Characteristics of Shiga Toxin-Producing Escherichia coli Circulating in Asymptomatic Food Handlers


 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Occurrence of STEC in Asymptomatic Food Handlers
2.2. Molecular Characteristics of Asymptomatic Food Handlers-Derived STEC Isolates
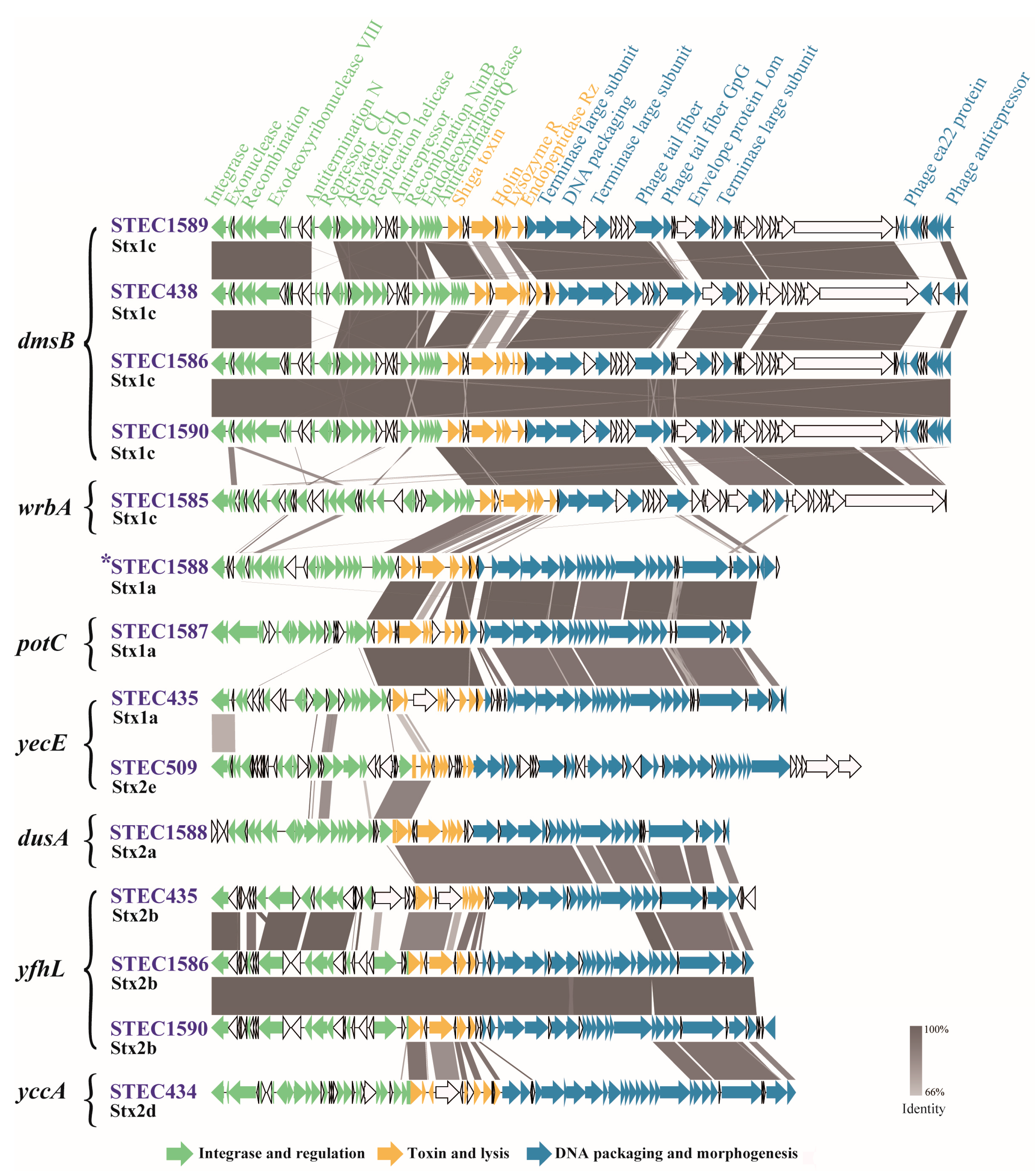
2.3. Genetic Features of Stx Prophages
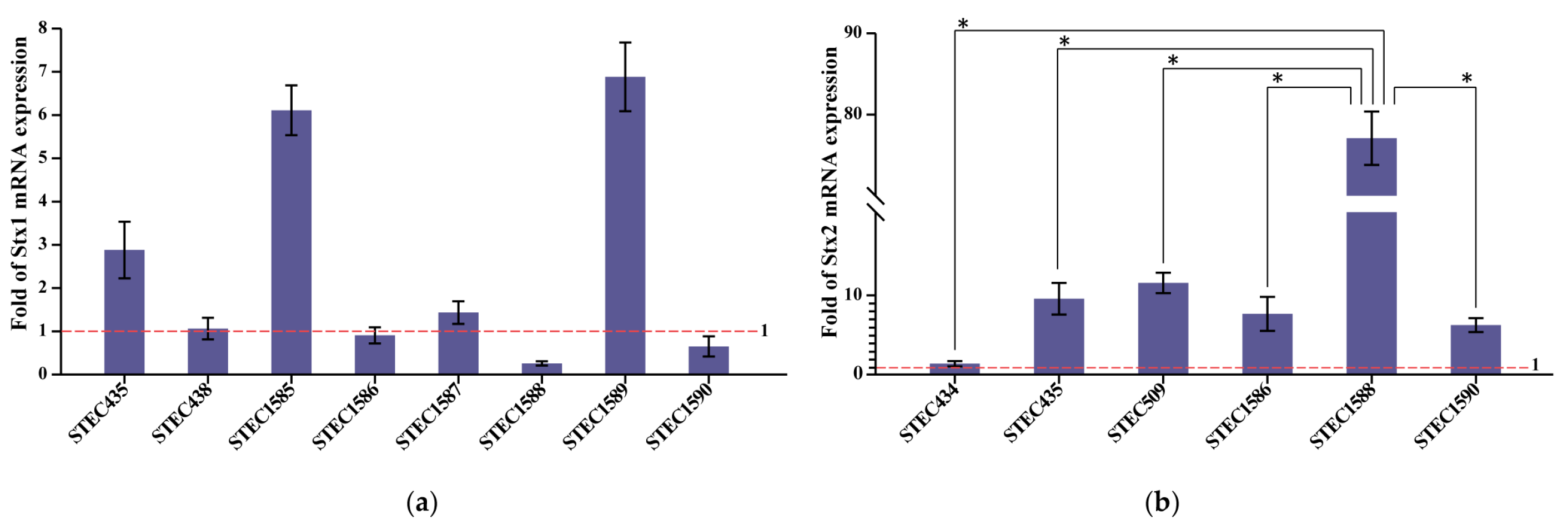
2.4. Variable Stx mRNA Expression Levels
2.5. Antimicrobial Resistance (AMR) and AMR Genes
2.6. Phylogenetic Relationships of STEC Strains from This Study and Other Sources
3. Discussion
4. Materials and Methods
4.1. Sample Collection and Bacterial Strains
4.2. Whole Genome Sequencing (WGS) and Assembling
4.3. WGS-Based Molecular Characterization
4.4. Genomic Characterization of Stx-Converting Prophages
4.5. RNA Extraction and Relative Quantification of Stx Expression
4.6. Antimicrobial Susceptibility Test
4.7. Phylogenomic Analysis
4.8. Statistical Analysis
4.9. Data Availability
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Smith, J.L.; Fratamico, P.M.; Gunther, N.W.t. Shiga toxin-producing Escherichia coli. Adv. Appl. Microbiol. 2014, 86, 145–197. [Google Scholar]
- Jones, G.; Mariani-Kurkdjian, P.; Cointe, A.; Bonacorsi, S.; Lefevre, S.; Weill, F.X.; Le Strat, Y. Sporadic Shiga toxin-producing Escherichia coli-associated pediatric Hemolytic Uremic Syndrome, France, 2012–2021. Emerg. Infect. Dis. 2023, 29, 2054–2064. [Google Scholar] [CrossRef]
- Scheutz, F.; Teel, L.D.; Beutin, L.; Pierard, D.; Buvens, G.; Karch, H.; Mellmann, A.; Caprioli, A.; Tozzoli, R.; Morabito, S.; et al. Multicenter evaluation of a sequence-based protocol for subtyping Shiga toxins and standardizing Stx nomenclature. J. Clin. Microbiol. 2012, 50, 2951–2963. [Google Scholar] [CrossRef]
- Koutsoumanis, K.; Allende, A.; Alvarez-Ordonez, A.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Herman, L.; Hilbert, F.; Lindqvist, R.; et al. Pathogenicity assessment of Shiga toxin-producing Escherichia coli (STEC) and the public health risk posed by contamination of food with STEC. Efsa J. 2020, 18, e05967. [Google Scholar]
- Bai, X.; Fu, S.; Zhang, J.; Fan, R.; Xu, Y.; Sun, H.; He, X.; Xu, J.; Xiong, Y. Identification and pathogenomic analysis of an Escherichia coli strain producing a novel Shiga toxin 2 subtype. Sci. Rep. 2018, 8, 6756. [Google Scholar] [CrossRef]
- Lacher, D.W.; Gangiredla, J.; Patel, I.; Elkins, C.A.; Feng, P.C. Use of the Escherichia coli Identification microarray for characterizing the health risks of Shiga toxin-producing Escherichia coli isolated from foods. J. Food Prot. 2016, 79, 1656–1662. [Google Scholar] [CrossRef]
- Yang, X.; Bai, X.; Zhang, J.; Sun, H.; Fu, S.; Fan, R.; He, X.; Scheutz, F.; Matussek, A.; Xiong, Y. Escherichia coli strains producing a novel Shiga toxin 2 subtype circulate in China. Int. J. Med. Microbiol. 2020, 310, 151377. [Google Scholar] [CrossRef]
- Gill, A.; Dussault, F.; McMahon, T.; Petronella, N.; Wang, X.; Cebelinski, E.; Scheutz, F.; Weedmark, K.; Blais, B.; Carrillo, C. Characterization of atypical Shiga toxin gene sequences and description of Stx2j, a new subtype. J. Clin. Microbiol. 2022, 60, e0222921. [Google Scholar] [CrossRef]
- Bai, X.N.; Scheutz, F.; Dahlgren, H.M.; Hedenstrom, I.; Jernberg, C. Characterization of clinical Escherichia coli strains producing a novel Shiga toxin 2 subtype in Sweden and Denmark. Microorganisms 2021, 9, 2374. [Google Scholar] [CrossRef]
- Murphy, K.C.; Ritchie, J.M.; Waldor, M.K.; Lobner-Olesen, A.; Marinus, M.G. Dam methyltransferase is required for stable lysogeny of the Shiga toxin (Stx2)-encoding bacteriophage 933W of enterohemorrhagic Escherichia coli O157:H7. J. Bacteriol. 2008, 190, 438–441. [Google Scholar] [CrossRef]
- Berger, P.; Kouzel, I.U.; Berger, M.; Haarmann, N.; Dobrindt, U.; Koudelka, G.B.; Mellmann, A. Carriage of Shiga toxin phage profoundly affects Escherichia coli gene expression and carbon source utilization. BMC Genom. 2019, 20, 504. [Google Scholar] [CrossRef]
- Bai, X.; Zhang, J.; Ambikan, A.; Jernberg, C.; Ehricht, R.; Scheutz, F.; Xiong, Y.; Matussek, A. Molecular characterization and comparative genomics of clinical hybrid Shiga toxin-producing and enterotoxigenic Escherichia coli (STEC/ETEC) strains in Sweden. Sci. Rep. 2019, 9, 5619. [Google Scholar] [CrossRef]
- Hazen, T.H.; Michalski, J.; Luo, Q.; Shetty, A.C.; Daugherty, S.C.; Fleckenstein, J.M.; Rasko, D.A. Comparative genomics and transcriptomics of Escherichia coli isolates carrying virulence factors of both enteropathogenic and enterotoxigenic E. coli. Sci. Rep. 2017, 7, 3513. [Google Scholar] [CrossRef]
- Rodriguez-Rubio, L.; Haarmann, N.; Schwidder, M.; Muniesa, M.; Schmidt, H. Bacteriophages of Shiga toxin-producing Escherichia coli and their contribution to pathogenicity. Pathogens 2021, 10, 404. [Google Scholar] [CrossRef]
- Kagambega, A.; Martikainen, O.; Siitonen, A.; Traore, A.S.; Barro, N.; Haukka, K. Prevalence of diarrheagenic Escherichia coli virulence genes in the feces of slaughtered cattle, chickens, and pigs in Burkina Faso. Microbiologyopen 2012, 1, 276–284. [Google Scholar] [CrossRef]
- Mercer, R.G.; Zheng, J.; Garcia-Hernandez, R.; Ruan, L.; Ganzle, M.G.; McMullen, L.M. Genetic determinants of heat resistance in Escherichia coli. Front. Microbiol. 2015, 6, 932. [Google Scholar] [CrossRef]
- Meng, Q.; Bai, X.; Zhao, A.; Lan, R.; Du, H.; Wang, T.; Shi, C.; Yuan, X.; Bai, X.; Ji, S.; et al. Characterization of Shiga toxin-producing Escherichia coli isolated from healthy pigs in China. BMC Microbiol. 2014, 14, 5. [Google Scholar] [CrossRef]
- Guh, A.; Phan, Q.; Nelson, R.; Purviance, K.; Milardo, E.; Kinney, S.; Mshar, P.; Kasacek, W.; Cartter, M. Outbreak of Escherichia coli O157 associated with raw milk, Connecticut, 2008. Clin. Infect. Dis. 2010, 51, 1411–1417. [Google Scholar] [CrossRef]
- Sethulekshmi, C.; Latha, C.; Anu, C.J. Occurrence and quantification of Shiga toxin-producing Escherichia coli from food matrices. Vet. World 2018, 11, 104–111. [Google Scholar] [CrossRef]
- Treacy, J.; Jenkins, C.; Paranthaman, K.; Jorgensen, F.; Mueller-Doblies, D.; Anjum, M.; Kaindama, L.; Hartman, H.; Kirchner, M.; Carson, T.; et al. Outbreak of Shiga toxin-producing Escherichia coli O157:H7 linked to raw drinking milk resolved by rapid application of advanced pathogen characterisation methods, England, August to October 2017. Eurosurveillance 2019, 24, 1800191. [Google Scholar] [CrossRef]
- Carlson-Banning, K.M.; Sperandio, V. Catabolite and oxygen regulation of enterohemorrhagic Escherichia coli virulence. mBio 2016, 7, e01852-16. [Google Scholar] [CrossRef] [PubMed]
- Kintz, E.; Brainard, J.; Vanderes, M.; Vivancos, R.; Byrne, L.; Butt, S.; Jenkins, C.; Elson, R.; Lake, I.; Hunter, P. Animal and environmental risk factors for sporadic Shiga toxin-producing Escherichia coli (STEC) infection in England: A case control study for O157, O26 and other STEC serotypes. Pathog. Glob. Health 2023, 117, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chen, X.; Zheng, S.; Yu, F.; Kong, H.; Yang, Q.; Cui, D.; Chen, N.; Lou, B.; Li, X.; et al. Serotypes, genotypes and antimicrobial resistance patterns of human diarrhoeagenic Escherichia coli isolates circulating in southeastern China. Clin. Microbiol. Infect. 2014, 20, 52–58. [Google Scholar] [CrossRef]
- Nuesch-Inderbinen, M.; Treier, A.; Stevens, M.J.A.; Stephan, R. Whole genome sequence-based characterisation of Shiga toxin-producing Escherichia coli isolated from game meat originating from several European countries. Sci. Rep. 2023, 13, 3247. [Google Scholar] [CrossRef]
- Riley, L.W.; Remis, R.S.; Helgerson, S.D.; McGee, H.B.; Wells, J.G.; Davis, B.R.; Hebert, R.J.; Olcott, E.S.; Johnson, L.M.; Hargrett, N.T.; et al. Hemorrhagic colitis associated with a rare Escherichia coli serotype. N. Engl. J. Med. 1983, 308, 681–685. [Google Scholar] [CrossRef] [PubMed]
- Grant, J.; Wendelboe, A.M.; Wendel, A.; Jepson, B.; Torres, P.; Smelser, C.; Rolfs, R.T. Spinach-associated Escherichia coli O157:H7 outbreak, Utah and New Mexico, 2006. Emerg. Infect. Dis. 2008, 14, 1633–1636. [Google Scholar] [CrossRef] [PubMed]
- Michino, H.; Araki, K.; Minami, S.; Takaya, S.; Sakai, N.; Miyazaki, M.; Ono, A.; Yanagawa, H. Massive outbreak of Escherichia coli O157:H7 infection in schoolchildren in Sakai City, Japan, associated with consumption of white radish sprouts. Am. J. Epidemiol. 1999, 150, 787–796. [Google Scholar] [CrossRef]
- Frank, C.; Werber, D.; Cramer, J.P.; Askar, M.; Faber, M.; an der Heiden, M.; Bernard, H.; Fruth, A.; Prager, R.; Spode, A.; et al. Epidemic profile of Shiga-toxin-producing Escherichia coli O104:H4 outbreak in Germany. N. Engl. J. Med. 2011, 365, 1771–1780. [Google Scholar] [CrossRef]
- Valilis, E.; Ramsey, A.; Sidiq, S.; DuPont, H.L. Non-O157 Shiga toxin-producing Escherichia coli-A poorly appreciated enteric pathogen: Systematic review. Int. J. Infect. Dis. 2018, 76, 82–87. [Google Scholar] [CrossRef]
- Xiong, Y.; Wang, P.; Lan, R.; Ye, C.; Wang, H.; Ren, J.; Jing, H.; Wang, Y.; Zhou, Z.; Bai, X.; et al. A novel Escherichia coli O157:H7 clone causing a major hemolytic uremic syndrome outbreak in China. PLoS ONE 2012, 7, e36144. [Google Scholar] [CrossRef]
- Zhu, W.; Guo, H.; Xu, J.; Wu, W.; Yi, Y.; Wang, J.; Duan, R.; Tong, J.; Du, Y. Enterohemorrhagic Escherichia coli O157:H7—Xuzhou City, Jiangsu Province, China, 2001–2021. China CDC Wkly. 2023, 5, 311–314. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Wang, Y.; Wang, C.; Tang, Y.; He, L.; Deng, J.; Pan, X. Complete genome sequences of two Shiga toxin-producing Escherichia coli O146:H10 strains recovered from a foodborne outbreak in China. Microbiol. Resour. Announc. 2021, 10, e0082521. [Google Scholar] [CrossRef] [PubMed]
- Balabanova, Y.; Gilsdorf, A.; Buda, S.; Burger, R.; Eckmanns, T.; Gartner, B.; Gross, U.; Haas, W.; Hamouda, O.; Hubner, J.; et al. Communicable diseases prioritized for surveillance and epidemiological research: Results of a standardized prioritization procedure in Germany, 2011. PLoS ONE 2011, 6, e25691. [Google Scholar] [CrossRef] [PubMed]
- Baba, H.; Kanamori, H.; Kudo, H.; Kuroki, Y.; Higashi, S.; Oka, K.; Takahashi, M.; Yoshida, M.; Oshima, K.; Aoyagi, T.; et al. Genomic analysis of Shiga toxin-producing Escherichia coli from patients and asymptomatic food handlers in Japan. PLoS ONE 2019, 14, e0225340. [Google Scholar] [CrossRef]
- Bizot, E.; Cointe, A.; Bechet, S.; Sobral, E.; Cohen, R.; Mariani-Kurkdjian, P.; Levy, C.; Bonacorsi, S. Shiga toxin-producing Escherichia coli carriage in 959 healthy French infants. Arch. Dis. Child. 2021, 106, 1239–1240. [Google Scholar] [CrossRef] [PubMed]
- Harries, M.; Dreesman, J.; Rettenbacher-Riefler, S.; Mertens, E. Faecal carriage of extended-spectrum beta-lactamase-producing Enterobacteriaceae and Shiga toxin-producing Escherichia coli in asymptomatic nursery children in Lower Saxony (Germany), 2014. Epidemiol. Infect. 2016, 144, 3540–3548. [Google Scholar] [CrossRef] [PubMed]
- Morita-Ishihara, T.; Iyoda, S.; Iguchi, A.; Ohnishi, M. Secondary Shiga toxin-producing Escherichia coli infection, Japan, 2010-2012. Emerg. Infect. Dis. 2016, 22, 2181–2184. [Google Scholar] [CrossRef]
- Quiros, P.; Martinez-Castillo, A.; Muniesa, M. Improving detection of Shiga toxin-producing Escherichia coli by molecular methods by reducing the interference of free Shiga toxin-encoding bacteriophages. Appl. Environ. Microbiol. 2015, 81, 415–421. [Google Scholar] [CrossRef]
- Gill, A.; Huszczynski, G.; Gauthier, M.; Blais, B. Evaluation of eight agar media for the isolation of Shiga toxin-producing Escherichia coli. J. Microbiol. Methods 2014, 96, 6–11. [Google Scholar] [CrossRef]
- Brooks, J.T.; Sowers, E.G.; Wells, J.G.; Greene, K.D.; Griffin, P.M.; Hoekstra, R.M.; Strockbine, N.A. Non-O157 Shiga toxin-producing Escherichia coli infections in the United States, 1983–2002. J. Infect. Dis. 2005, 192, 1422–1429. [Google Scholar] [CrossRef]
- Bai, X.; Hu, B.; Xu, Y.; Sun, H.; Zhao, A.; Ba, P.; Fu, S.; Fan, R.; Jin, Y.; Wang, H.; et al. Molecular and phylogenetic characterization of non-O157 Shiga toxin-producing Escherichia coli strains in China. Front. Cell Infect. Microbiol. 2016, 6, 143. [Google Scholar] [CrossRef] [PubMed]
- Mahanti, A.; Samanta, I.; Bandyopadhyay, S.; Joardar, S.N. Molecular characterization and antibiotic susceptibility pattern of caprine Shiga toxin producing-Escherichia coli (STEC) isolates from India. Iran. J. Vet. Res. 2015, 16, 31–35. [Google Scholar] [PubMed]
- Franci, T.; Sanso, A.M.; Bustamante, A.V.; Lucchesi, P.M.; Parma, A.E. Genetic characterization of non-O157 verocytotoxigenic Escherichia coli isolated from raw beef products using multiple-locus variable-number tandem repeat analysis. Foodborne Pathog. Dis. 2011, 8, 1019–1023. [Google Scholar] [CrossRef]
- Meichtri, L.; Miliwebsky, E.; Gioffre, A.; Chinen, I.; Baschkier, A.; Chillemi, G.; Guth, B.E.; Masana, M.O.; Cataldi, A.; Rodriguez, H.R.; et al. Shiga toxin-producing Escherichia coli in healthy young beef steers from Argentina: Prevalence and virulence properties. Int. J. Food Microbiol. 2004, 96, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Polifroni, R.; Etcheverria, A.I.; Sanz, M.E.; Cepeda, R.E.; Kruger, A.; Lucchesi, P.M.; Fernandez, D.; Parma, A.E.; Padola, N.L. Molecular characterization of Shiga toxin-producing Escherichia coli isolated from the environment of a dairy farm. Curr. Microbiol. 2012, 65, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Melton-Celsa, A.; Mohawk, K.; Teel, L.; O’Brien, A. Pathogenesis of Shiga-toxin producing Escherichia coli. Curr. Top. Microbiol. Immunol. 2012, 357, 67–103. [Google Scholar] [PubMed]
- Baranzoni, G.M.; Fratamico, P.M.; Gangiredla, J.; Patel, I.; Bagi, L.K.; Delannoy, S.; Fach, P.; Boccia, F.; Anastasio, A.; Pepe, T. Characterization of Shiga toxin subtypes and virulence genes in porcine Shiga toxin-producing Escherichia coli. Front. Microbiol. 2016, 7, 574. [Google Scholar] [CrossRef]
- Melton-Celsa, A.R. Shiga toxin (Stx) classification, structure, and function. Microbiol. Spectr. 2014, 2, 2–4. [Google Scholar] [CrossRef]
- Friedrich, A.W.; Bielaszewska, M.; Zhang, W.L.; Pulz, M.; Kuczius, T.; Ammon, A.; Karch, H. Escherichia coli harboring Shiga toxin 2 gene variants: Frequency and association with clinical symptoms. J. Infect. Dis. 2002, 185, 74–84. [Google Scholar] [CrossRef]
- Jelacic, J.K.; Damrow, T.; Chen, G.S.; Jelacic, S.; Bielaszewska, M.; Ciol, M.; Carvalho, H.M.; Melton-Celsa, A.R.; O’Brien, A.D.; Tarr, P.I. Shiga toxin-producing Escherichia coli in Montana: Bacterial genotypes and clinical profiles. J. Infect. Dis. 2003, 188, 719–729. [Google Scholar] [CrossRef]
- Bai, X.; Wang, H.; Xin, Y.; Wei, R.; Tang, X.; Zhao, A.; Sun, H.; Zhang, W.; Wang, Y.; Xu, Y.; et al. Prevalence and characteristics of Shiga toxin-producing Escherichia coli isolated from retail raw meats in China. Int. J. Food Microbiol. 2015, 200, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Yang, X.; Liu, Q.; Zhang, Y.; Jiang, D.; Jiao, H.; Yang, Y.; Xiong, Y.; Bai, X.; Hou, P. High prevalence and pathogenic potential of Shiga toxin-producing Escherichia coli strains in raw mutton and beef in Shandong, China. Curr. Res. Food Sci. 2022, 5, 1596–1602. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wu, Y.; Liu, Q.; Sun, H.; Luo, M.; Xiong, Y.; Matussek, A.; Hu, B.; Bai, X. Genomic characteristics of Stx2e-producing Escherichia coli strains derived from humans, animals, and meats. Pathogens 2021, 10, 1551. [Google Scholar] [CrossRef] [PubMed]
- Ethelberg, S.; Olsen, K.E.; Scheutz, F.; Jensen, C.; Schiellerup, P.; Enberg, J.; Petersen, A.M.; Olesen, B.; Gerner-Smidt, P.; Molbak, K. Virulence factors for hemolytic uremic syndrome, Denmark. Emerg. Infect. Dis. 2004, 10, 842–847. [Google Scholar] [CrossRef] [PubMed]
- Joseph, A.; Cointe, A.; Mariani Kurkdjian, P.; Rafat, C.; Hertig, A. Shiga toxin-associated hemolytic uremic syndrome: A narrative review. Toxins 2020, 12, 67. [Google Scholar] [CrossRef]
- Ichimura, K.; Shimizu, T.; Matsumoto, A.; Hirai, S.; Yokoyama, E.; Takeuchi, H.; Yahiro, K.; Noda, M. Nitric oxide-enhanced Shiga toxin production was regulated by Fur and RecA in enterohemorrhagic O157. Microbiologyopen 2017, 6, e00461. [Google Scholar] [CrossRef]
- Wagner, P.L.; Livny, J.; Neely, M.N.; Acheson, D.W.; Friedman, D.I.; Waldor, M.K. Bacteriophage control of Shiga toxin 1 production and release by Escherichia coli. Mol. Microbiol. 2002, 44, 957–970. [Google Scholar] [CrossRef]
- Waldor, M.K.; Friedman, D.I. Phage regulatory circuits and virulence gene expression. Curr. Opin. Microbiol. 2005, 8, 459–465. [Google Scholar] [CrossRef]
- Steyert, S.R.; Sahl, J.W.; Fraser, C.M.; Teel, L.D.; Scheutz, F.; Rasko, D.A. Comparative genomics and Stx phage characterization of LEE-negative Shiga toxin-producing Escherichia coli. Front. Cell Infect. Microbiol. 2012, 2, 133. [Google Scholar] [CrossRef]
- Nyong, E.C.; Zaia, S.R.; Allue-Guardia, A.; Rodriguez, A.L.; Irion-Byrd, Z.; Koenig, S.S.K.; Feng, P.; Bono, J.L.; Eppinger, M. Pathogenomes of atypical non-Shigatoxigenic Escherichia coli NSF/SF O157:H7/NM: Comprehensive phylogenomic analysis using closed genomes. Front. Microbiol. 2020, 11, 619. [Google Scholar] [CrossRef]
- Serra-Moreno, R.; Jofre, J.; Muniesa, M. Insertion site occupancy by Stx2 bacteriophages depends on the locus availability of the host strain chromosome. J. Bacteriol. 2007, 189, 6645–6654. [Google Scholar] [CrossRef] [PubMed]
- Oporto, B.; Ocejo, M.; Alkorta, M.; Marimon, J.M.; Montes, M.; Hurtado, A. Zoonotic approach to Shiga toxin-producing Escherichia coli: Integrated analysis of virulence and antimicrobial resistance in ruminants and humans. Epidemiol. Infect. 2019, 147, e164. [Google Scholar] [CrossRef]
- Mir, R.A.; Kudva, I.T. Antibiotic-resistant Shiga toxin-producing Escherichia coli: An overview of prevalence and intervention strategies. Zoonoses Public Health 2019, 66, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Day, M.; Doumith, M.; Jenkins, C.; Dallman, T.J.; Hopkins, K.L.; Elson, R.; Godbole, G.; Woodford, N. Antimicrobial resistance in Shiga toxin-producing Escherichia coli serogroups O157 and O26 isolated from human cases of diarrhoeal disease in England, 2015. J. Antimicrob. Chemother. 2017, 72, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Zhang, W.; Tang, X.; Xin, Y.; Xu, Y.; Sun, H.; Luo, X.; Pu, J.; Xu, J.; Xiong, Y.; et al. Shiga toxin-producing Escherichia coli in plateau pika (Ochotona curzoniae) on the Qinghai-Tibetan plateau, China. Front. Microbiol. 2016, 7, 375. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Liu, Q.; Bai, X.; Hu, B.; Jiang, D.; Jiao, H.; Lu, L.; Fan, R.; Hou, P.; Matussek, A.; et al. High prevalence and persistence of Escherichia coli strains producing Shiga toxin subtype 2k in goat herds. Microbiol. Spectr. 2022, 10, e0157122. [Google Scholar] [CrossRef]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Croucher, N.J.; Page, A.J.; Connor, T.R.; Delaney, A.J.; Keane, J.A.; Bentley, S.D.; Parkhill, J.; Harris, S.R. Rapid phylogenetic analysis of large samples of recombinant bacterial whole genome sequences using Gubbins. Nucleic Acids Res. 2015, 43, e15. [Google Scholar] [CrossRef]
- Xie, J.; Chen, Y.; Cai, G.; Cai, R.; Hu, Z.; Wang, H. Tree Visualization By One Table (tvBOT): A web application for visualizing, modifying and annotating phylogenetic trees. Nucleic Acids Res. 2023, 51, W587–W592. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sampling Site | No. of Samples | No. of STEC Isolates | Prevalence of STEC (%) |
|---|---|---|---|
| Guangdong | 1349 | 3 | 0.222 |
| Qinghai | 49 | 1 | 2.041 |
| Guangxi | 2589 | 6 | 0.232 |
| Total | 3987 | 10 | 0.251 |
| Strain | Serotype | Sequence Type | stx Subtype | Sampling Year | Antimicrobial Resistant Phenotypes | Antimicrobial Resistant Genes |
|---|---|---|---|---|---|---|
| STEC434 | O43:H2 | 937 | stx2d | 2014 | - | - |
| STEC435 | O91:H14 | 33 | stx1a+stx2b | 2013 | - | - |
| STEC438 | ONT:H21 | 40 | stx1c | 2014 | Streptomycin, Tetracycline | ant(3″)-Ia, tet(A) |
| STEC509 | O8:H19 | 21 | stx2e | 2016 | Tetracycline | tet(B) |
| STEC1585 | O178:H7 | 278 | stx1c | 2022 | - | - |
| STEC1586 | O112ab:H8 | 75 | stx1c+stx2b | 2022 | - | fosA7 |
| STEC1587 | O21:H25 | 971 | stx1a | 2022 | - | - |
| STEC1588 | O112ac:H19 | 2038 | stx1a+stx2a | 2022 | - | - |
| STEC1589 | O112ab:H8 | 75 | stx1c | 2022 | - | fosA7 |
| STEC1590 | O112ab:H8 | 75 | stx1c+stx2b | 2022 | - | fosA7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sui, X.; Yang, X.; Luo, M.; Wang, H.; Liu, Q.; Sun, H.; Jin, Y.; Wu, Y.; Bai, X.; Xiong, Y. Characteristics of Shiga Toxin-Producing Escherichia coli Circulating in Asymptomatic Food Handlers. Toxins 2023, 15, 640. https://doi.org/10.3390/toxins15110640
Sui X, Yang X, Luo M, Wang H, Liu Q, Sun H, Jin Y, Wu Y, Bai X, Xiong Y. Characteristics of Shiga Toxin-Producing Escherichia coli Circulating in Asymptomatic Food Handlers. Toxins. 2023; 15(11):640. https://doi.org/10.3390/toxins15110640
Chicago/Turabian StyleSui, Xinxia, Xi Yang, Ming Luo, Hua Wang, Qian Liu, Hui Sun, Yujuan Jin, Yannong Wu, Xiangning Bai, and Yanwen Xiong. 2023. "Characteristics of Shiga Toxin-Producing Escherichia coli Circulating in Asymptomatic Food Handlers" Toxins 15, no. 11: 640. https://doi.org/10.3390/toxins15110640
APA StyleSui, X., Yang, X., Luo, M., Wang, H., Liu, Q., Sun, H., Jin, Y., Wu, Y., Bai, X., & Xiong, Y. (2023). Characteristics of Shiga Toxin-Producing Escherichia coli Circulating in Asymptomatic Food Handlers. Toxins, 15(11), 640. https://doi.org/10.3390/toxins15110640