1. Introduction
The increasing occurrence of food/feed contaminants worldwide poses a huge threat to human and animal health. One of the major contaminants are mycotoxins, which annually cause enormous economic losses in the food industry and animal husbandry [
1,
2]. These low molecular weight metabolites produced by filamentous fungi (belonging to the phylum Ascomycota) contaminate various categories of foods and feeds [
3]. Two groups of mycotoxigenic fungi exist: field fungi (such as
Fusarium and
Aspergillus spp.) that infect crops before harvest, and storage fungi (such as
Penicillium spp.), which only occur after harvest [
4,
5]. According to a recent world survey based on around 97,000 analyses performed between January and December 2020 on more than 21,000 finished feed and raw commodity sources collected from 79 countries, the most prevalent mycotoxins were those produced by
Fusarium, affecting more than the 60% of tested samples [Biomin, Inzersdorf-Getzersdorf, Austria,
https://www.biomin.net/science-hub/world-mycotoxin-survey-impact-2021/, accessed on 4 January 2023], In 2020, Mesterhazy et al. highlighted how toxins are responsible for a loss of almost 700 mt during the harvest and storage of grains [
6]. At any stage of the food production process (in the field, during harvest, during drying and transport, as well as during storage), the fungal production of mycotoxins can occur by exposing consumers to the risk of contamination, either directly through food consumption or indirectly through feed [
7]. The most important mycotoxins are aflatoxins [mainly represented by aflatoxin B1 (AFB1), B2 (AFB2), G1 (AFG1), G2 (AFG2) and M1 (AFM1)], ochratoxins, fumonisins, trichothecenes, zearalenone, the emerging
Fusarium mycotoxins, ergot alkaloids,
Alternaria toxins, and patulin [
8]. Of the approximately 400 compounds identified as mycotoxins, 30 have received significant consideration due to their harmful effects on both human and animal health (including genotoxicity and endocrine disruption [
9]). Despite a huge number of published papers reporting the occurrence of mycotoxins on cereals and cereal-derived food products, in 2017, an analysis of 104 papers—from 2006 to 2016—was carried out, summarizing that mycotoxins are ubiquitously present in cereals and cereal-derived food products throughout the world [
10]. If Africa and Asia showed the highest incidence (%) of cereals contaminated by aflatoxins and ochratoxins, respectively, South and North America registered the highest level of fumonisins and Europe the highest percentage of deoxynivalenol (trichothecenes)/zearalenone contamination [
10].
Different physical, chemical and biological factors affect fungal colonization and mycotoxins production. Physical factors include environmental conditions such as temperature, relative humidity, pH, water activity, nutrients, insect infestation and other associated factors, which at specific rates are known to favor the growth of many types of fungi and the production of mycotoxins [
2]. Biological factors are mainly related to the interactions between the colonizing toxic fungal species and the host, thus including features such as fungal species, strain specificity, levels of inoculation, the nature of the substrate, strain variation, the instability of fungal toxic properties, and insect damage [
11].
Regulatory agencies have established strict legislative thresholds in order to keep the levels of mycotoxins in food/feed commodities under control. These limits range from below one to thousands of μg kg
−1, depending on the (i) mycotoxin, (ii) type of product and (iii) country considered [
12,
13]. Consequently, there is an urgent need to develop a feasible and highly sensitive analytical method for mycotoxins detection [
14] and reduce the contamination of mycotoxins in food/feed, in order to protect/preserve their quality and safety. Overall, the control of mycotoxin contamination follows two strategies: the prevention of their production (i.e., microbial inactivation) and detoxification (e.g., mycotoxin degradation [
3]). In pre-harvest, the control of mycotoxins is based on control of the contamination levels in crops to be used as food/feed components. Generally, these systems are based on preventive strategies (such as the use of resistant varieties, crop rotation, tillage and the management of irrigation) which aim to avoid the development of contamination, operating on the predisposing factors that facilitate the production of mycotoxins. Although pre-harvest approaches should be preferred, in the perspective of preventing mycotoxin contamination, the development of toxic fungi is inevitable under certain environmental conditions [
15]. Therefore, appropriate storage practices and other post-harvest control systems (at the microbiological, physical and chemical level) are necessary to minimize the final mycotoxin content of foods/feeds [
16]. These traditional methods for the elimination/inactivation of mycotoxins have some limitations regarding (potential) safety issues, loss in the nutritional value and the palatability of feeds, cost implication and limited efficacy [
17]. In recent decades, various detoxification approaches have demonstrated to be (i) highly effective in degrading mycotoxins into less toxic products, (ii) economically favorable and (iii) not environmentally harmful [
18]. Among these, cold plasma—containing reactive oxygen and nitrogen species and free radicals—has received attention in recent years for use on cereals during storage, due to its lethal effect on microorganisms and its potential to decontaminate surfaces and improve shelf-life [
19]. Nevertheless, their practical application in food/feed matrices is limited, since the degradation process under conditions of large-scale production is much more complex, and the experiments at lab-scale might not always reflect practices in industrial processing [
1]. Possible reasons for this are that the degradation process can be easily affected by multiple factors such as temperature, relative humidity, pH, water activity, nutrients, insect infestation and types of contamination [
2]. The relevance of studying naturally contaminated samples is consistent with the actual distribution conditions of mycotoxins in the field and/or in grains.
Since gas composition is considered one of the most important abiotic conditions that impacts fungal and pest growth, ozonation (i.e., the application of gaseous ozone, O
3) is a simple technology for controlling insects and reducing mycotoxins in stored products, which does not leave harmful residues after application. Being unstable, O
3 quickly degrades into oxygen (and related cytotoxic radicals) in a short period, oxidizing the vital cellular components (such as unsaturated lipids and proteins) of pathogenic microbes and storage pests by causing lysis and rapid cell death [
20]. Consequently, O
3 can inhibit fungal growth, sporulation and germination by offering a negligible loss of nutrients or sensory qualities in food/feed [
21], making it a suitable candidate as a residue-free fumigant. For this reason, the application of O
3 in food chains has been considered safe and effective by the WHO and is now recognized as a “green technology” for the fumigation of grains, fruits and vegetables [
22]. In fact, O
3-treated products are safe for consumption and their microbiological shelf life can be greatly enhanced. However, the efficacy of O
3 in fungi count reductions, mycotoxin degradation and insect control depend on the (i) method, concentration and timing of the O
3 application; (ii) microorganisms/contaminants to inactivate; (iii) the type and mass of food/feed processed; and (iv) other co-factors such as temperature, relative humidity and water activity [
16,
20]. Similarly, the use of a controlled or modified atmosphere by using a very high nitrogen (N
2) concentration is a valid alternative to chemical fumigation to control mycotoxigenic fungi contamination and pest challenge post-harvest [
23]. Its effects on different stored products (such as wheat, maize, corn and rye) are well documented [
2,
24]. In particular, a N
2-controlled atmosphere can control fungal growth and proliferation by improving the quality of stored products [
25]. The action of N
2 at high concentration is mainly due to the significant reduction in O
2 (1% or less [
26]) and offers several advantages at the economic and environmental level [
27]. A major advantage of N
2 is that all gas is free of pollutants, leaving no residue in food/feed. Consequently, N
2-treated products are safe for consumption and their microbiological shelf-life can be greatly enhanced [
28]. Consequently, a N
2-controlled atmosphere might represent an eco-friendly tool that could be transferred to a large-scale system for grain storage as an alternative strategy to the use of conventional residue-producing chemical fumigants [
29]. However, the efficacy of a N
2-controlled atmosphere in fungi count reductions, mycotoxin degradation and insect control depends on the (i) concentration of gas, (ii) the timing of the application, (iii) microorganisms/contaminants to inactive, (iv) the type and mass of food/feed processed, and (v) other co-factors such as temperature and water activity [
30].
The chickpea (
Cicer arietinum L.) is a legume of the family Fabaceae, subfamily Faboideae. It is one of the most cultivated pulses in terms of world production due to its low content of fat and sodium, absence of cholesterol and being an excellent source of both soluble and insoluble fiber, complex carbohydrates, vitamins, folate and minerals (such as calcium, phosphorus, iron and magnesium [
31]). With a worldwide production of more than 12 million tons per year [FAOSTAT,
https://www.atlasbig.com/en-in/countries-by-chickpea-production, accessed on 4 January 2023], chickpea represents one of the five leading pulses based on sales value. In 2021, India was the largest chickpea producer in the world, with around 11 million metric tons of production, followed by Turkey, with around 600,000 metric tons (
https://www.statista.com/statistics/722203/chickpeas-production-volume-by-country-worldwide/, accessed on 4 January 2023). Chickpea is often attacked by fungi pre- and post-harvest, significantly affecting its productivity. Many fungal genera/species commonly isolated from chickpea seeds and chickpea by-products are potential mycotoxin-producers, especially of aflatoxins, ochratoxin A and patulin, so there would be a potential risk of contamination [
32]. Another issue threatening chickpea quality is
Callosobruchus maculatus (Fab.) (Coleoptera: Chrysomelidae: Bruchinae [
33]), which is also known as the “cowpea weevil”. The granivorous larvae of cowpea weevil are the considerable causative agent of severe losses in the grain germination, weight and nutritional values of chickpea (in some cases reaching 60% of the grain [
34]). In addition,
C. maculatus can favor the occurrence of infections due to mycotoxigenic fungi (
Aspergillus and
Penicillium [
35]).
The aim of this work was to investigate the possibility of using a single pulse of O
3 or high N
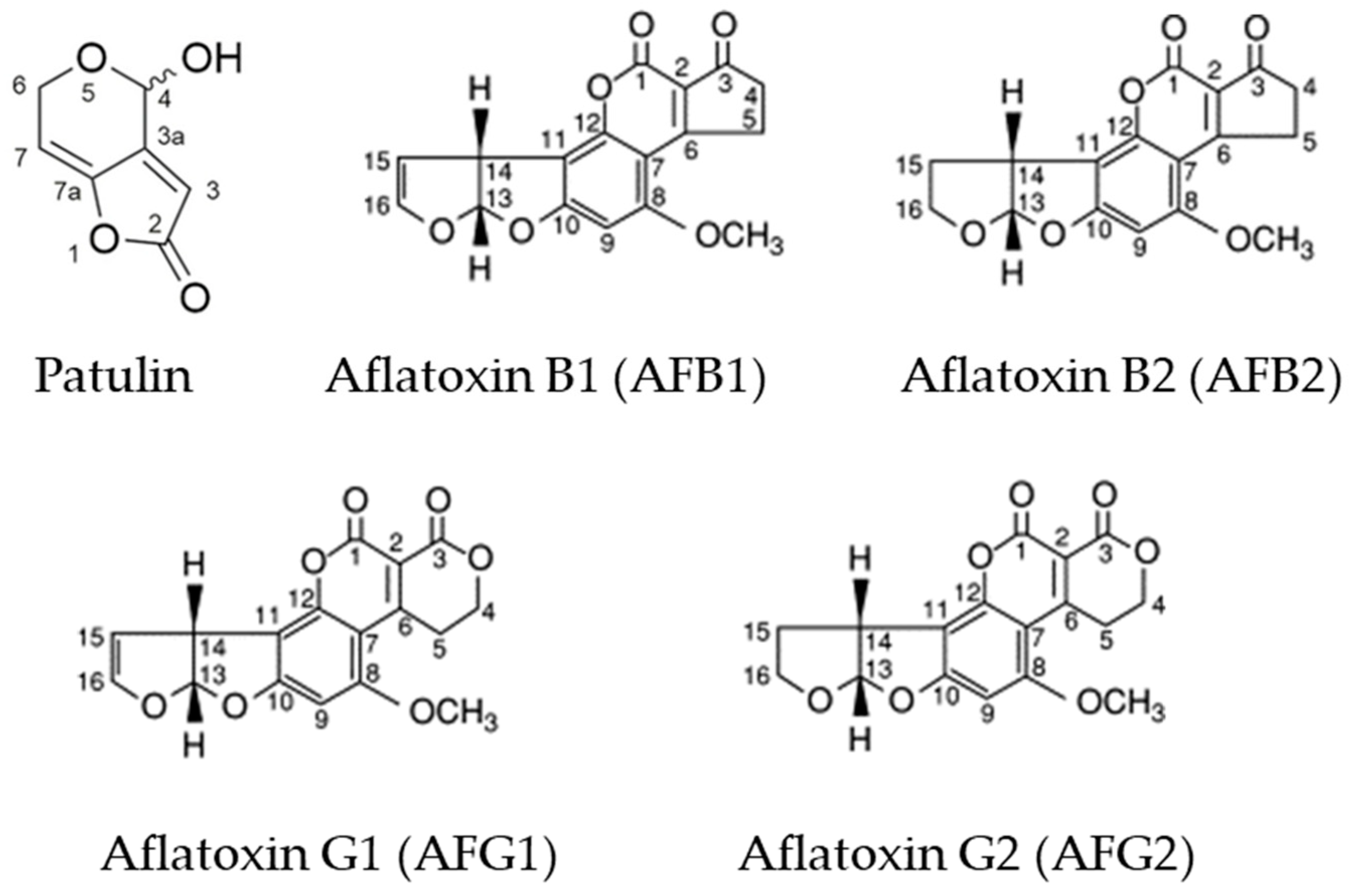
2 concentrations as storage technologies (at lab- and large-scale) for the purpose of (i) containing the fungal population present on the chickpea seeds surface; (ii) reducing the mycotoxins content (such as aflatoxins and patulin;
Figure 1); and (iii) limiting the
C. maculatus infestations (only in the case of N
2 treatment). We postulated that O
3 and N
2 can be an alternative to traditional chemical-based fumigants for controlling spoilage pathogens and insects in stored chickpea seeds.
3. Discussion
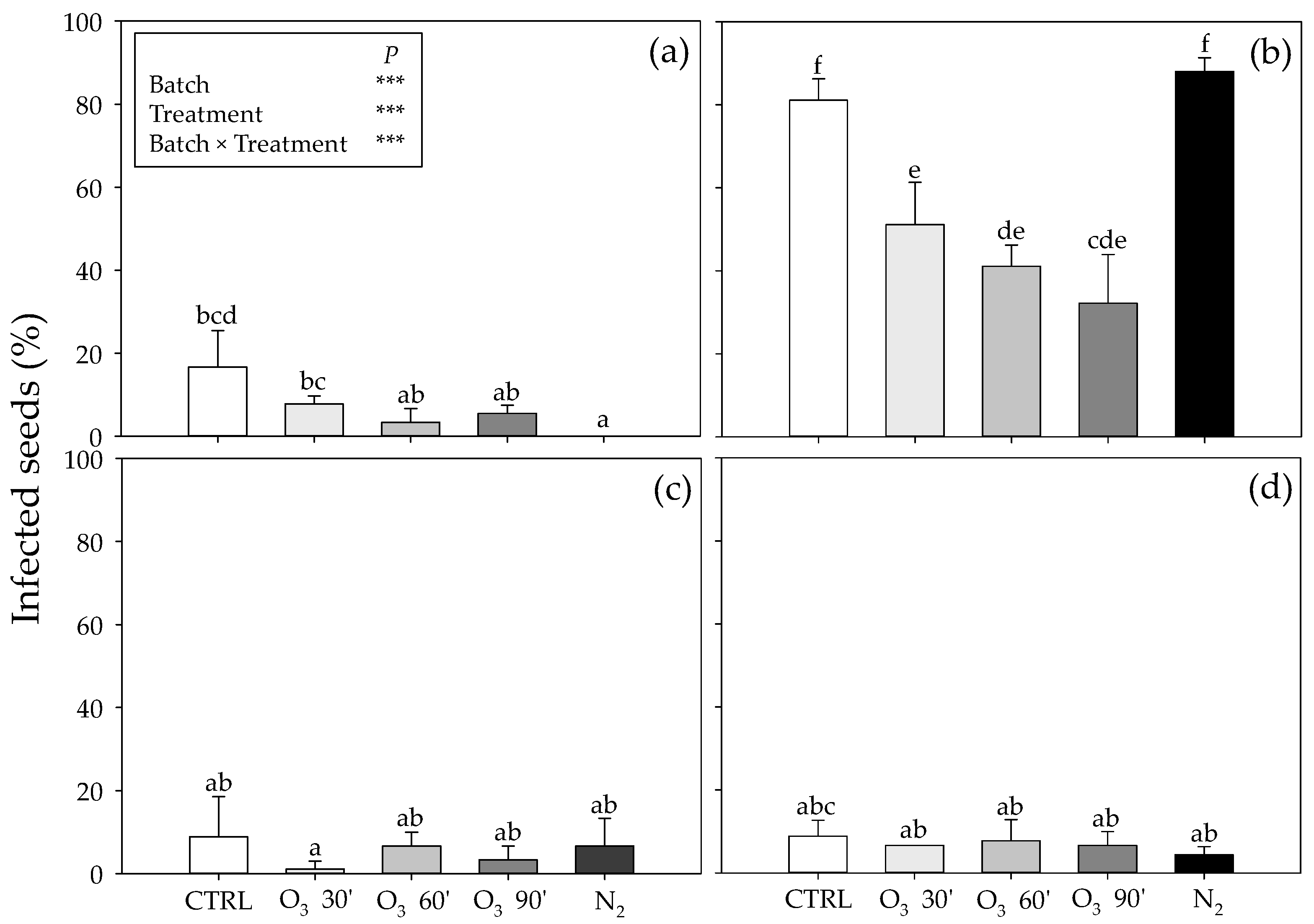
In the present study, the fungicidal efficacy of O
3 was only observed in batch n. 2 (the most infected seeds), as confirmed by the large reduction in
Penicillium contamination. According to Mendez et al. [
36], at the beginning of O
3 treatment, this gas reacts with a mass of grains and quickly decomposes. In its second phase, O
3 moves freely through the grains with little degradation. Moreover, O
3 reacts faster with the whole mass of grains when higher dosages are used. This was also confirmed by our results: a higher reduction in the naturally occurring fungal population was observed by using O
3 at 500 ppb for 90 min. To date, few studies have investigated the direct effect of O
3 on fungal growth and proliferation, preferring to focus on the reduction in mycotoxins in different products (such as wheat, corn, corn flour, peanuts and pistachio [
37]). Most studies have been carried out on aflatoxins. Using O
3 in patulin, degradation has only been studied in apple juice, apples and pear with brown rot, flour, and malt feed [
38]. In the present study, O
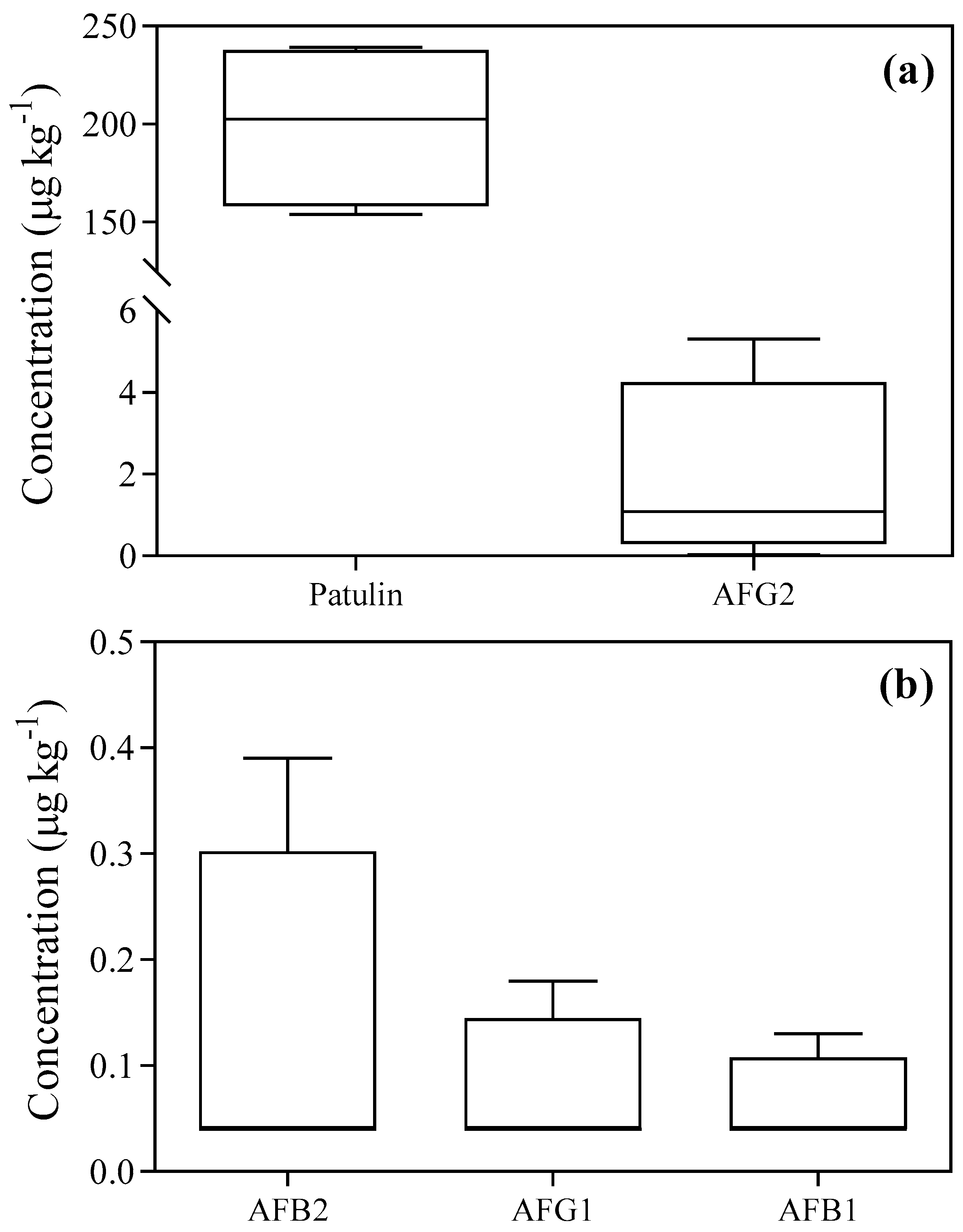
3 treatment induced a significant decrease in patulin in batch n. 2 (independently of its duration), and even more in the remaining batches. The mechanism of patulin degradation might be associated with the oxidation of a polyketide lactone on its structure, which made it highly susceptible to O
3 attack [
39]. The experimental conditions used in this study (e.g., O
3 and/or the timing of treatment) were sufficient, by attacking two conjugated ethylenic double bonds on the chemical structure of patulin and inducing its partial or full degradation [
40]. Similarly, all aflatoxins were easily attacked and degraded by O
3 (independently of its duration), confirming its efficacy as a detoxifying agent. The mechanism of AFB1 and AFG1 degradation might be associated with the oxidation of the C8-C9 double bond at the terminal furan, resulting in the production of primary ozonide [
26]. This product may rapidly rearrange to a molozonide derivative, yielding a variety of carbonyl compounds or organic acids. Since AFB2 and AFG2 lack a susceptible double bond for oxidation, their degradation requires higher levels of O
3 and/or longer exposure until the lactone ring is opened [
41]. The experimental conditions used in this study (e.g., O
3 concentration and/or the timing of treatment) were sufficient, by rapidly and effectively detoxifying aflatoxins without any difference in degradation rate between AFB1 and AFG1 with AFB2 and AFG2 (the most abundant aflatoxin in batches n. 2, 3 and 4). It is worth noting that the production of aflatoxins was not associated with the presence of the fungal itself, confirming that the absence of
Aspergillus spp. from chickpea seeds does not guarantee the absence of aflatoxins because of their resistant chemical nature [
42].
In the present study, high N
2 concentrations only induced a reduction in
Penicillium contamination in batch n. 1 confirming that certain fungal species might continue to grow, albeit at a greatly reduced rate under low O
2 concentrations [
43]. However, a significant decrease in patulin was observed in batches n. 3 and 4, and even more in the remaining ones, indicating that low O
2 concentrations may (partially or totally) depress patulin production by
Penicillium spp. on chickpea seeds. In addition, the experimental conditions used in this study (e.g., N
2 concentration and/or the timing of treatment, temperature, relative humidity and water activity) were sufficient, by totally removing aflatoxins. Consequently, it is possible to speculate that high N
2 concentrations are more effective in inhibiting selected mycotoxins (aflatoxins > patulin) than in preventing the development of mycotoxigenic fungi [
44]. Although a controlled atmosphere is used to control both mycotoxigenic fungi and insects in stored products, it has been documented that the experimental conditions sufficient for controlling fungal proliferation are not always effective against insect pests that can survive, due to the dependence on other environmental factors (e.g., temperature and humidity [
45]). This is in line with the results of the present work, where the same experimental conditions (e.g., N
2 concentration and/or the timing of treatment, temperature, relative humidity and water activity) that were partially effective for reducing the growth of
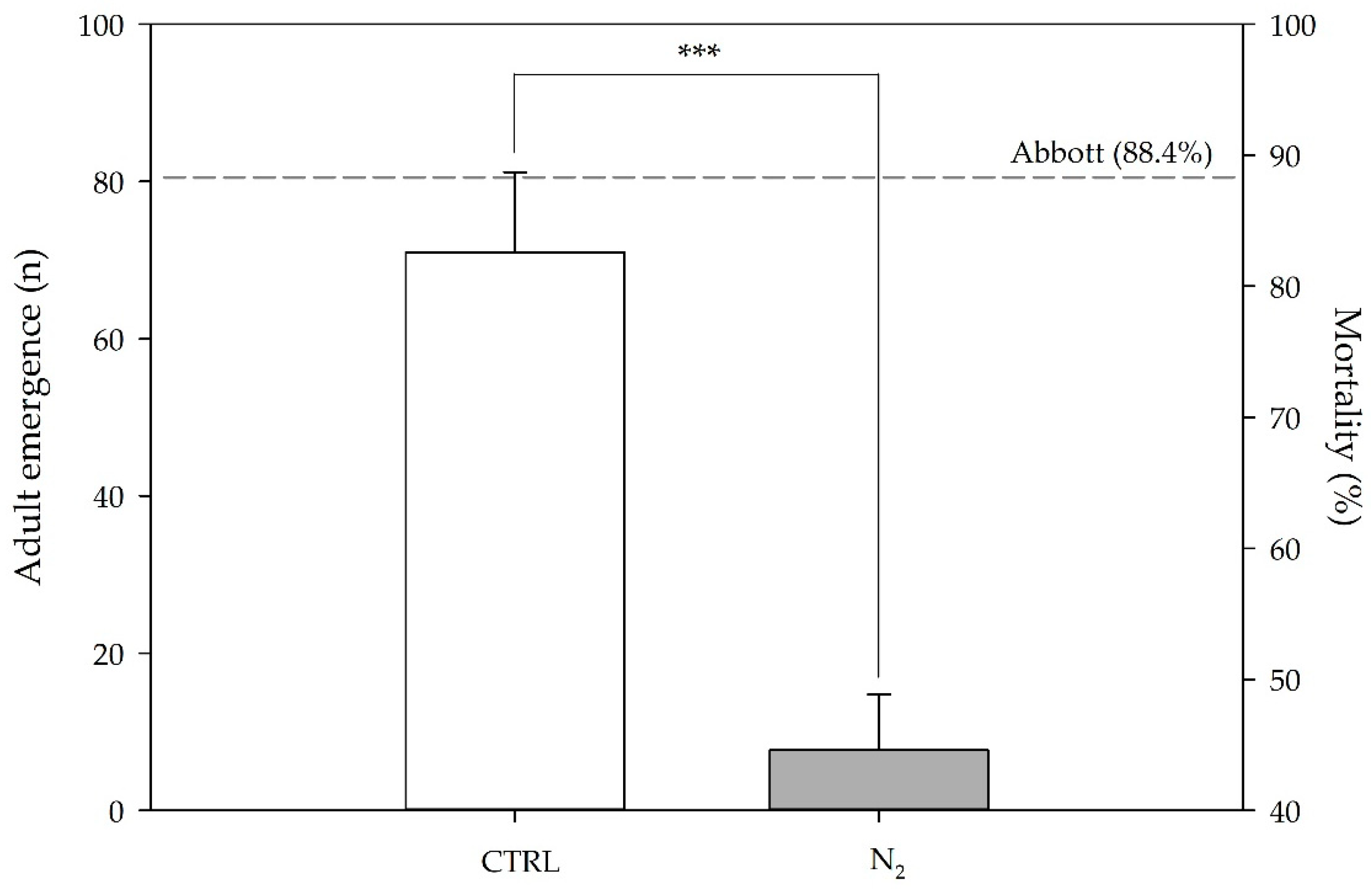
Penicillium spp. were effective for detoxifying mycotoxins (as previously reported) and limiting
C. maculatus infestation (as confirmed by the reduction in adult emergence already observed starting from the first day of the experiment, and the concomitant pest mortality). This is of relevance, since it is generally recognized that pest attack can damage grains and favor moisture accumulation by creating suitable conditions for fungal development and mycotoxin production (as observed in batch n. 2 [
46]). Consequently, it is possible to speculate that the experimental conditions used in this study were sufficient, by limiting the pest infestation and guaranteeing seed quality, as already reported in wheat [
23,
25]. In addition, the possibility of moving to a real-scale dimension, as here preliminary reported, with results comparable with those obtained in the 60 L lab-scale silos, marks a further step made in the direction of the scaling-up of the method. In fact, the set-up chosen for the present study is not only a suitable way to provide a proof-of-concept of its efficacy before scale-up, but also a valid choice for small and medium farms [
23,
25].
The findings of the present study are relevant in enhancing the shelf-life of chickpea seeds by controlling fungal growth, mycotoxin contamination and pest infestation with eco-friendly and low-cost storage practices (the mechanisms of action of O
3 and N
2 are summarized in
Figure 7).
In particular, O
3 can contain the fungal population present on the chickpea seeds surface and reduce the content of patulin and aflatoxins. This is a fundamental goal in the development of emerging new techniques, since these metabolites are recognized as a Group I carcinogen by the International Agency of Research on Cancer (Lyon, France), and their allowable levels in human foods and animal feedstuff are strictly regulated by governmental jurisdictions in about 100 countries [
47]. It is worth noting that the antimicrobial activity of O
3 is highly dependent on vegetable/fungus species, growth stage, concentration and timing of exposure. Improvements and innovations in O
3 generation and application systems will be evaluated more effectively in the future by facilitating the enhanced control of both the quality and safety parameters of ozonized foods/feeds. Similarly, a N
2-controlled atmosphere represents a valid sustainable method to limit mycotoxin accumulation and
C. maculatus infestation. This system requires low energetic costs, offers a negligible loss of nutrients or sensory qualities in food/feed, and demonstrates a reduced hazard to employees with no need for registration and no contamination of the environment [
23,
25]. Consequently, it can be considered as a promising alternative method that could be transferred to a large-scale grain storage system. For effective and safe use in processing, optimum O
3 and N
2 concentrations, contact time and other treatment conditions should be defined for foods and feeds. Here, a pilot test was conducted by offering scientific evidence to support the commercial application of these innovative strategies.
5. Materials and Methods
5.1. Reagents and Standards
Sodium hypochlorite, ethanol and streptomycin sulphate were supplied by Sigma-Aldrich (Milan, Italy). Potato Dextrose Agar was purchased from Biolife (Milan, Italy). Acetonitrile, methanol and water were HPLC-grade (Carlo Erba, Milan, Italy). Standards of patulin and aflatoxins were chromatographically pure and purchased from Sigma-Aldrich (Milan, Italy) and Romer Lab (Getzersdorf, Austria).
5.2. Raw Materials
Four C. arietinum batches, produced by a local farm located in Tuscany (Italy), were used in the present work. Before commercialization, all the batches (except in the case of n. 3) were submitted to a quality check by the producer, thus resulting in batches n. 1 and n. 4 being validated for sale, and batch n. 2 rendered non-compliant. All seeds were stored at 4 °C until submitted to a grain health test, mycotoxin determination and treatments under a controlled atmosphere.
5.3. Isolation and Identification of Fungal Contaminants Naturally Associated with Chickpea Seeds
To assess the presence of potential mycotoxigenic fungi naturally associated with chickpea seeds, all four batches were submitted to a grain health test. In detail, seeds from each batch were surface sterilized on a rotary shaker for 1 min in a solution containing NaClO (1% active chlorine) in 50% ethanol, then washed three times in sterile distilled water for 1 min each. After drying on filter paper, the seeds were transferred to 100 mm diameter Petri dishes containing Potato Dextrose Agar (PDA, 42 g L
−1) with the addition of 300 mg L
−1 of streptomycin sulphate. Since batch n. 2 showed a profuse development of
Mucor spp., the seeds from this sample were plated on PDA that contained the antibiotic, as previously described, and with hymexazol fungicide (at the final concentration of 300 mg L
−1; [
48]). For each batch, four replicates (each consisting of twenty-five seeds) were made. The plates were incubated at room temperature (24 ± 2 °C). Then, from the second to the tenth day of incubation, colonies morphologically attributable to
Aspergillus and
Penicillium spp., developing from seeds, were transferred to new PDA + streptomycin sulphate plates and incubated under the same conditions previously reported. Then, when sporulated, they were used for single-spore cultivation. The single-spore
Penicillium spp. colonies were used for molecular identification. Genomic DNA was extracted from each single-spore culture according to the Chelex 100 method [
49]. For molecular identification, the complete internal transcribed spacers (ITS) 1 and 2 sequences—including the 5.8S gene—of the nuclear ribosomal DNA were amplified and sequenced as described in Sarrocco et al. [
30]. All the sequences were then submitted to GeneBank (NCBI) to assign, where possible, the species for a preliminary evaluation of the risk of mycotoxin contamination that could occur on the seeds.
5.4. O3 and N2 Treatment Systems for Chickpea Seeds
5.4.1. O3 Treatment System for Chickpea Seeds
Chickpea seeds (300 g from each batch) were placed in two Perspex chambers (60 × 60 × 110 cm) in a controlled environment fumigation facility. The system was adapted by including commercial colanders (34 × 23 cm, stainless steel), collocated in the middle of the chambers, in which seeds were placed to allow their complete exposure to O
3, maintained in the dark throughout the whole period of the experiment (temperature 25 ± 1 °C, relative humidity (RH) 50 ± 5%). The fumigation system was continuously ventilated (two complete air changes per min) with charcoal-filtered air. Fumigation was performed by generating O
3 from pure oxygen by electrical discharge, using a Fisher 500 air-cooled apparatus (Fisher America Inc., Houston, TX, USA). The O
3 concentration was monitored with a Serinus 10 analyzer (Ecotech Acoem Group, Milan, Italy) set at 500 ± 50 ppb of O
3 (for O
3, 1 ppb = 1.96 μg m
−3, at 20 °C and 101.325 kPa) for 30, 60 and 90 min, in which the chickpea seeds were mixed every 15 min. At the end of each treatment, the samples were collected and immediately used for the subsequent analyses (fungal infection and mycotoxin contamination). The entire methodology was performed according to Marchica et al. [
50].
5.4.2. Nitrogen Treatment System for Chickpea Seeds
Nitrogen treatments were performed using a NitrosepAgri system (Eurosider sas, Grosseto, Italy) based on selective membrane (MNS, Membrane Nitrogen Separator) to separate N
2 from atmospheric air [
25]. The N
2-enriched atmosphere was driven into silos where it was maintained under a slight overpressure. All the environmental parameters, such as temperature and RH, were constantly monitored, and the N
2 percentage could be set, automatically maintained and quickly reintegrated if needed. To perform the experiments here reported, two lab-scale (20 and 60 L silos, respectively) prototypes, already described in Moncini et al. [
25], and one field-scale apparatus were used. For each chickpea batch, 300 g of seeds were transferred into a 1.5 L glass jar (9 cm diameter) that was closed with a micro-perforated nylon layer (350 μm pore size) to facilitate gas exchange and incubated under a 99% N
2 atmosphere for 21 consecutive days. For each treatment and for each batch, three replicates were made. The field-scale apparatus consisted of four 15 m
3 volume fiberglass silos connected to the MNS system. Each silos had a stainless steel (786 mm diameter) top hatch for grains charging, equipped with an overpressure valve and, on the bottom, with a stainless-steel ball (90 mm diameter) valve for discharging the product. The field-scale apparatus is located at the Azienda Agraria Macchiascandona (Castiglione della Pescaia, Grosseto, Italy).
5.5. Effect of O3 and N2 Treatments on Fungal Infection, Mycotoxin Contamination and Pest Survival
At the end of the O3 and N2 treatments, the seeds were collected and used to evaluate fungal contamination according to the grain health protocol. Mycotoxins determination was performed by using the clean-up aflatoxins and patulin (AFP) columns (OR SELL, S.p.a., Modena, Italy), according to the manufacturing protocols, with a few modifications. The samples were extracted by adding 50 mL of acetonitrile:water (ACN:H2O, 84:16 v/v) solution to 25 g of finely ground chickpea seeds, and vigorously vortexed for 6 min. The samples were centrifuged for 10 min at 12,000× g at room temperature, and the supernatants were filtered through Whatman® paper (Cytiva, Marlborough, MA, USA) and subsequently by using the clean-up AFP columns, which enable the contemporary purification of aflatoxins and patulin. The obtained solutions were equally separated, dried at 40 °C and finally resuspended in 400 µL of 45% methanol (v/v) or 75% ACN (v/v in HPLC-demineralized water) for aflatoxins and patulin separation, respectively. The separation was performed in a UHPLC Dionex UltiMate 3000 system (Thermo Scientific, Waltham, MA, USA) equipped with a ZORBAX Eclipse Plus C18 column (150 × 4.6 mm, 5 μm particle size, Agilent technologies, Santa Clara, CA, USA). Aflatoxins determination was carried out by using an UltiMate™ 3000 Fluorescence Detector (Thermo Scientific, Waltham, MA, USA) with excitation and emission at 362 and 420 nm, after post-column derivatization through a UVE™ Photochemical Reactor for Aflatoxin Analysis (254 nm lamp; 240 VAC, 50/60 Hz, LCTech, Obertaufkirchen, Germany). The run conditions were set at a flow rate of 0.8 mL min−1 of a mobile phase 45% methanol (v/v in HPLC-demineralized water), for 30 min at 30 °C. Patulin quantification was performed using a Dionex UV-Vis Detector (Dionex UVD 170 U UV-Vis detector, Thermo scientific, Waltham, MA, USA) at 276 nm, at a flow rate of 0.6 mL min−1, for 30 min at 30 °C, and the same mobile phase reported above. Known amounts of pure (Patulin HPLC standard) or mixed (Aflatoxin Mix 4 solution) standards were injected into the UHPLC system (range 0.1−100 ng mL−1), and the quantity of each mycotoxin was obtained by correlating the peak area to the related standard concentration by using the Chromeleon Chromatography Management System software, version 7.2.10-2019 (Thermo Scientific, Waltham, MA, USA). The sum of AFB1, AFB2, AFG1 and AFG2 was considered as a measure of the total aflatoxins content.
Stock cultures of
C. maculatus, kindly provided by Graham J. Holloway (University of Reading, Reading Berkshire, UK), were maintained in a climatic chamber (25 ± 1 °C, 70 ± 5% RH) on chickpea seeds in a 150 mL PP jar (5.5 cm diameter) that was closed with a micro-perforated nylon layer (350 μm pore size) to facilitate air exchange. To obtain chickpea infested with eggs, pest sub-cultures were set up by placing 100 unsexed adults into glass Petri dishes (14 cm diameter) containing 100 g of chickpea seeds. Adults were allowed to oviposit for 24 h in a climatic chamber under the same conditions as described before. At the end of the oviposition period, the adults were removed. Seeds with the addition of age-synchronized eggs were immediately used for the N
2-controlled atmosphere test [
51]. The lab-scale experiment was set up in six 60 L silos (three under a 99% N
2 atmosphere and three under an unmodified atmosphere used as control). Two bio-tests, each consisting of 15 g of chickpea seeds with the addition of 1-3 eggs (up to a total of 30 eggs) placed into a 30 mL PP jar (4.3 cm diameter) and covered with a micro-perforated nylon layer (350 μm pore size), were transferred into each of the 60 L silos, which were already full of chickpea seeds; one bio-test was placed at the center and the other 10 cm under the grains’ surface. The infected seeds were incubated for 24, 48, 72, 96 and 120 days. To avoid the opening of the silos at each sampling, the experiment was repeated each time, with three replicates for each experiment.
With respect to the field-scale experiment, two silos filled with 9 tons of chickpea seeds (the same batch as the laboratory) were used for the tests: one of these was automatically kept at a 99% N
2-controlled atmosphere, and the other one, with the lid partially opened and disconnected from the N
2 supply, was used as a control. In order to simulate a storage condition inside the full grain mass, three bio-tests previously placed into a jute bag filled with 8 kg of chickpea seeds were transferred into the silos. The test was carried out for 5 days and replicated three times. At the end of each experiment (lab-scale and large-scale test), the bio-tests were removed from the silos and their contents transferred into larger PP jars (like those used for pest stock cultures), closed with a micro-perforated nylon layer and placed in a climate chamber at 25 ± 2 °C, 70 ± 5% RH. The adults’ emergence was periodically recorded from 35 to 70 days after oviposition. The number of adults was registered, and mortality was corrected using Abbott’s formula [
52].
where N
c = No. of emerged adults in control and N
t = No. of emerged adults in treatment.
5.6. Statistical Analysis
The robustness of data among the replicates was verified according to the results of the Shapiro–Wilk test for normality and Levene’s tests for homogeneity of variance. Data were submitted to an analysis of variance (one-way or two-way ANOVA), and comparisons among the means were determined by Tukey’s HSD post hoc test or Student t-test by using JMP Pro 14 software (SAS Institute Inc., Cary, NC, USA), in order to evaluate the effect of the treatments (control silos; O3 exposure for 30, 60 and 90 min; and N2 exposure for 21 consecutive days), batch (B1-4), and their interaction. For all the analyses, p ≤ 0.05 was assumed as a significant level.


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






