Fusarium Head Blight on Wheat: Biology, Modern Detection and Diagnosis and Integrated Disease Management


Abstract
:1. Introduction
2. Fusarium Head Blight Epidemics on Wheat
3. The Pathogen
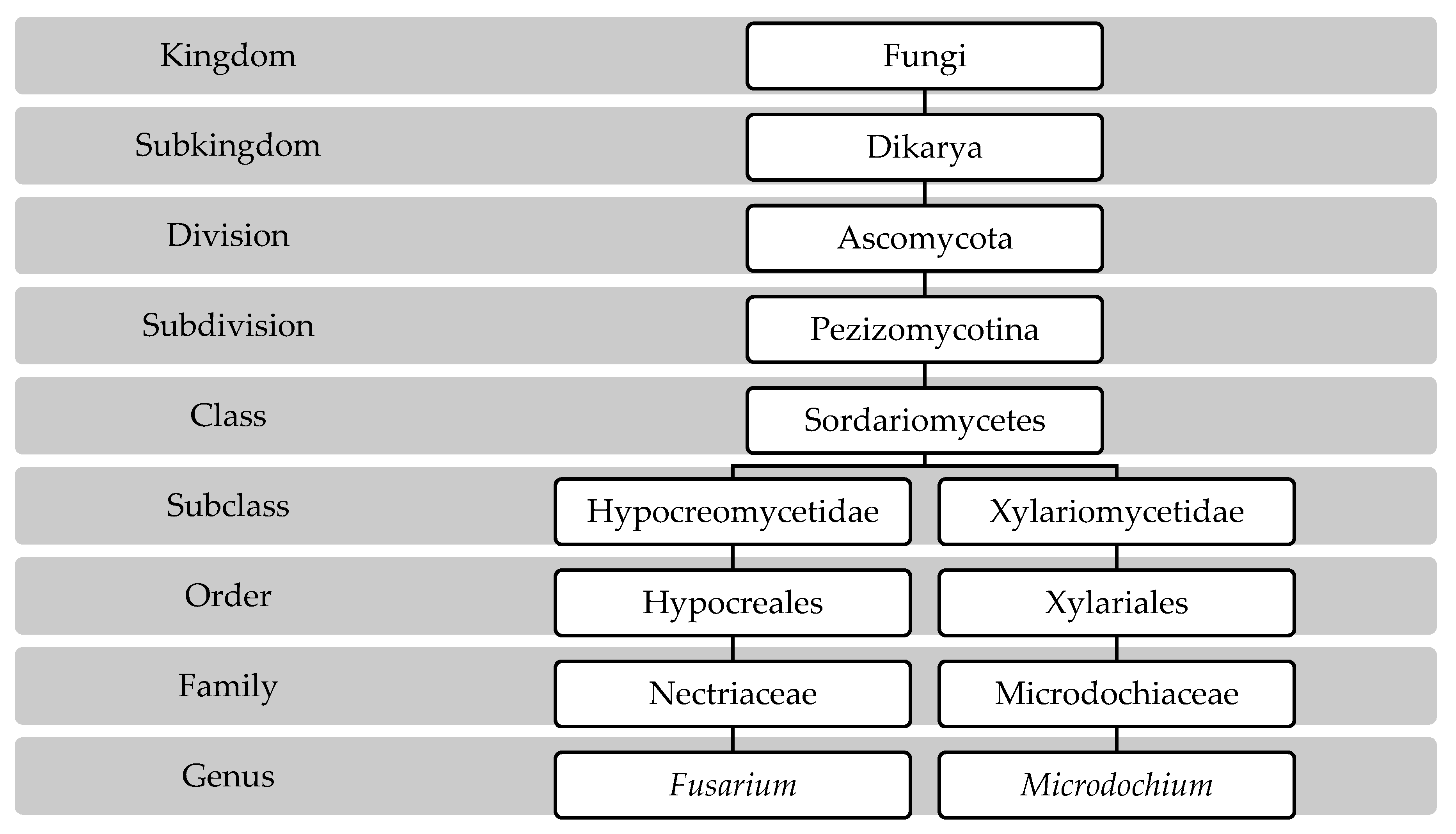
3.1. Pathogen Taxonomy
3.2. Fusarium Species Involved in FHB
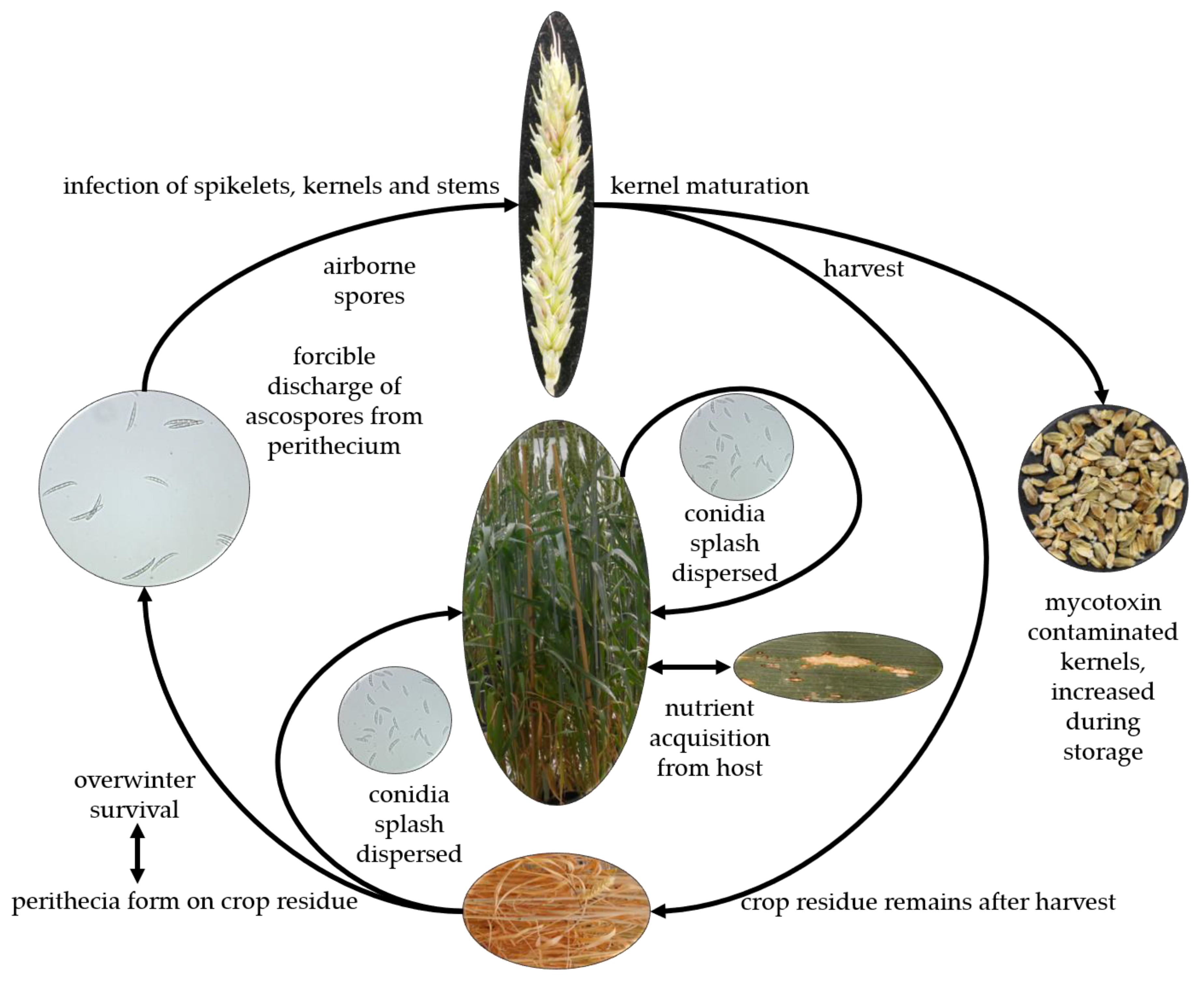
3.3. Life Cycle and Pathogenesis
3.4. Symptoms

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathogen | Teleomorph | Mycotoxin Profile | Reference | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Trichothecenes | Zearalenone | Fusaric Acid | Fumonisins | Emerging Toxins | |||||||||||||||||
| Trichothecenes A | Trichothecenes B | Enniatins | Beauvericin | Fusaproliferin | Moniliformin | ||||||||||||||||
| T-2 toxin | HT-2 toxin | DAS | MAS | NEO | NX-2 | NX-3 | DON | 3-ADON | 15-ADON | NIV | 4-ANIV | ||||||||||
| F. acuminatum Ellis & Everh. | + | + | + | + | + | + | + | [46,47,48] | |||||||||||||
| F. avenaceum (Fr.) Sacc. | + | + | + | + | [46,47,49,50] | ||||||||||||||||
| F. crookwellense L.W. Burgess, P.E. Nelson & Toussoun synonym F. cerealis (Cooke) Sacc. | - | + | + | + | [47,50,51,52] | ||||||||||||||||
| F. culmorum (W.G. Sm.) Sacc. | - | + | + | + | + | + | + | [34,39,53,54,55,56] | |||||||||||||
| F. equiseti (Corda) Sacc. | + | + | + | + | + | + | + | + | [34,48,57] | ||||||||||||
| F. graminearum Schwabe | + | + | + | + | + | + | + | + | + | [34,46,53,56,58,59,60] | |||||||||||
| F. lateritium Nees | + | + | [61,62] | ||||||||||||||||||
| F. oxysporum Schltdl. | - | + | + | [7,63] | |||||||||||||||||
| F. poae (Peck) Wollenw. | - | + | + | + | + | + | + | + | + | [7,46,49,57,64] | |||||||||||
| F. proliferatum (Matsush.) Nirenberg | + | + | + | + | + | + | + | [49,65,66] | |||||||||||||
| F. sambucinum Fuckel | + | + | + | + | + | + | + | + | + | + | [49,61,67] | ||||||||||
| F. semitectum Berk. & Ravenel | - | + | + | [68,69] | |||||||||||||||||
| F. sporotrichioides Sherb. | - | + | + | + | + | + | + | + | + | + | [49,57,70] | ||||||||||
| F. subglutinans (Wollenw. & Reinking) P.E. Nelson, Toussoun & Marasas | + | + | + | + | + | [7,61,71] | |||||||||||||||
| F. tricinctum (Corda) Sacc. | + | + | + | [49] | |||||||||||||||||
| F. verticillioides (Sacc.) Nirenberg synonym F. moniliforme J. Sheld. | + | + | + | [7,72] | |||||||||||||||||
| Microdochium nivale (Fr.) Samuels & I.C. Hallett | + | [10] | |||||||||||||||||||
| Microdochium majus (Wollenw.) Glynn & S.G.Edwards | + | [10] | |||||||||||||||||||
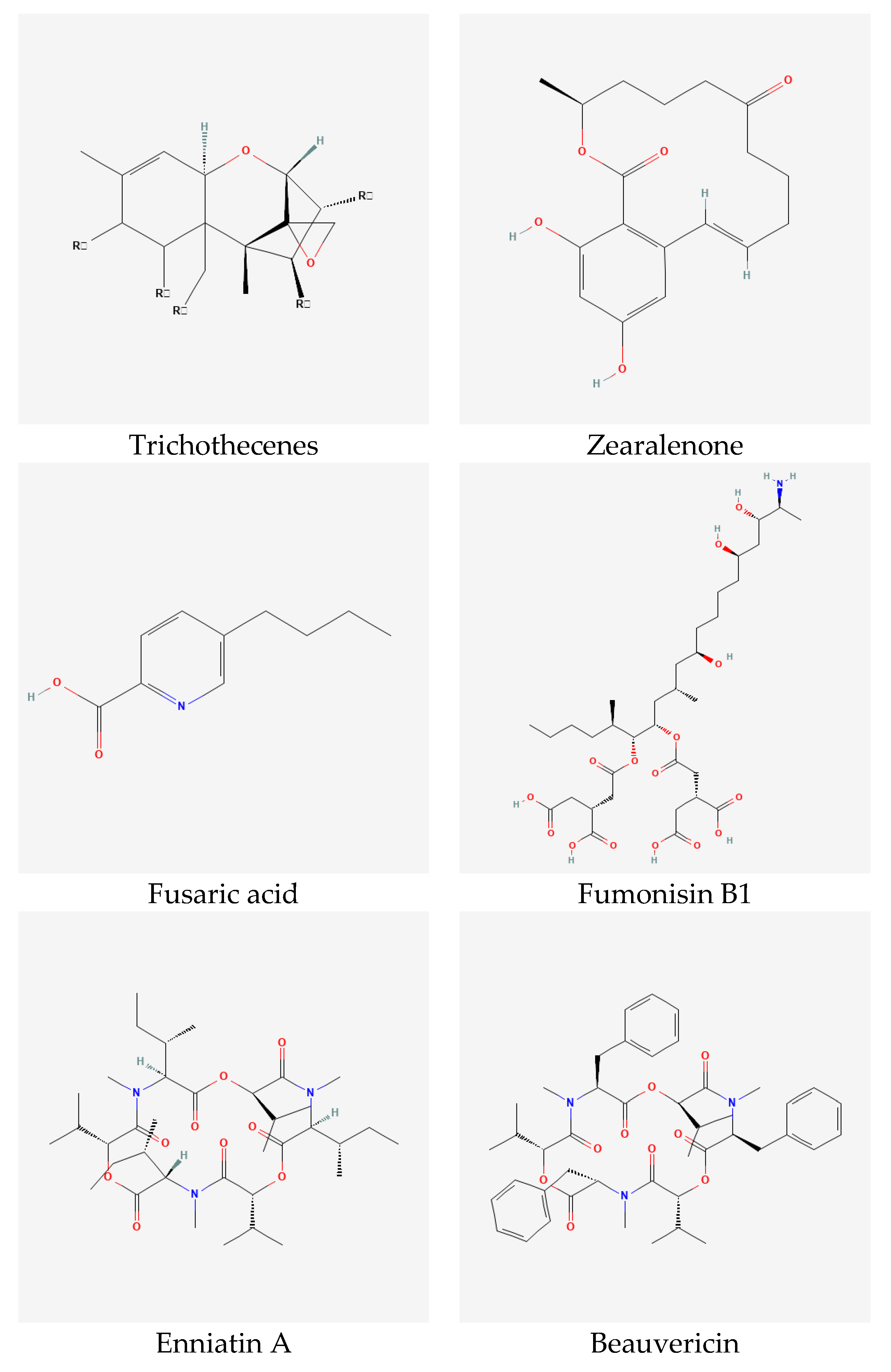
4. Mycotoxins
4.1. Trichothecenes
4.2. Zearalenone
4.3. Fusaric Acid
4.4. Fumonisins
4.5. Emerging Toxins
4.5.1. Enniatins and Beauvericin
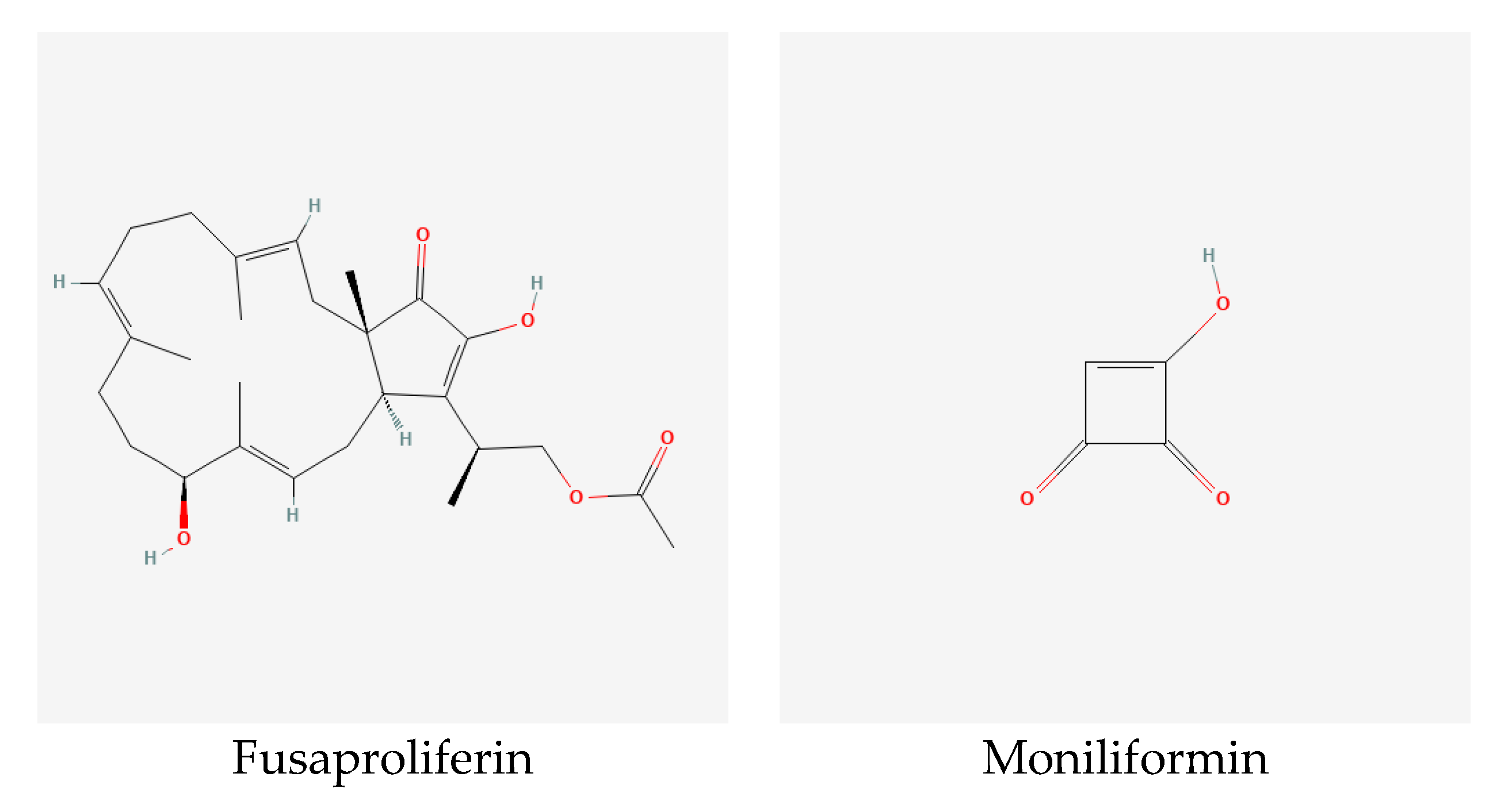
4.5.2. Fusaproliferin
4.5.3. Moniliformin
4.6. Masked Mycotoxins
5. Fusarium Head Blight Diagnosis on Wheat
5.1. Selective Media
5.2. Immunological Method
5.3. Molecular Method
| Pathogen | Primer Name | Primer Sequence (5′-3′) | Amplified Fragment | Reference |
|---|---|---|---|---|
| F. acuminatum | FAC-F FAC-R | GGGATATCGGGCCTCA GGGATATCGGCAAGATCG | 602 bp | [109] |
| F. avenaceum | Fave574 fwd Fave627 rev | TATGTTGTCACTGTCTCACACCACC AGAGGGATGTTAGCATGATGAAG | EF1α gene | [110] |
| F. crookwellense synonym F. cerealis | CRO-A F CRO-A R | CTCAGTGTCCACCGCGTTGCGTAG CTCAGTGTCCCAATCAAATAGTCC | 842 bp | [111] |
| F. culmorum | OPT18 F OPT18 R | GATGCCAGACCAAGACGAAG GATGCCAGACGCACTAAGAT | 472 bp | [112] |
| F. equiseti | Feq-F Feq-R | GGCCTGCCGATGCGTC CGATACTGAAACCGACCTC | 990 bp | [113] |
| F. graminearum | Fg16N F Fg16N R | ACAGATGACAAGATTCAGGCACA TTCTTTGACATCTGTTCAACCCA | 280 bp | [114] |
| F. lateritium | ||||
| F. oxysporum | FOF1 FOR1 | ACATACCACTTGTTGCCTCG CGCCAATCAATTTGAGGAACG | 340 bp | [115] |
| F. poae | Fp82 F Fp82 R | CAAGCAAACAGGCTCTTCACC TGTTCCACCTCAGTGACAGGTT | 220 bp | [116] |
| F. proliferatum | Fp3-F Fp4-R | CGGCCACCAGAGGATGTG CAACACGAATCGCTTCCTGAC | 230 bp | [117] |
| F. sambucinum | FSF1 FSR1 | ACATACCTTTATGTTGCCTCG GGAGTGTCAGACGACAGCT | 315 bp | [115] |
| F. semitectum | ||||
| F. sporotrichioides | Fspor F1 Lanspo R1 | CGCACAACGCAAACTCATC TACAAGAAGACGTGGCGATAT | 332 bp | [118] |
| F. subglutinans | 61-2F 61-2R | GGCCACTCAAGAGGCGAAAG GTCAGACCAGAGCAATGGGC | 445 bp | [119] |
| F. tricinctum | Ftri573 fwd Ftri630 rev | TTGGTATGTTGTCACTGTCTCACACTAT TGACAGAGATGTTAGCATGATGCA | EF1α gene | [110] |
| F. verticillioides synonym F. moniliforme | VERT1 VERT2 | GTCAGAATCCATGCCAGAACG CACCCGCAGCAATCCATCAG | 800 bp | [120] |
| Microdochium nivale | Y13N F Y13N R | ACCAGCCGATTTGTGGTTATG GGTCACGAGGCAGAGTTCG | 300 bp | [121] |
| Microdochium majus | Y13M F Y13M R | CTTGAGGCGGAAGATCGC ATCCCTTTTCCGGGGTTG | 220 bp | [121] |
6. Integrated Management of Fusarium Head Blight
6.1. Cultural Practices
6.2. Host Plant Resistance to Fusarium Head Blight
| QTL | Location | Source | Resistance Type | Reference |
|---|---|---|---|---|
| Fhb1 | 3BS | Sumai 3 and Nyubai | Type II | [132] |
| Fhb2 | 6BS | Sumai 3 | Type II | [133] |
| Fhb3 | 7AS | Leymus racemosus | Type II | [134] |
| Fhb4 | 4BL | Wangshuibai | Type I | [135] |
| Fhb5 | 5AS | Wangshuibai and Sumai 3 | Type I | [136] |
| Fhb6 | 1AS | Elymus tsukushiensis | Type II | [137] |
| Fhb7 | 7D | Thinopyrum ponticum | Type II | [138] |
6.3. Biological Control
6.4. Chemical Control
6.5. Predicting and Detecting Disease Incidence
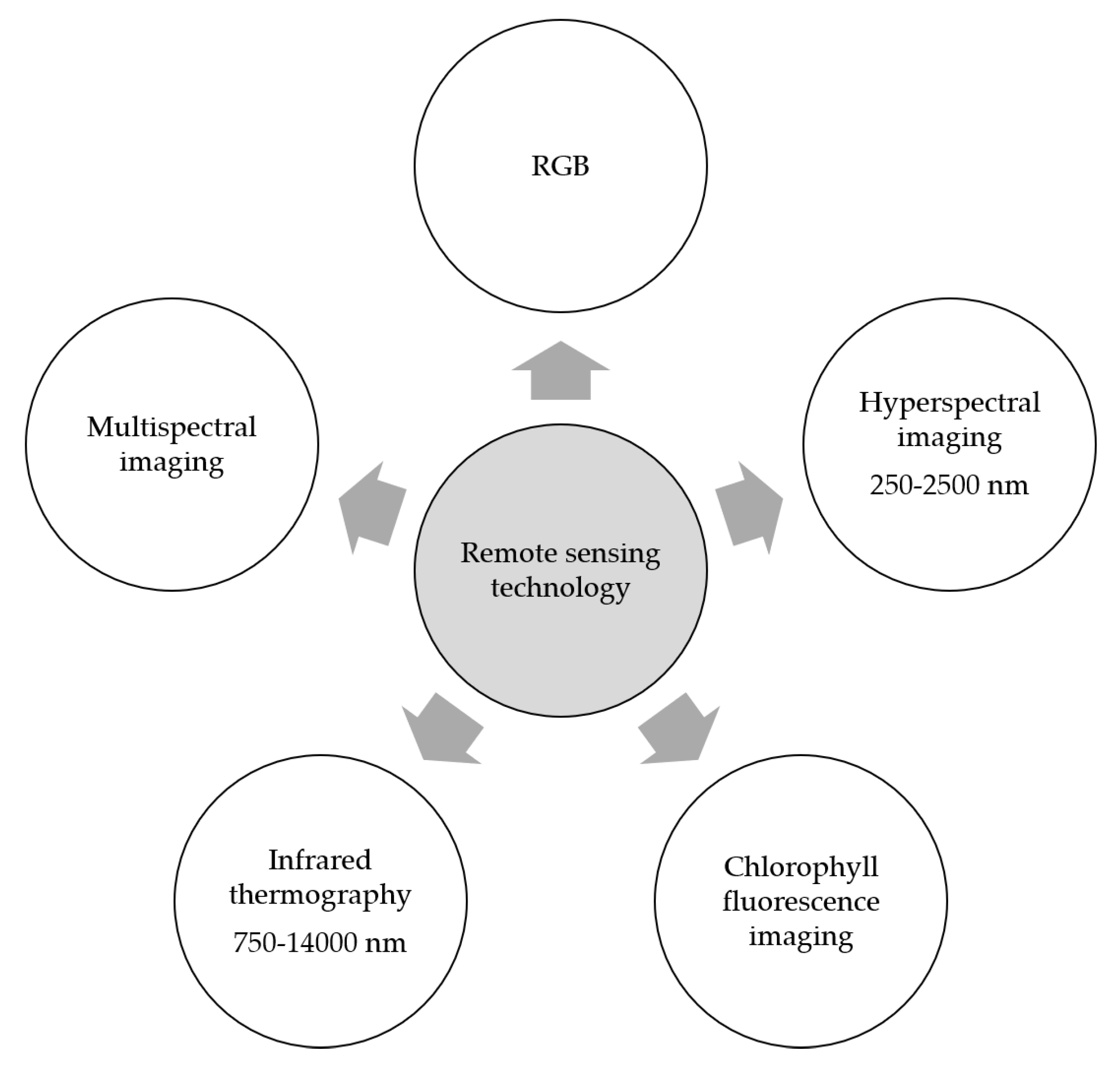
7. Remote Sensing for Monitoring and Phenotyping Fusarium Head Blight
7.1. Spectral Techniques
7.2. Infrared Thermography
7.3. Chlorophyll Fluorescence Imaging
8. Future Perspectives
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- FAO Food and Agriculture Organization of the United Nations FAOStat. Available online: https://www.fao.org/faostat/en/#data (accessed on 22 November 2022).
- CABI Centre for Agriculture and Bioscience International Crop Protection Compendium. Available online: https://www.cabi.org/publishing-products/crop-protection-compendium/ (accessed on 22 November 2022).
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The Global Burden of Pathogens and Pests on Major Food Crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef]
- Summerell, B.A. Resolving Fusarium: Current Status of the Genus. Annu. Rev. Phytopathol. 2019, 57, 323–339. [Google Scholar] [CrossRef]
- Parry, D.W.; Jenkinson, P.; McLeod, L. Fusarium Ear Blight (Scab) in Small Grain Cereals—A Review. Ptant Pathol. 1995, 44, 207–238. [Google Scholar] [CrossRef]
- McMullen, M.; Bergstrom, G.; de Wolf, E.; Dill-Macky, R.; Hershman, D.; Shaner, G.; van Sanford, D. A Unified Effort to Fight an Enemy of Wheat and Barley: Fusarium Head Blight. Plant Dis. 2012, 96, 1712–1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrigo, D.; Raiola, A.; Causin, R. Fusarium Toxins in Cereals: Occurrence, Legislation, Factors Promoting the Appearance and Their Management. Molecules 2016, 21, 627. [Google Scholar] [CrossRef] [Green Version]
- Smith, W. New Diseases of Wheat, Barley, and Rye-Grass. In Diseases of Field and Garden Crops; MacMillan and Co.: London, UK, 1884; pp. 208–213. [Google Scholar]
- McMullen, M.; Jones, R.; Gallenberg, D. Scab of Wheat and Barley: A Re-Emerging Disease of Devastating Impact. Plant Dis. 1997, 81, 1340–1348. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.M.; Nicholson, P.; Thomsett, M.A.; Simpson, D.; Cooke, B.M.; Doohan, F.M.; Brennan, J.; Monaghan, S.; Moretti, A.; Mule, G.; et al. Relationship between the Fungal Complex Causing Fusarium Head Blight of Wheat and Environmental Conditions. Phytopathology 2008, 98, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Mahlein, A.K.; Kuska, M.T.; Behmann, J.; Polder, G.; Walter, A. Hyperspectral Sensors and Imaging Technologies in Phytopathology: State of the Art. Annu. Rev. Phytopathol. 2018, 56, 535–558. [Google Scholar] [CrossRef] [PubMed]
- Furbank, R.T.; Tester, M. Phenomics—Technologies to Relieve the Phenotyping Bottleneck. Trends Plant Sci. 2011, 16, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Mahlein, A.K. Plant Disease Detection by Imaging Sensors—Parallels and Specific Demands for Precision Agriculture and Plant Phenotyping. Plant Dis. 2016, 100, 241–254. [Google Scholar] [CrossRef] [Green Version]
- Alisaac, E.; Behmann, J.; Kuska, M.T.; Dehne, H.W.; Mahlein, A.K. Hyperspectral Quantification of Wheat Resistance to Fusarium Head Blight: Comparison of Two Fusarium Species. Eur. J. Plant Pathol. 2018, 152, 869–884. [Google Scholar] [CrossRef]
- Alisaac, E.; Behmann, J.; Rathgeb, A.; Karlovsky, P.; Dehne, H.W.; Mahlein, A.K. Assessment of Fusarium Infection and Mycotoxin Contamination of Wheat Kernels and Flour Using Hyperspectral Imaging. Toxins 2019, 11, 556. [Google Scholar] [CrossRef] [Green Version]
- Mahlein, A.K.; Alisaac, E.; al Masri, A.; Behmann, J.; Dehne, H.W.; Oerke, E.C. Comparison and Combination of Thermal, Fluorescence, and Hyperspectral Imaging for Monitoring Fusarium Head Blight of Wheat on Spikelet Scale. Sensors 2019, 19, 2281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, J.D.; Culley, J.; Fraser, K.; Hubbard, S.; Meloche, F.; Ouellet, T.; Seaman, W.L.; Seifert, K.A.; Turkington, K.; Voldeng, H. Effect of Tillage Practice on Fusarium Head Blight of Wheat. Can. J. Plant Pathol. 1998, 20, 95–103. [Google Scholar] [CrossRef]
- Dill-Macky, R.; Jones, R.K. The Effect of Previous Crop Residues and Tillage on Fusarium Head Blight of Wheat. Plant Dis. 2000, 84, 71–76. [Google Scholar] [CrossRef] [Green Version]
- Shah, D.A.; de Wolf, E.D.; Paul, P.A.; Madden, L.v. Functional Data Analysis of Weather Variables Linked to Fusarium Head Blight Epidemics in the United States. Phytopathology 2019, 109, 96–110. [Google Scholar] [CrossRef] [Green Version]
- Obanor, F.; Neate, S.; Simpfendorfer, S.; Sabburg, R.; Wilson, P.; Chakraborty, S. Fusarium graminearum and Fusarium pseudograminearum Caused the 2010 Head Blight Epidemics in Australia. Plant Pathol. 2013, 62, 79–91. [Google Scholar] [CrossRef] [Green Version]
- Torres, A.M.; Palacios, S.A.; Yerkovich, N.; Palazzini, J.M.; Battilani, P.; Leslie, J.F.; Logrieco, A.F.; Chulze, S.N. Fusarium Head Blight and Mycotoxins in Wheat: Prevention and Control Strategies across the Food Chain. World Mycotoxin J. 2019, 12, 333–355. [Google Scholar] [CrossRef]
- Johns, L.E.; Bebber, D.P.; Gurr, S.J.; Brown, A. Emerging Health Threat and Cost of Fusarium Mycotoxins in European Wheat. Nat. Food 2022, 3, 1014–1019. [Google Scholar] [CrossRef]
- Zhu, Z.; Hao, Y.; Mergoum, M.; Bai, G.; Humphreys, G.; Cloutier, S.; Xia, X.; He, Z. Breeding Wheat for Resistance to Fusarium Head Blight in the Global North: China, USA, and Canada. Crop J. 2019, 7, 730–738. [Google Scholar] [CrossRef]
- Aboukhaddour, R.; Fetch, T.; McCallum, B.D.; Harding, M.W.; Beres, B.L.; Graf, R.J. Wheat Diseases on the Prairies: A Canadian Story. Plant Pathol. 2020, 69, 418–432. [Google Scholar] [CrossRef]
- Xue, A.G.; Chen, Y.; Seifert, K.; Guo, W.; Blackwell, B.A.; Harris, L.J.; Overy, D.P. Prevalence of Fusarium Species Causing Head Blight of Spring Wheat, Barley and Oat in Ontario during 2001–2017. Can. J. Plant Pathol. 2019, 41, 392–402. [Google Scholar] [CrossRef]
- Palazzini, J.; Fumero, V.; Yerkovich, N.; Barros, G.; Cuniberti, M.; Chulze, S. Correlation between Fusarium graminearum and Deoxynivalenol during the 2012/13 Wheat Fusarium Head Blight Outbreak in Argentina. Cereal. Res. Commun. 2015, 43, 627–637. [Google Scholar] [CrossRef] [Green Version]
- del Ponte, E.M.; Fernandes, J.M.C.; Pavan, W.; Baethgen, W.E. A Model-Based Assessment of the Impacts of Climate Variability on Fusarium Head Blight Seasonal Risk in Southern Brazil. J. Phytopathol. 2009, 157, 675–681. [Google Scholar] [CrossRef]
- Geiser, D.M.; Aoki, T.; Bacon, C.W.; Baker, S.E.; Bhattacharyya, M.K.; Brandt, M.E.; Brown, D.W.; Burgess, L.W.; Chulze, S.; Coleman, J.J.; et al. One Fungus, One Name: Defining the Genus Fusarium in a Scientifically Robust Way That Preserves Longstanding Use. Phytopathology 2013, 103, 400–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MycoBank An Initiative of the International Mycological Association (IMA) and the Westerdijk Fungal Biodiversity Institute MycoBank Database. Available online: https://www.mycobank.org/ (accessed on 22 November 2022).
- Haile, J.K.; N’Diaye, A.; Walkowiak, S.; Nilsen, K.T.; Clarke, J.M.; Kutcher, H.R.; Steiner, B.; Buerstmayr, H.; Pozniak, C.J. Fusarium Head Blight in Durum Wheat: Recent Status, Breeding Directions, and Future Research Prospects. Phytopathology 2019, 109, 1664–1675. [Google Scholar] [CrossRef]
- Nielsen, L.K.; Cook, D.J.; Edwards, S.G.; Ray, R.v. The Prevalence and Impact of Fusarium Head Blight Pathogens and Mycotoxins on Malting Barley Quality in UK. Int. J. Food Microbiol. 2014, 179, 38–49. [Google Scholar] [CrossRef] [Green Version]
- Oerke, E.C.; Meier, A.; Dehne, H.W.; Sulyok, M.; Krska, R.; Steiner, U. Spatial Variability of Fusarium Head Blight Pathogens and Associated Mycotoxins in Wheat Crops. Plant Pathol. 2010, 59, 671–682. [Google Scholar] [CrossRef]
- Spanic, V.; Lemmens, M.; Drezner, G. Morphological and Molecular Identification of Fusarium Species Associated with Head Blight on Wheat in East Croatia. Eur. J. Plant Pathol. 2010, 128, 511–516. [Google Scholar] [CrossRef]
- Alkadri, D.; Nipoti, P.; Döll, K.; Karlovsky, P.; Prodi, A.; Pisi, A. Study of Fungal Colonization of Wheat Kernels in Syria with a Focus on Fusarium Species. Int. J. Mol. Sci. 2013, 14, 5938–5951. [Google Scholar] [CrossRef] [PubMed]
- Trail, F. For Blighted Waves of Grain: Fusarium graminearum in the Postgenomics Era. Plant Physiol. 2009, 149, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Mourelos, C.A.; Malbrán, I.; Balatti, P.A.; Ghiringhelli, P.D.; Lori, G.A. Gramineous and Non-Gramineous Weed Species as Alternative Hosts of Fusarium graminearum, Causal Agent of Fusarium Head Blight of Wheat, in Argentina. Crop Prot. 2014, 65, 100–104. [Google Scholar] [CrossRef]
- Suproniene, S.; Kadziene, G.; Irzykowski, W.; Sneideris, D.; Ivanauskas, A.; Sakalauskas, S.; Serbiak, P.; Svegzda, P.; Auskalniene, O.; Jedryczka, M. Weed Species within Cereal Crop Rotations Can Serve as Alternative Hosts for Fusarium graminearum Causing Fusarium Head Blight of Wheat. Fungal. Ecol. 2019, 37, 30–37. [Google Scholar] [CrossRef]
- Dong, F.; Xu, J.; Zhang, X.; Wang, S.; Xing, Y.; Mokoena, M.P.; Olaniran, A.O.; Shi, J. Gramineous Weeds near Paddy Fields Are Alternative Hosts for the Fusarium graminearum Species Complex That Causes Fusarium Head Blight in Rice. Plant Pathol. 2020, 69, 433–441. [Google Scholar] [CrossRef]
- Kang, Z.; Buchenauer, H. Immunocytochemical Localization of Fusarium Toxins in Infected Wheat Spikes by Fusarium culmorum. Physiol. Mol. Plant Pathol. 1999, 55, 275–288. [Google Scholar] [CrossRef]
- Brown, N.A.; Urban, M.; van de Meene, A.M.L.; Hammond-Kosack, K.E. The Infection Biology of Fusarium graminearum: Defining the Pathways of Spikelet to Spikelet Colonisation in Wheat Ears. Fungal. Biol. 2010, 114, 555–571. [Google Scholar] [CrossRef] [PubMed]
- Divon, H.H.; Bøe, L.; Tveit, M.M.N.; Klemsdal, S.S. Infection Pathways and Penetration Modes of Fusarium langsethiae. Eur. J. Plant Pathol. 2019, 154, 259–271. [Google Scholar] [CrossRef]
- Dweba, C.C.; Figlan, S.; Shimelis, H.A.; Motaung, T.E.; Sydenham, S.; Mwadzingeni, L.; Tsilo, T.J. Fusarium Head Blight of Wheat: Pathogenesis and Control Strategies. Crop Prot. 2017, 91, 114–122. [Google Scholar] [CrossRef]
- Scherm, B.; Balmas, V.; Spanu, F.; Pani, G.; Delogu, G.; Pasquali, M.; Migheli, Q. Fusarium culmorum: Causal Agent of Foot and Root Rot and Head Blight on Wheat. Mol. Plant Pathol. 2013, 14, 323–341. [Google Scholar] [CrossRef]
- Stenglein, S.A. Fusarium poae a Pathogen That Needs More Attention. J. Plant Pathol. 2009, 91, 25–36. [Google Scholar]
- Stenglein, S.A.; Dinolfo, M.I.; Barros, G.; Bongiorno, F.; Chulze, S.N.; Moreno, M.V. Fusarium poae Pathogenicity and Mycotoxin Accumulation on Selected Wheat and Barley Genotypes at a Single Location in Argentina. Plant Dis. 2014, 98, 1733–1738. [Google Scholar] [CrossRef] [Green Version]
- Beccari, G.; Arellano, C.; Covarelli, L.; Tini, F.; Sulyok, M.; Cowger, C. Effect of Wheat Infection Timing on Fusarium Head Blight Causal Agents and Secondary Metabolites in Grain. Int. J. Food Microbiol. 2019, 290, 214–225. [Google Scholar] [CrossRef]
- Bottalico, A.; Perrone, G. Toxigenic Fusarium Species and Mycotoxins Associated with Head Blight in Small-Grain Cereals in Europe. Eur. J. Plant Pathol. 2002, 108, 611–624. [Google Scholar] [CrossRef]
- Marín, P.; Moretti, A.; Ritieni, A.; Jurado, M.; Vázquez, C.; González-Jaén, M.T. Phylogenetic Analyses and Toxigenic Profiles of Fusarium equiseti and Fusarium acuminatum Isolated from Cereals from Southern Europe. Food Microbiol. 2012, 31, 229–237. [Google Scholar] [CrossRef] [Green Version]
- Beccari, G.; Colasante, V.; Tini, F.; Senatore, M.T.; Prodi, A.; Sulyok, M.; Covarelli, L. Causal Agents of Fusarium Head Blight of Durum Wheat (Triticum durum Desf.) in Central Italy and Their in vitro Biosynthesis of Secondary Metabolites. Food Microbiol. 2018, 70, 17–27. [Google Scholar] [CrossRef]
- Glenn, A.E. Mycotoxigenic Fusarium Species in Animal Feed. Anim. Feed Sci. Technol. 2007, 137, 213–240. [Google Scholar] [CrossRef]
- Chandler, E.A.; Simpson, D.R.; Thomsett, M.A.; Nicholson, P. Development of PCR Assays to Tri7 and Tri13 Trichothecene Biosynthetic Genes, and Characterisation of Chemotypes of Fusarium graminearum, Fusarium culmorum and Fusarium cerealis. Physiol. Mol. Plant Pathol. 2003, 62, 355–367. [Google Scholar] [CrossRef]
- Vesonder, R.F.; Golifiski, P.; Plattner, R.; Zietkiewicz, D.L. Mycotoxin Formation by Different Geographic Isolates of Fusarium crookwellense. Mycopathologia 1991, 113, 11–14. [Google Scholar] [CrossRef] [PubMed]
- Alisaac, E.; Rathgeb, A.; Karlovsky, P.; Mahlein, A.K. Fusarium Head Blight: Effect of Infection Timing on Spread of Fusarium graminearum and Spatial Distribution of Deoxynivalenol within Wheat Spikes. Microorganisms 2021, 9, 79. [Google Scholar] [CrossRef]
- Basler, R. Diversity of Fusarium Species Isolated from UK Forage Maize and the Population Structure of F. graminearum from Maize and Wheat. PeerJ 2016, 4, e2143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasquali, M.; Beyer, M.; Logrieco, A.; Audenaert, K.; Balmas, V.; Basler, R.; Boutigny, A.L.; Chrpová, J.; Czembor, E.; Gagkaeva, T.; et al. A European Database of Fusarium graminearum and F. culmorum Trichothecene Genotypes. Front Microbiol. 2016, 7, 406. [Google Scholar] [CrossRef] [Green Version]
- Siou, D.; Gélisse, S.; Laval, V.; Elbelt, S.; Repinçay, C.; Bourdat-Deschamps, M.; Suffert, F.; Lannou, C. Interactions between Head Blight Pathogens: Consequences for Disease Development and Toxin Production in Wheat Spikes. Appl. Environ. Microbiol. 2015, 81, 957–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Covarelli, L.; Beccari, G.; Prodi, A.; Generotti, S.; Etruschi, F.; Meca, G.; Juan, C.; Mañes, J. Biosynthesis of Beauvericin and Enniatins Invitro by Wheat Fusarium Species and Natural Grain Contamination in an Area of Central Italy. Food Microbiol. 2015, 46, 618–626. [Google Scholar] [CrossRef] [PubMed]
- Varga, E.; Wiesenberger, G.; Hametner, C.; Ward, T.J.; Dong, Y.; Schöfbeck, D.; Mccormick, S.; Broz, K.; Stückler, R.; Schuhmacher, R.; et al. New Tricks of an Old Enemy: Isolates of Fusarium graminearum Produce a Type A Trichothecene Mycotoxin. Environ. Microbiol. 2015, 17, 2588–2600. [Google Scholar] [CrossRef] [PubMed]
- Crippin, T.; Renaud, J.B.; Sumarah, M.W.; Miller, J.D. Comparing Genotype and Chemotype of Fusarium graminearum from Cereals in Ontario, Canada. PLoS ONE 2019, 14, e0216735. [Google Scholar] [CrossRef]
- Lofgren, L.; Riddle, J.; Dong, Y.; Kuhnem, P.R.; Cummings, J.A.; del Ponte, E.M.; Bergstrom, G.C.; Kistler, H.C. A High Proportion of NX-2 Genotype Strains Are Found among Fusarium graminearum Isolates from Northeastern New York State. Eur. J. Plant Pathol. 2018, 150, 791–796. [Google Scholar] [CrossRef]
- Jestoi, M. Emerging Fusarium-Mycotoxins Fusaproliferin, Beauvericin, Enniatins, and Moniliformin—A Review. Crit. Rev. Food Sci. Nutr. 2008, 48, 21–49. [Google Scholar] [CrossRef]
- Stanciu, O.; Juan, C.; Miere, D.; Loghin, F.; Mañes, J. Presence of Enniatins and Beauvericin in Romanian Wheat Samples: From Raw Material to Products for Direct Human Consumption. Toxins 2017, 9, 189. [Google Scholar] [CrossRef] [Green Version]
- Shi, W.; Tan, Y.; Wang, S.; Gardiner, D.M.; de Saeger, S.; Liao, Y.; Wang, C.; Fan, Y.; Wang, Z.; Wu, A. Mycotoxigenic Potentials of Fusarium Species in Various Culture Matrices Revealed by Mycotoxin Profiling. Toxins 2017, 9, 6. [Google Scholar] [CrossRef] [Green Version]
- Covarelli, L.; Beccari, G.; Prodi, A.; Generotti, S.; Etruschi, F.; Juan, C.; Ferrer, E.; Mañes, J. Fusarium Species, Chemotype Characterisation and Trichothecene Contamination of Durum and Soft Wheat in an Area of Central Italy. J. Sci. Food Agric. 2015, 95, 540–551. [Google Scholar] [CrossRef]
- Guo, Z.; DolÏl, K.; Dastjerdi, R.; Karlovsky, P.; Dehne, H.W.; Altincicek, B. Effect of Fungal Colonization of Wheat Grains with Fusarium spp. on Food Choice, Weight Gain and Mortality of Meal Beetle Larvae (Tenebrio molitor). PLoS ONE 2014, 9, e100112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Z.; Pfohl, K.; Karlovsky, P.; Dehne, H.W.; Altincicek, B. Dissemination of Fusarium proliferatum by Mealworm Beetle Tenebrio molitor. PLoS ONE 2018, 13, e0204602. [Google Scholar] [CrossRef] [PubMed]
- Fraeyman, S.; Croubels, S.; Devreese, M.; Antonissen, G. Emerging Fusarium and Alternaria Mycotoxins: Occurrence, Toxicity and Toxicokinetics. Toxins 2017, 9, 228. [Google Scholar] [CrossRef] [Green Version]
- Barros, G.; Zanon, M.S.A.; Palazzini, J.M.; Haidukowski, M.; Pascale, M.; Chulze, S. Trichothecenes and Zearalenone Production by Fusarium equiseti and Fusarium semitectum Species Isolated from Argentinean Soybean. Food Addit. Contam.—Part A 2012, 29, 1436–1442. [Google Scholar] [CrossRef] [PubMed]
- Ezekiel, C.N.; Odebode, A.C.; Fapohunda, S.O. Zearalenone Production by Naturally Occurring Fusarium Species on Maize, Wheat and Soybeans from Nigeria. J. Biol. Environ. Sci. 2008, 2, 77–82. [Google Scholar]
- Nazari, L.; Pattori, E.; Terzi, V.; Morcia, C.; Rossi, V. Influence of Temperature on Infection, Growth, and Mycotoxin Production by Fusarium langsethiae and F. sporotrichioides in Durum Wheat. Food Microbiol. 2014, 39, 19–26. [Google Scholar] [CrossRef]
- Jajić, I.; Dudaš, T.; Krstović, S.; Krska, R.; Sulyok, M.; Bagi, F.; Savić, Z.; Guljaš, D.; Stankov, A. Emerging Fusarium Mycotoxins Fusaproliferin, Beauvericin, Enniatins, and Moniliformin in Serbian Maize. Toxins 2019, 11, 357. [Google Scholar] [CrossRef] [Green Version]
- Mylona, K.; Garcia-Cela, E.; Sulyok, M.; Medina, A.; Magan, N. Influence of Two Garlic-Derived Compounds, Propyl Propane Thiosulfonate (PTS) and Propyl Propane Thiosulfinate (PTSO), on Growth and Mycotoxin Production by Fusarium Species in vitro and in Stored Cereals. Toxins 2019, 11, 495. [Google Scholar] [CrossRef] [Green Version]
- Foroud, N.A.; Baines, D.; Gagkaeva, T.Y.; Thakor, N.; Badea, A.; Steiner, B.; Bürstmayr, M.; Bürstmayr, H. Trichothecenes in Cereal Grains—An Update. Toxins 2019, 11, 634. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Kistler, H.C.; Ma, Z. Fusarium graminearum Trichothecene Mycotoxins: Biosynthesis, Regulation, and Management. Annu. Rev. Phytopathol. 2019, 57, 15–39. [Google Scholar] [CrossRef] [Green Version]
- McCormick, S.P.; Stanley, A.M.; Stover, N.A.; Alexander, N.J. Trichothecenes: From Simple to Complex Mycotoxins. Toxins 2011, 3, 802–814. [Google Scholar] [CrossRef] [PubMed]
- Goswami, R.S.; Kistler, H.C. Pathogenicity and in Planta Mycotoxin Accumulation among Members of the Fusarium graminearum Species Complex on Wheat and Rice. Phytopathology 2005, 95, 1397–1404. [Google Scholar] [CrossRef] [Green Version]
- Spanic, V.; Zdunic, Z.; Drezner, G.; Sarkanj, B. The Pressure of Fusarium Disease and Its Relation with Mycotoxins in the Wheat Grain and Malt. Toxins 2019, 11, 198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogelgsang, S.; Beyer, M.; Pasquali, M.; Jenny, E.; Musa, T.; Bucheli, T.D.; Wettstein, F.E.; Forrer, H.R. An Eight-Year Survey of Wheat Shows Distinctive Effects of Cropping Factors on Different Fusarium Species and Associated Mycotoxins. Eur. J. Agron. 2019, 105, 62–77. [Google Scholar] [CrossRef]
- Kuzdraliński, A.; Solarska, E.; Muszyńska, M. Deoxynivalenol and Zearalenone Occurence in Beers Analysed by an Enzyme-Linked Immunosorbent Assay Method. Food Control 2013, 29, 22–24. [Google Scholar] [CrossRef]
- Tralamazza, S.M.; Bemvenuti, R.H.; Zorzete, P.; de Souza Garcia, F.; Corrêa, B. Fungal Diversity and Natural Occurrence of Deoxynivalenol and Zearalenone in Freshly Harvested Wheat Grains from Brazil. Food Chem. 2016, 196, 445–450. [Google Scholar] [CrossRef]
- Stanciu, O.; Juan, C.; Berrada, H.; Miere, D.; Loghin, F.; Mañes, J. Study on Trichothecene and Zearalenone Presence in Romanian Wheat Relative to Weather Conditions. Toxins 2019, 11, 163. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Yang, C.; Mao, Y.; Hong, X.; Du, D. Zearalenone Contamination in Corn, Corn Products, and Swine Feed in China in 2016–2018 as Assessed by Magnetic Bead Immunoassay. Toxins 2019, 11, 451. [Google Scholar] [CrossRef] [Green Version]
- McLean, M. The Phytotoxicity of Selected Mycotoxins on Mature, Germinating Zea mays Embryos. Mycopathologia 1995, 132, 173–183. [Google Scholar] [CrossRef]
- Bertero, A.; Moretti, A.; Spicer, L.J.; Caloni, F. Fusarium Molds and Mycotoxins: Potential Species-Specific Effects. Toxins 2018, 10, 244. [Google Scholar] [CrossRef] [Green Version]
- López-Díaz, C.; Rahjoo, V.; Sulyok, M.; Ghionna, V.; Martín-Vicente, A.; Capilla, J.; di Pietro, A.; López-Berges, M.S. Fusaric Acid Contributes to Virulence of Fusarium oxysporum on Plant and Mammalian Hosts. Mol. Plant Pathol. 2018, 19, 440–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dall’Asta, C.; Battilani, P. Fumonisins and Their Modified Forms, a Matter of Concern in Future Scenario? World Mycotoxin J. 2016, 9, 727–739. [Google Scholar] [CrossRef]
- Alshannaq, A.; Yu, J.H. Occurrence, Toxicity, and Analysis of Major Mycotoxins in Food. Int. J. Environ. Res. Public Health 2017, 14, 632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, M.S.; Wink, M. Exposure, Occurrence, and Chemistry of Fumonisins and Their Cryptic Derivatives. Compr. Rev. Food Sci. Food Saf. 2018, 17, 769–791. [Google Scholar] [CrossRef] [Green Version]
- Scott, P.M. Recent Research on Fumonisins: A Review. Food Addit. Contam.—Part A 2012, 29, 242–248. [Google Scholar] [CrossRef]
- Kovalsky, P.; Kos, G.; Nährer, K.; Schwab, C.; Jenkins, T.; Schatzmayr, G.; Sulyok, M.; Krska, R. Co-Occurrence of Regulated, Masked and Emerging Mycotoxins and Secondary Metabolites in Finished Feed and Maize–an Extensive Survey. Toxins 2016, 8, 863. [Google Scholar] [CrossRef] [Green Version]
- Stanciu, O.; Juan, C.; Miere, D.; Loghin, F.; Mañes, J. Occurrence and Co-Occurrence of Fusarium Mycotoxins in Wheat Grains and Wheat Flour from Romania. Food Control 2017, 73, 147–155. [Google Scholar] [CrossRef]
- Prosperini, A.; Berrada, H.; Ruiz, M.J.; Caloni, F.; Coccini, T.; Spicer, L.J.; Perego, M.C.; Lafranconi, A. A Review of the Mycotoxin Enniatin B. Front. Public Health 2017, 5, 304. [Google Scholar] [CrossRef]
- Ivanova, L.; Fæste, C.K.; van Pamel, E.; Daeseleire, E.; Callebaut, A.; Uhlig, S. Presence of Enniatin B and Its Hepatic Metabolites in Plasma and Liver Samples from Broilers and Eggs from Laying Hens. World Mycotoxin J. 2014, 7, 167–175. [Google Scholar] [CrossRef]
- Tedjiotsop Feudjio, F.; Dornetshuber, R.; Lemmens, M.; Hoffmann, O.; Lemmens-Gruber, R.; Berger, W. Beauvericin and Enniatin: Emerging Toxins and/or Remedies? World Mycotoxin J. 2010, 3, 415–430. [Google Scholar] [CrossRef]
- Rychlik, M.; Humpf, H.U.; Marko, D.; Dänicke, S.; Mally, A.; Berthiller, F.; Klaffke, H.; Lorenz, N. Proposal of a Comprehensive Definition of Modified and Other Forms of Mycotoxins Including “Masked” Mycotoxins. Mycotoxin Res. 2014, 30, 197–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berthiller, F.; Crews, C.; Dall’Asta, C.; Saeger, S.D.; Haesaert, G.; Karlovsky, P.; Oswald, I.P.; Seefelder, W.; Speijers, G.; Stroka, J. Masked Mycotoxins: A Review. Mol. Nutr. Food Res. 2013, 57, 165–186. [Google Scholar] [CrossRef] [PubMed]
- Gratz, S.W. Do Plant-Bound Masked Mycotoxins Contribute to Toxicity? Toxins 2017, 9, 85. [Google Scholar] [CrossRef]
- Zhang, Z.; Nie, D.; Fan, K.; Yang, J.; Guo, W.; Meng, J.; Zhao, Z.; Han, Z. A Systematic Review of Plant-Conjugated Masked Mycotoxins: Occurrence, Toxicology, and Metabolism. Crit. Rev. Food Sci. Nutr. 2020, 60, 1523–1537. [Google Scholar] [CrossRef]
- Bragulat, M.R.; Martínez, E.; Castellá, G.; Cabañes, F.J. Selective Efficacy of Culture Media Recommended for Isolation and Enumeration of Fusarium spp. J. Food Prot. 2004, 67, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Thrane, U. Comparison of Three Selective Media for Detecting Fusarium Species in Foods: A Collaborative Study. Int. J. Food Microbiol. 1996, 29, 149–156. [Google Scholar] [CrossRef]
- Jung, B.; Lee, S.; Ha, J.; Park, J.C.; Han, S.S.; Hwang, I.; Lee, Y.W.; Lee, J. Development of a Selective Medium for the Fungal Pathogen Fusarium graminearum Using Toxoflavin Produced by the Bacterial Pathogen Burkholderia glumae. Plant Pathol. J. (Faisalabad) 2013, 29, 446–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, N.S.; Schwarz, P.; Dahleen, L.S.; Neate, S.M.; Horsley, R.; Glenn, A.E.; O’Donnell, K. ELISA Analysis for Fusarium in Barley: Development of Methodology and Field Assessment. Crop Sci. 2006, 46, 2636–2642. [Google Scholar] [CrossRef]
- Gan, Z.; Marquardt, R.R.; Abramson, D.; Clear, R.M. The Characterization of Chicken Antibodies Raised against Fusarium spp. by Enzyme-Linked Immunosorbent Assay and Immunoblotting. Int. J. Food Microbiol. 1997, 38, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Brunner, K.; Farnleitner, A.; Mach, R.L. Novel Methods for the Quantification of Pathogenic Fungi in Crop Plants: Quantitative PCR and ELISA Accurately Determine Fusarium Biomass. In Plant Pathology; Cumagun, C.J.R., Ed.; IntechOpen Limited: London, UK, 2012; pp. 203–218. [Google Scholar]
- Henson, J.M.; French, R. The Polymerase Chain Reaction and Plant Disease Diagnosis. Annu. Rev. Phytopathol. 1993, 31, 81–109. [Google Scholar] [CrossRef]
- Schaad, N.W.; Frederick, R.D. Real-Time PCR and Its Application for Rapid Plant Disease Diagnostics. Can. J. Plant Pathol. 2002, 24, 250–258. [Google Scholar] [CrossRef]
- Kuzdraliński, A.; Kot, A.; Szczerba, H.; Nowak, M.; Muszyńska, M. A Review of Conventional PCR Assays for the Detection of Selected Phytopathogens of Wheat. J. Mol. Microbiol. Biotechnol. 2017, 27, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.Y.; Kang, M.R.; Cho, E.J.; Kim, H.K.; Yun, S.H. Specific PCR Detection of Four Quarantine Fusarium Species in Korea. Plant Pathol. J. (Faisalabad) 2010, 26, 409–416. [Google Scholar] [CrossRef] [Green Version]
- Williams, K.J.; Dennis, J.I.; Smyl, C.; Wallwork, H. The Application of Species-Specific Assays Based on the Polymerase Chain Reaction to Analyse Fusarium Crown Rot of Durum Wheat. Australas. Plant Pathol. 2002, 31, 119–127. [Google Scholar] [CrossRef]
- Nicolaisen, M.; Suproniene, S.; Nielsen, L.K.; Lazzaro, I.; Spliid, N.H.; Justesen, A.F. Real-Time PCR for Quantification of Eleven Individual Fusarium Species in Cereals. J. Microbiol. Methods 2009, 76, 234–240. [Google Scholar] [CrossRef]
- Yoder, W.T.; Christianson, L.M. Species-Specific Primers Resolve Members of Fusarium Section Fusarium. Taxonomic Status of the Edible “Quorn” Fungus Reevaluated. Fungal Genet. Biol. 1998, 23, 68–80. [Google Scholar] [CrossRef]
- Schilling, A.G.; Möller, E.M.; Geiger, H.H. Polymerase Chain Reaction-Based Assays for Species-Specific Detection of Fusarium culmorum, F. graminearum and F. avenaceum. Phytopathology 1996, 86, 515–522. [Google Scholar] [CrossRef]
- Jurado, M.; Vázquez, C.; Patiño, B.; González-Jaén, M.T. PCR Detection Assays for the Trichothecene-Producing Species Fusarium graminearum, Fusarium culmorum, Fusarium poae, Fusarium equiseti and Fusarium sporotrichioides. Syst. Appl. Microbiol. 2005, 28, 562–568. [Google Scholar] [CrossRef]
- Nicholson, P.; Simpson, D.R.; Weston, G.; Rezanoor, H.N.; Lees, A.K.; Parry, D.W.; Joyce, D. Detection and Quantification of Fusarium culmorum and Fusarium graminearum in Cereals Using PCR Assays. Physiol. Mol. Plant Pathol. 1998, 53, 17–37. [Google Scholar] [CrossRef]
- Mishra, P.K.; Fox, R.T.V.; Culham, A. Development of a PCR-Based Assay for Rapid and Reliable Identification of Pathogenic Fusaria. FEMS Microbiol. Lett. 2003, 218, 329–332. [Google Scholar] [CrossRef]
- Parry, D.W.; Nicholson, P. Development of a PCR Assay to Detect Fusarium poae in Wheat. Plant Pathol. 1996, 45, 383–391. [Google Scholar] [CrossRef]
- Jurado, M.; Vázquez, C.; Marín, S.; Sanchis, V.; Teresa González-Jaén, M. PCR-Based Strategy to Detect Contamination with Mycotoxigenic Fusarium Species in Maize. Syst. Appl. Microbiol. 2006, 29, 681–689. [Google Scholar] [CrossRef]
- Wilson, A.; Simpson, D.; Chandler, E.; Jennings, P.; Nicholson, P. Development of PCR Assays for the Detection and Differentiation of Fusarium sporotrichioides and Fusarium langsethiae. FEMS Microbiol. Lett. 2004, 233, 69–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Möller, E.M.; Chełkowski, J.; Geiger, H.H. Species-Specific PCR Assays for the Fungal Pathogens Fusarium moniliforme and Fusarium subglutinans and Their Application to Diagnose Maize Ear Rot Disease. J. Phytopathol. 1999, 147, 497–508. [Google Scholar] [CrossRef]
- Patiño, B.; Mirete, S.; González-Jaén, M.T.; Mulé, G.; Rodríguez, M.T.; Vázquez, C. PCR Detection Assay of Fumonisin-Producing Fusarium verticillioides Strains. J. Food Prot. 2004, 67, 1278–1283. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, P.; Lees, A.K.; Maurin, N.; Parry, D.W.; Rezanoor, H.N. Development of a PCR Assay to Identify and Quantify Microdochium nivale var. nivale and Microdochium nivale var. majus in Wheat. Physiol. Mol. Plant Pathol. 1996, 48, 257–271. [Google Scholar] [CrossRef]
- Nelson, R. International Plant Pathology: Past and Future Contributions to Global Food Security. Phytopathology 2020, 110, 245–253. [Google Scholar] [CrossRef] [Green Version]
- Cowger, C.; Smith, J.; Boos, D.; Bradley, C.A.; Ransom, J.; Bergstrom, G.C. Managing a Destructive, Episodic Crop Disease: A National Survey of Wheat and Barley Growers’ Experience with Fusarium Head Blight. Plant Dis. 2020, 104, 634–648. [Google Scholar] [CrossRef]
- Salgado, J.D.; Madden, L.V.; Paul, P.A. Efficacy and Economics of Integrating In-Field and Harvesting Strategies to Manage Fusarium Head Blight of Wheat. Plant Dis. 2014, 98, 1407–1421. [Google Scholar] [CrossRef] [Green Version]
- Blandino, M.; Haidukowski, M.; Pascale, M.; Plizzari, L.; Scudellari, D.; Reyneri, A. Integrated Strategies for the Control of Fusarium Head Blight and Deoxynivalenol Contamination in Winter Wheat. Field Crops Res. 2012, 133, 139–149. [Google Scholar] [CrossRef] [Green Version]
- McKee, G.; Cowger, C.; Dill-Macky, R.; Friskop, A.; Gautam, P.; Ransom, J.; Wilson, W. Disease Management and Estimated Effects on DON (Deoxynivalenol) Contamination in Fusarium Infested Barley. Agriculture 2019, 9, 155. [Google Scholar] [CrossRef] [Green Version]
- Mesterházy, Á. Types and Components of Resistance to Fusarium Head Blight of Wheat. Plant Breed. 1995, 114, 377–386. [Google Scholar] [CrossRef]
- Steiner, B.; Buerstmayr, M.; Michel, S.; Schweiger, W.; Lemmens, M.; Buerstmayr, H. Breeding Strategies and Advances in Line Selection for Fusarium Head Blight Resistance in Wheat. Trop. Plant Pathol. 2017, 42, 165–174. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, H.W.; Christensen, J.J. Factors Affecting Resistance of Wheat to Scab Caused by Gibberella zeae. Phytopathology 1963, 53, 831–838. [Google Scholar]
- Mesterházy, Á.; Bartók, T.; Mirocha, C.G.; Komoróczy, R. Nature of Wheat Resistance to Fusarium Head Blight and the Role of Deoxynivalenol for Breeding. Plant Breed. 1999, 118, 97–110. [Google Scholar] [CrossRef]
- Bai, G.; Su, Z.; Cai, J. Wheat Resistance to Fusarium Head Blight. Can. J. Plant Pathol. 2018, 40, 336–346. [Google Scholar] [CrossRef]
- Cuthbert, P.A.; Somers, D.J.; Thomas, J.; Cloutier, S.; Brulé-Babel, A. Fine Mapping Fhb1, a Major Gene Controlling Fusarium Head Blight Resistance in Bread Wheat (Triticum aestivum L.). Theor. Appl. Genet. 2006, 112, 1465–1472. [Google Scholar] [CrossRef]
- Cuthbert, P.A.; Somers, D.J.; Brulé-Babel, A. Mapping of Fhb2 on Chromosome 6BS: A Gene Controlling Fusarium Head Blight Field Resistance in Bread Wheat (Triticum aestivum L.). Theor. Appl. Genet. 2007, 114, 429–437. [Google Scholar] [CrossRef]
- Qi, L.L.; Pumphrey, M.O.; Friebe, B.; Chen, P.D.; Gill, B.S. Molecular Cytogenetic Characterization of Alien Introgressions with Gene Fhb3 for Resistance to Fusarium Head Blight Disease of Wheat. Theor. Appl. Genet. 2008, 117, 1155–1166. [Google Scholar] [CrossRef]
- Xue, S.; Li, G.; Jia, H.; Xu, F.; Lin, F.; Tang, M.; Wang, Y.; An, X.; Xu, H.; Zhang, L.; et al. Fine Mapping Fhb4, a Major QTL Conditioning Resistance to Fusarium Infection in Bread Wheat (Triticum aestivum L.). Theor. Appl. Genet. 2010, 121, 147–156. [Google Scholar] [CrossRef]
- Xue, S.; Xu, F.; Tang, M.; Zhou, Y.; Li, G.; An, X.; Lin, F.; Xu, H.; Jia, H.; Zhang, L.; et al. Precise Mapping Fhb5, a Major QTL Conditioning Resistance to Fusarium Infection in Bread Wheat (Triticum aestivum L.). Theor. Appl. Genet. 2011, 123, 1055–1063. [Google Scholar] [CrossRef]
- Cainong, J.C.; Bockus, W.W.; Feng, Y.; Chen, P.; Qi, L.; Sehgal, S.K.; Danilova, T.V.; Koo, D.H.; Friebe, B.; Gill, B.S. Chromosome Engineering, Mapping, and Transferring of Resistance to Fusarium Head Blight Disease from Elymus tsukushiensis into Wheat. Theor. Appl. Genet. 2015, 128, 1019–1027. [Google Scholar] [CrossRef]
- Guo, J.; Zhang, X.; Hou, Y.; Cai, J.; Shen, X.; Zhou, T.; Xu, H.; Ohm, H.W.; Wang, H.; Li, A.; et al. High-Density Mapping of the Major FHB Resistance Gene Fhb7 Derived from Thinopyrum ponticum and Its Pyramiding with Fhb1 by Marker-Assisted Selection. Theor. Appl. Genet. 2015, 128, 2301–2316. [Google Scholar] [CrossRef]
- Wegulo, S.N.; Baenziger, P.S.; Hernandez Nopsa, J.; Bockus, W.W.; Hallen-Adams, H. Management of Fusarium Head Blight of Wheat and Barley. Crop Prot. 2015, 73, 100–107. [Google Scholar] [CrossRef]
- Palazzini, J.M.; Groenenboom-de Haas, B.H.; Torres, A.M.; Köhl, J.; Chulze, S.N. Biocontrol and Population Dynamics of Fusarium spp. on Wheat Stubble in Argentina. Plant Pathol. 2013, 62, 859–866. [Google Scholar] [CrossRef]
- Comby, M.; Gacoin, M.; Robineau, M.; Rabenoelina, F.; Ptas, S.; Dupont, J.; Profizi, C.; Baillieul, F. Screening of Wheat Endophytes as Biological Control Agents against Fusarium Head Blight Using Two Different in vitro Tests. Microbiol. Res. 2017, 202, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.I.; Schisler, D.A.; Boehm, M.J.; Slininger, P.J.; Bothast, R.J. Selection and Evaluation of Microorganisms for Biocontrol of Fusarium Head Blight of Wheat Incited by Gibberella zeae. Plant Dis. 2001, 85, 1253–1258. [Google Scholar] [CrossRef] [Green Version]
- Khan, N.I.; Schisler, D.A.; Boehm, M.J.; Lipps, P.E.; Slininger, P.J. Field Testing of Antagonists of Fusarium Head Blight Incited by Gibberella zeae. Biol. Control 2004, 29, 245–255. [Google Scholar] [CrossRef]
- Schisler, D.A.; Khan, N.I.; Boehm, M.J.; Slininger, P.J. Greenhouse and Field Evaluation of Biological Control of Fusarium Head Blight on Durum Wheat. Plant Dis. 2002, 86, 1350–1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Zhu, Y.; Ma, D.; Xu, W.; Zhou, J.; Yan, H.; Yang, L.; Yin, J. Screening, Identification, and Optimization of Fermentation Conditions of an Antagonistic Endophyte to Wheat Head Blight. Agronomy 2019, 9, 476. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Selvaraj, J.N.; Xing, F.; Zhou, L.; Wang, Y.; Song, H.; Tan, X.; Sun, L.; Sangare, L.; Folly, Y.M.E.; et al. Antagonistic Action of Bacillus subtilis Strain SG6 on Fusarium graminearum. PLoS ONE 2014, 9, e92486. [Google Scholar] [CrossRef] [PubMed]
- Zachetti, V.G.L.; Cendoya, E.; Nichea, M.J.; Chulze, S.N.; Ramirez, M.L. Preliminary Study on the Use of Chitosan as an Eco-Friendly Alternative to Control Fusarium Growth and Mycotoxin Production on Maize and Wheat. Pathogens 2019, 8, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.R.; Doohan, F.M. Comparison of the Efficacy of Chitosan with That of a Fluorescent Pseudomonad for the Control of Fusarium Head Blight Disease of Cereals and Associated Mycotoxin Contamination of Grain. Biol. Control 2009, 48, 48–54. [Google Scholar] [CrossRef]
- Drakopoulos, D.; Luz, C.; Torrijos, R.; Meca, G.; Weber, P.; Bänziger, I.; Voegele, R.T.; Six, J.; Vogelgsang, S. Use of Botanicals to Suppress Different Stages of the Life Cycle of Fusarium graminearum. Phytopathology 2019, 109, 2116–2123. [Google Scholar] [CrossRef] [Green Version]
- Wegulo, S.N.; Bockus, W.W.; Nopsa, J.H.; de Wolf, E.D.; Eskridge, K.M.; Peiris, K.H.S.; Dowell, F.E. Effects of Integrating Cultivar Resistance and Fungicide Application on Fusarium Head Blight and Deoxynivalenol in Winter Wheat. Plant Dis. 2011, 95, 554–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Angelo, D.L.; Bradley, C.A.; Ames, K.A.; Willyerd, K.T.; Madden, L.V.; Paul, P.A. Efficacy of Fungicide Applications during and after Anthesis against Fusarium Head Blight and Deoxynivalenol in Soft Red Winter Wheat. Plant Dis. 2014, 98, 1387–1397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, P.A.; Bradley, C.A.; Madden, L.V.; Dalla Lana, F.; Bergstrom, G.C.; Dill-Macky, R.; Esker, P.D.; Wise, K.A.; McMullen, M.; Grybauskas, A.; et al. Meta-Analysis of the Effects of QoI and DMI Fungicide Combinations on Fusarium Head Blight and Deoxynivalenol in Wheat. Plant Dis. 2018, 102, 2602–2615. [Google Scholar] [CrossRef] [Green Version]
- Lehoczki-Krsjak, S.; Varga, M.; Mesterházy, Á. Distribution of Prothioconazole and Tebuconazole between Wheat Ears and Flag Leaves Following Fungicide Spraying with Different Nozzle Types at Flowering. Pest. Manag. Sci. 2015, 71, 105–113. [Google Scholar] [CrossRef]
- Mesterházy, Á.; Tóth, B.; Varga, M.; Bartók, T.; Szabó-Hevér, Á.; Farády, L.; Lehoczki-Krsjak, S. Role of Fungicides, Application of Nozzle Types, and the Resistance Level of Wheat Varieties in the Control of Fusarium Head Blight and Deoxynivalenol. Toxins 2011, 3, 1453–1483. [Google Scholar] [CrossRef] [Green Version]
- Behmann, J.; Mahlein, A.K.; Rumpf, T.; Römer, C.; Plümer, L. A Review of Advanced Machine Learning Methods for the Detection of Biotic Stress in Precision Crop Protection. Precis. Agric. 2015, 16, 239–260. [Google Scholar] [CrossRef]
- Bauriegel, E.; Giebel, A.; Geyer, M.; Schmidt, U.; Herppich, W.B. Early Detection of Fusarium Infection in Wheat Using Hyper-Spectral Imaging. Comput. Electron. Agric. 2011, 75, 304–312. [Google Scholar] [CrossRef]
- Lancashire, P.D.; Bleiholder, H.; van den Boom, T.; Langelüddeke, P.; Stauss, R.; Weber, E.; Witzenberger, A. A Uniform Decimal Code for Growth Stages of Crops and Weeds. Ann. Appl. Biol. 1991, 119, 561–601. [Google Scholar] [CrossRef]
- Menesatti, P.; Antonucci, F.; Pallottino, F.; Giorgi, S.; Matere, A.; Nocente, F.; Pasquini, M.; D’Egidio, M.G.; Costa, C. Laboratory vs. in-Field Spectral Proximal Sensing for Early Detection of Fusarium Head Blight Infection in Durum Wheat. Biosyst. Eng. 2013, 114, 289–293. [Google Scholar] [CrossRef]
- Femenias, A.; Gatius, F.; Ramos, A.J.; Sanchis, V.; Marín, S. Use of Hyperspectral Imaging as a Tool for Fusarium and Deoxynivalenol Risk Management in Cereals: A Review. Food Control 2020, 108, 106819. [Google Scholar] [CrossRef]
- Ma, H.; Huang, W.; Jing, Y.; Pignatti, S.; Laneve, G.; Dong, Y.; Ye, H.; Liu, L.; Guo, A.; Jiang, J. Identification of Fusarium Head Blight in Winter Wheat Ears Using Continuous Wavelet Analysis. Sensors 2020, 20, 20. [Google Scholar] [CrossRef] [Green Version]
- Jin, X.; Jie, L.; Wang, S.; Qi, H.J.; Li, S.W. Classifying Wheat Hyperspectral Pixels of Healthy Heads and Fusarium Head Blight Disease Using a Deep Neural Network in the Wild Field. Remote Sens. 2018, 10, 395. [Google Scholar] [CrossRef] [Green Version]
- Qiu, R.; Yang, C.; Moghimi, A.; Zhang, M.; Steffenson, B.J.; Hirsch, C.D. Detection of Fusarium Head Blight in Wheat Using a Deep Neural Network and Color Imaging. Remote Sens. 2019, 11, 2658. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.Y.; Chen, G.; Yin, X.; Hu, R.J.; Gu, C.Y.; Pan, Z.G.; Zhou, X.G.; Chen, Y. Integrating Spectral and Image Data to Detect Fusarium Head Blight of Wheat. Comput. Electron. Agric. 2020, 175, 105588. [Google Scholar] [CrossRef]
- Zhang, H.; Huang, L.; Huang, W.; Dong, Y.; Weng, S.; Zhao, J.; Ma, H.; Liu, L. Detection of Wheat Fusarium Head Blight Using UAV-Based Spectral and Image Feature Fusion. Front. Plant Sci. 2022, 13, 1004427. [Google Scholar] [CrossRef]
- Liu, L.; Dong, Y.; Huang, W.; Du, X.; Ma, H. Monitoring Wheat Fusarium Head Blight Using Unmanned Aerial Vehicle Hyperspectral Imagery. Remote Sens. 2020, 12, 3811. [Google Scholar] [CrossRef]
- Ma, H.; Huang, W.; Dong, Y.; Liu, L.; Guo, A. Using UAV-Based Hyperspectral Imagery to Detect Winter Wheat Fusarium Head Blight. Remote Sens. 2021, 13, 3024. [Google Scholar] [CrossRef]
- Oerke, E.C.; Steiner, U.; Dehne, H.W.; Lindenthal, M. Thermal Imaging of Cucumber Leaves Affected by Downy Mildew and Environmental Conditions. J. Exp. Bot. 2006, 57, 2121–2132. [Google Scholar] [CrossRef]
- Murchie, E.H.; Lawson, T. Chlorophyll Fluorescence Analysis: A Guide to Good Practice and Understanding Some New Applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, N.R. Chlorophyll Fluorescence: A Probe of Photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauriegel, E.; Giebel, A.; Herppich, W.B. Hyperspectral and Chlorophyll Fluorescence Imaging to Analyse the Impact of Fusarium culmorum on the Photosynthetic Integrity of Infected Wheat Ears. Sensors 2011, 11, 3765–3779. [Google Scholar] [CrossRef] [Green Version]
- Bauriegel, E.; Herppich, W.B. Hyperspectral and Chlorophyll Fluorescence Imaging for Early Detection of Plant Diseases, with Special Reference to Fusarium Spec. Infections on Wheat. Agriculture 2014, 4, 32–57. [Google Scholar] [CrossRef] [Green Version]
- Bock, C.H.; Barbedo, J.G.A.; del Ponte, E.M.; Bohnenkamp, D.; Mahlein, A.K. From Visual Estimates to Fully Automated Sensor-Based Measurements of Plant Disease Severity: Status and Challenges for Improving Accuracy. Phytopathol. Res. 2020, 2, 9. [Google Scholar] [CrossRef] [Green Version]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alisaac, E.; Mahlein, A.-K. Fusarium Head Blight on Wheat: Biology, Modern Detection and Diagnosis and Integrated Disease Management. Toxins 2023, 15, 192. https://doi.org/10.3390/toxins15030192
Alisaac E, Mahlein A-K. Fusarium Head Blight on Wheat: Biology, Modern Detection and Diagnosis and Integrated Disease Management. Toxins. 2023; 15(3):192. https://doi.org/10.3390/toxins15030192
Chicago/Turabian StyleAlisaac, Elias, and Anne-Katrin Mahlein. 2023. "Fusarium Head Blight on Wheat: Biology, Modern Detection and Diagnosis and Integrated Disease Management" Toxins 15, no. 3: 192. https://doi.org/10.3390/toxins15030192
APA StyleAlisaac, E., & Mahlein, A. -K. (2023). Fusarium Head Blight on Wheat: Biology, Modern Detection and Diagnosis and Integrated Disease Management. Toxins, 15(3), 192. https://doi.org/10.3390/toxins15030192