Gut–Kidney Axis Investigations in Animal Models of Chronic Kidney Disease

 , , , ,
, , , ,
Abstract
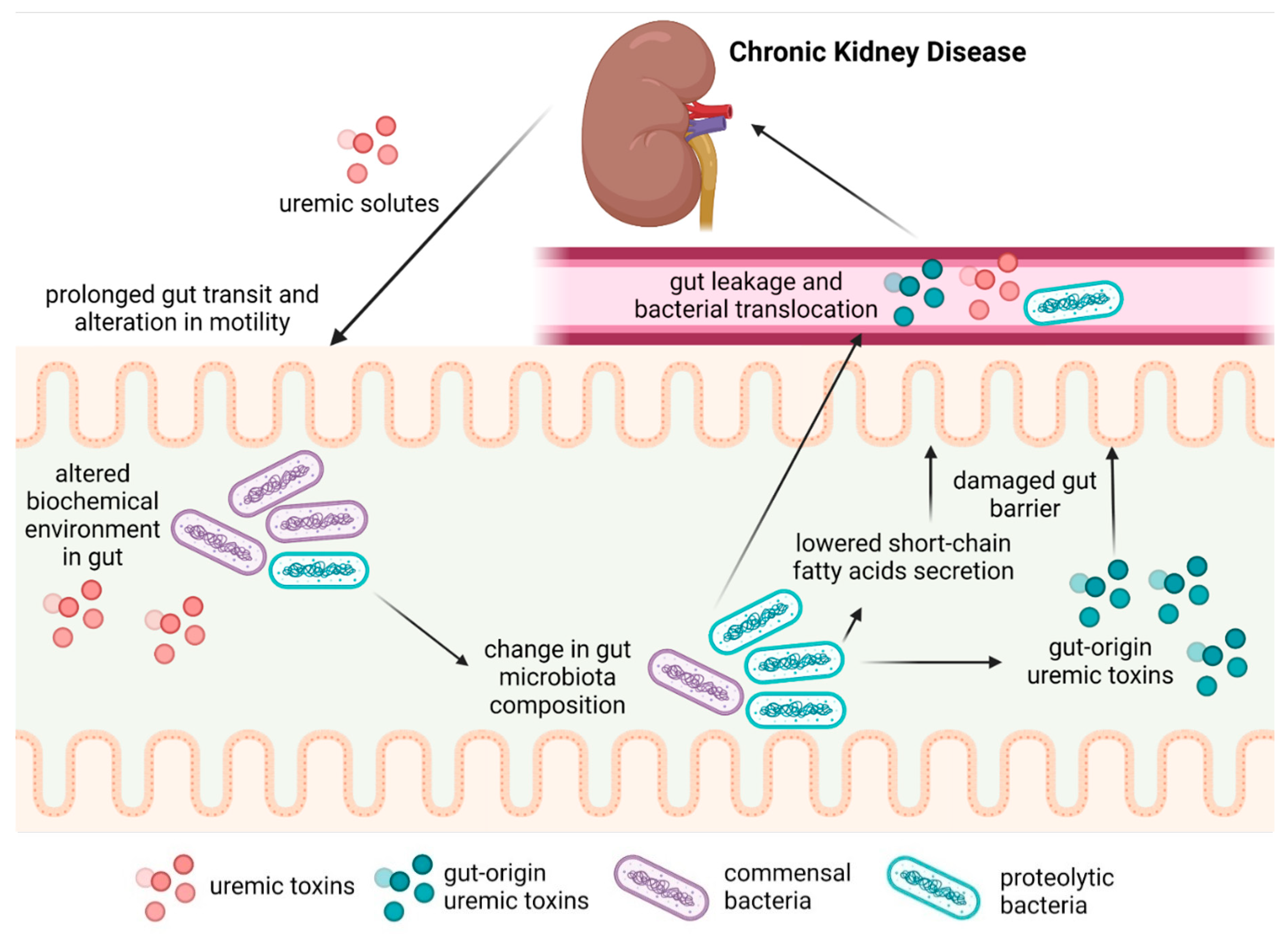
:1. Introduction
2. Materials and Methods
3. CKD Models Used to Assess the Gut–Kidney Axis
4. Motility Dysfunction and Gut Barrier
5. Gut Microbiota Alterations in CKD
6. Adaptation of CKD Animal Models to GM–Kidney Research
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Boer, I.H.; Caramori, M.L.; Chan, J.C.N.; Heerspink, H.J.L.; Hurst, C.; Khunti, K.; Liew, A.; Michos, E.D.; Navaneethan, S.D.; Olowu, W.A.; et al. KDIGO 2020 Clinical Practice Guideline for Diabetes Management in Chronic Kidney Disease. Kidney Int. 2020, 98, S1–S115. [Google Scholar] [CrossRef] [PubMed]
- Alméras, C.; Argilés, A. The General Picture of Uremia. Semin Dial. 2009, 22, 329–333. [Google Scholar] [CrossRef]
- Duranton, F.; Cohen, G.; De Smet, R.; Rodriguez, M.; Jankowski, J.; Vanholder, R.; Argiles, A.; European Uremic Toxin Work Group. Normal and Pathologic Concentrations of Uremic Toxins. J. Am. Soc. Nephrol. 2012, 23, 1258–1270. [Google Scholar] [CrossRef] [PubMed]
- Cosola, C.; Rocchetti, M.T.; di Bari, I.; Acquaviva, P.M.; Maranzano, V.; Corciulo, S.; Di Ciaula, A.; Di Palo, D.M.; La Forgia, F.M.; Fontana, S.; et al. An Innovative Synbiotic Formulation Decreases Free Serum Indoxyl Sulfate, Small Intestine Permeability and Ameliorates Gastrointestinal Symptoms in a Randomized Pilot Trial in Stage IIIb-IV CKD Patients. Toxins 2021, 13, 334. [Google Scholar] [CrossRef]
- Strid, H.; Simrén, M.; Stotzer, P.-O.; Ringström, G.; Abrahamsson, H.; Björnsson, E.S. Patients with Chronic Renal Failure Have Abnormal Small Intestinal Motility and a High Prevalence of Small Intestinal Bacterial Overgrowth. Digestion 2003, 67, 129–137. [Google Scholar] [CrossRef]
- Wang, F.; Jiang, H.; Shi, K.; Ren, Y.; Zhang, P.; Cheng, S. Gut Bacterial Translocation Is Associated with Microinflammation in End-Stage Renal Disease Patients: Bacterial Translocation in ESRD. Nephrology 2012, 17, 733–738. [Google Scholar] [CrossRef]
- Chiavaroli, L.; Mirrahimi, A.; Sievenpiper, J.L.; Jenkins, D.J.A.; Darling, P.B. Dietary Fiber Effects in Chronic Kidney Disease: A Systematic Review and Meta-Analysis of Controlled Feeding Trials. Eur. J. Clin. Nutr. 2015, 69, 761–768. [Google Scholar] [CrossRef]
- Hsu, C.-K.; Su, S.-C.; Chang, L.-C.; Shao, S.-C.; Yang, K.-J.; Chen, C.-Y.; Chen, Y.-T.; Wu, I.-W. Effects of Low Protein Diet on Modulating Gut Microbiota in Patients with Chronic Kidney Disease: A Systematic Review and Meta-Analysis of International Studies. Int. J. Med. Sci. 2021, 18, 3839–3850. [Google Scholar] [CrossRef]
- Kortman, G.A.M.; Reijnders, D.; Swinkels, D.W. Oral Iron Supplementation: Potential Implications for the Gut Microbiome and Metabolome in Patients with CKD. Hemodial. Int. 2017, 21, S28–S36. [Google Scholar] [CrossRef]
- Yoshifuji, A.; Wakino, S.; Irie, J.; Matsui, A.; Hasegawa, K.; Tokuyama, H.; Hayashi, K.; Itoh, H. Oral Adsorbent AST-120 Ameliorates Gut Environment and Protects against the Progression of Renal Impairment in CKD Rats. Clin. Exp. Nephrol. 2018, 22, 1069–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, D.; Zhao, W.; Lin, Z.; Wu, J.; Lin, H.; Li, Y.; Song, J.; Zhang, J.; Peng, H. The Effects of Hemodialysis and Peritoneal Dialysis on the Gut Microbiota of End-Stage Renal Disease Patients, and the Relationship Between Gut Microbiota and Patient Prognoses. Front. Cell. Infect. Microbiol. 2021, 11, 579386. [Google Scholar] [CrossRef]
- Fouque, D.; Kalantar-Zadeh, K.; Kopple, J.; Cano, N.; Chauveau, P.; Cuppari, L.; Franch, H.; Guarnieri, G.; Ikizler, T.A.; Kaysen, G.; et al. A Proposed Nomenclature and Diagnostic Criteria for Protein—Energy Wasting in Acute and Chronic Kidney Disease. Kidney Int. 2008, 73, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.; Piceno, Y.M.; DeSantis, T.Z.; Pahl, M.; Andersen, G.L.; Vaziri, N.D. Expansion of Urease- and Uricase-Containing, Indole- and p-Cresol-Forming and Contraction of Short Chain Fatty Acid-Producing Intestinal Microbiota in ESRD. Am. J. Nephrol. 2014, 39, 230–237. [Google Scholar] [CrossRef]
- He, J.; Zhang, P.; Shen, L.; Niu, L.; Tan, Y.; Chen, L.; Zhao, Y.; Bai, L.; Hao, X.; Li, X.; et al. Short-Chain Fatty Acids and Their Association with Signalling Pathways in Inflammation, Glucose and Lipid Metabolism. Int. J. Mol. Sci. 2020, 21, 6356. [Google Scholar] [CrossRef]
- Huang, Y.; Xin, W.; Xiong, J.; Yao, M.; Zhang, B.; Zhao, J. The Intestinal Microbiota and Metabolites in the Gut-Kidney-Heart Axis of Chronic Kidney Disease. Front. Pharmacol. 2022, 13, 837500. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, H.P.; Ferré, J.P.; Watson, A.D.; Brown, C.A.; Serthelon, J.P.; Laroute, V.; Concordet, D.; Toutain, P.L. Small Bowel Motility and Colonic Transit Are Altered in Dogs with Moderate Renal Failure. Am. J. Physiol Regul Integr Comp. Physiol 2001, 281, R230–R238. [Google Scholar] [CrossRef]
- Da Graça, J.R.V.; Parente, C.C.; Fiúza, R.F.; Da Silva, P.A.F.; Mota, B.T.; Salles, L.D.; Silva, C.M.D.S.; da Silva, M.T.B.; Oliveira, R.; Dos Santos, A.A. Subtotal Nephrectomy Inhibits the Gastric Emptying of Liquid in Awake Rats. Physiol. Rep. 2015, 3, e12291. [Google Scholar] [CrossRef]
- Wang, S.C.; Lu, K.Y.; Chen, S.M.; Young, T.K. Gastric Emptying and Intestinal Transit of Liquid and Solid Markers in Rats with Chronic Uremia. Chin. J. Physiol. 2001, 44, 81–87. [Google Scholar] [PubMed]
- Yu, C.; Tan, S.; Wang, Z.; Deng, B.; Li, J.; Wang, Q.; Zhou, C.; Kang, X.; Yu, Z.; Zhuang, S. Chronic Kidney Disease Elicits an Intestinal Inflammation Resulting in Intestinal Dysmotility Associated with the Activation of Inducible Nitric Oxide Synthesis in Rat. Digestion 2018, 97, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Hoibian, E.; Florens, N.; Koppe, L.; Vidal, H.; Soulage, C.O. Distal Colon Motor Dysfunction in Mice with Chronic Kidney Disease: Putative Role of Uremic Toxins. Toxins 2018, 10, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, A.; Krieg, R.; Massey, H.D.; Carl, D.; Ghosh, S.; Gehr, T.W.B.; Ghosh, S.S. Sodium Butyrate Ameliorates Insulin Resistance and Renal Failure in CKD Rats by Modulating Intestinal Permeability and Mucin Expression. Nephrol. Dial. Transplant. 2019, 34, 783–794. [Google Scholar] [CrossRef]
- Huang, Y.; Zhou, J.; Wang, S.; Xiong, J.; Chen, Y.; Liu, Y.; Xiao, T.; Li, Y.; He, T.; Li, Y.; et al. Indoxyl Sulfate Induces Intestinal Barrier Injury through IRF1-DRP1 Axis-Mediated Mitophagy Impairment. Theranostics 2020, 10, 7384–7400. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Lim, S.Y.; Ko, Y.S.; Lee, H.Y.; Oh, S.W.; Kim, M.G.; Cho, W.Y.; Jo, S.K. Intestinal Barrier Disruption and Dysregulated Mucosal Immunity Contribute to Kidney Fibrosis in Chronic Kidney Disease. Nephrol. Dial. Transplant. 2019, 34, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Vaziri, N.D.; Yuan, J.; Khazaeli, M.; Masuda, Y.; Ichii, H.; Liu, S. Oral Activated Charcoal Adsorbent (AST-120) Ameliorates Chronic Kidney Disease-Induced Intestinal Epithelial Barrier Disruption. Am. J. Nephrol. 2013, 37, 518–525. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, P.; Jiang, H.; Cheng, S. Gut Bacterial Translocation Contributes to Microinflammation in Experimental Uremia. Dig. Dis. Sci. 2012, 57, 2856–2862. [Google Scholar] [CrossRef] [PubMed]
- De Almeida Duarte, J.B.; de Aguilar-Nascimento, J.E.; Nascimento, M.; Nochi, R.J. Bacterial Translocation in Experimental Uremia. Urol. Res. 2004, 32, 266–270. [Google Scholar] [CrossRef]
- Vaziri, N.D.; Yuan, J.; Nazertehrani, S.; Ni, Z.; Liu, S. Chronic Kidney Disease Causes Disruption of Gastric and Small Intestinal Epithelial Tight Junction. Am. J. Nephrol. 2013, 38, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Yoshifuji, A.; Wakino, S.; Irie, J.; Tajima, T.; Hasegawa, K.; Kanda, T.; Tokuyama, H.; Hayashi, K.; Itoh, H. Gut Lactobacillus Protects against the Progression of Renal Damage by Modulating the Gut Environment in Rats. Nephrol. Dial. Transplant. 2016, 31, 401–412. [Google Scholar] [CrossRef]
- Vaziri, N.D.; Yuan, J.; Rahimi, A.; Ni, Z.; Said, H.; Subramanian, V.S. Disintegration of Colonic Epithelial Tight Junction in Uremia: A Likely Cause of CKD-Associated Inflammation. Nephrol. Dial. Transplant. 2012, 27, 2686–2693. [Google Scholar] [CrossRef]
- Vaziri, N.D.; Nunes, A.C.F.; Said, H.; Khazaeli, M.; Liu, H.; Zhao, Y.; Jing, W.; Cogburn, K.; Alikhani, L.; Lau, W.L. Route of Intestinal Absorption and Tissue Distribution of Iron Contained in the Novel Phosphate Binder Ferric Citrate. Nephrol. Dial. Transplant. 2020, 35, 1136–1144. [Google Scholar] [CrossRef] [PubMed]
- Lau, W.L.; Liu, S.-M.; Pahlevan, S.; Yuan, J.; Khazaeli, M.; Ni, Z.; Chan, J.Y.; Vaziri, N.D. Role of Nrf2 Dysfunction in Uremia-Associated Intestinal Inflammation and Epithelial Barrier Disruption. Dig. Dis. Sci. 2015, 60, 1215–1222. [Google Scholar] [CrossRef]
- Vaziri, N.D.; Liu, S.-M.; Lau, W.L.; Khazaeli, M.; Nazertehrani, S.; Farzaneh, S.H.; Kieffer, D.A.; Adams, S.H.; Martin, R.J. High Amylose Resistant Starch Diet Ameliorates Oxidative Stress, Inflammation, and Progression of Chronic Kidney Disease. PLoS ONE 2014, 9, e114881. [Google Scholar] [CrossRef] [PubMed]
- Hung, T.V.; Suzuki, T. Dietary Fermentable Fibers Attenuate Chronic Kidney Disease in Mice by Protecting the Intestinal Barrier. J. Nutr. 2018, 148, 552–561. [Google Scholar] [CrossRef]
- Chen, L.; Chen, D.-Q.; Liu, J.-R.; Zhang, J.; Vaziri, N.D.; Zhuang, S.; Chen, H.; Feng, Y.-L.; Guo, Y.; Zhao, Y.-Y. Unilateral Ureteral Obstruction Causes Gut Microbial Dysbiosis and Metabolome Disorders Contributing to Tubulointerstitial Fibrosis. Exp. Mol. Med. 2019, 51, 1–18. [Google Scholar] [CrossRef]
- Hsu, C.-N.; Yang, H.-W.; Hou, C.-Y.; Chang-Chien, G.-P.; Lin, S.; Tain, Y.-L. Maternal Adenine-Induced Chronic Kidney Disease Programs Hypertension in Adult Male Rat Offspring: Implications of Nitric Oxide and Gut Microbiome Derived Metabolites. Int. J. Mol. Sci. 2020, 21, 7237. [Google Scholar] [CrossRef]
- Yano, H.; Tamura, Y.; Kobayashi, K.; Tanemoto, M.; Uchida, S. Uric Acid Transporter ABCG2 Is Increased in the Intestine of the 5/6 Nephrectomy Rat Model of Chronic Kidney Disease. Clin. Exp. Nephrol. 2014, 18, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Ji, C.; Deng, Y.; Yang, A.; Lu, Z.; Chen, Y.; Liu, X.; Han, L.; Zou, C. Rhubarb Enema Improved Colon Mucosal Barrier Injury in 5/6 Nephrectomy Rats May Associate With Gut Microbiota Modification. Front. Pharmacol. 2020, 11, 1092. [Google Scholar] [CrossRef] [PubMed]
- Mishima, E.; Fukuda, S.; Shima, H.; Hirayama, A.; Akiyama, Y.; Takeuchi, Y.; Fukuda, N.N.; Suzuki, T.; Suzuki, C.; Yuri, A.; et al. Alteration of the Intestinal Environment by Lubiprostone Is Associated with Amelioration of Adenine-Induced CKD. J. Am. Soc. Nephrol. 2015, 26, 1787–1794. [Google Scholar] [CrossRef]
- Coe, F.L. Uric Acid and Calcium Oxalate Nephrolithiasis. Kidney Int. 1983, 24, 392–403. [Google Scholar] [CrossRef]
- Koeda, T.; Wakaki, K.; Koizumi, F.; Yokozawa, T.; Oura, H. Early Changes of Proximal Tubules in the Kidney of Adenine-Ingesting Rats, with Special Reference to Biochemical and Electron Microscopic Studies. Nihon Jinzo Gakkai Shi. 1988, 30, 239–246. [Google Scholar] [PubMed]
- Yokozawa, T.; Zheng, P.D.; Oura, H.; Koizumi, F. Animal Model of Adenine-Induced Chronic Renal Failure in Rats. Nephron 1986, 44, 230–234. [Google Scholar] [CrossRef]
- Brenner, B.M.; Lawler, E.V.; Mackenzie, H.S. The Hyperfiltration Theory: A Paradigm Shift in Nephrology. Kidney Int. 1996, 49, 1774–1777. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Klimova, E.; Aparicio-Trejo, O.E.; Tapia, E.; Pedraza-Chaverri, J. Unilateral Ureteral Obstruction as a Model to Investigate Fibrosis-Attenuating Treatments. Biomolecules 2019, 9, 141. [Google Scholar] [CrossRef]
- Ucero, A.C.; Benito-Martin, A.; Izquierdo, M.C.; Sanchez-Niño, M.D.; Sanz, A.B.; Ramos, A.M.; Berzal, S.; Ruiz-Ortega, M.; Egido, J.; Ortiz, A. Unilateral Ureteral Obstruction: Beyond Obstruction. Int. Urol. Nephrol. 2014, 46, 765–776. [Google Scholar] [CrossRef]
- Meijers, B.; Farré, R.; Dejongh, S.; Vicario, M.; Evenepoel, P. Intestinal Barrier Function in Chronic Kidney Disease. Toxins 2018, 10, 298. [Google Scholar] [CrossRef]
- Wu, M.-J.; Chang, C.-S.; Cheng, C.-H.; Chen, C.-H.; Lee, W.-C.; Hsu, Y.-H.; Shu, K.-H.; Tang, M.-J. Colonic Transit Time in Long-Term Dialysis Patients. Am. J. Kidney Dis. 2004, 44, 322–327. [Google Scholar] [CrossRef] [PubMed]
- Vanuytsel, T.; Tack, J.; Farre, R. The Role of Intestinal Permeability in Gastrointestinal Disorders and Current Methods of Evaluation. Front. Nutr. 2021, 8, 717925. [Google Scholar] [CrossRef]
- Lemesch, S.; Ribitsch, W.; Schilcher, G.; Spindelböck, W.; Hafner-Gießauf, H.; Marsche, G.; Pasterk, L.; Payerl, D.; Schmerböck, B.; Tawdrous, M.; et al. Mode of Renal Replacement Therapy Determines Endotoxemia and Neutrophil Dysfunction in Chronic Kidney Disease. Sci. Rep. 2016, 6, 34534. [Google Scholar] [CrossRef]
- Munford, R.S. Endotoxemia—Menace, Marker, or Mistake? J. Leukoc. Biol. 2016, 100, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Berg, R.D.; Garlington, A.W. Translocation of Certain Indigenous Bacteria from the Gastrointestinal Tract to the Mesenteric Lymph Nodes and Other Organs in a Gnotobiotic Mouse Model. Infect. Immun. 1979, 23, 403–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bossola, M.; Sanguinetti, M.; Scribano, D.; Zuppi, C.; Giungi, S.; Luciani, G.; Torelli, R.; Posteraro, B.; Fadda, G.; Tazza, L. Circulating Bacterial-Derived DNA Fragments and Markers of Inflammation in Chronic Hemodialysis Patients. Clin. J. Am. Soc. Nephrol. 2009, 4, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.-K.; Lim, P.-S.; Jin, J.-S.; Wu, M.-Y.; Chen, C.-H. Impaired Gut Epithelial Tight Junction Expression in Hemodialysis Patients Complicated with Intradialytic Hypotension. Biomed. Res. Int. 2018, 2018, 2670312. [Google Scholar] [CrossRef]
- Greenbaum, D.; Colangelo, C.; Williams, K.; Gerstein, M. Comparing Protein Abundance and MRNA Expression Levels on a Genomic Scale. Genome Biol. 2003, 4, 117. [Google Scholar] [CrossRef]
- Wang, S.; Lv, D.; Jiang, S.; Jiang, J.; Liang, M.; Hou, F.; Chen, Y. Quantitative Reduction in Short-Chain Fatty Acids, Especially Butyrate, Contributes to the Progression of Chronic Kidney Disease. Clin. Sci. 2019, 133, 1857–1870. [Google Scholar] [CrossRef]
- Peterson, L.W.; Artis, D. Intestinal Epithelial Cells: Regulators of Barrier Function and Immune Homeostasis. Nat. Rev. Immunol. 2014, 14, 141–153. [Google Scholar] [CrossRef]
- Nigam, S.K.; Bush, K.T. Uraemic Syndrome of Chronic Kidney Disease: Altered Remote Sensing and Signalling. Nat. Rev. Nephrol. 2019, 15, 301–316. [Google Scholar] [CrossRef]
- Matsuo, H.; Tsunoda, T.; Ooyama, K.; Sakiyama, M.; Sogo, T.; Takada, T.; Nakashima, A.; Nakayama, A.; Kawaguchi, M.; Higashino, T.; et al. Hyperuricemia in Acute Gastroenteritis Is Caused by Decreased Urate Excretion via ABCG2. Sci. Rep. 2016, 6, 31003. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Ning, X.; Liu, B.; Dong, R.; Bai, M.; Sun, S. Specific Alterations in Gut Microbiota in Patients with Chronic Kidney Disease: An Updated Systematic Review. Ren. Fail. 2021, 43, 102–112. [Google Scholar] [CrossRef]
- Van der Worp, H.B.; Howells, D.W.; Sena, E.S.; Porritt, M.J.; Rewell, S.; O’Collins, V.; Macleod, M.R. Can Animal Models of Disease Reliably Inform Human Studies? PLoS Med. 2010, 7, e1000245. [Google Scholar] [CrossRef] [PubMed]
- Belzung, C.; Lemoine, M. Criteria of Validity for Animal Models of Psychiatric Disorders: Focus on Anxiety Disorders and Depression. Biol. Mood Anxiety Disord. 2011, 1, 9. [Google Scholar] [CrossRef] [Green Version]
- Bebarta, V.; Luyten, D.; Heard, K. Emergency Medicine Animal Research: Does Use of Randomization and Blinding Affect the Results? Acad. Emerg. Med. 2003, 10, 684–687. [Google Scholar] [CrossRef]
- Becker, G.J.; Hewitson, T.D. Animal Models of Chronic Kidney Disease: Useful but Not Perfect. Nephrol. Dial. Transplant. 2013, 28, 2432–2438. [Google Scholar] [CrossRef] [PubMed]
- Treuting, P.M.; Valasek, M.A.; Dintzis, S.M. 11—Upper gastrointestinal tract. In Comparative Anatomy and Histology; Treuting, P.M., Dintzis, S.M., Eds.; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2017; pp. 155–175. ISBN 978-0-12-381361-9. [Google Scholar]
- Kararli, T.T. Comparison of the Gastrointestinal Anatomy, Physiology, and Biochemistry of Humans and Commonly Used Laboratory Animals. Biopharm. Drug Dispos. 1995, 16, 351–380. [Google Scholar] [CrossRef]
- Horn, C.C.; Kimball, B.A.; Wang, H.; Kaus, J.; Dienel, S.; Nagy, A.; Gathright, G.R.; Yates, B.J.; Andrews, P.L.R. Why Can’t Rodents Vomit? A Comparative Behavioral, Anatomical, and Physiological Study. PLoS ONE 2013, 8, e60537. [Google Scholar] [CrossRef]
- Bo, T.-B.; Zhang, X.-Y.; Kohl, K.D.; Wen, J.; Tian, S.-J.; Wang, D.-H. Coprophagy Prevention Alters Microbiome, Metabolism, Neurochemistry, and Cognitive Behavior in a Small Mammal. ISME J. 2020, 14, 2625–2645. [Google Scholar] [CrossRef]
- Jarett, J.K.; Kingsbury, D.D.; Dahlhausen, K.E.; Ganz, H.H. Best Practices for Microbiome Study Design in Companion Animal Research. Front. Vet. Sci. 2021, 8, 644836. [Google Scholar] [CrossRef] [PubMed]
- Al-Asmakh, M.; Sohail, M.U.; Al-Jamal, O.; Shoair, B.M.; Al-Baniali, A.Y.; Bouabidi, S.; Nasr, S.; Bawadi, H. The Effects of Gum Acacia on the Composition of the Gut Microbiome and Plasma Levels of Short-Chain Fatty Acids in a Rat Model of Chronic Kidney Disease. Front. Pharmacol. 2020, 11, 569402. [Google Scholar] [CrossRef]
- Sueyoshi, M.; Fukunaga, M.; Mei, M.; Nakajima, A.; Tanaka, G.; Murase, T.; Narita, Y.; Hirata, S.; Kadowaki, D. Effects of Lactulose on Renal Function and Gut Microbiota in Adenine-Induced Chronic Kidney Disease Rats. Clin. Exp. Nephrol. 2019, 23, 908–919. [Google Scholar] [CrossRef] [PubMed]
- Jia, T.; Olauson, H.; Lindberg, K.; Amin, R.; Edvardsson, K.; Lindholm, B.; Andersson, G.; Wernerson, A.; Sabbagh, Y.; Schiavi, S.; et al. A Novel Model of Adenine-Induced Tubulointerstitial Nephropathy in Mice. BMC Nephrol. 2013, 14, 116. [Google Scholar] [CrossRef]
- Tirelle, P.; Breton, J.; Riou, G.; Déchelotte, P.; Coëffier, M.; Ribet, D. Comparison of Different Modes of Antibiotic Delivery on Gut Microbiota Depletion Efficiency and Body Composition in Mouse. BMC Microbiol. 2020, 20, 340. [Google Scholar] [CrossRef]
- Serbanescu, M.A.; Mathena, R.P.; Xu, J.; Santiago-Rodriguez, T.; Hartsell, T.L.; Cano, R.J.; Mintz, C.D. General Anesthesia Alters the Diversity and Composition of the Intestinal Microbiota in Mice. Anesth. Analg. 2019, 129, e126–e129. [Google Scholar] [CrossRef]
- Wang, F.; Meng, J.; Zhang, L.; Johnson, T.; Chen, C.; Roy, S. Morphine Induces Changes in the Gut Microbiome and Metabolome in a Morphine Dependence Model. Sci. Rep. 2018, 8, 3596. [Google Scholar] [CrossRef]
- Vaziri, N.D.; Yuan, J.; Norris, K. Role of Urea in Intestinal Barrier Dysfunction and Disruption of Epithelial Tight Junction in CKD. Am. J. Nephrol. 2013, 37, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.; Fan, R.; Zhao, L.; Tong, C.; Qian, X.; Zhang, M.; Xiao, R.; Ma, W. Trans-Fatty Acids Alter the Gut Microbiota in High-Fat-Diet-Induced Obese Rats. Br. J. Nutr. 2020, 124, 1251–1263. [Google Scholar] [CrossRef] [PubMed]
- Olson, C.A.; Vuong, H.E.; Yano, J.M.; Liang, Q.Y.; Nusbaum, D.J.; Hsiao, E.Y. The Gut Microbiota Mediates the Anti-Seizure Effects of the Ketogenic Diet. Cell 2018, 173, 1728–1741.e13. [Google Scholar] [CrossRef]
- Dias, M.F.; Reis, M.P.; Acurcio, L.B.; Carmo, A.O.; Diamantino, C.F.; Motta, A.M.; Kalapothakis, E.; Nicoli, J.R.; Nascimento, A.M.A. Changes in Mouse Gut Bacterial Community in Response to Different Types of Drinking Water. Water Res. 2018, 132, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Whipple, B.; Agar, J.; Zhao, J.; Pearce, D.A.; Kovács, A.D. The Acidified Drinking Water-Induced Changes in the Behavior and Gut Microbiota of Wild-Type Mice Depend on the Acidification Mode. Sci. Rep. 2021, 11, 2877. [Google Scholar] [CrossRef]
- Zhou, K.; Liu, W.; Chen, Z.; Yang, D.; Qiu, Z.; Feng, H.; Li, C.; Jin, M.; Li, J.; Xu, Q.; et al. The Effect of Different Drinking Water in Culture Medium on Feces Microbiota Diversity. J. Water Health 2020, 19, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Ambery, A.G.; Tackett, L.; Penque, B.A.; Hickman, D.L.; Elmendorf, J.S. Effect of Corncob Bedding on Feed Conversion Efficiency in a High-Fat Diet-Induced Prediabetic Model in C57Bl/6J Mice. J. Am. Assoc. Lab. Anim. Sci. 2014, 53, 449–451. [Google Scholar] [PubMed]
- Gregor, A.; Fragner, L.; Trajanoski, S.; Li, W.; Sun, X.; Weckwerth, W.; König, J.; Duszka, K. Cage Bedding Modifies Metabolic and Gut Microbiota Profiles in Mouse Studies Applying Dietary Restriction. Sci. Rep. 2020, 10, 20835. [Google Scholar] [CrossRef] [PubMed]
- Bidot, W.A.; Ericsson, A.C.; Franklin, C.L. Effects of Water Decontamination Methods and Bedding Material on the Gut Microbiota. PLoS ONE 2018, 13, e0198305. [Google Scholar] [CrossRef]
- Carter, R.L.; Lipman, N.S. Feed and bedding. In Management of Animal Care and Use Programs in Research, Education, and Testing; Weichbrod, R.H., Thompson, G.A. (Heidbrink), Norton, J.N., Eds.; CRC Press: Boca Raton, FL, USA; Taylor & Francis: Abingdon, UK, 2018; ISBN 978-1-4987-4844-5. [Google Scholar]
- Chen, S.; Zheng, Y.; Zhou, Y.; Guo, W.; Tang, Q.; Rong, G.; Hu, W.; Tang, J.; Luo, H. Gut Dysbiosis with Minimal Enteritis Induced by High Temperature and Humidity. Sci. Rep. 2019, 9, 18686. [Google Scholar] [CrossRef]
- Kim, Y.-M.; Snijders, A.M.; Brislawn, C.J.; Stratton, K.G.; Zink, E.M.; Fansler, S.J.; Metz, T.O.; Mao, J.-H.; Jansson, J.K. Light-Stress Influences the Composition of the Murine Gut Microbiome, Memory Function, and Plasma Metabolome. Front. Mol. Biosci. 2019, 6, 108. [Google Scholar] [CrossRef] [PubMed]
- David, J.M.; Knowles, S.; Lamkin, D.M.; Stout, D.B. Individually Ventilated Cages Impose Cold Stress on Laboratory Mice: A Source of Systemic Experimental Variability. J. Am. Assoc. Lab. Anim. Sci. 2013, 52, 738–744. [Google Scholar] [PubMed]
- Deloris Alexander, A.; Orcutt, R.P.; Henry, J.C.; Baker, J.; Bissahoyo, A.C.; Threadgill, D.W. Quantitative PCR Assays for Mouse Enteric Flora Reveal Strain-Dependent Differences in Composition That Are Influenced by the Microenvironment. Mamm. Genome 2006, 17, 1093–1104. [Google Scholar] [CrossRef] [PubMed]
- Beura, L.K.; Hamilton, S.E.; Bi, K.; Schenkel, J.M.; Odumade, O.A.; Casey, K.A.; Thompson, E.A.; Fraser, K.A.; Rosato, P.C.; Filali-Mouhim, A.; et al. Normalizing the Environment Recapitulates Adult Human Immune Traits in Laboratory Mice. Nature 2016, 532, 512–516. [Google Scholar] [CrossRef] [PubMed]
- Foster, K.R.; Schluter, J.; Coyte, K.Z.; Rakoff-Nahoum, S. The Evolution of the Host Microbiome as an Ecosystem on a Leash. Nature 2017, 548, 43–51. [Google Scholar] [CrossRef]
- Rosenberg, E.; Zilber-Rosenberg, I. The Hologenome Concept of Evolution after 10 Years. Microbiome 2018, 6, 78. [Google Scholar] [CrossRef]
- Van de Guchte, M.; Blottière, H.M.; Doré, J. Humans as Holobionts: Implications for Prevention and Therapy. Microbiome 2018, 6, 81. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Study | Species | CKD Induction | Study Strengths | Study Weaknesses | Main Results |
|---|---|---|---|---|---|
| Lefebvre, (2001) [16] | Beagle male dogs | SNx | colony animals, confirmed CKD, individual cages | no randomization, carnivorous, no treated control | intestinal transit time↑ |
| da Graça, (2015) [17] | Wistar male rats | SNx | randomization, omnivorous colony animals | no diet description | small intestinal transit↓ |
| Wang, (2001) [18] | Sprague– Dawley male rats | SNx | omnivorous colony animals, confirmed CKD, individual cages | no randomization, no-sham control, tap water, no diet description | small intestinal transit↓ (no change when fasting) |
| Yu, (2018) [19] | Sprague– Dawley male rats | SNx | randomization, omnivorous colony animals | no diet description | small intestinal transit↓ inflammation, oxidative stress↑ |
| Hoibian, (2018) [20] | C57Bl/6 JRj male mice | adenine diet | randomization, omnivorous colony animals | diet-induced CKD | total intestinal transit↓ colon motility↓ |
| Gonzalez, (2019) [21] | rats | SNx | omnivorous, individual cages | no randomization, unknown rat line and sex, no diet description | gut barrier functions↑ diabetes↑ LPS↑ |
| Huang, (2020) [22] | Balb/c male mice | SNx | randomization, blind testing, omnivorous colony animals | no housing description | GM alteration↑ gut injury↑ mRNA tight junction↓ gut permeability↑ |
| Yang, (2019) [23] | C57BL/6 male mice | SNx | omnivorous colony animals, SPF environment | no randomization, no diet description | GM alteration↑ claudin-1↓ claudin-2↑ gut permeability↑ inflammation↑ |
| Vaziri, (2013) [24] | Sprague– Dawley male rats | adenine diet | randomization, omnivorous colony animals | diet-induced CKD, no treated control | tight junctions↓ endotoxemia↑ inflammation, oxidative stress↑ |
| Wang, (2012) [25] | Sprague– Dawley male rats | SNx | randomization, omnivorous colony animals | no housing and diet description | Intestinal permeability↑ bacterial translocation↑ inflammation↑ |
| de Almeida Duarte, (2004) [26] | Wistar male rats | SNx | randomization, omnivorous colony animals | tap water | gut injury↑ bacterial translocation↑ |
| Vaziri, (2013) [27] | Sprague– Dawley male rats | SNx | randomization, omnivorous colony animals | no diet description | tight junctions↓ oxidative stress↑ |
| Yoshifuji, (2016) [28] | SHR male rats | SNx | randomization, omnivorous colony animals | no treated control, no housing and diet description | GM alteration↑ tight junctions↓ inflammation↑ |
| Vaziri, (2012) [29] | Sprague– Dawley male rats | SNx adenine diet | randomization, omnivorous colony animals, two CKD models | no diet description | tight junctions↓ mRNA tight junctions↑ inflammation↑ |
| Vaziri, (2020) [30] | Sprague– Dawley male rats | SNx | randomization, omnivorous colony animals | tight junctions↓ | |
| Lau, (2015) [31] | Sprague– Dawley male rats | SNx | randomization, omnivorous colony animals | no treated control, no housing and diet description | tight junctions↓ inflammation↑ oxidative stress↑ |
| Vaziri, (2014) [32] | Sprague– Dawley male rats | adenine diet | randomization, omnivorous colony animals | diet-induced CKD, no treated control | tight junctions↓ |
| Hung, (2018) [33] | ICR male mice | adenine diet | omnivorous colony animals, distilled water | diet-induced CKD, no treated control | GM alteration↑ gut barrier functions↓ inflammation↑ gut permeability↑ |
| Chen, (2019) [34] | Sprague– Dawley male rats | UUO | randomization, omnivorous colony animals | no housing and diet description | GM alteration↑ tight junctions↓ uremic toxins↑ SCFAs↓ inflammation↑ oxidative stress↑ |
| Hsu, (2020) [35] | Sprague– Dawley female rats | adenine diet | omnivorous colony animals | no randomization, diet-induced CKD | GM alteration↑ uremic toxins↑ SCFAs↓ |
| Yano, (2014) [36] | Wistar male rats | SNx | omnivorous colony animals | no randomization, no housing and diet description | mRNA ABCG2↑ uricase activity (stable) uric acid (stable) |
| Ji, (2020) [37] | Sprague– Dawley male rats | SNx | randomization, omnivorous colony animals, SPF environment | no treated control, no diet description | GM alteration↑ gut injury↑ tight junctions↓ inflammation↑ LPS↑ |
| Mishima, (2014) [38] | C57BL/6 male mice | adenine diet | randomization, omnivorous colony animals | diet-induced CKD, no housing description | GM alteration↑ gut barrier functions↓ uremic toxins↑ |
| Characteristic | Adenine-Enriched Diet-Induced CKD | Surgery-Induced CKD |
|---|---|---|
| gut motility | reduced [20] | reduced [17,18,19] (increased for dogs [16]) |
| leaky gut | not verified | induced [21,22,23] |
| bacterial translocation | induced [24] | induced [25,26] |
| tight junction | lower protein concentration [29,32,33,74] | lower protein concentration [21,22,23,28,29,30,31] structural alteration observed [22] inconclusive changes in tight junction mRNA [22,29] |
| transcellular transport in the intestine | not checked | increased expression of the ABCG2 urate transporter in the ileum [36] |
| gut microbiota | Metabolic changes: increased uremic toxins production, reduced SCFAs production [35] Composition changes: no similarity to the gut microbiota of CKD patients [35] | Metabolic changes: increased uremic toxins production, reduced SCFAs production [34,37] Composition changes: no similarity to the gut microbiota of CKD patients [34,37] |
| main disruptors of the intestinal microbiota | modified diet [38,68,69,70] | anesthetics, analgesics, antibiotics [71,72,73] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bartochowski, P.; Gayrard, N.; Bornes, S.; Druart, C.; Argilés, A.; Cordaillat-Simmons, M.; Duranton, F. Gut–Kidney Axis Investigations in Animal Models of Chronic Kidney Disease. Toxins 2022, 14, 626. https://doi.org/10.3390/toxins14090626
Bartochowski P, Gayrard N, Bornes S, Druart C, Argilés A, Cordaillat-Simmons M, Duranton F. Gut–Kidney Axis Investigations in Animal Models of Chronic Kidney Disease. Toxins. 2022; 14(9):626. https://doi.org/10.3390/toxins14090626
Chicago/Turabian StyleBartochowski, Piotr, Nathalie Gayrard, Stéphanie Bornes, Céline Druart, Angel Argilés, Magali Cordaillat-Simmons, and Flore Duranton. 2022. "Gut–Kidney Axis Investigations in Animal Models of Chronic Kidney Disease" Toxins 14, no. 9: 626. https://doi.org/10.3390/toxins14090626
APA StyleBartochowski, P., Gayrard, N., Bornes, S., Druart, C., Argilés, A., Cordaillat-Simmons, M., & Duranton, F. (2022). Gut–Kidney Axis Investigations in Animal Models of Chronic Kidney Disease. Toxins, 14(9), 626. https://doi.org/10.3390/toxins14090626