The Mechanism of Ochratoxin Contamination of Artificially Inoculated Licorice Roots



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
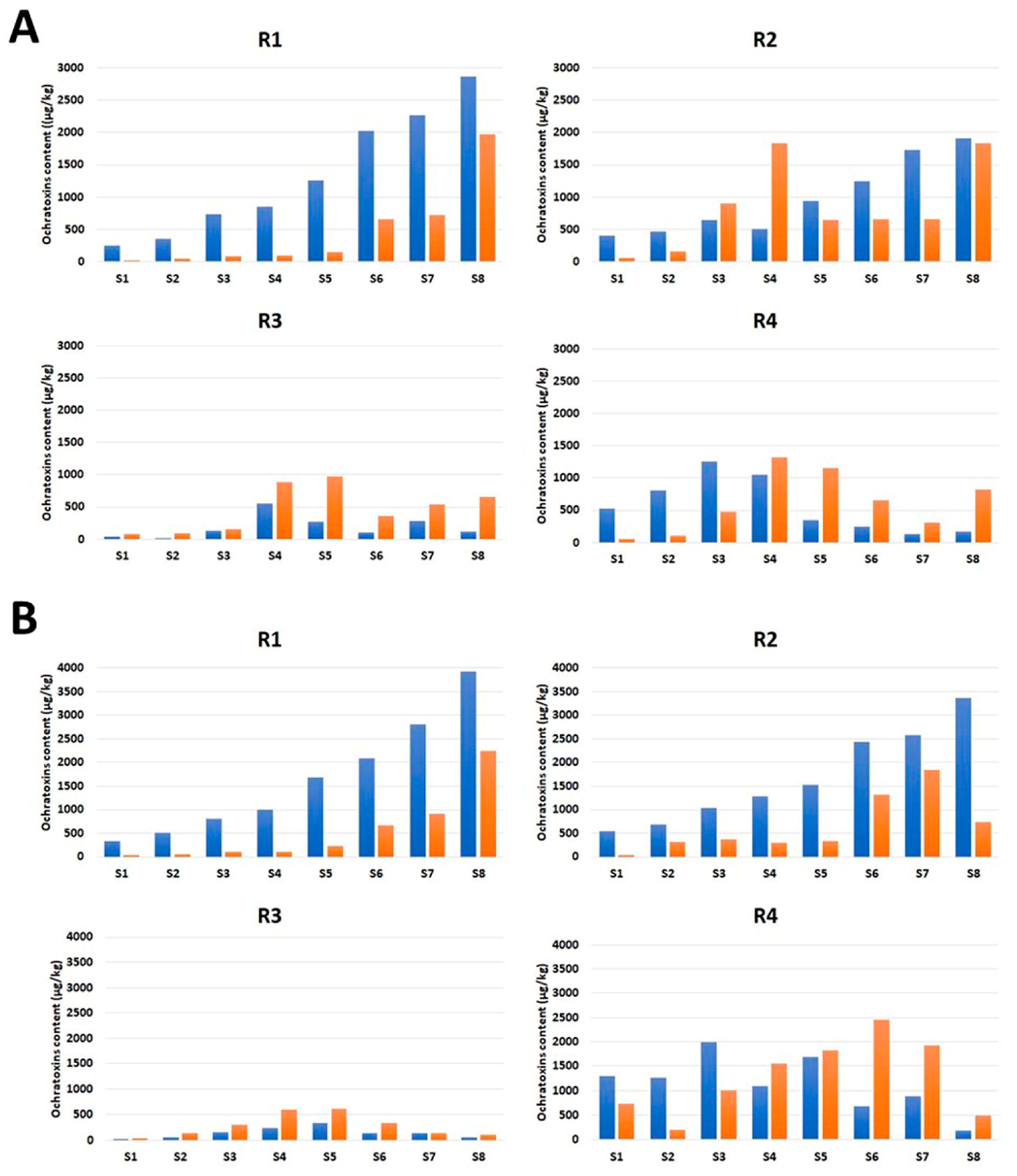
2.1. Susceptibility of Different Licorice Root Parts to OT Contamination
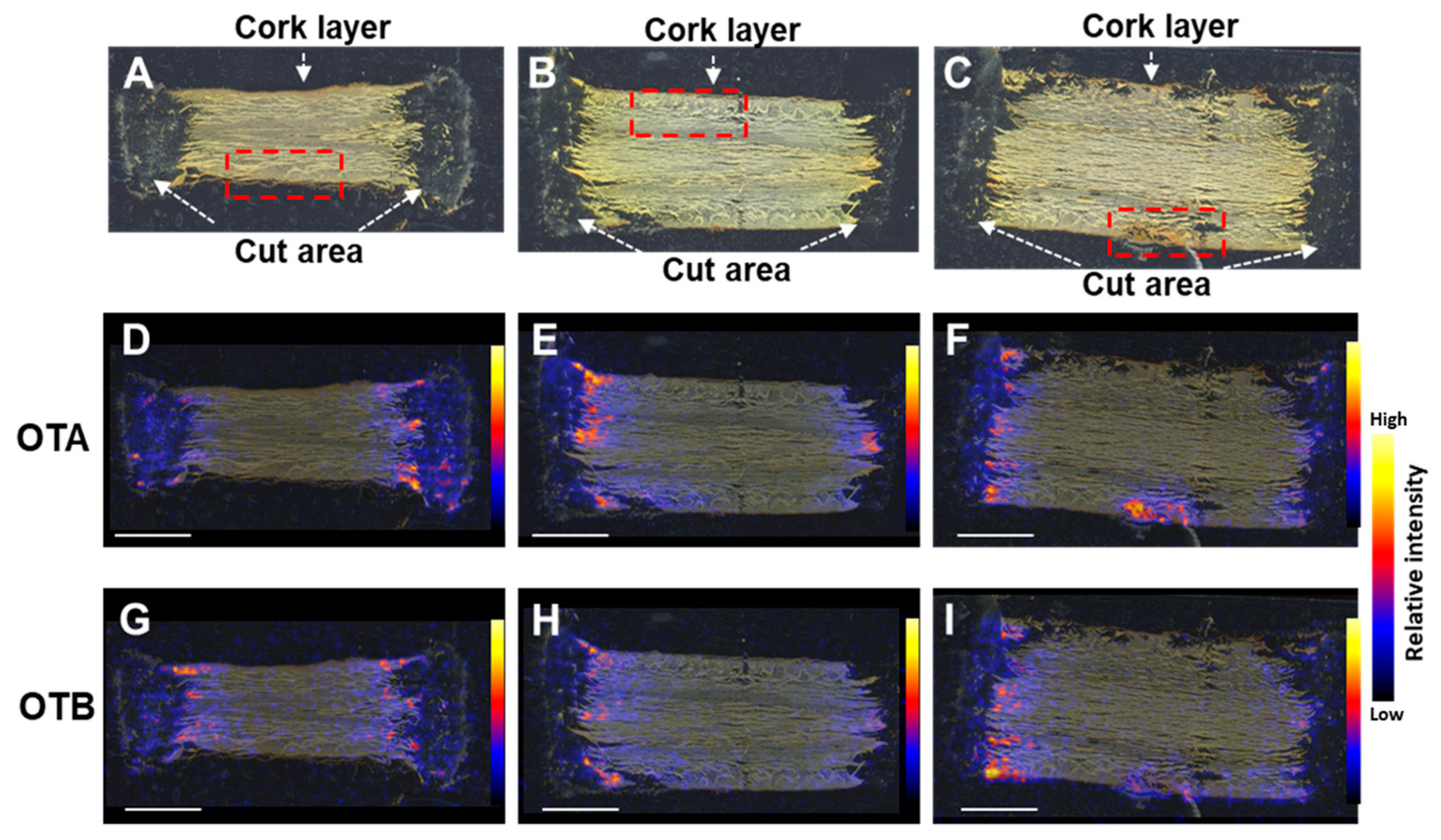
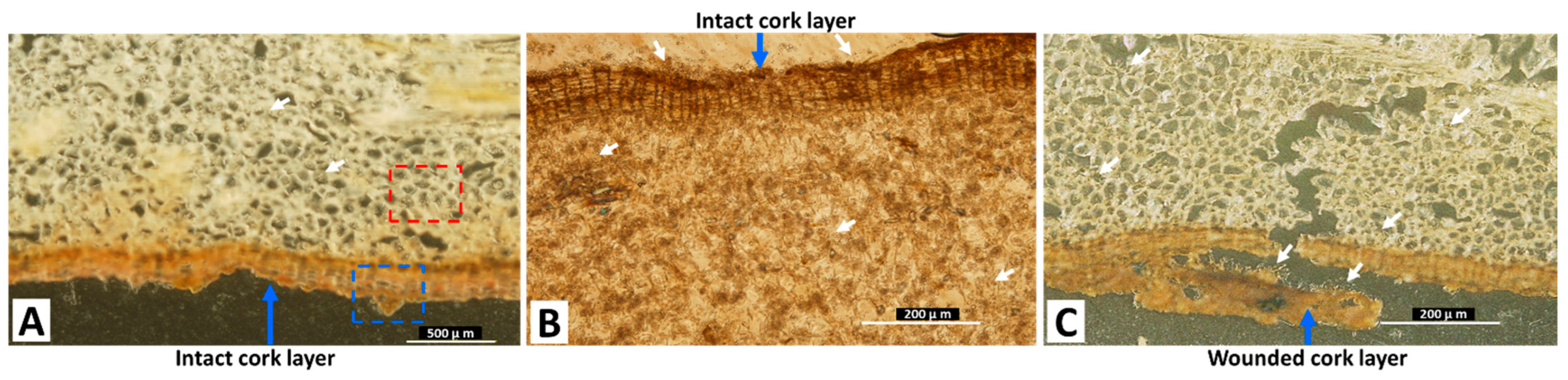
2.2. OT Localization in Contaminated Licorice Roots
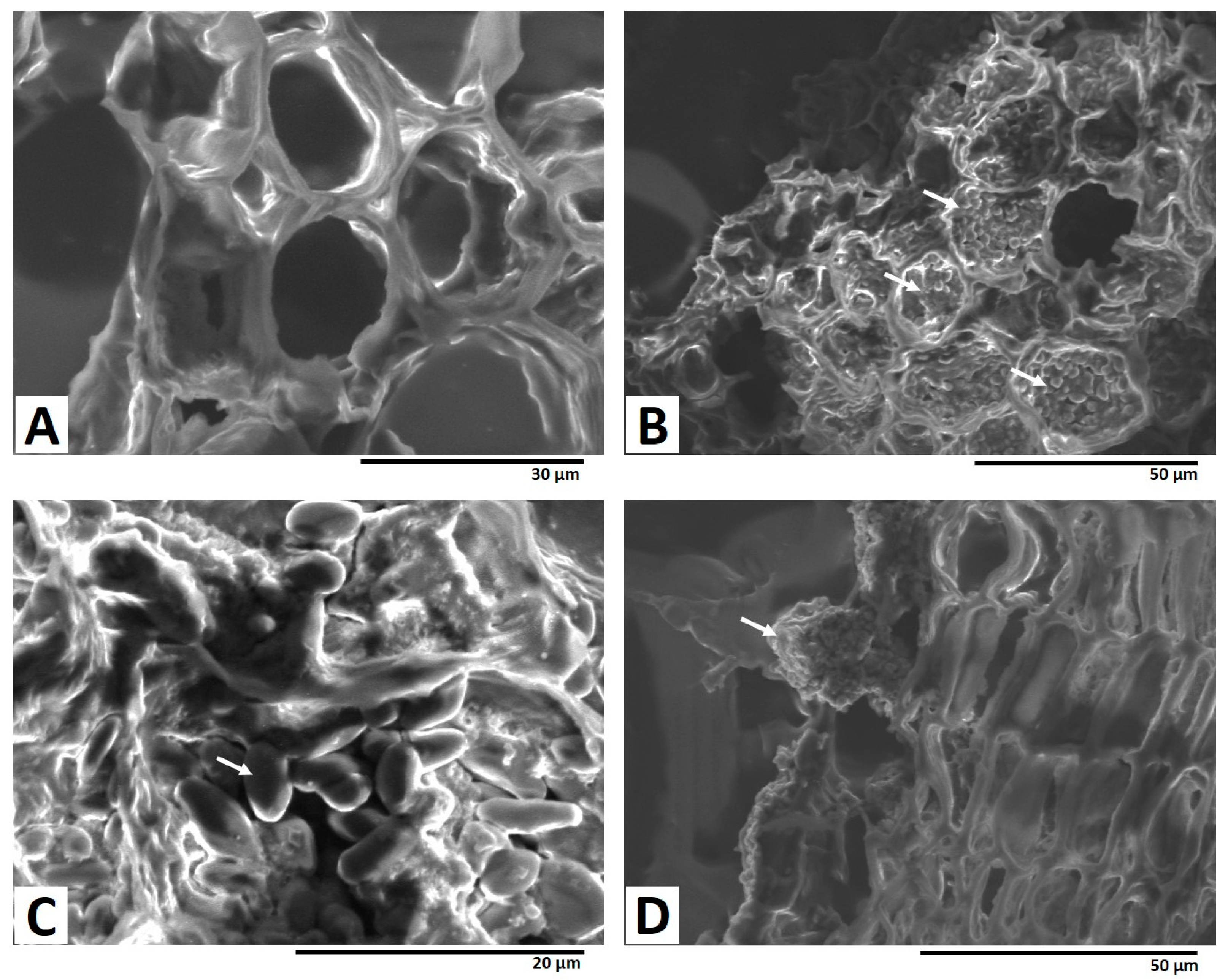
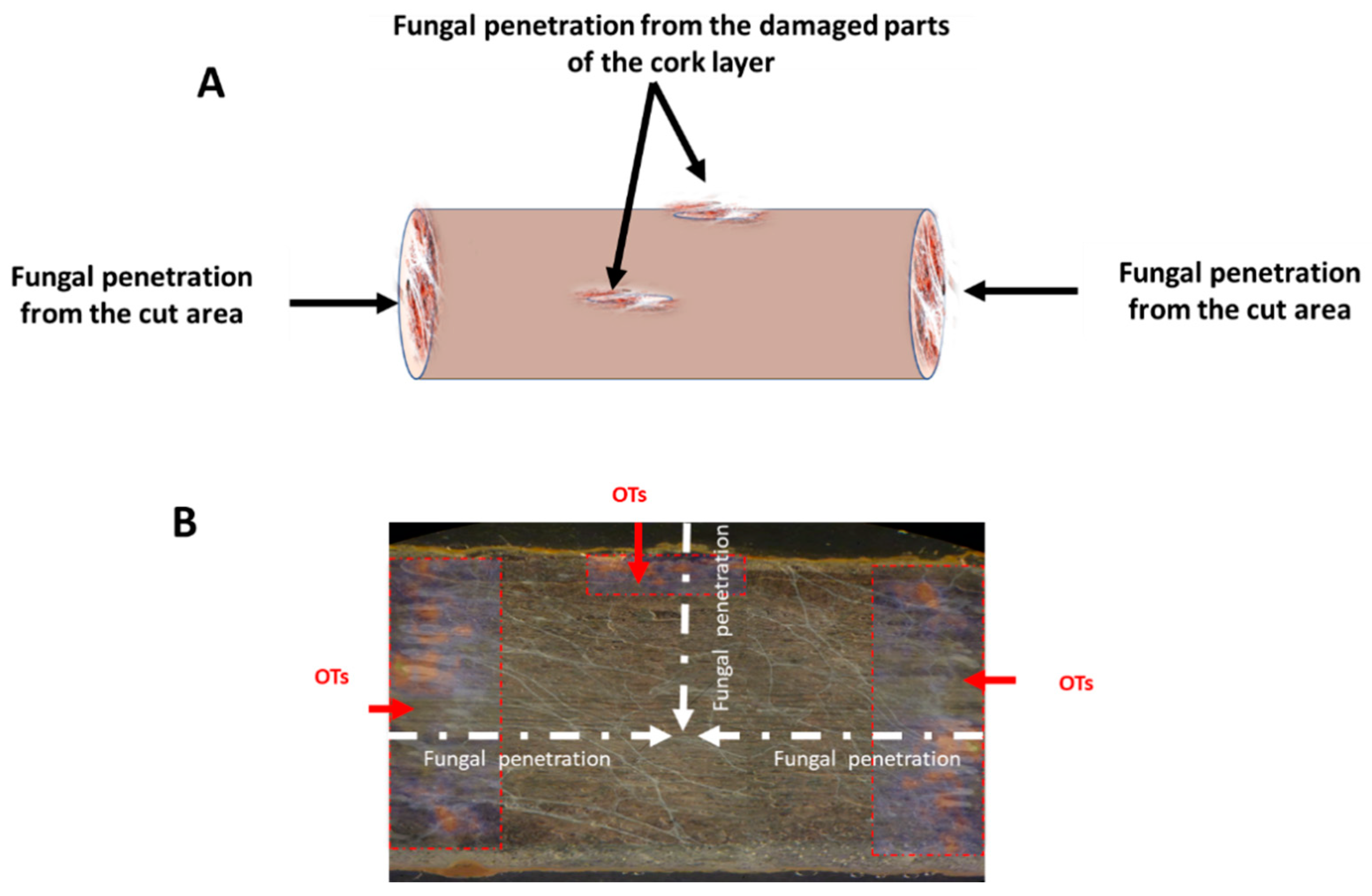
2.3. Path of Fungal Mycelial Penetration of Licorice Root
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. OT Productivity of A. Westerdijkiae Strain NIHS-7241 Isolated from Licorice
5.2. Sample Preparation
5.3. OT Quantification by HPLC
5.4. Thin Section Preparation
5.5. DESI-MS/MSI
5.6. Light Microscope (LM) and Scanning Electron Microscopy (SEM)
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pallarés, N.; Berrada, H.; Font, G.; Ferrer, E. Mycotoxins occurrence in medicinal herbs dietary supplements and exposure assessment. J. Food Sci. Technol. 2022, 59, 2830–2841. [Google Scholar]
- Ałtyn, I.; Twarużek, M. Review: Mycotoxin contamination concerns of herbs and medicinal plants. Toxins 2020, 12, 182. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Guo, W.; Zheng, Y.; Zhou, J.; Liu, T.; Chen, W.; Liang, D.; Zhao, M.; Zhu, Y.; Wu, Q.; et al. Occurrence and characterization of fungi and mycotoxins in contaminated medicinal herbs. Toxins 2020, 12, 30. [Google Scholar] [CrossRef] [Green Version]
- Reda, F.M.; El-Saadony, M.T.; El-Rayes, T.K.; Farahat, M.; Attia, G.; Alagawany, M. Dietary effect of licorice (Glycyrrhiza glabra) on quail performance, carcass, blood metabolites and intestinal microbiota. Poult Sci. 2021, 100, 101266. [Google Scholar]
- Cerulli, A.; Masullo, M.; Montoro, P.; Piacent, S. Licorice (Glycyrrhiza glabra, G. uralensis, and G. inflata) and their constituents as active cosmeceutical ingredients. Cosmetics 2022, 9, 7. [Google Scholar] [CrossRef]
- Chen, A.J.; Huang, L.F.; Wang, L.Z.; Tang, D.; Cai, F.; Gao, W.W. Occurrence of toxigenic fungi in ochratoxin A contaminated liquorice root. Food Addit. Contam. 2011, 28, 1091–1097. [Google Scholar]
- Kitagawa, I. Licorice root. A natural sweetener and an important ingredient in Chinese medicine. Pure Appl. Chem. 2002, 74, 1189–1198. [Google Scholar]
- Su, C.; Hu, Y.; Gao, D.; Luo, Y.I.; Chen, A.J.; Jiao, X.; Gao, W. Occurrence of toxigenic fungi and mycotoxins on root herbs from Chinese markets. J. Food Prot. 2018, 81, 754–761. [Google Scholar] [CrossRef]
- Pickova, D.; Ostry, V.; Malir, J.; Toman, J.; Malir, F. R review on mycotoxins and microfungi in spices in the light of the last five years. Toxins 2020, 12, 789. [Google Scholar] [CrossRef]
- Gil-Serna, J.; Patiño, B.; Cortes, L.; Gonzalez-Jaen, M.T.; Vazquez, C. Aspergillus steynii and Aspergillus westerdijkiae as potential risk of OTA contamination in food products in warm climates. Food Microbiol. 2015, 46, 168–175. [Google Scholar]
- Wakhungu, C.N.; Okoth, S.; Wachira, P.; Otieno, N.A. Mycotoxins contaminating herbs and spices in Africa: A review. Afr. J. Bio. Sci. 2021, 3, 10–28. [Google Scholar]
- Lerda, D.; Ambrosio, M.; Kunsagi, Z.; Stroka, J. Determination of ochratoxin A in licorice and licorice extracts by high-performance liquid chromatography coupled with fluorescence detection: Collaborative study. J. AOAC Int. 2013, 96, 331–340. [Google Scholar]
- Awuchi, C.G.; Ondari, E.N.; Ogbonna, C.U.; Upadhyay, A.K.; Baran, K.; Okpala, C.O.R.; Korzeniowska, M.; Guiné, R.P.F. Mycotoxins affecting animals, foods, humans and plants: Types, occurrence, toxicities, action mechanisms, prevention and detoxification strategies—A revisit. Foods 2021, 10, 1279. [Google Scholar]
- Chen, A.J.; Tang, D.; Zhou, Y.Q.; Sun, B.D.; Li, X.J.; Wang, L.Z.; Gao, W.W. Identification of ochratoxin A producing fungi associated with fresh and dry liquorice. PLoS ONE 2013, 8, e78285. [Google Scholar]
- Schrenk, D.; Bodin, L.; Chipman, J.K.; D-Mazoc, J.; Grasl-Kraupp, B.; Hogstrand, C.; Hoogenboom, L.; Leblanc, J.-C.; Nebbia, C.S.; Nielsen, E.; et al. Risk assessment of ochratoxin A in food. EFSA 2020, 18, 6113. [Google Scholar]
- Pietri, A.; Rastelli, S.; Terenzio Bertuzzi, T. Ochratoxin A and aflatoxins in liquorice products. Toxins 2010, 2, 758–770. [Google Scholar] [CrossRef] [Green Version]
- Bresch, H.; Urbanek, M.; Nusser, M. Ochratoxin A in food containing liquorice. Nahrung 2000, 44, 276–278. [Google Scholar]
- Arino, A.; Herrera, M.; Estopanan, G.; Juan, T. High levels of ochratoxin A in licorice and derived products. Int. J. Food Microbiol. 2007, 114, 366–369. [Google Scholar]
- Louppis, A.P.; Constantinou, M.S. Application of a validated method for the identification and quantification of mycotoxins in wines using UPLC-MS/MS. Separations 2022, 9, 102. [Google Scholar]
- Scott, P.M.; Truckses, M.W. Application of immunoaffinity columns to mycotoxin analysis. J. AOAC Int. 1997, 80, 941–949. [Google Scholar]
- Elamin, A.; Sakuda, S. Evaluation of the susceptibility of Ziziphus jujuba var. spinosa fruit to aflatoxin contamination and infection of aflatoxigenic fungus based on ripening stages and fruit parts. JSM Mycotoxins 2021, 71, 63–67. [Google Scholar]
- Hickert, S.; Cramer, B.; Letzel, M.C.; Humpf, H.-U. Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry imaging of ochratoxin A and fumonisins in mold-infected food. Rapid Commun. Mass Spectrom. 2016, 30, 2508–2516. [Google Scholar]
- Heussner, A.H.; Bingle, L.E.H. Comparative ochratoxin toxicity: A review of the available data. Toxins 2015, 7, 4253–4282. [Google Scholar]
- Davis, N.; Searcy, J.; Diener, U. Production of ochratoxin A by Aspergillus ochraceus in a semisynthetic medium. Appl. Microbiol. 1969, 17, 742–744. [Google Scholar]
- Wongworapat, K.; Ho, M.H.T.; Soontornjanagit, M.; Kawamura, O. Occurrence of ochratoxin A and ochratoxin B in commercial coffee in Vietnam and Thailand. JSM Mycotoxins 2016, 66, 1–6. [Google Scholar]
- Remiro, R.; Ibáñez-Vea, M.; González-Peñas, E.; Lizarraga, E. Validation of a liquid chromatography method for the simultaneous quantification of ochratoxin A and its analogues in red wines. J. Chromatogr. A. 2010, 1217, 8249–8256. [Google Scholar]
- Parussoloa, G.; Oliveirab, M.S.; Garciaa, M.V.; Bernardia, A.O.; Lemosa, J.G.; Stefanelloa, A.; Mallmannb, C.A.; Copetti, M.V. Ochratoxin A production by Aspergillus westerdijkiae in Italian-type salami. Food Microbiol. 2019, 83, 134–140. [Google Scholar]
- Coton, M.; Auffret, A.; Poirier, E.; Debaets, S.; Coton, E.; Dantigny, P. Production and migration of ochratoxin A and citrinin in Comté cheese by an isolate of Penicillium verrucosum selected among Penicillium spp. mycotoxin producers in YES medium. Food Microbiol. 2019, 82, 551–559. [Google Scholar]
- Szulc, J.; Ruman, T. Laser ablation remote-electrospray ionisation mass spectrometry (LARESI MSI) imaging-new method for detection and spatial localization of metabolites and mycotoxins produced by moulds. Toxins 2020, 12, 720. [Google Scholar] [CrossRef]
- Bani, M.; Pérez-De-Luque, A.; Rubiales, D.; Rispail, N. Physical and chemical barriers in root tissues contribute to quantitative resistance to Fusarium oxysporum f. sp. pisi in Pea. Front. Plant Sci. 2018, 9, 199. [Google Scholar] [CrossRef]
- Fatima, A.; Gupta, V.K.; Luqman, S.; Negi, A.S.; Kumar, J.K.; Shanker, K.; Saikia, D.; Srivastava, S.; Darokar, M.P.; Suman, P.S.; et al. Antifungal activity of Glycyrrhiza glabra extracts and its active constituent glabridin. Phytother Res. 2009, 23, 1190–1193. [Google Scholar] [CrossRef]
- Shuping, D.S.S.; Eloff, J.N. The use of plants to protect plants and food against fungal pathogens: A review. Afr J Tradit Complement Altern Med. 2017, 14, 120–127. [Google Scholar] [CrossRef] [Green Version]
- Mendgen, K.; Hahn, M.; Deising, H. Morphogenesis and mechanisms of penetration by plant pathogenic fungi. Annu. Rev. Phytopathol. 1996, 34, 364–386. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, R.T. Review: Cork development: What lies within. Plants 2022, 11, 2671. [Google Scholar] [CrossRef]
- Enomoto, H. Adhesive film applications help to prepare strawberry fruit sections for desorption electrospray ionization-mass spectrometry imaging. Biosci. Biotechnol. Biochem. 2021, 85, 1341–1347. [Google Scholar] [CrossRef]
- Enomoto, H. Distribution analysis of jasmonic acid-related compounds in developing Glycine max L. (soybean) seeds using mass spectrometry imaging and liquid chromatography-mass spectrometry. Phytochem. Anal. 2022, 33, 194–203. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K.; Mega, X. Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Frank, J.M.; Houbraken, J.A.M.P.; Kuijpers, A.F.A.; Samson, R.A. New ochratoxin A producing species of Aspergillus section Circumdati. Stud. Mycol. 2004, 50, 23–43. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elamin, A.; Enomoto, H.; Watanabe, M.; Sakuda, S. The Mechanism of Ochratoxin Contamination of Artificially Inoculated Licorice Roots. Toxins 2023, 15, 219. https://doi.org/10.3390/toxins15030219
Elamin A, Enomoto H, Watanabe M, Sakuda S. The Mechanism of Ochratoxin Contamination of Artificially Inoculated Licorice Roots. Toxins. 2023; 15(3):219. https://doi.org/10.3390/toxins15030219
Chicago/Turabian StyleElamin, Abdelrahman, Hirofumi Enomoto, Maiko Watanabe, and Shohei Sakuda. 2023. "The Mechanism of Ochratoxin Contamination of Artificially Inoculated Licorice Roots" Toxins 15, no. 3: 219. https://doi.org/10.3390/toxins15030219
APA StyleElamin, A., Enomoto, H., Watanabe, M., & Sakuda, S. (2023). The Mechanism of Ochratoxin Contamination of Artificially Inoculated Licorice Roots. Toxins, 15(3), 219. https://doi.org/10.3390/toxins15030219