Scholar] [CrossRef] [Green Version]
- Tejero, P.; Martín, A.; Rodríguez, A.; Galván, A.I.; Ruiz-Moyano, S.; Hernández, A. In Vitro Biological Control of Aspergillus flavus by Hanseniaspora opuntiae L479 and Hanseniaspora uvarum L793, Producers of Antifungal Volatile Organic Compounds. Toxins 2021, 13, 663. [Google Scholar] [CrossRef]
- Bhatnagar, D.; Cary, J.W.; Ehrlich, K.; Yu, J.; Cleveland, T.E. Understanding the genetics of regulation of aflatoxin production and Aspergillus flavus development. Mycopathologia 2006, 162, 155. [Google Scholar] [CrossRef]
- Rapid Alert System for Food and Feed (RASFF). Available online: https://webgate.ec.europa.eu/rasff-window/screen/search (accessed on 9 February 2020).
- Pickova, D.; Ostry, V.; Toman, J.; Malir, F. Aflatoxins: History, Significant Milestones, Recent Data on Their Toxicity and Ways to Mitigation. Toxins 2021, 13, 399. [Google Scholar] [CrossRef] [PubMed]
- Jallow, A.; Xie, H.; Tang, X.; Qi, Z.; Li, P. Worldwide aflatoxin contamination of agricultural products and foods: From occurrence to control. Compr. Rev. Food Sci. Food Saf. 2021, 20, 2332–2381. [Google Scholar] [CrossRef] [PubMed]
- Denning, D.W.; Pleuvry, A.; Cole, D.C. Global burden of chronic pulmonary aspergillosis as a sequel to pulmonary tuberculosis. Bull World Health Organ 2011, 89, 864–872. [Google Scholar] [CrossRef]
- Denning, D.W.; Pleuvry, A.; Cole, D.C. Global burden of allergic bronchopulmonary aspergillosis with asthma and its complication chronic pulmonary aspergillosis in adults. Med. Mycol. J. 2013, 51, 361–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, D.; Moss, R.; Kurup, V.; Knutsen, A.; Greenberger, P.; Judson, M.; Denning, D.; Crameri, R.; Brody, A.; Light, M.; et al. Allergic bronchopulmonary aspergillosis in cystic fibrosis-state of the art: Cystic Fibrosis Foundation consensus conference. Clin. Infect. Dis. 2003, 37, S225–S264. [Google Scholar] [CrossRef] [PubMed]
- Winters, B.; Custer, J.; Galvagno, S.M.; Colantuoni, E.; Kapoor, S.G.; Lee, H.; Goode, V.; Robinson, K.; Nakhasi, A.; Pronovost, P.; et al. Diagnostic errors in the intensive care unit: A systematic review of autopsy studies. BMJ Qual. Saf. 2012, 21, 894–902. [Google Scholar] [CrossRef]
- Dhanasekaran, D.; Shanmugapriya, S.; Thajuddin, N.; Panneerselvam, A. Aflatoxins and aflatoxicosis in human and animals. In Aflatoxins: Biochemistry and Molecular Biology; Guevara-González, R.G., Ed.; BoD–Books on Demand: London, UK, 2011; Volume 10, pp. 221–254. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Aflatoxins; WHO: Geneva, Switzerland, 2018; Available online: https://www.who.int/foodsafety/FSDigest_Aflatoxins_EN.pdf (accessed on 19 December 2021).
- Grace, D.; Lindahl, J.F.; Atherstone, C.; Kang’ethe, E.; Nelson, F.; Wesonga, T.; Manyong, V. Aflatoxin Standards for Feed. Building an Aflatoxin Safe East African Community Technical Policy Paper 7; International Institute of Tropical Agriculture: Ibadan, Nigeria, 2015; Available online: https://cgspace.cgiar.org/bitstream/handle/10568/75535/Aflatoxin%20standards%20for%20feed.pdf?sequence=1 (accessed on 19 December 2021).
- Chandra, P. Aflatoxins: Food Safety, Human Health Hazards and Their Prevention. In Aflatoxins—Occurrence, Detoxification, Determination and Health Risks; IntechOpen: London, UK, 2021. [Google Scholar]
- Kimanya, M.E.; Routledge, M.N.; Mpolya, E.; Ezekiel, C.N.; Shirima, C.P.; Gong, Y.Y. Estimating the risk of aflatoxin-induced liver cancer in Tanzania based on biomarker data. PLoS ONE 2021, 16, e0247281. [Google Scholar] [CrossRef]
- Alkhersan, R.N.; Khudor, M.H.; Abbas, B.A. Rapid detection of aflatoxigenic producing atrains of Aspergillus flavus from poultry fees by uv light and ammonia. Bas. J. Vet. Res. 2016, 14, 169–178. [Google Scholar]
- Iram, W.; Anjum, T.; Jabeen, R.; Abbas, M. Isolation of stored maize mycoflora, identification of aflatoxigenic fungi and its inhibition using medicinal plant extracts. Int. J. Agric. Biol. 2018, 20, 2149–2160. [Google Scholar] [CrossRef]
- Usman, M.; Javed, M.R.; Mehmood, M.A.; Huma, T.; Ijaz, A. Isolation of aflatoxigenic Aspergillus flavus from animal-feed and exploration of the genetic basis of aflatoxin biosynthesis. Pak. Vet. J. 2019, 39, 541–547. [Google Scholar] [CrossRef]
- Yazdani, D.; Zainal, A.M.; Tan, Y.; Kamaruzaman, S. Evaluation of the detection techniques of toxigenic Aspergillus isolates. Afr. J. Biotechnol. 2010, 9, 7654–7659. [Google Scholar] [CrossRef]
- Abbas, H.K.; Shier, W.; Horn, B.; Weaver, M.A. Cultural methods for aflatoxin detection. J. Toxicol. Toxin Rev. 2004, 23, 295–315. [Google Scholar] [CrossRef]
- Levin, R.E. PCR detection of aflatoxin producing fungi and its limitations. Int. J. Food Microbiol. 2012, 156, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Bintvihok, A.; Treebonmuang, S.; Srisakwattana, K.; Nuanchun, W.; Patthanachai, K.; Usawang, S. A rapid and sensitive detection of aflatoxin-producing fungus using an optimized polymerase chain reaction (PCR). Toxicol. Res. 2016, 32, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.-Y.; Shim, W.-B.; Kim, J.-H.; Park, S.-J.; Kang, S.-J.; Nam, B.-S.; Chung, D.H. Detection of aflatoxin-producing molds in Korean fermented foods and grains by multiplex PCR. J. Food Prot. 2004, 67, 2622–2626. [Google Scholar] [CrossRef] [PubMed]
- Segura, G.G.; Arevabini, C.A.M.; Heinzen de Abreu, M.; Bitencourt, T.A.; Nishimura, F.G.; Beleboni, R.; Marins, M.; Fachin, A.L. The molecular and analytical methods reveal aflatoxin.B1-producing Aspergillus flavus isolated from ready-to-eat peanut samples are resistant to the antifungal agent methyl-thiophanate. J. Food Sci. Technol. 2020, 5, 1–7. [Google Scholar] [CrossRef]
- Azeez, A.Z.; Hindi, M.T.; Khudiar, M.M.; Saadi, A.S.A.; Khadim, N.I. Determination of Aflatoxins in Feed by HPLC and PCR. J. Food Pharm. Sci. 2020, 8, 315–323. [Google Scholar] [CrossRef]
- Yahyaraeyat, R.; Khosravi, A.; Shahbazzadeh, D.; Khalaj, V. The potential effects of Zataria multiflora Boiss essential oil on growth, aflatoxin production and transcription of aflatoxin biosynthesis pathway genes of toxigenic Aspergillus parasiticus. Braz. J. Microbiol. 2013, 44, 649–655. [Google Scholar] [CrossRef] [Green Version]
- Mayer, Z.; Färber, P.; Geisen, R. Monitoring the production of aflatoxin B1 in wheat by measuring the concentration of nor-1 mRNA. Appl. Environ. Microbiol. 2003, 69, 1154–1158. [Google Scholar] [CrossRef] [Green Version]
- Degola, F.; Berni, E.; Dall’Asta, C.; Spotti, E.; Marchelli, R.; Ferrero, I.; Restivo, F.M. A multiplex RT-PCR approach to detect aflatoxigenic strains of Aspergillus flavus. J. Appl. Microbiol. 2007, 103, 409–417. [Google Scholar] [CrossRef]
- Rodrigues, P.; Soares, C.; Kozakiewicz, Z.; Paterson, R.; Lima, N.; Venâncio, A. Identification and Characterization of Aspergillus flavus and Aflatoxins. In Communicating Current Research and Educational Topics and Trends in Applied Microbiology; Formatex: Badajoz, Spain, 2007; pp. 527–534. Available online: http://hdl.handle.net/10198/933 (accessed on 19 December 2021).
- Jonathan, G.; Abdul-Lateef, N.; Hull, A.; Chikwem, J.; Asemoloye, M.; Odeseye, A. Characterization and detection of aflatoxigenic genes in fungi associated with kulikuli (groundnut cake) from Nigeria. J. Sci. 2019, 8. Available online: https://www.researchgate.net/profile/John-Chikwem/publication/341495114_Characterization_and_detection_of_aflatoxigenic_genes_in_fungi_associated_with_kulikuli_groundnut_cake_from_Nigeria/links/5ec435ae92851c11a87775f1/Characterization-and-detection-of-aflatoxigenic-genes-in-fungi-associated-with-kulikuli-groundnut-cake-from-Nigeria.pdf (accessed on 19 December 2021).
- Al-Hmoud, N.; Ibrahim, M.A.; Al-Rousan, H.; Alseyah, A. The prevalence of aflatoxinogenic Aspergillus parasiticus in Jordan. Int. J. Microbiol. 2012, 2012, 675361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abd El-Aziz, A.R.; Shehata, S.M.; Hisham, S.M.; Alobathani, A.A. Molecular profile of aflatoxigenic and non-aflatoxigenic isolates of Aspergillus flavus isolated from stored maize. Saudi. J. Biol. Sci. 2021, 28, 1383–1391. [Google Scholar] [CrossRef] [PubMed]
- Cruz, P.; Buttner, M.P. Development and evaluation of a real-time quantitative PCR assay for Aspergillus flavus. Mycologia 2008, 100, 683–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sardiñas, N.; Vazquez, C.; Gil-Serna, J.; González-Jaén, M.T.; Patiño, B. Specific detection of Aspergillus parasiticus in wheat flour using a highly sensitive PCR assay. Food Addit. Contam. 2010, 27, 853–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, P.; Orbach, M.J.; Cotty, P.J. Aspergillus texensis: A Novel Aflatoxin Producer with S Morphology from the United States. Toxins 2018, 10, 513. [Google Scholar] [CrossRef] [Green Version]
- Ehrlich, K.C.; Chang, P.-K.; Yu, J.; Cotty, P.J. Aflatoxin biosynthesis cluster gene cypA is required for G aflatoxin formation. Appl. Environ. Microbiol. 2004, 70, 6518–6524. [Google Scholar] [CrossRef] [Green Version]
- Probst, C.; Callicott, K.; Cotty, P.J. Deadly strains of Kenyan Aspergillus are distinct from other aflatoxin producers. Eur. J. Plant Pathol. 2012, 132, 419–429. [Google Scholar] [CrossRef]
- Fakhoury, A.; Woloshuk, C.P. Amy1, the α-amylase gene of Aspergillus flavus: Involvement in aflatoxin biosynthesis in maize kernels. APS 1999, 89, 908–914. [Google Scholar] [CrossRef] [Green Version]
- Beyene, A.M.; Du, X.; Schrunk, D.E.; Ensley, S.; Rumbeiha, W.K. High-performance liquid chromatography and Enzyme-Linked Immunosorbent Assay techniques for detection and quantification of aflatoxin B 1 in feed samples: A comparative study. BMC Res. Notes 2019, 12, 492. [Google Scholar] [CrossRef]
- Ostadrahimi, A.; Ashrafnejad, F.; Kazemi, A.; Sargheini, N.; Mahdavi, R.; Farshchian, M.; Mahluji, S. Aflatoxin in raw and salt-roasted nuts (pistachios, peanuts and walnuts) sold in markets of Tabriz, Iran. Jundishapur J. Microbiol. 2014, 7, e8674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huybrechts, B. Evaluation of Immunoassay Kits for Aflatoxin Determination in Corn & Rice; Agrochemical Research Centre, NRL: Brussels, Belgium, 2011; Available online: https://www.favv-afsca.be/laboratoires/laboratoiresagrees/_documents/2012-05-04_Evaluation_immunoassay_kits_aflatoxin.pdf (accessed on 16 March 2020).
- Roman, B.E.; Driksna, D.; Abouzied, M.M.; Klein, F.; Rice, J. Validation of MAX Aqueous Extraction on Veratox® for Total Aflatoxin ELISA Test Kit. J. AOAC Int. 2017, 100, 1131–1133. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Pillai, M.; Gautam, S.; Hajare, S. Mycotoxins Immunological techniques for detection and analysis. In Encyclopedia of Food Microbiology; Elsevier: Amsterdam, The Netherlands, 2014; pp. 869–879. [Google Scholar] [CrossRef]
- Anfossi, L.; Baggiani, C.; Giovannoli, C.; Giraudi, G. Lateral flow immunoassays for aflatoxins B and G and for aflatoxin M1. In Aflatoxins: Recent Advances and Future Prospects; IntechOpen: London, UK, 2013; Volume 15, pp. 315–339. [Google Scholar]
- Chauhan, R.; Solanki, P.R.; Singh, J.; Mukherjee, I.; Basu, T.; Malhotra, B. A novel electrochemical piezoelectric label free immunosensor for aflatoxin B1 detection in groundnut. Food Control 2015, 52, 60–70. [Google Scholar] [CrossRef]
- Moon, J.; Byun, J.; Kim, H.; Lim, E.-K.; Jeong, J.; Jung, J.; Kang, T. On-Site Detection of Aflatoxin B1 in Grains by a Palm-Sized Surface Plasmon Resonance Sensor. Sensors 2018, 18, 598. [Google Scholar] [CrossRef] [Green Version]
- Wacoo, A.P.; Wendiro, D.; Vuzi, P.C.; Hawumba, J.F. Methods for detection of aflatoxins in agricultural food crops. J. Appl. Chem. 2014, 2014, 706291. [Google Scholar] [CrossRef] [Green Version]
- Bhardwaj, H.; Sumana, G.; Marquette, C.A. A label-free ultrasensitive microfluidic surface Plasmon resonance biosensor for Aflatoxin B1 detection using nanoparticles integrated gold chip. Food Chem. 2020, 307, 125530. [Google Scholar] [CrossRef]
- Kaminiaris, M.D.; Mavrikou, S.; Georgiadou, M.; Paivana, G.; Tsitsigiannis, D.I.; Kintzios, S. An Impedance Based Electrochemical Immunosensor for Aflatoxin B1 Monitoring in Pistachio Matrices. Chemosensors 2020, 8, 121. [Google Scholar] [CrossRef]
- Abera, B.D.; Falco, A.; Ibba, P.; Cantarella, G.; Petti, L.; Lugli, P. Development of Flexible Dispense-Printed Electrochemical Immunosensor for Aflatoxin M1 Detection in Milk. Sensors 2019, 19, 3912. [Google Scholar] [CrossRef] [Green Version]
- Benedetti, S.; Iametti, S.; Bonomi, F.; Mannino, S. Head space sensor array for the detection of aflatoxin M1 in raw ewe’s milk. J. Food Prot. 2005, 68, 1089–1092. [Google Scholar] [CrossRef]
- Kasoju, A.; Shahdeo, D.; Khan, A.A.; Shrikrishna, N.S.; Mahari, S.; Alanazi, A.M.; Bhat, M.A.; Giri, J.; Gandhi, S. Fabrication of microfluidic device for Aflatoxin M1 detection in milk samples with specific aptamers. Sci. Rep. 2020, 10, 4627. [Google Scholar] [CrossRef] [Green Version]
- Kasoju, A.; Shrikrishna, N.S.; Shahdeo, D.; Khan, A.A.; Alanazi, A.M.; Gandhi, S. Microfluidic paper device for rapid detection of aflatoxin B1 using an aptamer based colorimetric assay. R. Soc. Chem. 2020, 10, 11843–11850. [Google Scholar] [CrossRef]
- Ahmadi, S.F.; Hojjatoleslamy, M.; Kiani, H.; Molavi, H. Monitoring of Aflatoxin M1 in milk using a novel electrochemical aptasensor based on reduced graphene oxide and gold nanoparticles. Food Chemi. 2022, 373, 131321. [Google Scholar] [CrossRef] [PubMed]
- Espinosa-Calderón, A.; Contreras-Medina, L.M.; Muñoz-Huerta, R.F.; Millán-Almaraz, J.R.; González, R.G.G.; Torres-Pacheco, I. Methods for detection and quantification of aflatoxins. In Aflatoxins; IntechOpen: London, UK, 2011; pp. 109–128. [Google Scholar] [CrossRef]
- Santiago, M.; Strobel, S. Thin layer chromatography. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 2013; Volume 533, pp. 303–324. [Google Scholar] [CrossRef]
- Malviya, R.; Bansal, V.; Pal, O.P.; Sharma, P.K. High performance liquid chromatography: A short review. J. Glob. Pharma Technol. 2010, 2, 22–26. Available online: https://www.researchgate.net/profile/Rishabha-Malviya/publication/235987484_High_performance_liquid_chromatography_A_short_review/links/02e7e5153e680408ac000000/High-performance-liquid-chromatography-A-short-review.pdf (accessed on 19 December 2021).
- Al-Bukhaiti, W.Q.; Noman, A.; Qasim, A.S.; Al-Farga, A. Gas chromatography: Principles, advantages and applications in food analysis. Int. J. Agric. Innov. Res. 2017, 6, 123–128. Available online: https://ijair.org/administrator/components/com_jresearch/files/publications/IJAIR_2467_FINAL.pdf (accessed on 19 December 2021).
- Bashiry, M.; Javanmardi, F.; Sadeghi, E.; Shokri, S.; Hossieni, H.; Oliveira, C.A.; Khaneghah, A.M. The prevalence of aflatoxins in commercial baby food products: A global systematic review, meta-analysis, and risk assessment study. Trends Food Sci. Technol. 2021, 114, 100–115. [Google Scholar] [CrossRef]
- Omar, S.S.; Haddad, M.A.; Parisi, S. Validation of HPLC and Enzyme-Linked Immunosorbent Assay (ELISA) Techniques for Detection and Quantification of Aflatoxins in Different Food Samples. Foods 2020, 9, 661. [Google Scholar] [CrossRef] [PubMed]
- Kortei, N.K.; Annan, T.; Akonor, P.T.; Richard, S.A.; Annan, H.A.; Kyei-Baffour, V.; Akuamoa, F.; Akpaloo, P.G.; Esua-Amoafo, P. The occurrence of aflatoxins and human health risk estimations in randomly obtained maize from some markets in Ghana. Sci. Rep. 2021, 11, 4295. [Google Scholar] [CrossRef]
- Khalil, R.; Yassin, M.; Yaseen, S.J. Detection some Aflatoxins in some locally marketed Raw Peanuts. Res. J. Pharm. Technol. 2021, 14, 1431–1437. [Google Scholar] [CrossRef]
- Jiménez Medina, M.L.; Lafarga, T.; Garrido Frenich, A.; Romero-González, R. Natural Occurrence, Legislation, and Determination of Aflatoxins Using Chromatographic Methods in Food: A Review (from 2010 to 2019). Food Rev. Int. 2021, 37, 244–275. [Google Scholar] [CrossRef]
- Vaz, A.; Cabral Silva, A.C.; Rodrigues, P.; Venâncio, A. Detection Methods for Aflatoxin M1 in Dairy Products. Microorganisms 2020, 8, 246. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Banerjee, K. A Review: Sample Preparation and Chromatographic Technologies for Detection of Aflatoxins in Foods. Toxins 2020, 12, 539. [Google Scholar] [CrossRef] [PubMed]
- Abbas, M. Chromatographic Techniques for Estimation of Aflatoxins in Food Commodities. In Aflatoxins; Intech Open: London, UK, 2021. [Google Scholar]
- AlFaris, N.A.; Wabaidur, S.M.; Alothman, Z.A.; Altamimi, J.Z.; Aldayel, T.S. Fast and efficient immunoaffinity column cleanup and liquid chromatography–tandem mass spectrometry method for the quantitative analysis of aflatoxins in baby food and feeds. J. Sep. Sci. 2020, 43, 2079–2087. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.E.; Gill, B.D.; Indyk, H.E.; Rhemrev, R.; Pazdanska, M.; Mackay, N.; Marley, E. Determination of Aflatoxin M1 in Liquid Milk, Cheese, and Selected Milk Proteins by Automated Online Immunoaffinity Cleanup with Liquid Chromatography—Fluorescence Detection. J. AOAC Int. 2021, 104, 719–724. [Google Scholar] [CrossRef]
- Wu, Q.; Xu, H. Design and development of an on-line fluorescence spectroscopy system for detection of aflatoxin in pistachio nuts. Postharvest Biol. Technol. 2020, 159, 111016. [Google Scholar] [CrossRef]
- Chavez, R.A.; Cheng, X.; Stasiewicz, M.J. A Review of the Methodology of Analyzing Aflatoxin and Fumonisin in Single Corn Kernels and the Potential Impacts of These Methods on Food Security. Foods 2020, 9, 297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, X.; Zhang, Q.; Zhang, W.; Mao, J.; Li, P. Control of Aflatoxigenic Molds by Antagonistic Microorganisms: Inhibitory Behaviors, Bioactive Compounds, Related Mechanisms, and Influencing Factors. Toxins 2020, 12, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cotty, P.; Bayman, P. Competitive exclusion of a toxigenic strain of Aspergillus flavus by an atoxigenic strain. Phytopathology 1993, 83, 1283–1287. Available online: https://www.apsnet.org/publications/phytopathology/backissues/Documents/1993Articles/Phyto83n12_1283.PDF (accessed on 19 December 2021). [CrossRef]
- Okun, D.O.; Khamis, F.M.; Muluvi, G.M.; Ngeranwa, J.J.; Ombura, F.O.; Yongo, M.O.; Kenya, E.U. Distribution of indigenous strains of atoxigenic and toxigenic Aspergillus flavus and Aspergillus parasiticus in maize and peanuts agro-ecological zones of Kenya. Agric. Food Secur. 2015, 4, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Cotty, P.J. Development of Aspergillus Flavus AF36. 2005. Available online: https://biopesticide.ucr.edu/abstracts/assets/Cotty_abstract.pdf (accessed on 19 December 2021).
- Dorner, J.; Lamb, M.C. Development and commercial use of afla-Guard®, an aflatoxin biocontrol agent. Mycotoxin Res. 2006, 22, 33–38. [Google Scholar] [CrossRef]
- Abbas, H.K.; Zablotowicz, R.M.; Bruns, H.A.; Abel, C.A. Biocontrol of aflatoxin in corn by inoculation with non-aflatoxigenic Aspergillus flavus isolates. Biocontrol Sci. Technol. 2006, 16, 437–449. [Google Scholar] [CrossRef]
- Mauro, A.; Garcia-Cela, E.; Pietri, A.; Cotty, P.J.; Battilani, P. Biological Control Products for Aflatoxin Prevention in Italy: Commercial Field Evaluation of Atoxigenic Aspergillus flavus Active Ingredients. Toxins 2018, 10, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senghor, A.; Ortega-Beltran, A.; Atehnkeng, J.; Jarju, P.; Cotty, P.; Bandyopadhyay, R. Aflasafe SN01 is the first biocontrol product approved for aflatoxin mitigation in two nations, Senegal and The Gambia. Plant Dis. 2021, 105, 1461–1473. [Google Scholar] [CrossRef] [PubMed]
- Agbetiameh, D.; Ortega-Beltran, A.; Awuah, R.T.; Atehnkeng, J.; Elzein, A.; Cotty, P.J.; Bandyopadhyay, R. Field efficacy of two atoxigenic biocontrol products for mitigation of aflatoxin contamination in maize and groundnut in Ghana. Biol. Control 2020, 150, 104351. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, R.; Ortega-Beltran, A.; Akande, A.; Mutegi, C.; Atehnkeng, J.; Kaptoge, L.; Senghor, A.; Adhikari, B.; Cotty, P.J. Biological control of aflatoxins in Africa: Current status and potential challenges in the face of climate change. World Mycotoxin J. 2016, 9, 771–789. [Google Scholar] [CrossRef] [Green Version]
- Zanon, M.S.A.; Barros, G.G.; Chulze, S.N. Non-aflatoxigenic Aspergillus flavus as potential biocontrol agents to reduce aflatoxin contamination in peanuts harvested in Northern Argentina. Int. J. Food Microbiol. 2016, 231, 63–68. [Google Scholar] [CrossRef]
- Xu, D.; Wang, H.; Zhang, Y.; Yang, Z.; Sun, X. Inhibition of non-toxigenic Aspergillus niger FS10 isolated from Chinese fermented soybean on growth and aflatoxin B1 production by Aspergillus flavus. Food Control 2013, 32, 359–365. [Google Scholar] [CrossRef]
- Yin, Y.-n.; Yan, L.-y.; Jiang, J.-h.; Ma, Z.H. Biological control of aflatoxin contamination of crops. J. Zhejiang Univ. Sci. 2008, 9, 787–792. [Google Scholar] [CrossRef]
- Konlambigue, M.; Ortega-Beltran, A.; Bandyopadhyay, R.; Shanks, T.; Landreth, E.; Jacob, O. Lessons Learned on Scaling Aflasafe® through Commercialization in Sub-Saharan Africa; Intl Food Policy Res Inst: Washington, DC, USA, 2020. [Google Scholar] [CrossRef]
- Mahuku, G.S. AR-NAFAKA Project Aflatoxin Management: 2016–2017 Progress. In Proceedings of the Africa RISING—NAFAKA Scaling Project End-of-Project Phase Review Meeting; Dar es Salaam, Tanzania, 3–4 July 2017, International Institute of Tropical Agriculture: Ibadan, Nigeria, 2017; Available online: https://cgspace.cgiar.org/bitstream/handle/10568/89001/ar_mahuku_jul2017.pdf?sequence=1&isAllowed=y (accessed on 19 December 2021).
- Atehnkeng, J.; Donner, M.; Ojiambo, P.S.; Ikotun, B.; Augusto, J.; Cotty, P.J.; Bandyopadhyay, R. Environmental distribution and genetic diversity of vegetative compatibility groups determine biocontrol strategies to mitigate aflatoxin contamination of maize by Aspergillus flavus. Microb. Biotechnol. 2016, 9, 75–88. [Google Scholar] [CrossRef] [Green Version]
- Grubisha, L.C.; Cotty, P. Genetic analysis of the Aspergillus flavus vegetative compatibility group to which a biological control agent that limits aflatoxin contamination in US crops belongs. ASM 2015, 81, 5889–5899. [Google Scholar] [CrossRef] [Green Version]
- Abbas, H.K.; Accinelli, C.; Shier, W.T. Biological control of aflatoxin contamination in US crops and the use of bioplastic formulations of Aspergillus flavus biocontrol strains to optimize application strategies. J. Agric. Food Chem. 2017, 65, 7081–7087. [Google Scholar] [CrossRef]
- Li, Q.Z.; He, Z.M. Advances in research of the structural gene characteristics of the aflatoxin biosynthetic gene cluster. J. Plant Sci. Phytopathol. 2018, 2, 68–82. [Google Scholar] [CrossRef] [Green Version]
- Donner, M.; Atehnkeng, J.; Sikora, R.; Bandyopadhyay, R.; Cotty, P. Molecular characterization of atoxigenic strains for biological control of aflatoxins in Nigeria. Food Addit. Contam. 2010, 27, 576–590. [Google Scholar] [CrossRef] [PubMed]
- Callicott, K.; Cotty, P. Method for monitoring deletions in the aflatoxin biosynthesis gene cluster of Aspergillus flavus with multiplex PCR. Lett. Appl. Microbiol. 2015, 60, 60–65. [Google Scholar] [CrossRef]
- Pitt, J.; Hocking, A.D. Mycotoxins in Australia: Biocontrol of aflatoxin in peanuts. Mycopathologia 2006, 162, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Xing, F.; Wang, L.; Liu, X.; Selvaraj, J.N.; Wang, Y.; Zhao, Y.; Liu, Y. Aflatoxin B1 inhibition in Aspergillus flavus by Aspergillus niger through down-regulating expression of major biosynthetic genes and AFB1 degradation by atoxigenic A. flavus. Int. J. Food Microbiol. 2017, 256, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Peles, F.; Sipos, P.; Kovács, S.; Győri, Z.; Pócsi, I.; Pusztahelyi, T. Biological Control and Mitigation of Aflatoxin Contamination in Commodities. Toxins 2021, 13, 104. [Google Scholar] [CrossRef]
- Mostafa, A.A.; Al-Rahmah, A.N.; Abdel-Megeed, A.; Sayed, S.R.; Hatamleh, A.A. Antagonistic activities of some fungal strains against the toxigenic Aspergillus flavus isolate and its aflatoxins productivity. J. Pure Appl. Microbiol. 2013, 7, 169–178. Available online: https://www.researchgate.net/profile/Ashraf-A-Mostafa/publication/287535880_Antagonistic_Activities_of_Some_Fungal_Strains_against_the_Toxigenic_Aspergillus_flavus_Isolate_and_its_Aflatoxins_Productivity/links/5fa6b86f458515157bf45a72/Antagonistic-Activities-of-Some-Fungal-Strains-against-the-Toxigenic-Aspergillus-flavus-Isolate-and-its-Aflatoxins-Productivity.pdf (accessed on 19 December 2021).
- Bernáldez, V.; Rodríguez, A.; Martín, A.; Lozano, D.; Córdoba, J.J. Development of a multiplex qPCR method for simultaneous quantification in dry-cured ham of an antifungal-peptide Penicillium chrysogenum strain used as protective culture and aflatoxin-producing moulds. Food Control 2014, 36, 257–265. [Google Scholar] [CrossRef]
- Kong, Q.; Chi, C.; Yu, J.; Shan, S.; Li, Q.; Li, Q.; Guan, B.; Nierman, W.C.; Bennett, J.W. The inhibitory effect of Bacillus megaterium on aflatoxin and cyclopiazonic acid biosynthetic pathway gene expression in Aspergillus flavus. Appl. Microbiol. Biotechnol. 2014, 98, 5161–5172. [Google Scholar] [CrossRef]
- Siahmoshteh, F.; Hamidi-Esfahani, Z.; Spadaro, D.; Shams-Ghahfarokhi, M.; Razzaghi-Abyaneh, M. Unraveling the mode of antifungal action of Bacillus subtilis and Bacillus amyloliquefaciens as potential biocontrol agents against aflatoxigenic Aspergillus parasiticus. Food Control 2018, 89, 300–307. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, Q.; Chen, Z.-Y.; Liu, H.; Li, P. Investigation of Pseudomonas fluorescens strain 3JW1 on preventing and reducing aflatoxin contaminations in peanuts. PLoS ONE 2017, 12, e0178810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akocak, P.B.; Churey, J.J.; Worobo, R.W. Antagonistic effect of chitinolytic Pseudomonas and Bacillus on growth of fungal hyphae and spores of aflatoxigenic Aspergillus flavus. Food Biosci. 2015, 10, 48–58. [Google Scholar] [CrossRef]
- Mannaa, M.; Oh, J.; Kim, K.D. Microbe-mediated control of Aspergillus flavus in stored rice grains with a focus on aflatoxin inhibition and biodegradation. Ann. Appl. Biol. 2017, 171, 376–392. [Google Scholar] [CrossRef]
- Ahlberg, S.; Joutsjoki, V.; Laurikkala, S.; Varmanen, P.; Korhonen, H. Aspergillus flavus growth inhibition by Lactobacillus strains isolated from traditional fermented Kenyan milk and maize products. Arch. Microbiol. 2017, 199, 457–464. [Google Scholar] [CrossRef]
- Gerez, C.L.; Torres, M.J.; Font de Valdez, G.; Rollán, G. Control of spoilage fungi by lactic acid bacteria. Biol. Control 2013, 64, 231–237. [Google Scholar] [CrossRef]
- Ghanbari, R.; Molaee Aghaee, E.; Rezaie, S.; Jahed Khaniki, G.; Alimohammadi, M.; Soleimani, M.; Noorbakhsh, F. The inhibitory effect of lactic acid bacteria on aflatoxin production and expression of aflR gene in Aspergillus parasiticus. J. Food Saf. 2018, 38, e12413. [Google Scholar] [CrossRef]
- Wang, K.; Yan, P.S.; Cao, L.-x.; Ding, Q.-l.; Shao, C.; Zhao, T.F. Potential of chitinolytic Serratia marcescens strain JPP1 for biological control of Aspergillus parasiticus and aflatoxin. Biomed Res. Int. 2013, 2013, 397142. [Google Scholar] [CrossRef] [Green Version]
- Sipos, P.; Peles, F.; Brassó, D.L.; Béri, B.; Pusztahelyi, T.; Pócsi, I.; Győri, Z. Physical and Chemical Methods for Reduction in Aflatoxin Content of Feed and Food. Toxins 2021, 13, 204. [Google Scholar] [CrossRef]
- Tahir, N.I.; Hussain, S.; Javed, M.; Rehman, H.; Shahzady, T.G.; Parveen, B.; Ali, K.G. Nature of aflatoxins: Their extraction, analysis, and control. J. Food Saf. 2018, 38, e12561. [Google Scholar] [CrossRef]
- Samarajeewa, U.; Sen, A.; Cohen, M.; Wei, C. Detoxification of aflatoxins in foods and feeds by physical and chemical methods. J. Food Prot. 1990, 53, 489–501. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
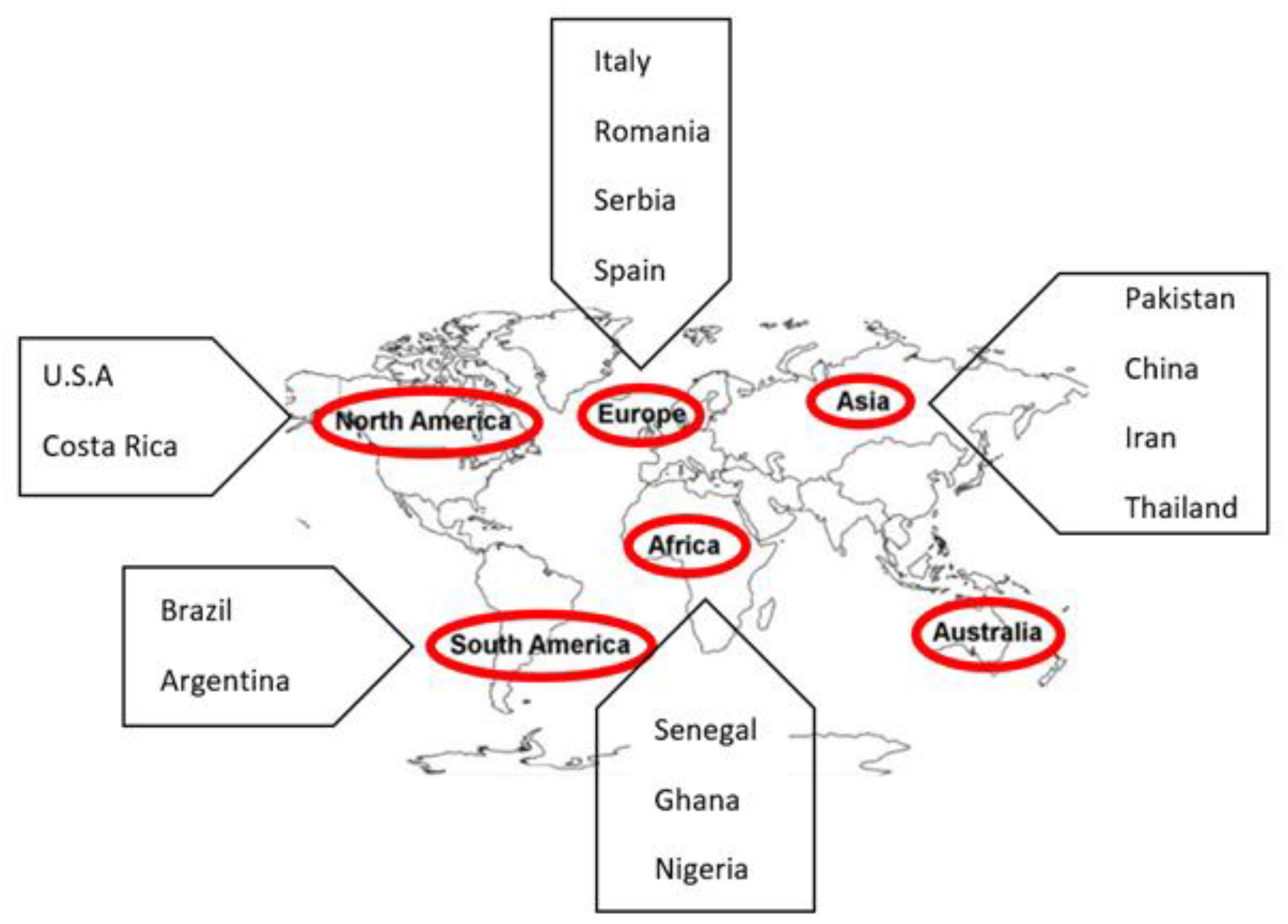{kind=link}
| Aflatoxin | Aflatoxin Producing Aspergillus spp. | Host/Affected Entity |
|---|---|---|
| B (B1, B2) | A. arachidicola, A. bombycis, A. flavus, A. minisclerotigenes, A. nomius, A. ochraceoroseus, A. oryzae, A. parasiticus, A. parvisclerotigenus, A. pseudotamarii, A. rambellii, A. tamarii, A. toxicarius, A. versicolor, Emericella astellata, E. venezuelensis. | Cottonseed, Dairy products, Figs, Fruit juices (apple, guava), Maize, Maize flour, Meat, Oilseed rape, Peanuts, Peanut butter, Pea, Pistachio, Rice, Sorghum, Sunflower seed, Spices |
| B2a | Hydroxylated metabolite of aflatoxin B1 | - |
| B3 (Parasiticol) | Aflatoxin G1 metabolite, naturally produced by: A. flavus, A. mottae, A. nomius, A. novoparasiticus, A. parasiticus | Same as aflatoxin B1 and G1 |
| G (G1, G2) | A. arachidicola, A. bombycis, A. minisclerotigenes. A. nomius, A. parasiticus, A. pseudotamarii, A. terreus, A. toxicarius, A. versicolor | Cottonseed, Dairy products, Figs, Fruit juices (apple, guava), Maize, Maize flour, Meat, Oilseed rape, Peanuts, Peanut butter, Pea, Pistachio, Rice, Sorghum, Sunflower seed, Spices |
| G2a | Hydroxylated metabolite of aflatoxin G1, also naturally produced by A. flavus | - |
| M (M1, M2) | Hydroxylated metabolite of aflatoxin B1 and B2, respectively | Dairy products, Milk, Meat |
| M2a | Aflatoxin M1 derivative | Dairy products, Milk |
| GM1 | Hydroxylated metabolite of aflatoxin G1, naturally produced by A. flavus, also produced by A. parasiticus in vitro | Dairy products, Milk |
| GM2 | Naturally produced by A. flavus and A. parasiticus and yeast, derived from aflatoxin G2 | Dairy products, Milk |
| GM2a | Aflatoxin GM1 metabolite | Dairy products, Milk |
| P1 | Metabolite ofaflatoxin B1 (demethylated) | Dairy products, Excreted in animals and human urine |
| Q1 | Metabolite of aflatoxin B1 (hydroxylated) | Present in the meat of cattle that feeds on aflatoxin-contaminated food |
| Q2a | Acid hydration of aflatoxin Q1 | - |
| Aflatoxicol R0 | Metabolite of aflatoxin B1, also naturally produced by A. flavus and A. parasiticus | Present in bird feed, also exists in birds that feed on aflatoxin-contaminated food |
| Aflatoxicol M1 | Aflatoxin B1, aflatoxin R0, or aflatoxin M1 metabolite | Dairy products, Milk |
| Aflatoxicol H1 | Aflatoxin B1 and aflatoxin Q1 metabolite | Dairy products, Milk |
| Aspertoxin | A. flavus and A. parasiticus | Crops and Plants |
| S. No | Primer | Amplified Gene | Sequence | Size (bp) | References |
|---|---|---|---|---|---|
| 1. | nor1 | nor-1 | 5′-ACCGCTACGCCGGCACTCTCGGCAC-3′ | 400 | [45] |
| nor2 | - | 5′-GTTGGCCGCCAGCTTCGACACTCCG-3′ | - | ||
| 2. | ver1 | ver-1/aflM | 5′-GCCGCACGCGGAGAAAGTGGT-3′ | 537 | [45] |
| ver2 | - | 5′-GGGGATATACTCCCGCGACACAGCC-3′ | - | ||
| 3. | omt1 | omtA/aflP | 5′-GTGGACGGACCTAGTCCGACATCAC-3′ | 797 | [46] |
| omt2 | - | 5′-GTCGGCGCCACGCACTGGGTTGGGG-3′ | - | ||
| 4. | Omt 208 | omtA | 5′-GGCCCGGTTCCTTGGCTCCTAAGC-3′ | 1024 | [47] |
| Omt-1232 | - | 5′-CGCCCCAGTGAGACCCTTCCTCG-3′ | - | ||
| 5. | VER-496 | ver-1 | 5′-ATGTCGGATAATCACCGTTTAGATGGC-3′ | 895 | [47] |
| VER-1391 | - | 5′-CGAAAAGCGCCACCATCCACCCCAATG-3′ | - | ||
| 6. | APA-450 | apa-2 | 5′-TATCTCCCCCCGGGCATCTCCCGG -3′ | 1032 | [47] |
| APA-1482 | - | 5′-CCGTCAGACAGCCACTGGACACGG-3′ | - | ||
| 7. | aflR660 | aflR | 5′-CGCGCTCCCAGTCCCCTTCATT-3′ | 630 | [45] |
| aflR1249 | - | 5′-CTTGTTCCCCGAGATGACCA-3′ | - | ||
| 8. | ord1501 | ord1 | 5′-TTAAGGCAGGGGAATACAAG -3′ | 610 | [48] |
| ord2226 | - | 5′-GACGCCCAAAGCCGAACACAAA-3′ | - | ||
| 9. | tub440-F | ß-tubulin | 5′-GGTAACCAAATAGGTGCCGCT -3′ | 1300 | [49] |
| tub1740-R | - | 5′-TAGGTCTGGTTCTTGCTCTGGATG-3′ | - | ||
| 10. | nortaq-1 | nor-1 | 5′-GTCCAAGCAACAGGCCAAGT -3′ | 66 | [50] |
| nortaq-2 | - | 5′-TCGTGCATGTTGGTGATGGT-3′ | - | ||
| norprobe | - | 5′-TGTCTTGATCGGCGCCCG-3′ | - | ||
| 11. | aflR1-F | aflR | 5′-AACCGCATCCACAATCTCAT-3′ | 798 | [45] |
| aflR1-R | - | 5′-AGTGCAGTTCGCTCAGAACA-3′ | - | ||
| 12. | Tub1-F | tub1 | 5′-GTCCGGTGCTGGTAACAACT -3′ | 1498 | [45] |
| Tub1-R | - | 5′-GGAGGTGGAGTTTCCAATGA-3′ | - | ||
| 13. | Nor1-F | aflD | 5′-ACGGATCACTTAGCCAGCAC-3′ | 990 | [51] |
| NoR1-R | - | 5′-CTACCAGGGGAGTTGAGATCC-3′ | - | ||
| 14. | OmtB(F)-F | aflO | 5′-GCCTTGACATGGAAACCATC-3′ | 1333 | [51] |
| OmtB(F)-R | - | 5′-CCAAGATGGCCTGCTCTTTA-3′ | - | ||
| 15. | Ord-gF | aflQ | 5′-TTAAGGCAGCGGAATACAAG-3′ | 719 | [51] |
| Ord-gR | - | 5′-GACGCCCAAAGCCGAACACAAA-3′ | - | ||
| 16. | Omt1-F | aflP | 5′-GCCTTGCAAACACACTTTCA-3′ | 1490 | [52] |
| Omt1R | - | 5′-AGTTGTTGAACGCCCCAGT-3′ | - | ||
| 17. | aflR-F1 | aflR | 5′-TGACCCACCTCTTCCCCCACG-3′ | 300 | [51] |
| aflR-R | - | 5′-CCGTCAGACAGCCACTGGACACGG-3′ | - | ||
| 18. | aflj-F/AP-F | aflJ | 5′-AGTCAAAGGTTGAATACC-3′ | 840 | [53] |
| aflj-R/AP-R | - | 5′-GCTCAGCCATGACCTTGACTG-3′ | - | ||
| 19. | omtBII-F | omt-B | 5′-ATGTGCTTGGGITGCTGTGG-3′ | 611 | [54] |
| omtBII-R | - | 5′-GGATGTGGTYATGCGATTGAG-3′ | - | ||
| 20. | AF138287 | ITS1-5. 8rRNA | 5′-CTCCCACCCGTGTTTACTGT-3′ | 199 | [55] |
| AF027863 | - | 5′-GCGTTCTTCATCGATGCCT-3′ | - | ||
| 21. | Asp1S | 5.8-28S rDNA | 5′-ATGCCTGTCCGAGCGT-3′ | - | [56] |
| AflR2 | - | 5′-TTAAGTTCAGCGGGTATRCCb-3′ | - | ||
| 22. | AflP-F | aflP | 5′-CATGCTCCATCATGGTGACT-3′ | - | [4] |
| AflP-R | - | 5′-CCGCCGCTTTGATCTAGG-3′ | - | ||
| 23. | FVAVIQ1 | ITS2 rDNA | 5′-GTCGTCCCCTCTCCGG-3′ | - | [57] |
| FLAQ2 | - | 5′-CTGGAAAAAGATTGATTTGCG-3′ | - | ||
| PARQ2 | - | 5′-GAAAAAATGGTTGTTTTGCG-3′ | - | ||
| 24. | cmd424 | Calmodulin | 5′-GGCCTTCTCCCTATTCGTAA-3′ | 613 | [58] |
| cmd6374 | - | 5′-CTCGCGGATCATCTCATC-3′ | - | ||
| 25. | cmd2F3 | Calmodulin | 5′-GGCTGGATGTGTGTAAATC-3′ | 811 | [58] |
| cmd2R3 | Calmodulin | 5′-ATTGGTCGCATTTGAAGGG-3′ | - | ||
| 26. | niaDF3 | niaD | 5′-CGGACGATAAGCAACAACAC-3′ | 795 | [58] |
| niaDAR3 | - | 5′-GGATGAACACCCGTTAATCTGA-3′ | - | ||
| 27. | niaDBF3 | niaD | 5′-ACGGCCGACAGAAGTGCTGA-3′ | 794 | [58] |
| niaDBR3 | niaD | 5′-TGGGCGAAGAGACTCCCCGT-3′ | - | ||
| 28. | niaDCF | Nitrate reductase | 5′-GCAGCCCAATGGTCACTACGGC-3′ | - | [58] |
| niaDCR | Nitrate reductase | 5′-GGCTGCACGCCCAATGCTTC-3′ | - | ||
| 29. | AP17295 | norB-cypA | 5′-GTGCCCAGCATCTTGGTCCACC-3′ | 1839(no deletion) 903 (L strain) 323 (L+S strain) | [59] |
| AP35515 | - | 5′-AAGGACTTGATGATTCCTC-3′ | - | ||
| 30. | CP-5F6 | norB-cypA | 5′-GGGACCCTTTTCCGGTGCGG-3′ | 3053(no deletion) 2134 (L strain) 1549(L+S strain) 836(LAF)7 | [60] |
| CP-R6 | - | 5′-GGCGGCCCCTCAGCAAACAT-3′ | - | ||
| 31. | Taka-amylaseF8 | amyB/amy1 | 5′-GGATCGATTTGCAAGGACGG-3′ | 1168 | [61] |
| Taka-amylaseR8 | - | 5′-TAGAGGTCGTCCATGCTGCC-3′ | - |
| S. No | Product/Strain Name | Country | References |
|---|---|---|---|
| 1. | AF36 | U.S. | [97] |
| 2. | Afla-Guard (strain NRRL21882) | U.S. | [98] |
| 3. | CT3 (unregistered) | Southern U.S. | [99] |
| 4. | K49 (unregistered) | Southern U.S. | [99] |
| 5. | AF-X1 | Italy | [100] |
| 6. | Aflasafe SN01 | Senegal and The Gambia | [101] |
| 7. | Aflasafe GH01 | Ghana | [102] |
| 8. | Aflasafe GH02 | Ghana | [102] |
| 9. | Aflasafe | Nigeria | [103] |
| 10. | Aflasafe KE01 | Kenya | [103] |
| 11. | AR27 (unregistered) | Northern Argentina | [104] |
| 12. | AR100G (unregistered) | Northern Argentina | [104] |
| 13. | AFCHG2 (unregistered) | Northern Argentina | [104] |
| 14. | FS10 (unregistered) | China | [105] |
| 15. | AF051 (unregistered) | China | [106] |
| 16. | BN30 (unregistered) | Africa | [106] |
| 17. | Aflasafe BF01 | Burkina Faso | [107] |
| 18. | Aflasafe TZ01 | Tanzania | [108] |
| 19. | Aflasafe TZ02 | Tanzania | [108] |
| 20. | Aflasafe ZM01 & ZM02 | Zambia | [108] |
| 21. | Aflasafe MW01 & MWMZ01 | Malawi | [108] |
| 22. | Aflasafe MZ01 & MWMZ01 | Mozambique | [108] |
| S. no | Panel | Marker | Sequence | Size (bp) |
|---|---|---|---|---|
| 01. | Sugar Cluster | SC01 | 5′-ATACCTCATGATCTGGTGCACGG 5′-CTTCGCAGCGACAATGATACGTC | 883 |
| 02. | IC01 | 5′-GTCCCCAGGTACGATAGGTCTCT 5′-GCTGGATATTCCAAGGAGTGGCT | 742 | |
| 03. | AC01 | 5′-GACTGCCACCCTATCACTCTTCC 5′-TGGCTCGACTGGGTATGAAATCC | 613 | |
| 04. | AC02 | 5′-GCATTGCCAGCATCGGTTTCATA 5′-AGGCAGACCGTACTAAGTGATGC | 487 | |
| 05. | AC03 | 5′-CATGATGGAGCATGACATTCGGC 5′-GCGCCACCATATCTTCTCAGTCT | 387 | |
| 06. | AC04 | 5′-TTTAACCCTTCAYGCCTCGAACT 5′-TGCGTARCTAATCTCATCGGGTT | 297 | |
| 07. | AC05 | 5′-TGCTGAGCGAGTAGGTAGTAGGT 5′-CCGGATCATCCCTCCAAATCTGT | 194 | |
| 08. | iac | 5′-GCTAGGGCGGGTCACGTTTTGCG 5′-GGCGTTGTTTAAGGGGAACCGACCC | 115 | |
| 09. | Aflatoxin Cluster | AC06 | 5′-CCTGTGAGGGACACAAAGACACT 5′-AAGAATAGCGGTGACATCCAGCA | 1427 |
| 10. | AC07 | 5′-GAGGACAGGTTGTGTTGCTGTTG 5′-GTTCACGAGCTATCCTCAGCCAT | 1092 | |
| 11. | AC08 | 5′-GAACTGAGCCATTTCCATCAGCG 5′-GTCTTGTACAGGGAACGTGGTGA | 897 | |
| 12. | AC09 | 5′-AACGCTTCAACGTGGAGGACATA 5′-AATAGCGTTGGCGTTGAAGTCAC | 736 | |
| 13. | AC10 | 5′-CCCGCATTTTTCTCGATCCCTTG 5′-GCGACGACCAGTCATTATGAAGC | 633 | |
| 14. | AC11 | 5′-GTCAGACCACAGTGAGTGCTTCT 5′-AAGCTGACTGGGAGAATGTTGCT | 536 | |
| 15. | AC12 | 5′-CCCCTCAACTTCTGTCGTCCTAC 5′-GCTGGGTAGCGAACAATCCAATG | 425 | |
| 16. | AC13 | 5′-GCACACAGCAGAGGCATTTCTAC 5′-AATCTATCTAGCCATCGCCACCG | 330 | |
| 17. | IC02 | 5′-GCCTGCTAGGCTTGGAACTATGT 5′-CGCAATGCTAGTATGCCCTTGTC | 209 | |
| 18. | iac | 5′-GCTAGGGCGGGTCACGTTTTGCG 5′-GGCGTTGTTTAAGGGGAACCGACCC | 115 | |
| 19. | CPA Cluster | CC01 | 5′-GACACTCGTACCATCTATGCACC 5′-GATCCCTGATCCATTCCACCTTG | 1219 |
| 20. | CC02 | 5′-ACGATACGAGCTTTAGTGCAAGG 5′-GATATAGACCTCAGGGTCGAGCA | 925 | |
| 21. | CC03 | 5′-AGAGCTGCGCACTCCATTT 5′-TGCCCAGGCAATAGGAAGTA | 821 | |
| 22. | CC04 | 5′-ACCTCAACAATTACACCGGATGG 5′-GTTGTAGCTCAACGTCACTAGCA | 648 | |
| 23. | ST01 | 5′-TATCTATCTGGGATACGGGCTGG 5′-TATGCCGTTGCTATCCAATGAGG | 521 | |
| 24. | ST02 | 5′-AAGTCAGATTCCGCGGTATGAAG 5′-TCATCGCATTAATCGAGGCAGTT | 416 | |
| 25. | ST03 | 5′-CCTCCTGCACAAAAATACTCCCA 5′-GATCAGATCTTTGAGCGTAGCGT | 320 | |
| 26. | ST04 | 5′-TCATGTTTCGGATCGGAGATTGG 5′-ACATTCCAAGTGAGAGATGTGGC | 234 | |
| 27. | iac | 5′-GCTAGGGCGGGTCACGTTTTGCG 5′-GGCGTTGTTTAAGGGGAACCGACCC | 115 | |
| 28. | Sub-telomere | ST05 | 5′-ACTGGTGTTGGATAGAGCTCAGA 5′-TGGAAGGTTCTCCGGATACTTGA | 908 |
| 29. | ST06 | 5′-TACTCCGTTGCTGTCATTGGATG 5′-CGAATTCTTGGTTGAGCAGCTTG | 782 | |
| 30. | ST07 | 5′-TGCTGAATAACAACCTCGACCAG 5′-CAGGCTGGTATAGCACCAATGTT | 684 | |
| 31. | ST08 | 5′-GGTTTCGTCTTGCCTTCTTCTCA 5′-AGCAAAGTGATGCCGTTCAAATG | 584 | |
| 32. | ST09 | 5′-CGTACTTTGTTACGGCGTACATC 5′-GCTGTTTCGCGTTAGTTGGTAAC | 512 | |
| 33. | ST10 | 5′-GCCCGTAAATGAGGTGCAGATAA 5′-TTTGGGTGTGCTTCTTCATGCTA | 404 | |
| 34. | ST11 | 5′-GGGGACTTAGTCGCGAATGGTTA 5′-TATGAAGGCCACCAACTGAGGAC | 285 | |
| 35. | ST12 | 5′-AATGACGACACTTGAGGCACAG 5′-TCGGCTCCGTGACACCATATTA | 185 | |
| 36. | iac | 5′-GCTAGGGCGGGTCACGTTTTGCG 5′-GGCGTTGTTTAAGGGGAACCGACCC | 115 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shabeer, S.; Asad, S.; Jamal, A.; Ali, A. Aflatoxin Contamination, Its Impact and Management Strategies: An Updated Review. Toxins 2022, 14, 307. https://doi.org/10.3390/toxins14050307
Shabeer S, Asad S, Jamal A, Ali A. Aflatoxin Contamination, Its Impact and Management Strategies: An Updated Review. Toxins. 2022; 14(5):307. https://doi.org/10.3390/toxins14050307
Chicago/Turabian StyleShabeer, Saba, Shahzad Asad, Atif Jamal, and Akhtar Ali. 2022. "Aflatoxin Contamination, Its Impact and Management Strategies: An Updated Review" Toxins 14, no. 5: 307. https://doi.org/10.3390/toxins14050307
APA StyleShabeer, S., Asad, S., Jamal, A., & Ali, A. (2022). Aflatoxin Contamination, Its Impact and Management Strategies: An Updated Review. Toxins, 14(5), 307. https://doi.org/10.3390/toxins14050307