Recovery from the Neuroparalysis Caused by the Micrurus nigrocinctus Venom Is Accelerated by an Agonist of the CXCR4 Receptor

 , , ,
, , ,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
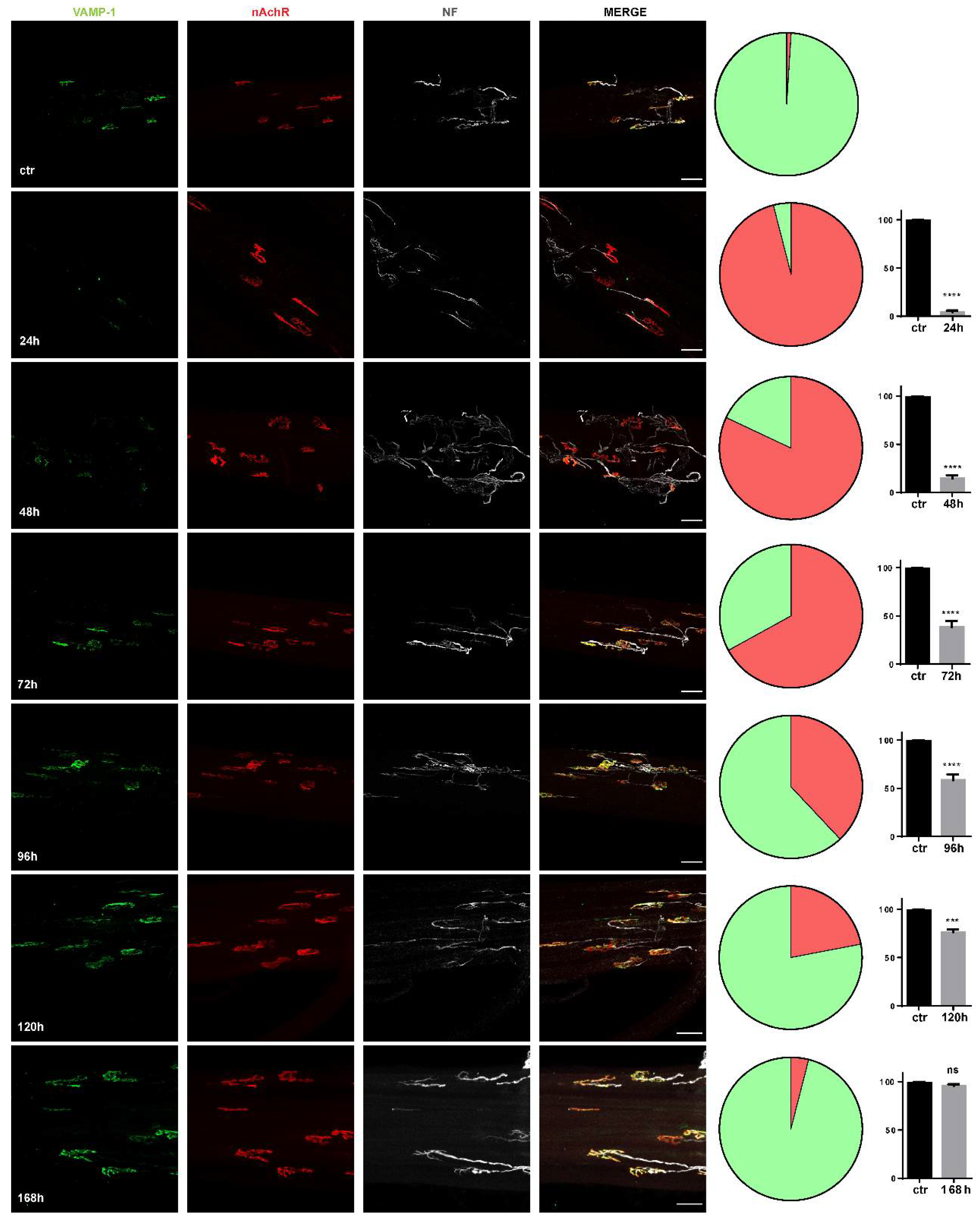
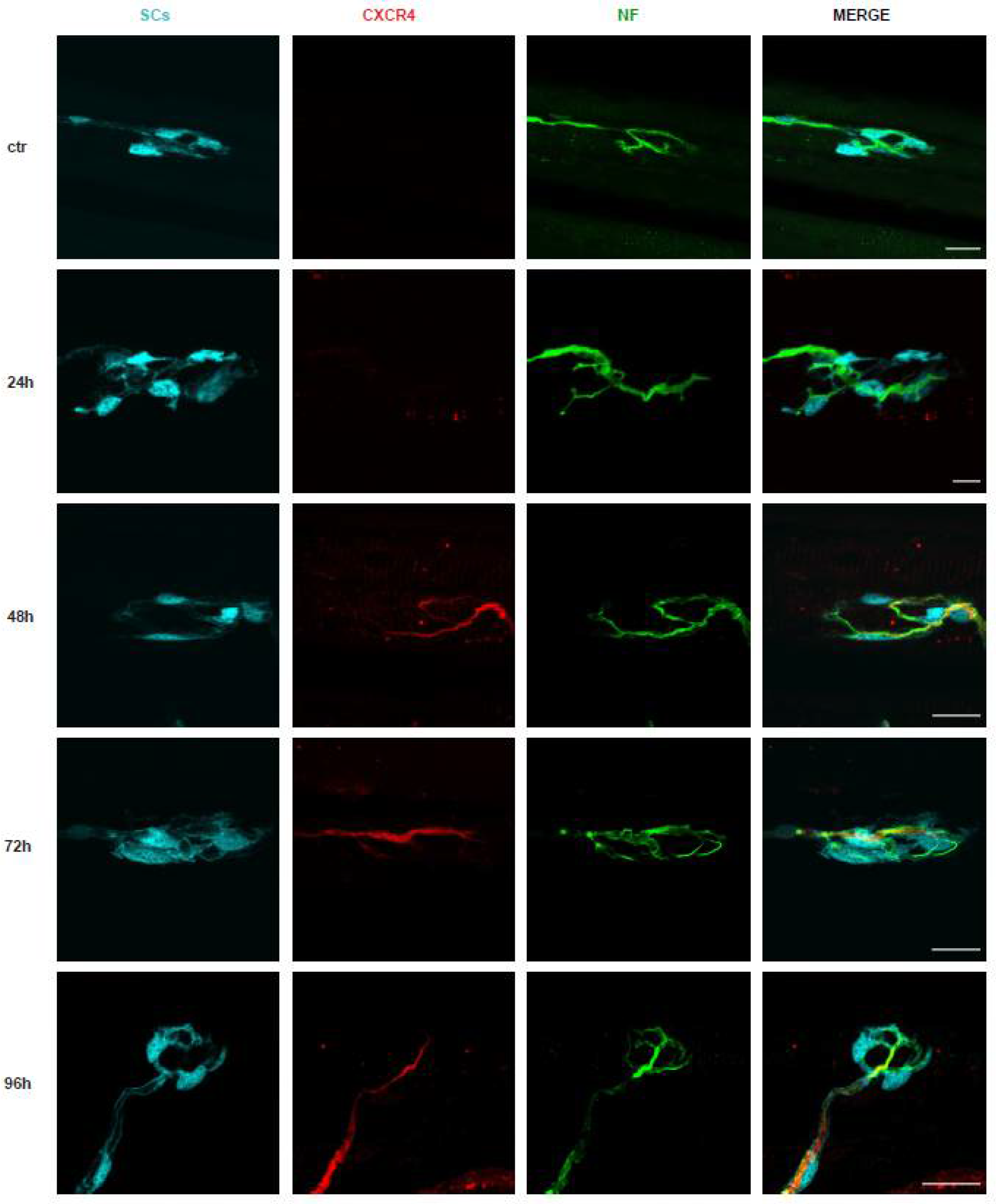
2.1. The Degeneration of the Motor Axon Terminals Induced by M. nigrocinctus Venom Is Accompanied by Expression of the CXCR4 Receptor
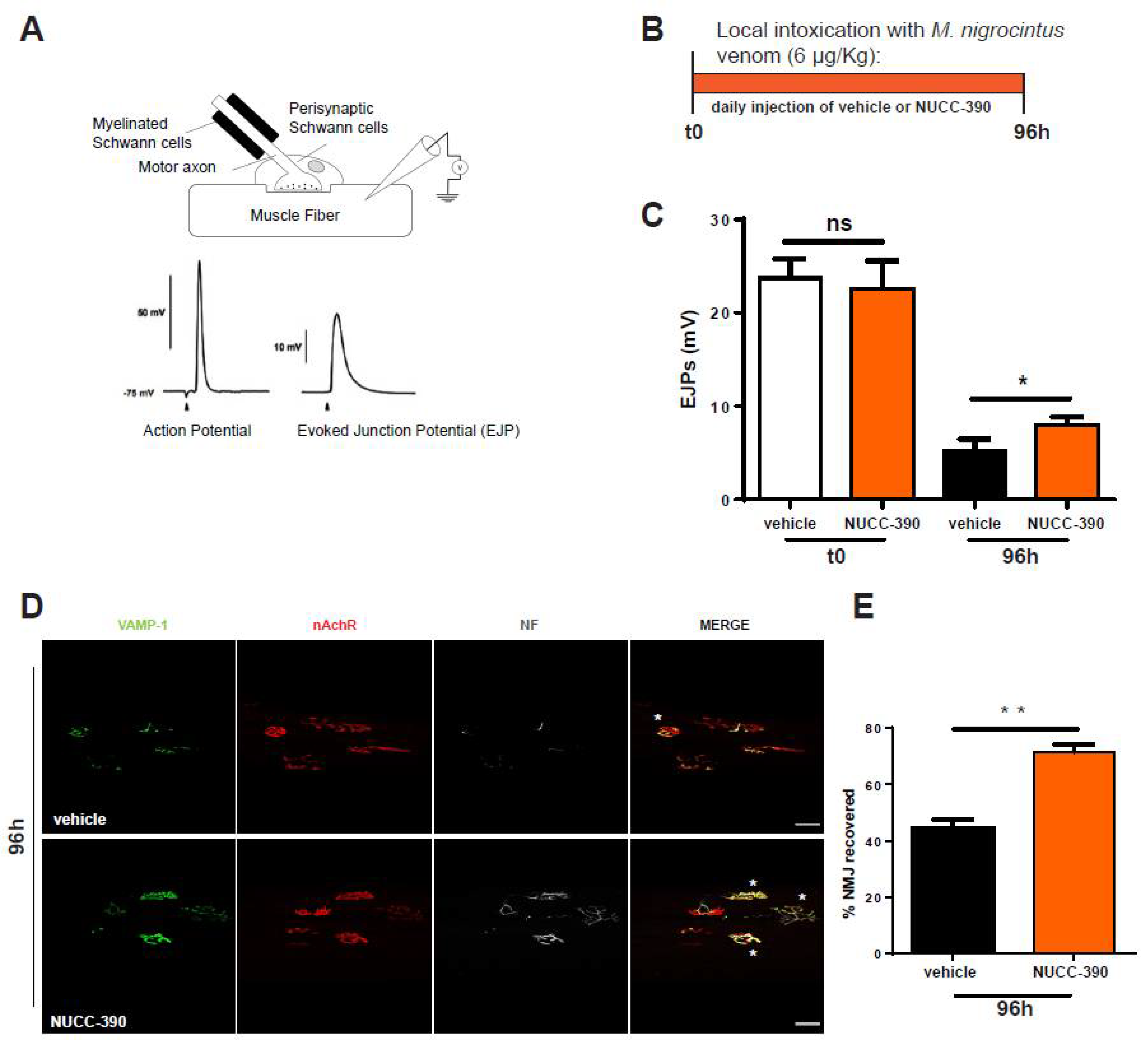
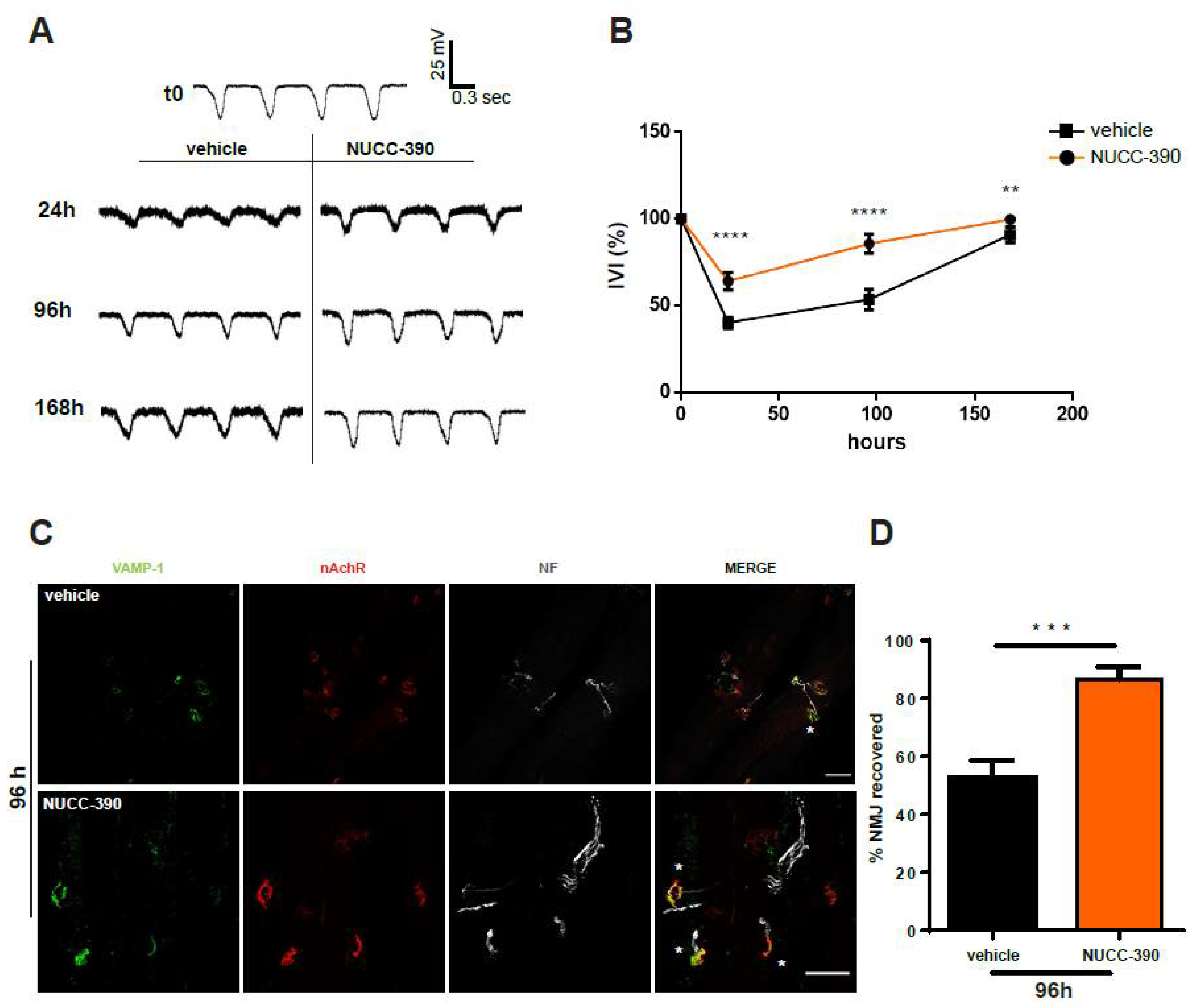
2.2. NUCC-390 Stimulates the Recovery of the NMJ Morphology and Function after Degeneration of the Motor Axon Terminals Induced by M. nigrocinctus Venom
2.3. NUCC-390 Stimulates Rehabilitation of the Neuromuscular Junction after Degeneration of the Axon Terminals Induced by M. nigrocinctus Venom
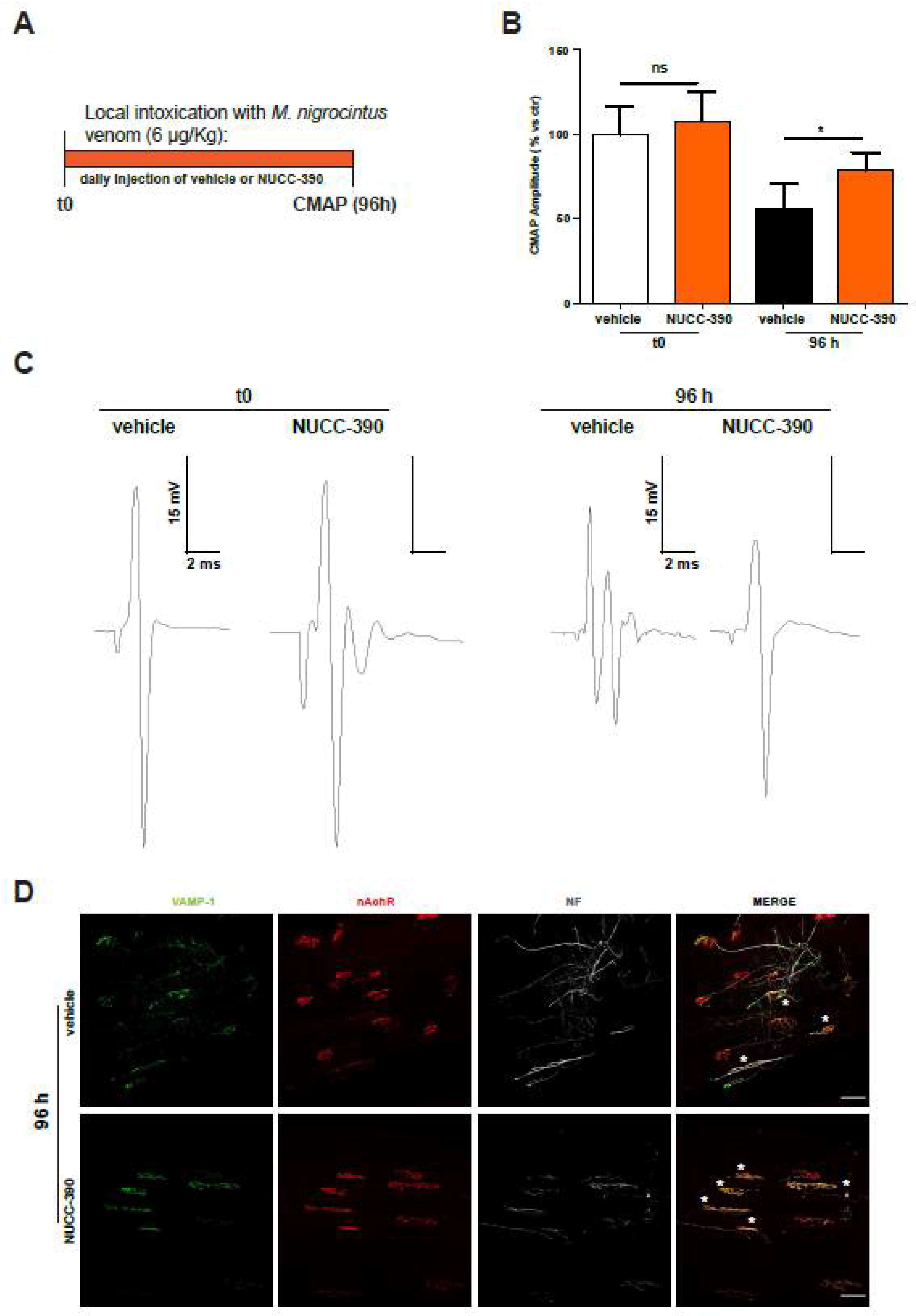
2.4. NUCC-390 Stimulates Restoration of Respiratory Function Impaired by M. nigrocinctus Venom
3. Discussion
4. Materials and Methods
4.1. Antibodies, Reagents, and Toxins
4.2. Evoked Junctional Potentials (EJPs)
4.3. Compound Muscle Action Potential (CMAP)
4.4. Ventilation (IVI) Recordings
4.5. Immunohistochemistry
4.6. Statistical Analysis
4.7. Ethical Statement
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Neglected Tropical Diseases. Available online: http://www.who.int/neglected_diseases/diseases/en/ (accessed on 21 June 2022).
- Williams, D.; Gutiérrez, J.M.; Harrison, R.; Warrell, D.A.; White, J.; Winkel, K.D.; Gopalakrishnakone, P. The Global Snake Bite Initiative: An antidote for snakebite. Lancet 2010, 375, 89–91. [Google Scholar] [CrossRef]
- Kasturiratne, A.; Wickremasinghe, A.R.; de Silva, N.; Gunawardena, N.K.; Pathmeswaran, A.; Premaratna, R.; Savioli, L.; Lalloo, D.G.; de Silva, H.J. The global burden of snakebite: A literature analysis and modelling based on regional estimates of envenoming and deaths. PLoS Med. 2008, 5, 1591–1604. [Google Scholar] [CrossRef] [Green Version]
- Harrison, R.A.; Hargreaves, A.; Wagstaff, S.C.; Faragher, B.; Lalloo, D.G. Snake envenoming: A disease of poverty. PLoS Negl. Trop. Dis. 2009, 3, e569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez, J.M.; Calvete, J.; Habib, A.G.; Harrison, R.A.; Williams, D.J.; Warrell, D.A. Snakebite envenoming. Nat. Rev. Dis. Primers 2017, 3, 17063. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Williams, D.; Fan, H.W.; Warrell, D.A. Snakebite envenoming from a global perspective: Towards an integrated approach. Toxicon 2010, 56, 1223–1235. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M. Current challenges for confronting the public health problem of snakebite envenoming in Central America. J. Venom Anim. Toxins Incl. Trop. Dis. 2014, 20, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, D.J.; Faiz, M.A.; Abela-Ridder, B.; Ainsworth, S.; Bulfone, T.C.; Nickerson, A.D.; Habib, A.G.; Junghanss, T.; Fan, H.W.; Turner, M.; et al. Strategy for a globally coordinated response to a priority neglected tropical disease: Snakebite envenoming. PLoS Negl. Trop. Dis. 2019, 13, e0007059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz, L.S.; Vargas, R.; Lopes, A.A. Snakebite envenomation and death in the developing world. Ethn. Dis. 2009, 19, 42. [Google Scholar]
- Seifert, S.A.; Armitage, J.O.; Sanchez, E.E. Snake Envenomation. N. Engl. J. Med. 2022, 386, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Warrell, D.A. Snake bite. Lancet 2010, 375, 77–88. [Google Scholar] [CrossRef]
- Tasoulis, T.; Isbister, G.K. A Review and Database of Snake Venom Proteomes. Toxins 2019, 9, e290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Silva, J.N., Jr.; Buononato, M.A.; Pires, M.G.; Feitosa, D.T. New World coral snake: An overview. In Advances in Coral Snake Biology: With an Emphasis in South America New World Coral Snakes: An Overview; Eagle Mountain Publishing: Eagle Mountain, UT, USA, 2021; pp. 115–139. [Google Scholar]
- Lomonte, B.; Rey-Suárez, P.; Fernández, J.; Sasa, M.; Pla, D.; Vargas, N.; Bénard-Valle, M.; Sanz, L.; Corrêa-Netto, C.; Núñez, V.; et al. Venoms of Micrurus coral snakes: Evolutionary trends in compositional patterns emerging from proteomic analyses. Toxicon 2016, 122, 7–25. [Google Scholar] [CrossRef]
- Bolaños, R. Las serpientes venenosas de Centroamérica y el problema del ofidismo. Primera parte. Aspectos zoológicos, epidemiológicos y biomédicos. Rev. Cost. Cienc. Méd. 1982, 3, 165–184. [Google Scholar]
- Gutiérrez, J.M. Snakebite envenomation in Central America. Epidemiology, Pathophysiology and treatment. In Handbook of Venoms and Toxins of Reptiles, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2021; pp. 543–558. [Google Scholar]
- Fernández, J.; Alape-Girón, A.; Angulo, Y.; Sanz, L.; Gutiérrez, J.; Calvete, J.; Lomonte, B. Venomic and antivenomic analyses of the Central American Coral Snake, Micrurus nigrocinctus (Elapidae). J. Proteom. Res. 2011, 10, 1816–1827. [Google Scholar] [CrossRef] [PubMed]
- Bucaretchi, F.; Capitani, E.M.; Vieira, R.J.; Rodrigues, C.K.; Zannin, M.; Da Silva, N.J.; Casais-e-Silva, L.L.; Hyslop, S. Coral snake bites (Micrurus spp.) in Brazil: A review of literature reports. Clin. Toxicol. 2016, 54, 222–234. [Google Scholar] [CrossRef]
- Negro, S.; Lessi, F.; Duregotti, E.; Aretini, P.; La Ferla, M.; Franceschi, S.; Menicagli, M.; Bergamin, E.; Radice, E.; Thelen, M.; et al. CXCL12α/SDF-1 from perisynaptic Schwann cells promotes regeneration of injured motor axon terminals. EMBO Mol. Med. 2017, 9, 1000–1010. [Google Scholar] [CrossRef] [PubMed]
- Negro, S.; Zanetti, G.; Mattarei, A.; Valentini, A.; Megighian, A.; Tombesi, G.; Zugno, A.; Dianin, V.; Pirazzini, M.; Fillo, S.; et al. An Agonist of the CXCR4 Receptor Strongly Promotes Regeneration of Degenerated Motor Axon Terminals. Cells 2019, 8, 1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stazi, M.; D’Este, G.; Mattarei, A.; Negro, S.; Lista, F.; Rigoni, M.; Megighian, A.; Montecucco, C. An agonist of the CXCR4 receptor accelerates the recovery from the peripheral neuroparalysis induced by Taipan snake envenomation. PLoS Negl. Trop. Dis. 2020, 14, e0008547. [Google Scholar] [CrossRef] [PubMed]
- Stazi, M.; Fabris, F.; Tan, K.Y.; Megighian, A.; Rubini, A.; Mattarei, A.; Negro, S.; D’Este, G.; Lista, F.; Rossetto, O.; et al. An agonist of the CXCR4 receptor is therapeutic for the neuroparalysis induced by Bungarus snakes envenoming. Clin. Transl. Med. 2022, 12, e651. [Google Scholar] [CrossRef]
- Rossetto, O.; Gorza, L.; Schiavo, G.; Schiavo, N.; Scheller, R.H.; Montecucco, C. VAMP/synaptobrevin isoforms 1 and 2 are widely and differentially expressed in nonneuronal tissues. J. Cell Biol. 1996, 32, 167–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Pena y Valenzuela, I.; Akaaboune, M. The Metabolic Stability of the Nicotinic Acetylcholine Receptor at the Neuromuscular Junction. Cells 2021, 10, 358. [Google Scholar] [CrossRef]
- Rey-Suárez, P.; Floriano, R.S.; Rostelato-Ferreira, S.; Saldarriaga-Córdoba, M.; Núñez, V.; Rodrigues-Simioni, L.; Lomonte, B. Mipartoxin-I, a novel three-finger toxin, is the major neurotoxic component in the venom of the redtail coral snake Micrurus mipartitus (Elapidae). Toxicon 2012, 60, 851–863. [Google Scholar] [CrossRef] [Green Version]
- Terra, A.L.; Moreira-Dill, L.S.; Simões-Silva, R.; Monteiro, J.R.; Cavalcante, W.L.; Gallacci, M.; Barros, N.B.; Nicolete, R.; Teles, C.B.; Medeiros, P.S.; et al. Biological characterization of the Amazon coral Micrurus spixii snake venom: Isolation of a new neurotoxic phospholipase A2. Toxicon 2015, 103, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Floriano, R.S.; Schezaro-Ramos, R.; Silva, N.J.; Bucaretchi, F.; Rowan, E.G.; Hyslop, S. Neurotoxicity of Micrurus lemniscatus lemniscatus (South American coralsnake) venom in vertebrate neuromuscular preparations in vitro and neutralization by antivenom. Arch. Toxicol. 2019, 93, 2065–2086. [Google Scholar] [CrossRef] [PubMed]
- Bénard-Valle, M.; Neri-Castro, E.; Yañez-Mendoza, M.F.; Lomonte, B.; Olvera, A.; Zamudio, F.; Restano-Cassulini, R.; Possani, L.D.; Jiménez-Ferrer, E.; Alagón, A. Functional, proteomic and transcriptomic characterization of the venom from Micrurus browni browni: Identification of the first lethal multimeric neurotoxin in coral snake venom. J. Proteom. 2020, 225, e103863. [Google Scholar] [CrossRef]
- Rigoni, M.; Caccin, P.; Gschmeissner, S.; Koster, G.; Postle, A.D.; Rossetto, O.; Schiavo, G.; Montecucco, C. Equivalent effects of snake PLA2 neurotoxins and lysophospholipid-fatty acid mixtures. Science 2005, 310, 1678–1680. [Google Scholar] [CrossRef] [PubMed]
- Rigoni, M.; Pizzo, P.; Schiavo, G.; Weston, A.E.; Zatti, G.; Caccin, P.; Rossetto, O.; Pozzan, T.; Montecucco, C. Calcium influx and mitochondrial alterations at synapses exposed to snake neurotoxin or their phospholipid hydrolysis products. J. Biol. Chem. 2007, 282, 11238–11245. [Google Scholar] [CrossRef] [Green Version]
- Rigoni, M.; Paoli, M.; Milanesi, E.; Caccin, P.; Rasola, A.; Bernardi, P.; Montecucco, C. Snake phospholipase A2 neurotoxins enter neurons, bind specifically to mitochondria and open their transition pores. J. Biol. Chem. 2008, 283, 34013–34020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paoli, M.; Rigoni, M.; Koster, G.; Rossetto, O.; Montecucco, C.; Postle, A.D. Mass Spectrometry Analysis of the Phospholipase A2 Activity of Snake Presynaptic Neurotoxins in Cultured Neurons. J. Neurochem. 2009, 111, 737–744. [Google Scholar] [CrossRef]
- Tedesco, E.; Rigoni, M.; Caccin, P.; Grishin, E.; Rossetto, O.; Montecucco, C. Calcium overload in nerve terminals of cultured neurons intoxicated by alpha-latrotoxin and snake PLA2 neurotoxins. Toxicon 2009, 54, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Mishra, R.K.; Shum, A.K.; Platanias, L.C.; Miller, R.J.; Schiltz, G.E. Discovery and characterization of novel small-molecule CXCR4 receptor agonists and antagonists. Sci. Rep. 2016, 6, 30155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trevett, A.J.; Lalloo, D.G.; Nwokolo, N.C.; Naraqi, S.; Kevau, I.H.; Theakston, R.D.; Warrell, D.A. Electrophysiological findings in patients envenomed following the bite of a Papuan taipan (Oxyuranus scutellatus canni). Trans. R. Soc. Trop. Med. Hyg. 1995, 89, 415–417. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stazi, M.; Fabris, F.; Fernández, J.; D'Este, G.; Rigoni, M.; Megighian, A.; Gutiérrez, J.M.; Lomonte, B.; Montecucco, C. Recovery from the Neuroparalysis Caused by the Micrurus nigrocinctus Venom Is Accelerated by an Agonist of the CXCR4 Receptor. Toxins 2022, 14, 531. https://doi.org/10.3390/toxins14080531
Stazi M, Fabris F, Fernández J, D'Este G, Rigoni M, Megighian A, Gutiérrez JM, Lomonte B, Montecucco C. Recovery from the Neuroparalysis Caused by the Micrurus nigrocinctus Venom Is Accelerated by an Agonist of the CXCR4 Receptor. Toxins. 2022; 14(8):531. https://doi.org/10.3390/toxins14080531
Chicago/Turabian StyleStazi, Marco, Federico Fabris, Julián Fernández, Giorgia D'Este, Michela Rigoni, Aram Megighian, José María Gutiérrez, Bruno Lomonte, and Cesare Montecucco. 2022. "Recovery from the Neuroparalysis Caused by the Micrurus nigrocinctus Venom Is Accelerated by an Agonist of the CXCR4 Receptor" Toxins 14, no. 8: 531. https://doi.org/10.3390/toxins14080531
APA StyleStazi, M., Fabris, F., Fernández, J., D'Este, G., Rigoni, M., Megighian, A., Gutiérrez, J. M., Lomonte, B., & Montecucco, C. (2022). Recovery from the Neuroparalysis Caused by the Micrurus nigrocinctus Venom Is Accelerated by an Agonist of the CXCR4 Receptor. Toxins, 14(8), 531. https://doi.org/10.3390/toxins14080531