Impacts of Climate Change on the Biogeography of Three Amnesic Shellfish Toxin Producing Diatom Species

 ,
,  and
and 
Abstract
:1. Introduction
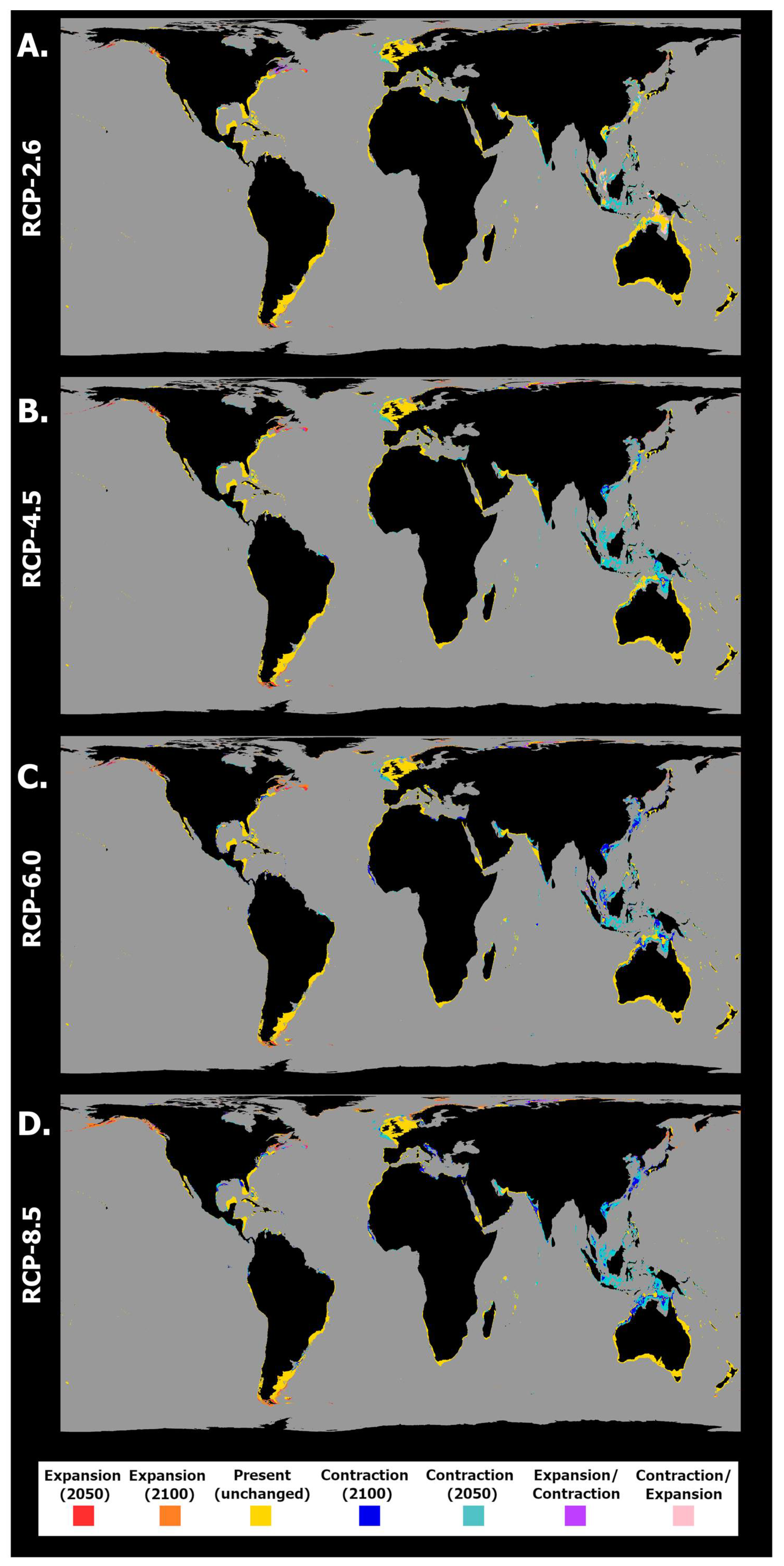
2. Results
2.1. Variable Contributions
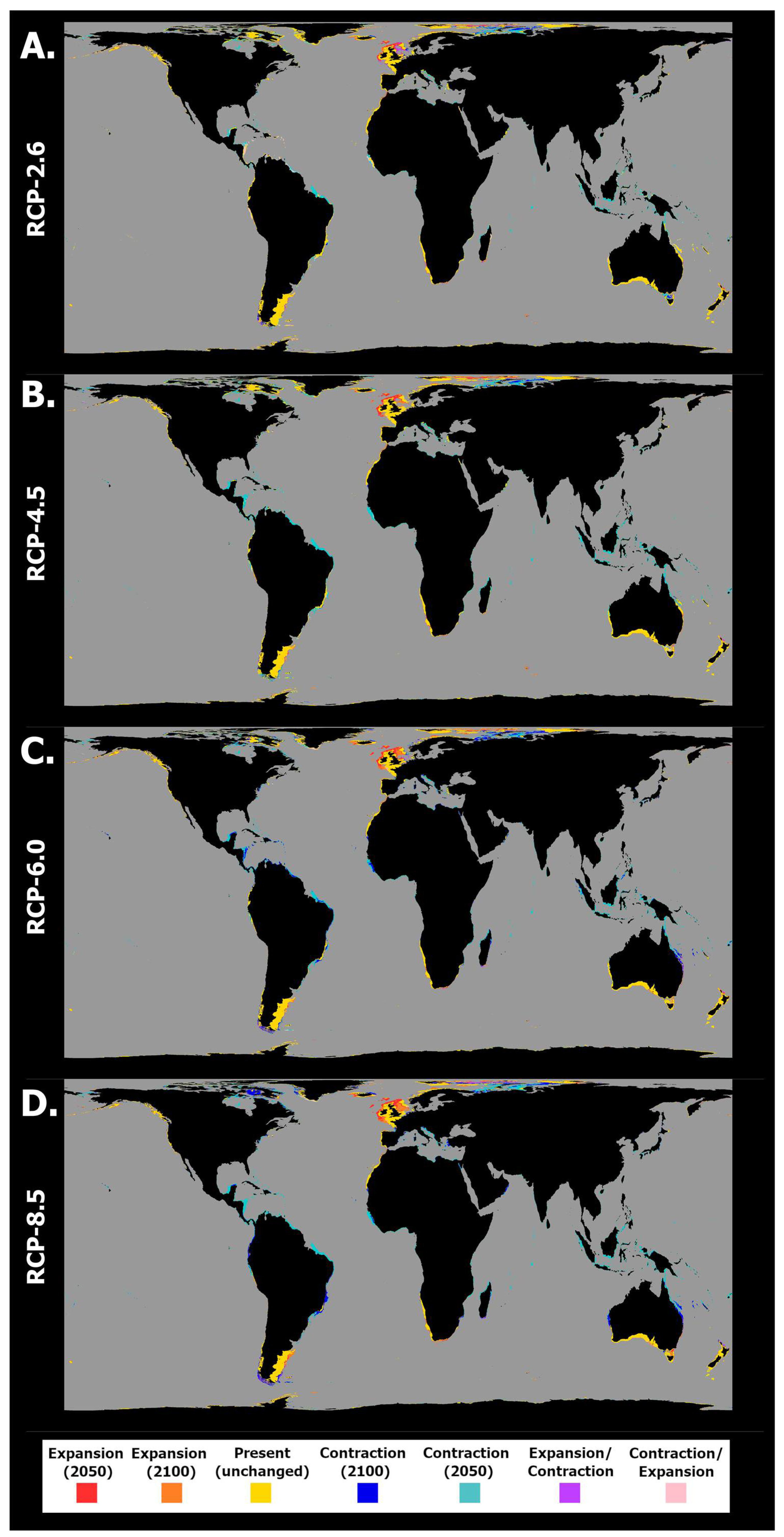
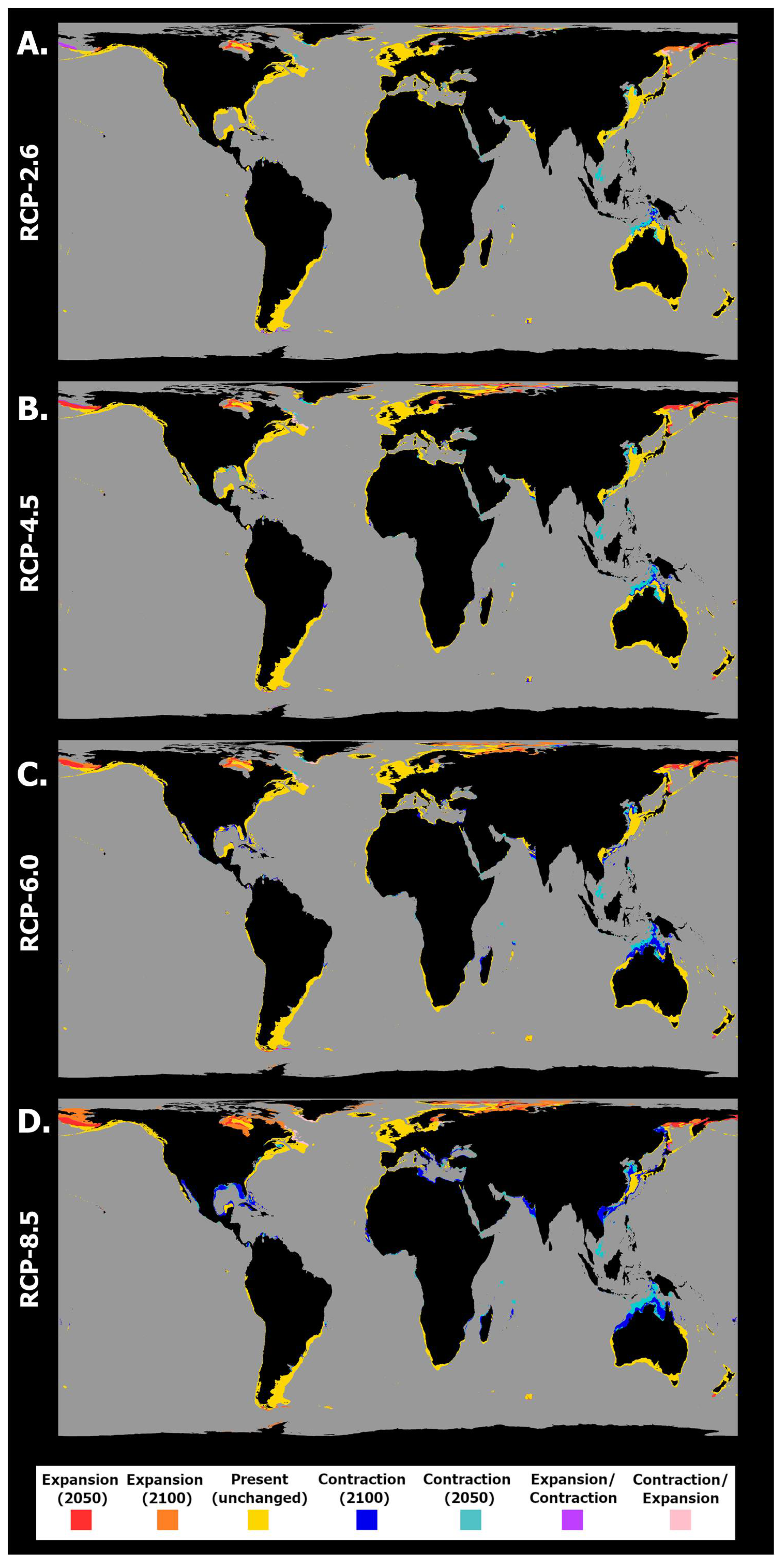
2.2. General Patterns of Habitat Suitability
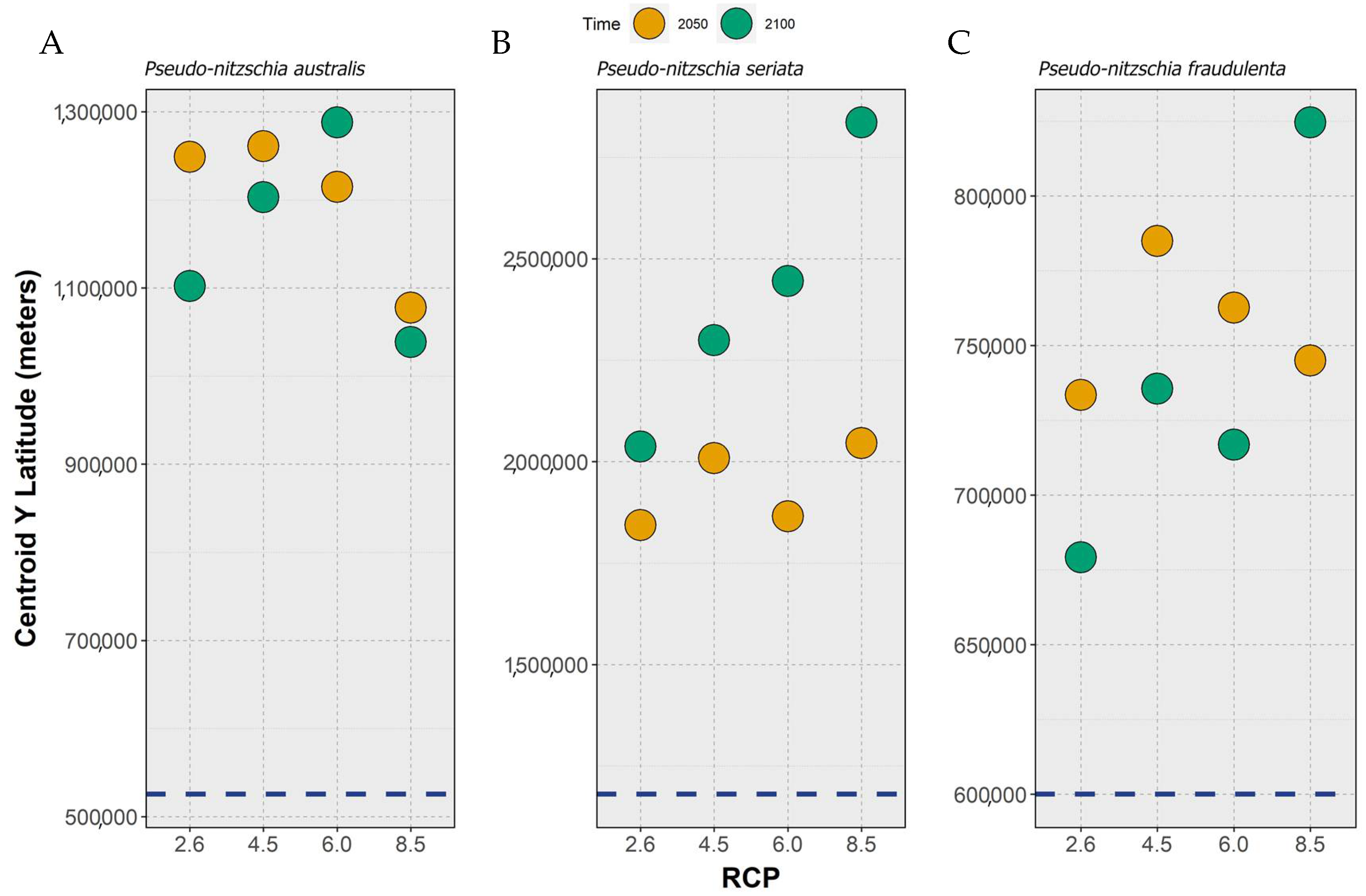
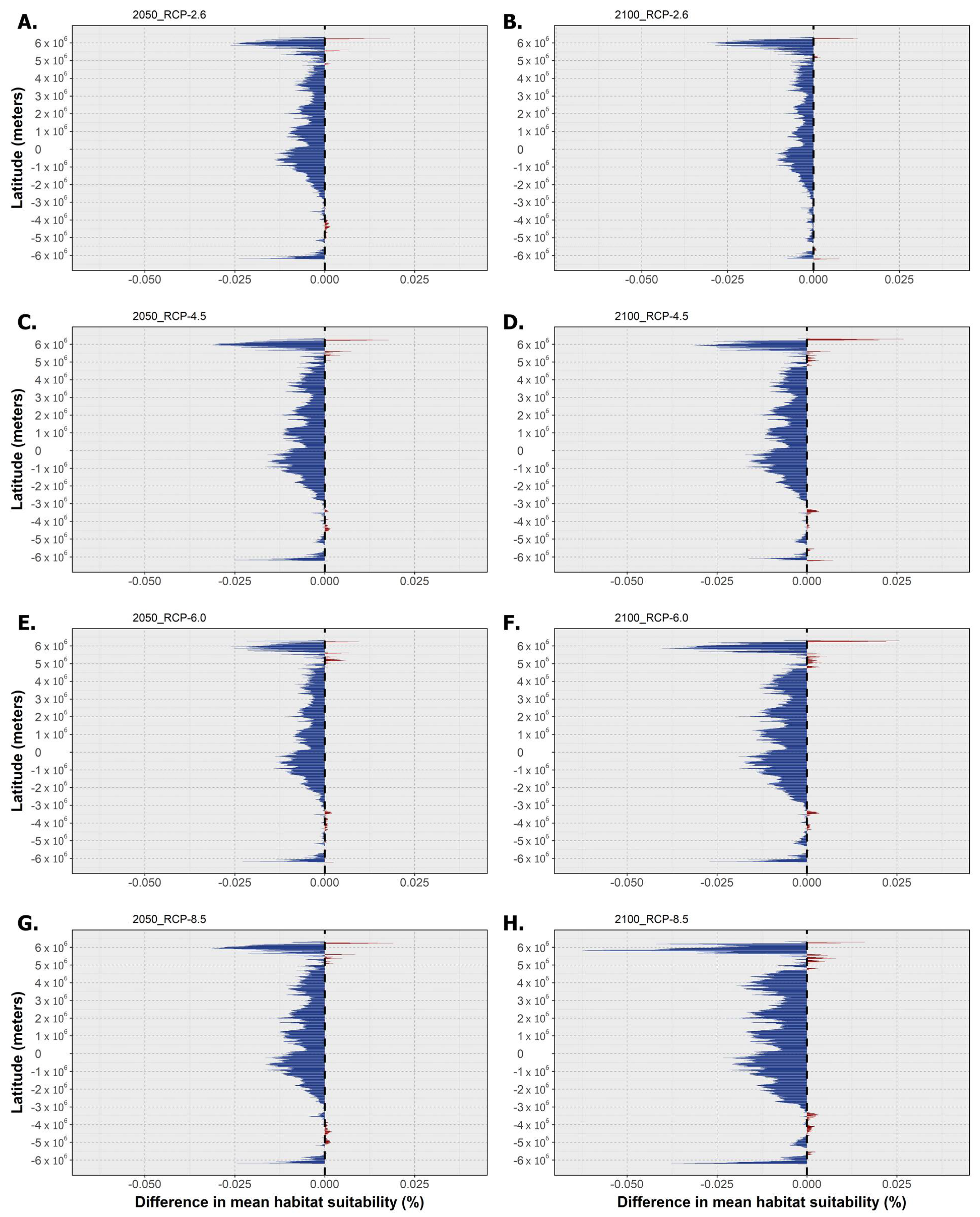
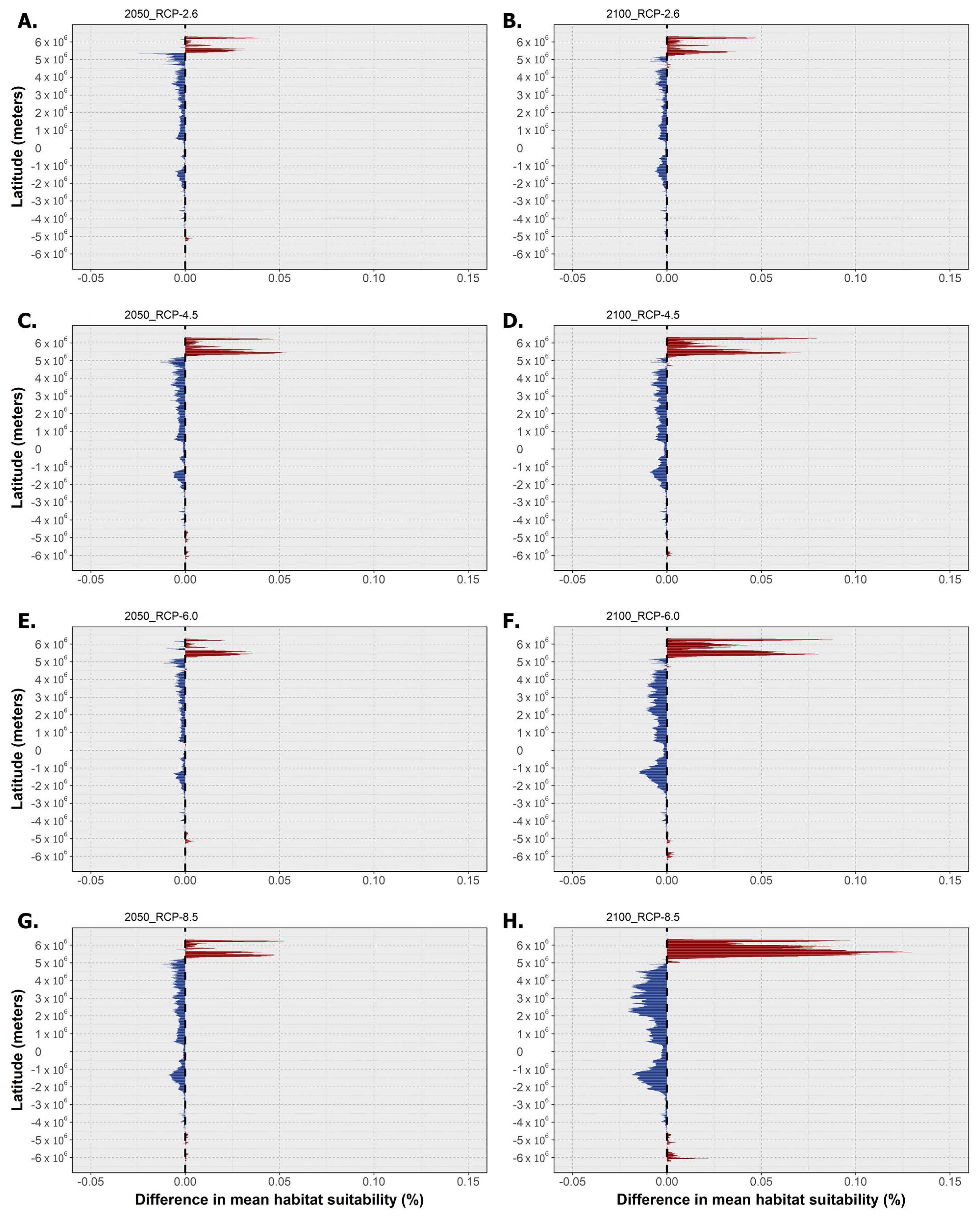
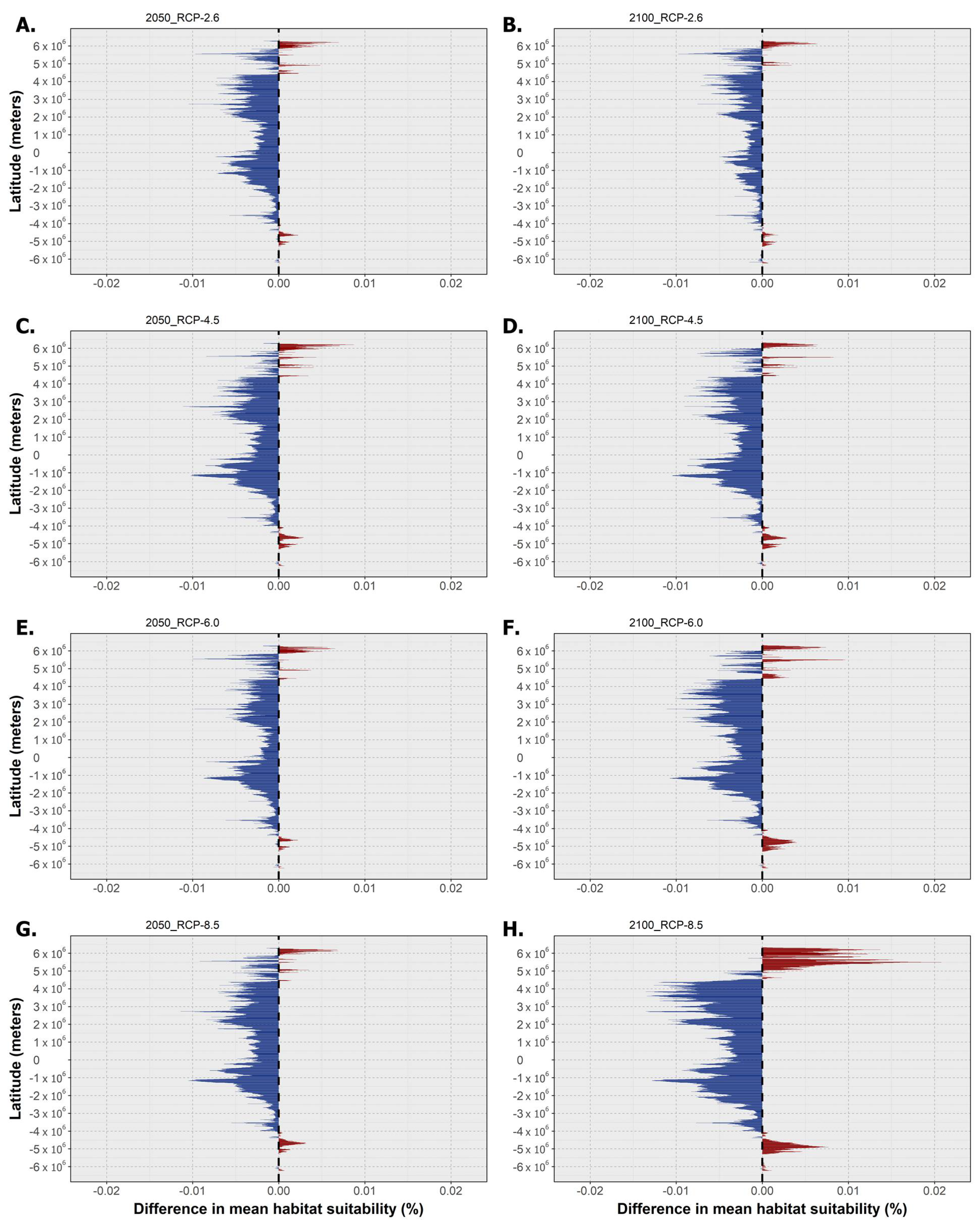
2.3. Habitat Suitability and Occurrence Distribution: Latitudinal Trends
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Data Collection and Curation
5.2. Environmental Predictors
5.3. Premodeling Procedures
5.4. Modeling
5.5. Postanalysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bindoff, N.; Cheung, W.; Kairo, J.G.; Aristegui, J.; Guinder, V.; Hallberg, R.; Hilmi, N.; Jiao, N.; Karim, M.; Levin, L.; et al. Changing Ocean, Marine Ecosystems, and Dependent Communities (09 SROCC Ch05 FINAL-1). In IPCC Special Report on the Ocean and Cryosphere in a Changing Climate; IPCC: Geneva, Switzerland, 2019; pp. 447–588. [Google Scholar]
- Arias, P.; Bellouin, N.; Coppola, E.; Jones, R.; Krinner, G.; Marotzke, J.; Naik, V.; Palmer, M.; Plattner, G.K.; Rogelj, J.; et al. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. In The Intergovernmental Panel on Climate Change AR6; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Kroeker, K.J.; Kordas, R.L.; Crim, R.; Hendriks, I.E.; Ramajo, L.; Singh, G.S.; Duarte, C.M.; Gattuso, J.-P. Impacts of Ocean Acidification on Marine Organisms: Quantifying Sensitivities and Interaction with Warming. Glob. Chang. Biol. 2013, 19, 1884–1896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sampaio, E.; Santos, C.; Rosa, I.C.; Ferreira, V.; Pörtner, H.-O.; Duarte, C.M.; Levin, L.A.; Rosa, R. Impacts of Hypoxic Events Surpass Those of Future Ocean Warming and Acidification. Nat. Ecol. Evol. 2021, 5, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Townhill, B.L.; Tinker, J.; Jones, M.; Pitois, S.; Creach, V.; Simpson, S.D.; Dye, S.; Bear, E.; Pinnegar, J.K. Harmful Algal Blooms and Climate Change: Exploring Future Distribution Changes. ICES J. Mar. Sci. 2018, 75, 1882–1893. [Google Scholar] [CrossRef] [Green Version]
- Gobler, C.J. Climate Change and Harmful Algal Blooms: Insights and Perspective. Harmful Algae 2020, 91, 101731. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, J.; González, P.M.; Puntarulo, S. The Phycotoxin Domoic Acid as a Potential Factor for Oxidative Alterations Enhanced by Climate Change. Front. Plant Sci. 2020, 11, 576971. [Google Scholar] [CrossRef]
- Zohdi, E.; Abbaspour, M. Harmful Algal Blooms (Red Tide): A Review of Causes, Impacts and Approaches to Monitoring and Prediction. Int. J. Environ. Sci. Technol. 2019, 16, 1789–1806. [Google Scholar] [CrossRef]
- Sha, J.; Xiong, H.; Li, C.; Lu, Z.; Zhang, J.; Zhong, H.; Zhang, W.; Yan, B. Harmful Algal Blooms and Their Eco-Environmental Indication. Chemosphere 2021, 274, 129912. [Google Scholar] [CrossRef]
- Glibert, P.M. Eutrophication, Harmful Algae and Biodiversity—Challenging Paradigms in a World of Complex Nutrient Changes. Mar. Pollut. Bull. 2017, 124, 591–606. [Google Scholar] [CrossRef]
- Kudela, R.M.; Raine, R.; Pitcher, G.C.; Gentien, P.; Berdalet, E.; Enevoldsen, H.; Urban, E. Establishment, Goals, and Legacy of the Global Ecology and Oceanography of Harmful Algal Blooms (GEOHAB) Programme. In Global Ecology and Oceanography of Harmful Algal Blooms; Glibert, P.M., Berdalet, E., Burford, M.A., Pitcher, G.C., Zhou, M., Eds.; Ecological Studies; Springer International Publishing: Cham, Switzerland, 2018; pp. 27–49. ISBN 978-3-319-70069-4. [Google Scholar]
- Berdalet, E.; Kudela, R.M.; Banas, N.S.; Bresnan, E.; Burford, M.A.; Davidson, K.; Gobler, C.J.; Karlson, B.; Lim, P.T.; Mackenzie, L.; et al. GlobalHAB: Fostering International Coordination on Harmful Algal Bloom Research in Aquatic Systems. In Global Ecology and Oceanography of Harmful Algal Blooms; Springer: Cham, Switzerland, 2018; pp. 425–447. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, S.; Zhang, H.; Lin, S. Omics Study of Harmful Algal Blooms in China: Current Status, Challenges, and Future Perspectives. Harmful Algae 2021, 107, 102079. [Google Scholar] [CrossRef]
- Work, T.M.; Moeller, P.D.R.; Beauchesne, K.R.; Dagenais, J.; Breeden, R.; Rameyer, R.; Walsh, W.J.; Abecassis, M.; Kobayashi, D.R.; Conway, C.; et al. Pufferfish Mortality Associated with Novel Polar Marine Toxins in Hawaii. Dis. Aquat. Org. 2017, 123, 87–99. [Google Scholar] [CrossRef] [Green Version]
- Costa, P.R. Impact and Effects of Paralytic Shellfish Poisoning Toxins Derived from Harmful Algal Blooms to Marine Fish. Fish Fish. 2016, 17, 226–248. [Google Scholar] [CrossRef]
- Broadwater, M.H.; Van Dolah, F.M.; Fire, S.E. Vulnerabilities of Marine Mammals to Harmful Algal Blooms. In Harmful Algal Blooms; John Wiley & Sons, Ltd.: New York, NY, USA, 2018; pp. 191–222. ISBN 978-1-118-99467-2. [Google Scholar]
- Van Hemert, C.; Dusek, R.J.; Smith, M.M.; Kaler, R.; Sheffield, G.; Divine, L.M.; Kuletz, K.J.; Knowles, S.; Lankton, J.S.; Hardison, D.R.; et al. Investigation of Algal Toxins in a Multispecies Seabird Die-Off in the Bering and Chukchi Seas. J. Wildl. Dis. 2020, 57, 399–407. [Google Scholar] [CrossRef]
- Lu, Y.; Yuan, J.; Lu, X.; Su, C.; Zhang, Y.; Wang, C.; Cao, X.; Li, Q.; Su, J.; Ittekkot, V.; et al. Major Threats of Pollution and Climate Change to Global Coastal Ecosystems and Enhanced Management for Sustainability. Environ. Pollut. 2018, 239, 670–680. [Google Scholar] [CrossRef] [Green Version]
- Dickey, R.W.; Plakas, S.M. Ciguatera: A Public Health Perspective. Toxicon 2010, 56, 123–136. [Google Scholar] [CrossRef]
- Masó, M.; Garcés, E. Harmful Microalgae Blooms (HAB); Problematic and Conditions That Induce Them. Mar. Pollut. Bull. 2006, 53, 620–630. [Google Scholar] [CrossRef]
- Lopes, V.M.; Costa, P.R.; Rosa, R. Effects of Harmful Algal Bloom Toxins on Marine Organisms. In Ecotoxicology of Marine Organisms; CRC Press: Boca Raton, FL, USA, 2019; pp. 42–88. ISBN 978-1-315-26749-4. [Google Scholar]
- Wells, M.L.; Karlson, B.; Wulff, A.; Kudela, R.; Trick, C.; Asnaghi, V.; Berdalet, E.; Cochlan, W.; Davidson, K.; De Rijcke, M.; et al. Future HAB Science: Directions and Challenges in a Changing Climate. Harmful Algae 2020, 91, 101632. [Google Scholar] [CrossRef]
- Jeffery, B.; Barlow, T.; Moizer, K.; Paul, S.; Boyle, C. Amnesic Shellfish Poison. Food Chem. Toxicol. 2004, 42, 545–557. [Google Scholar] [CrossRef]
- Costa, P.R.; Garrido, S. Domoic Acid Accumulation in the Sardine Sardina Pilchardus and Its Relationship to Pseudo-Nitzschia Diatom Ingestion. Mar. Ecol. Prog. Ser. 2004, 284, 261–268. [Google Scholar] [CrossRef] [Green Version]
- Costa, P.R.; Rosa, R.; Pereira, J.; Sampayo, M.A.M. Detection of Domoic Acid, the Amnesic Shellfish Toxin, in the Digestive Gland of Eledone Cirrhosa and E. Moschata (Cephalopoda, Octopoda) from the Portuguese Coast. Aquat. Living Resour. 2005, 18, 395–400. [Google Scholar] [CrossRef]
- Hasle, G.R. Are Most of the Domoic Acid-Producing Species of the Diatom Genus Pseudo-Nitzschia Cosmopolites? Harmful Algae 2002, 1, 137–146. [Google Scholar] [CrossRef]
- Trainer, V.L.; Bates, S.S.; Lundholm, N.; Thessen, A.E.; Cochlan, W.P.; Adams, N.G.; Trick, C.G. Pseudo-Nitzschia Physiological Ecology, Phylogeny, Toxicity, Monitoring and Impacts on Ecosystem Health. Harmful Algae 2012, 14, 271–300. [Google Scholar] [CrossRef] [Green Version]
- Doucette, G.J.; King, K.L.; Thessen, A.E.; Dortch, Q. The Effect of Salinity on Domoic Acid Production by the Diatom Pseudo-nitzschia multiseries. Nova Hedwig. 2008, 133, 31–46. [Google Scholar]
- Hallegraeff, G.M. Ocean Climate Change, Phytoplankton Community Responses, and Harmful Algal Blooms: A Formidable Predictive Challenge1. J. Phycol. 2010, 46, 220–235. [Google Scholar] [CrossRef]
- Barton, A.D.; Irwin, A.J.; Finkel, Z.V.; Stock, C.A. Anthropogenic Climate Change Drives Shift and Shuffle in North Atlantic Phytoplankton Communities. Proc. Natl. Acad. Sci. USA 2016, 113, 2964–2969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutkiewicz, S.; Morris, J.J.; Follows, M.J.; Scott, J.; Levitan, O.; Dyhrman, S.T.; Berman-Frank, I. Impact of Ocean Acidification on the Structure of Future Phytoplankton Communities. Nat. Clim Chang. 2015, 5, 1002–1006. [Google Scholar] [CrossRef]
- Gobler, C.J.; Doherty, O.M.; Hattenrath-Lehmann, T.K.; Griffith, A.W.; Kang, Y.; Litaker, R.W. Ocean Warming since 1982 Has Expanded the Niche of Toxic Algal Blooms in the North Atlantic and North Pacific Oceans. Proc. Natl. Acad. Sci. USA 2017, 114, 4975–4980. [Google Scholar] [CrossRef] [Green Version]
- Bresnan, E.; Davidson, K.; Edwards, M.; Fernand, L.; Gowen, R.; Hall, A.; Kennington, K.; McKinney, A.; Milligan, S.; Raine, R. Impacts of Climate Change on Harmful Algal Blooms. MCCIP Sci. Rev. 2013, 236–243. [Google Scholar]
- Edwards, M.; Johns, D.G.; Leterme, S.C.; Svendsen, E.; Richardson, A.J. Regional Climate Change and Harmful Algal Blooms in the Northeast Atlantic. Limnol. Oceanogr. 2006, 51, 820–829. [Google Scholar] [CrossRef] [Green Version]
- Bates, S.S.; Hubbard, K.A.; Lundholm, N.; Montresor, M.; Leaw, C.P. Pseudo-Nitzschia, Nitzschia, and Domoic Acid: New Research since 2011. Harmful Algae 2018, 79, 3–43. [Google Scholar] [CrossRef]
- Trainer, V.L.; Kudela, R.M.; Hunter, M.V.; Adams, N.G.; McCabe, R.M. Climate Extreme Seeds a New Domoic Acid Hotspot on the US West Coast. Front. Clim. 2020, 2, 571836. [Google Scholar] [CrossRef]
- Trainer, V.L.; Moore, S.K.; Hallegraeff, G.; Kudela, R.M.; Clement, A.; Mardones, J.I.; Cochlan, W.P. Pelagic Harmful Algal Blooms and Climate Change: Lessons from Nature’s Experiments with Extremes. Harmful Algae 2020, 91, 101591. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.R.; Moore, S.K.; Tomlinson, M.C.; Silke, J.; Cusack, C.K. Chapter 17—Living with Harmful Algal Blooms in a Changing World: Strategies for Modeling and Mitigating Their Effects in Coastal Marine Ecosystems. In Coastal and Marine Hazards, Risks, and Disasters; Shroder, J.F., Ellis, J.T., Sherman, D.J., Eds.; Hazards and Disasters Series; Elsevier: Boston, MA, USA, 2015; pp. 495–561. ISBN 978-0-12-396483-0. [Google Scholar]
- Ralston, D.K.; Moore, S.K. Modeling Harmful Algal Blooms in a Changing Climate. Harmful Algae 2020, 91, 101729. [Google Scholar] [CrossRef] [PubMed]
- Hannah, L.; Midgley, G.F.; Lovejoy, T.; Bond, W.J.; Bush, M.; Lovett, J.C.; Scott, D.; Woodward, F.I. Conservation of Biodiversity in a Changing Climate. Conserv. Biol. 2002, 16, 264–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, J. Species Distribution Modeling. Geogr. Compass 2010, 4, 490–509. [Google Scholar] [CrossRef]
- Araújo, M.B.; Pearson, R.G.; Rahbek, C. Equilibrium of Species’ Distributions with Climate. Ecography 2005, 28, 693–695. [Google Scholar] [CrossRef]
- Araújo, M.B.; Guisan, A. Five (or so) Challenges for Species Distribution Modelling. J. Biogeogr. 2006, 33, 1677–1688. [Google Scholar] [CrossRef]
- Fitzpatrick, M.C.; Hargrove, W.W. The Projection of Species Distribution Models and the Problem of Non-Analog Climate. Biodivers. Conserv. 2009, 18, 2255. [Google Scholar] [CrossRef]
- Heikkinen, R.K.; Luoto, M.; Araújo, M.B.; Virkkala, R.; Thuiller, W.; Sykes, M.T. Methods and Uncertainties in Bioclimatic Envelope Modelling under Climate Change. Prog. Phys. Geogr. Earth Environ. 2006, 30, 751–777. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel Methods Improve Prediction of Species’ Distributions from Occurrence Data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, N.E.; Edwards, T.C., Jr.; Graham, C.H.; Pearman, P.B.; Svenning, J.-C. New Trends in Species Distribution Modelling. Ecography 2010, 33, 985–989. [Google Scholar] [CrossRef]
- Fabrizio, M.C.; Henderson, M.J.; Rose, K.; Petitgas, P. Editorial: Habitat and Distribution Models of Marine and Estuarine Species: Advances for a Sustainable Future. Front. Mar. Sci. 2022, 9, 1050548. [Google Scholar] [CrossRef]
- Borges, F.O.; Santos, C.P.; Paula, J.R.; Mateos-Naranjo, E.; Redondo-Gomez, S.; Adams, J.B.; Caçador, I.; Fonseca, V.F.; Reis-Santos, P.; Duarte, B.; et al. Invasion and Extirpation Potential of Native and Invasive Spartina Species Under Climate Change. Front. Mar. Sci. 2021, 8, 696333. [Google Scholar] [CrossRef]
- Sinclair, S.; White, M.; Newell, G. How Useful Are Species Distribution Models for Managing Biodiversity under Future Climates? Ecol. Soc. 2010, 15, 8. [Google Scholar] [CrossRef]
- Boavida-Portugal, J.; Rosa, R.; Calado, R.; Pinto, M.; Boavida-Portugal, I.; Araújo, M.B.; Guilhaumon, F. Climate Change Impacts on the Distribution of Coastal Lobsters. Mar. Biol. 2018, 165, 186. [Google Scholar] [CrossRef]
- Borges, F.O.; Guerreiro, M.; Santos, C.P.; Paula, J.R.; Rosa, R. Projecting Future Climate Change Impacts on the Distribution of the “Octopus Vulgaris Species Complex”. Front. Mar. Sci. 2022, 9, 2462. [Google Scholar] [CrossRef]
- Anderson, C.R.; Berdalet, E.; Kudela, R.M.; Cusack, C.K.; Silke, J.; O’Rourke, E.; Dugan, D.; McCammon, M.; Newton, J.A.; Moore, S.K.; et al. Scaling Up from Regional Case Studies to a Global Harmful Algal Bloom Observing System. Front. Mar. Sci. 2019, 6, 250. [Google Scholar] [CrossRef]
- Borges, F.O.; Lopes, V.M.; Amorim, A.; Santos, C.F.; Costa, P.R.; Rosa, R. Projecting Future Climate Change-Mediated Impacts in Three Paralytic Shellfish Toxins-Producing Dinoflagellate Species. Biology 2022, 11, 1424. [Google Scholar] [CrossRef]
- Palma, S.; Mouriño, H.; Silva, A.; Barão, M.I.; Moita, M.T. Can Pseudo-Nitzschia Blooms Be Modeled by Coastal Upwelling in Lisbon Bay? Harmful Algae 2010, 9, 294–303. [Google Scholar] [CrossRef]
- Schnetzer, A.; Jones, B.H.; Schaffner, R.A.; Cetinic, I.; Fitzpatrick, E.; Miller, P.E.; Seubert, E.L.; Caron, D.A. Coastal Upwelling Linked to Toxic Pseudo-Nitzschia Australis Blooms in Los Angeles Coastal Waters, 2005–2007. J. Plankton Res. 2013, 35, 1080–1092. [Google Scholar] [CrossRef] [Green Version]
- Torres Palenzuela, J.M.; González Vilas, L.; Bellas, F.M.; Garet, E.; González-Fernández, Á.; Spyrakos, E. Pseudo-Nitzschia Blooms in a Coastal Upwelling System: Remote Sensing Detection, Toxicity and Environmental Variables. Water 2019, 11, 1954. [Google Scholar] [CrossRef] [Green Version]
- Sorte, C.J.B.; Williams, S.L.; Carlton, J.T. Marine Range Shifts and Species Introductions: Comparative Spread Rates and Community Impacts. Glob. Ecol. Biogeogr. 2010, 19, 303–316. [Google Scholar] [CrossRef]
- Hastings, R.A.; Rutterford, L.A.; Freer, J.J.; Collins, R.A.; Simpson, S.D.; Genner, M.J. Climate Change Drives Poleward Increases and Equatorward Declines in Marine Species. Curr. Biol. 2020, 30, 1572–1577.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, M.K.; Kremer, C.T.; Klausmeier, C.A.; Litchman, E. A Global Pattern of Thermal Adaptation in Marine Phytoplankton. Science 2012, 338, 1085–1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Continuous Plankton Recorder Survey Team. Continuous Plankton Records: Plankton Atlas of the North Atlantic Ocean (1958–1999). II. Biogeographical Charts. Mar. Ecol. Prog. Ser. 2004, 11–75. [Google Scholar]
- Chivers, W.J.; Walne, A.W.; Hays, G.C. Mismatch between Marine Plankton Range Movements and the Velocity of Climate Change. Nat. Commun. 2017, 8, 14434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldrich, Á.M.; Molinet, C.; Reguera, B.; Espinoza-González, O.; Pizarro, G.; Rodríguez-Villegas, C.; Opazo, D.; Mejías, P.; Díaz, P.A. Interannual Variability in Mesoscale Distribution of Dinophysis Acuminata and D. Acuta in Northwestern Patagonian Fjords. Harmful Algae 2022, 115, 102228. [Google Scholar] [CrossRef]
- Oziel, L.; Baudena, A.; Ardyna, M.; Massicotte, P.; Randelhoff, A.; Sallée, J.-B.; Ingvaldsen, R.B.; Devred, E.; Babin, M. Faster Atlantic Currents Drive Poleward Expansion of Temperate Phytoplankton in the Arctic Ocean. Nat. Commun. 2020, 11, 1705. [Google Scholar] [CrossRef] [Green Version]
- Hennon, G.M.M.; Dyhrman, S.T. Progress and Promise of Omics for Predicting the Impacts of Climate Change on Harmful Algal Blooms. Harmful Algae 2020, 91, 101587. [Google Scholar] [CrossRef]
- Chen, B.; Laws, E.A. Is There a Difference of Temperature Sensitivity between Marine Phytoplankton and Heterotrophs? Limnol. Oceanogr. 2017, 62, 806–817. [Google Scholar] [CrossRef]
- Jin, P.; Agustí, S. Fast Adaptation of Tropical Diatoms to Increased Warming with Trade-Offs. Sci. Rep. 2018, 8, 17771. [Google Scholar] [CrossRef] [Green Version]
- Moore, S.K.; Mantua, N.J.; Salathé, E.P. Past Trends and Future Scenarios for Environmental Conditions Favoring the Accumulation of Paralytic Shellfish Toxins in Puget Sound Shellfish. Harmful Algae 2011, 10, 521–529. [Google Scholar] [CrossRef]
- Casabianca, S.; Penna, A.; Pecchioli, E.; Jordi, A.; Basterretxea, G.; Vernesi, C. Population Genetic Structure and Connectivity of the Harmful Dinoflagellate Alexandrium Minutum in the Mediterranean Sea. Proc. R. Soc. B Biol. Sci. 2012, 279, 129–138. [Google Scholar] [CrossRef]
- Beck, J.; Böller, M.; Erhardt, A.; Schwanghart, W. Spatial Bias in the GBIF Database and Its Effect on Modeling Species’ Geographic Distributions. Ecol. Inform. 2014, 19, 10–15. [Google Scholar] [CrossRef]
- Lürling, M. Grazing Resistance in Phytoplankton. Hydrobiologia 2021, 848, 237–249. [Google Scholar] [CrossRef]
- Harðardóttir, S.; Pančić, M.; Tammilehto, A.; Krock, B.; Møller, E.F.; Nielsen, T.G.; Lundholm, N. Dangerous Relations in the Arctic Marine Food Web: Interactions between Toxin Producing Pseudo-Nitzschia Diatoms and Calanus Copepodites. Mar. Drugs 2015, 13, 3809–3835. [Google Scholar] [CrossRef] [Green Version]
- Woods, A. Domoic Acid Production in Pseudo-Nitzschia (Bacillariophyceae) as a General Response to Unbalanced Growth: The Role of Photo-Oxidative Stress. Capstone Projects and Master’s Theses, California State University, Monterey Bay, Seaside, CA, USA, 2016. [Google Scholar]
- Trainor, A.M.; Schmitz, O.J. Infusing Considerations of Trophic Dependencies into Species Distribution Modelling. Ecol. Lett. 2014, 17, 1507–1517. [Google Scholar] [CrossRef]
- Trainor, A.M.; Schmitz, O.J.; Ivan, J.S.; Shenk, T.M. Enhancing Species Distribution Modeling by Characterizing Predator-Prey Interactions. Ecol. Appl. 2014, 24, 204–216. [Google Scholar] [CrossRef]
- Glibert, P.M.; Icarus Allen, J.; Artioli, Y.; Beusen, A.; Bouwman, L.; Harle, J.; Holmes, R.; Holt, J. Vulnerability of Coastal Ecosystems to Changes in Harmful Algal Bloom Distribution in Response to Climate Change: Projections Based on Model Analysis. Glob. Chang. Biol. 2014, 20, 3845–3858. [Google Scholar] [CrossRef] [Green Version]
- Telenius, A. Biodiversity Information Goes Public: GBIF at Your Service. Nord. J. Bot. 2011, 29, 378–381. [Google Scholar] [CrossRef]
- Shipley, B.R.; Bach, R.; Do, Y.; Strathearn, H.; McGuire, J.L.; Dilkina, B. MegaSDM: Integrating Dispersal and Time-Step Analyses into Species Distribution Models. Ecography 2022, 2022, e05450. [Google Scholar] [CrossRef]
- RStudio Team. RStudio: Integrated Development for R; RStudio, Inc.: Boston, MA, USA, 2022. [Google Scholar]
- Laruelle, G.G.; Cai, W.-J.; Hu, X.; Gruber, N.; Mackenzie, F.T.; Regnier, P. Continental Shelves as a Variable but Increasing Global Sink for Atmospheric Carbon Dioxide. Nat. Commun. 2018, 9, 454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sbrocco, E.J.; Barber, P.H. MARSPEC: Ocean Climate Layers for Marine Spatial Ecology. Ecology 2013, 94, 979. [Google Scholar] [CrossRef]
- IPCC. IPCC, 2014: Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Core Writing Team, Pachauri, R.K., Meyer, L.A., Eds.; IPCC: Geneva, Switzerland, 2014; p. 151.
- Schwalm, C.R.; Glendon, S.; Duffy, P.B. RCP8.5 Tracks Cumulative CO2 Emissions. Proc. Natl. Acad. Sci. USA 2020, 117, 19656–19657. [Google Scholar] [CrossRef] [PubMed]
- Tyberghein, L.; Verbruggen, H.; Pauly, K.; Troupin, C.; Mineur, F.; De Clerck, O. Bio-ORACLE: A Global Environmental Dataset for Marine Species Distribution Modelling. Glob. Ecol. Biogeogr. 2012, 21, 272–281. [Google Scholar] [CrossRef]
- Assis, J.; Tyberghein, L.; Bosch, S.; Verbruggen, H.; Serrão, E.A.; De Clerck, O. Bio-ORACLE v2.0: Extending Marine Data Layers for Bioclimatic Modelling. Glob. Ecol. Biogeogr. 2018, 27, 277–284. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A Statistical Explanation of MaxEnt for Ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Elith, J.; Graham, C.H.; Lehmann, A.; Leathwick, J.; Ferrier, S. Sample Selection Bias and Presence-Only Distribution Models: Implications for Background and Pseudo-Absence Data. Ecol. Appl. 2009, 19, 181–197. [Google Scholar] [CrossRef] [Green Version]
- Boakes, E.H.; McGowan, P.J.K.; Fuller, R.A.; Chang-qing, D.; Clark, N.E.; O’Connor, K.; Mace, G.M. Distorted Views of Biodiversity: Spatial and Temporal Bias in Species Occurrence Data. PLoS Biol. 2010, 8, e1000385. [Google Scholar] [CrossRef]
- Beck, J.; Ballesteros-Mejia, L.; Nagel, P.; Kitching, I.J. Online Solutions and the ‘Wallacean Shortfall’: What Does GBIF Contribute to Our Knowledge of Species’ Ranges? Divers. Distrib. 2013, 19, 1043–1050. [Google Scholar] [CrossRef]
- Varela, S.; Anderson, R.P.; García-Valdés, R.; Fernández-González, F. Environmental Filters Reduce the Effects of Sampling Bias and Improve Predictions of Ecological Niche Models. Ecography 2014, 37, 1084–1091. [Google Scholar] [CrossRef]
- Peterson, A.; Soberón, J. Species Distribution Modeling and Ecological Niche Modeling: Getting the Concepts Right. Nat. Conserv. 2012, 10, 1–6. [Google Scholar] [CrossRef]
- Barbet-Massin, M.; Jiguet, F.; Albert, C.H.; Thuiller, W. Selecting Pseudo-Absences for Species Distribution Models: How, Where and How Many? Methods Ecol. Evol. 2012, 3, 327–338. [Google Scholar] [CrossRef]
- Lobo, J.M.; Jiménez-Valverde, A.; Hortal, J. The Uncertain Nature of Absences and Their Importance in Species Distribution Modelling. Ecography 2010, 33, 103–114. [Google Scholar] [CrossRef]
- Kramer-Schadt, S.; Niedballa, J.; Pilgrim, J.D.; Schröder, B.; Lindenborn, J.; Reinfelder, V.; Stillfried, M.; Heckmann, I.; Scharf, A.K.; Augeri, D.M.; et al. The Importance of Correcting for Sampling Bias in MaxEnt Species Distribution Models. Divers. Distrib. 2013, 19, 1366–1379. [Google Scholar] [CrossRef]
- Radosavljevic, A.; Anderson, R.P. Making Better Maxent Models of Species Distributions: Complexity, Overfitting and Evaluation. J. Biogeogr. 2014, 41, 629–643. [Google Scholar] [CrossRef]
- Elith, J.; Kearney, M.; Phillips, S. The Art of Modelling Range-Shifting Species. Methods Ecol. Evol. 2010, 1, 330–342. [Google Scholar] [CrossRef]
- Raes, N.; ter Steege, H. A Null-Model for Significance Testing of Presence-Only Species Distribution Models. Ecography 2007, 30, 727–736. [Google Scholar] [CrossRef]
- Araújo, M.B.; New, M. Ensemble Forecasting of Species Distributions. Trends Ecol. Evol. 2007, 22, 42–47. [Google Scholar] [CrossRef]
- Liu, C.; Berry, P.M.; Dawson, T.P.; Pearson, R.G. Selecting Thresholds of Occurrence in the Prediction of Species Distributions. Ecography 2005, 28, 385–393. [Google Scholar] [CrossRef]
- Early, R.; Sax, D.F. Analysis of Climate Paths Reveals Potential Limitations on Species Range Shifts. Ecol. Lett. 2011, 14, 1125–1133. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | #1 | #2 | #3 | #4 |
|---|---|---|---|---|
| P. australis | Bathymetry 71.4% | Temperature Maximum 9.7% | Temperature Range 5.4% | Temperature Minimum 3.4% |
| P. seriata | Bathymetry 81.9% | Temperature Minimum 9.5% | Temperature Range 4.7% | Temperature Mean 0.9% |
| P. fraudulenta | Bathymetry 61.9% | Salinity Minimum 15.2% | Temperature Mean 5.4% | Temperature Maximum 4.6% |
| Species | Accurate prediction | Overprediction | Underprediction |
|---|---|---|---|
| P. australis | W Atlantic (Gulf of Mexico, Argentina, Gulf of Maine, South Brazil) | Southern Ocean | Bay of Fundy Gulf of Alaska Bering Strait |
| N Atlantic (Celt Sea, Ireland, Scotland, Galicia, Portugal, Morocco) | Arctic Ocean | ||
| SE Atlantic (Benguela, Namibia) | Indonesia | ||
| Eastern Pacific (Mexico, Baja California, USA, Alaska, Canada, Peru, Chile) | Indian Ocean (Oman) | ||
| Bering Strait and Bering Sea | NW Pacific (Okhotsk Sea, Sea of Japan, Eastern China Sea) | ||
| Oceania (Australia, New Zealand) | Mediterranean Sea | ||
| Atlantic (northern South America) | |||
| P. seriata | NW Pacific (Okhotsk Sea, Sea of Japan, Bering Sea) | Southern Hemisphere | Singapore |
| NW Atlantic (Gulf of St. Lawrence, Greenland) | |||
| Gulf of Mexico | |||
| Arctic Ocean | |||
| Black Sea | |||
| P. fraudulenta | NW Atlantic (USA, Canada) | Madagascar Indonesia SE Pacific Red Sea Arctic Ocean | Marmara Sea (Turkey) |
| NE Atlantic (Morocco, Celt Sea, Scotland) | |||
| North Sea | |||
| Mediterranean Sea (NW Mediterranean, Adriatic, Morocco) | |||
| Gulf of Mexico | |||
| SW Atlantic (South Brazil, Argentina) | |||
| NE Pacific (Washington, Gulf of California, Baja California, Mexico, Chile) | |||
| NW Pacific (Okhotsk Sea, Sea of Japan) | |||
| Oceania (Australia, New Zealand) | |||
| Indian Ocean (Pakistan) |
| Species | Precuration | Curated | Post-Environmental Filtering |
|---|---|---|---|
| Pseudo-nitzschia australisA | 57 | 32 | 30 |
| Pseudo-nitzschia seriataB | 1997 | 834 | 782 |
| Pseudo-nitzschia fraudulentaC | 162 | 129 | 124 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borges, F.O.; Lopes, V.M.; Santos, C.F.; Costa, P.R.; Rosa, R. Impacts of Climate Change on the Biogeography of Three Amnesic Shellfish Toxin Producing Diatom Species. Toxins 2023, 15, 9. https://doi.org/10.3390/toxins15010009
Borges FO, Lopes VM, Santos CF, Costa PR, Rosa R. Impacts of Climate Change on the Biogeography of Three Amnesic Shellfish Toxin Producing Diatom Species. Toxins. 2023; 15(1):9. https://doi.org/10.3390/toxins15010009
Chicago/Turabian StyleBorges, Francisco O., Vanessa M. Lopes, Catarina Frazão Santos, Pedro Reis Costa, and Rui Rosa. 2023. "Impacts of Climate Change on the Biogeography of Three Amnesic Shellfish Toxin Producing Diatom Species" Toxins 15, no. 1: 9. https://doi.org/10.3390/toxins15010009
APA StyleBorges, F. O., Lopes, V. M., Santos, C. F., Costa, P. R., & Rosa, R. (2023). Impacts of Climate Change on the Biogeography of Three Amnesic Shellfish Toxin Producing Diatom Species. Toxins, 15(1), 9. https://doi.org/10.3390/toxins15010009