Freshwater Cyanobacterial Toxins, Cyanopeptides and Neurodegenerative Diseases


 , ,
, ,
Abstract
:1. Introduction
2. Cyanobacterial Toxins
2.1. Microcystins (MC) Family
2.2. BMAA (β-N-methylamino-l-alanine) and Isomers
2.3. Other Cyanobacterial Neurotoxins
2.4. Cyanopeptides
2.5. Chronic Effects of Cyanobacterial Toxins
2.6. Stability of Cyanotoxins
2.7. Current Cyanotoxins Analytical Methods
3. Toxin Exposure Pathways
3.1. Transport of Cyanotoxins in Freshwater and Marine Systems
3.2. Toxin Exposure Pathways: Oral (Drinking Water)
3.3. Toxin Exposure Pathways: Oral (Food)
3.4. Toxin Exposure Pathways: Air (Aerosolization)
3.5. Natural Model of Toxin Exposure
3.6. Cyanotoxins and Infections
4. Mechanisms of Brain Toxicity
4.1. Neurodevelopmental Effects
4.2. Blood-Brain Barrier (BBB)
4.3. Glia
5. Cyanotoxins, Cyanopeptides and Neurodegenerative Diseases
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murray, S.A.; Mihali, T.K.; Neilan, B.A. Extraordinary conservation, gene loss, and positive selection in the evolution of an ancient neurotoxin. Mol. Biol. Evol. 2011, 28, 1173–1182. [Google Scholar] [CrossRef] [Green Version]
- Chorus, I.; Welker, M. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management, 2nd ed.; Taylor & Francis: Boca Raton, FL, USA, 2021. [Google Scholar] [CrossRef]
- Kozlowsky-Suzuki, B.; Wilson, A.E.; da Silva Ferrao-Filho, A. Biomagnification or biodilution of microcystins in aquatic foodwebs? Meta-analyses of laboratory and field studies. Harmful Algae 2012, 18, 47–55. [Google Scholar] [CrossRef]
- Pybus, M.; Hobson, D.; Onderka, D. Mass mortality of bats due to probable blue-green algal toxicity. J. Wildl. Dis. 1986, 22, 449–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahmood, N.A.; Carmichael, W.W.; Pfahler, D. Anticholinesterase poisonings in dogs from a cyanobacterial (blue-green algae) bloom dominated by Anabaena flos-aquae. Am. J. Vet. Res. 1988, 49, 500–503. [Google Scholar]
- Henriksen, P.; Carmichael, W.W.; An, J.; Moestrup, O. Detection of an anatoxin-a (s)-like anticholinesterase in natural blooms and cultures of cyanobacteria/blue-green algae from Danish lakes and in the stomach contents of poisoned birds. Toxicon 1997, 35, 901–913. [Google Scholar] [CrossRef] [PubMed]
- Mez, K.; Beattie, K.A.; Codd, G.A.; Hanselmann, K.; Hauser, B.; Naegeli, H.; Preisig, H.R. Identification of a microcystin in benthic cyanobacteria linked to cattle deaths on alpine pastures in Switzerland. Eur. J. Phycol. 1997, 32, 111–117. [Google Scholar] [CrossRef]
- Saker, M.; Thomas, A.; Norton, J. Cattle mortality attributed to the toxic cyanobacterium Cylindrospermopsis raciborskii in an outback region of north Queensland. Environ. Toxicol. 1999, 14, 179–182. [Google Scholar] [CrossRef]
- Codd, G.A.; Lindsay, J.; Young, F.M.; Morrison, L.F.; Metcalf, J.S. Harmful cyanobacteria: From mass mortalities to management measures. In Harmful Cyanobacteria; Huisman, J., Matthijs, H.C.P., Visser, P.M., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 1–23. [Google Scholar] [CrossRef]
- Miller, M.A.; Kudela, R.M.; Mekebri, A.; Crane, D.; Oates, S.C.; Tinker, M.T.; Staedler, M.; Miller, W.A.; Toy-Choutka, S.; Dominik, C.; et al. Evidence for a novel marine harmful algal bloom: Cyanotoxin (microcystin) transfer from land to sea otters. PLoS ONE 2010, 5, e12576. [Google Scholar] [CrossRef] [PubMed]
- Chatziefthimiou, A.D.; Richer, R.; Rowles, H.; Powell, J.T.; Metcalf, J.S. Cyanotoxins as a potential cause of dog poisonings in desert environments. Vet. Rec. 2014, 174, 484–485. [Google Scholar] [CrossRef]
- Danil, K.; Berman, M.; Frame, E.; Preti, A.; Fire, S.E.; Leighfield, T.; Carretta, J.; Carter, M.L.; Lefebvre, K. Marine algal toxins and their vectors in southern California cetaceans. Harmful Algae 2021, 103, 102000. [Google Scholar] [CrossRef] [PubMed]
- Rattner, B.A.; Wazniak, C.E.; Lankton, J.S.; McGowan, P.C.; Drovetski, S.V.; Egerton, T.A. Review of harmful algal bloom effects on birds with implications for avian wildlife in the Chesapeake Bay region. Harmful Algae 2022, 120, 102319. [Google Scholar] [CrossRef]
- Pouria, S.; de Andrade, A.; Barbosa, J.; Cavalcanti, R.; Barreto, V.; Ward, C.; Preiser, W.; Poon, G.K.; Neild, G.; Codd, G. Fatal microcystin intoxication in haemodialysis unit in Caruaru, Brazil. Lancet 1998, 352, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, W.W.; Azevedo, S.; An, J.S.; Molica, R.; Jochimsen, E.M.; Lau, S.; Rinehart, K.L.; Shaw, G.R.; Eaglesham, G.K. Human fatalities from cyanobacteria: Chemical and biological evidence for cyanotoxins. Environ. Health Perspect. 2001, 109, 663–668. [Google Scholar] [CrossRef]
- Briand, J.-F.; Jacquet, S.; Bernard, C.; Humbert, J.-F. Health hazards for terrestrial vertebrates from toxic cyanobacteria in surface water ecosystems. Vet. Res. 2003, 34, 361–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLellan, N.L.; Manderville, R.A. Toxic mechanisms of microcystins in mammals. Toxicol. Res. 2017, 6, 391–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, S.H.; Duivenvoorden, L.J.; Fabbro, L.D. A decision-making framework for ecological impacts associated with the accumulation of cyanotoxins (cylindrospermopsin and microcystin). Lakes Reserv. Res. Manag. 2005, 10, 25–37. [Google Scholar] [CrossRef]
- Paerl, H.W.; Huisman, J. Blooms like it hot. Science 2008, 320, 57–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moe, S.J.; Haande, S.; Couture, R.-M. Climate change, cyanobacteria blooms and ecological status of lakes: A Bayesian network approach. Ecol. Model. 2016, 337, 330–347. [Google Scholar] [CrossRef] [Green Version]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef]
- Taranu, Z.E.; Gregory-Eaves, I.; Leavitt, P.R.; Bunting, L.; Buchaca, T.; Catalan, J.; Domaizon, I.; Guilizzoni, P.; Lami, A.; McGowan, S. Acceleration of cyanobacterial dominance in north temperate-subarctic lakes during the Anthropocene. Ecol. Lett. 2015, 18, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Trout-Haney, J.V.; Wood, Z.T.; Cottingham, K.L. Presence of the cyanotoxin microcystin in arctic lakes of Southwestern Greenland. Toxins 2016, 8, 256. [Google Scholar] [CrossRef] [Green Version]
- Ho, J.C.; Michalak, A.M.; Pahlevan, N. Widespread global increase in intense lake phytoplankton blooms since the 1980s. Nature 2019, 574, 667–670. [Google Scholar] [CrossRef] [PubMed]
- Gobler, C.J. Climate change and harmful algal blooms: Insights and perspective. Harmful Algae 2020, 91, 101731. [Google Scholar] [CrossRef]
- Bowling, L.; Baker, P. Major cyanobacterial bloom in the Barwon-Darling River, Australia, in 1991, and underlying limnological conditions. Mar. Freshw. Res. 1996, 47, 643–657. [Google Scholar] [CrossRef]
- Heisler, J.; Glibert, P.M.; Burkholder, J.M.; Anderson, D.M.; Cochlan, W.; Dennison, W.C.; Dortch, Q.; Gobler, C.J.; Heil, C.A.; Humphries, E.; et al. Eutrophication and harmful algal blooms: A scientific consensus. Harmful Algae 2008, 8, 3–13. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Cyanobacterial Toxins: Microcystin-LR in Drinking-Water; No. WHO/SDE/WSH/03.04/57; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Andersson, M.; Karlsson, O.; Banack, S.A.; Brandt, I. Transfer of developmental neurotoxin β-N-methylamino-L-alanine (BMAA) via milk to nursed offspring: Studies by mass spectrometry and image analysis. Toxicol. Lett. 2016, 258, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Kulpys, J.; Paulauskas, E.; Pilipavicius, V.; Stankevicius, R. Influence of cyanobacteria Arthrospira (Spirulina) platensis biomass additives towards the body condition of lactation cows and biochemical milk indexes. Agric. Res. 2009, 7, 823–835. [Google Scholar]
- Christaki, E.; Florou-Paneri, P.; Bonos, E. Microalgae: A novel ingredient in nutrition. Int. J. Food Sci. 2011, 62, 794–799. [Google Scholar] [CrossRef]
- McGorum, B.C.; Pirie, R.S.; Glendinning, L.; McLachlan, G.; Metcalf, J.S.; Banack, S.A.; Cox, P.A.; Codd, G.A. Grazing livestock are exposed to terrestrial cyanobacteria. Vet. Res. 2015, 46, 16. [Google Scholar] [CrossRef] [Green Version]
- Andersson, M.; Karlsson, O.; Bergström, U.; Brittebo, E.B.; Brandt, I. Maternal transfer of the cyanobacterial neurotoxin β-N-methylamino-L-alanine (BMAA) via milk to suckling offspring. PLoS ONE 2013, 8, e78133. [Google Scholar] [CrossRef] [Green Version]
- Nunn, P.B. 50 years of research on α-amino-β-methylaminopropionic acid (β-methylaminoalanine). Phytochemistry 2017, 144, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.; Karlsson, O.; Brandt, I. The environmental neurotoxin β-N-methylamino-l-alanine (l-BMAA) is deposited into birds’ eggs. Ecotoxicol. Environ. Saf. 2018, 147, 720–724. [Google Scholar] [CrossRef] [PubMed]
- Puddick, J.; van Ginkel, R.; Page, C.D.; Murray, J.S.; Greenhough, H.E.; Bowater, J.; Selwood, A.I.; Wood, S.A.; Prinsep, M.R.; Truman, P. Acute toxicity of dihydroanatoxin-a from Microcoleus autumnalis in comparison to anatoxin-a. Chemosphere 2021, 263, 127937. [Google Scholar] [CrossRef]
- Sano, T.; Takagi, H.; Morrison, L.F.; Metcalf, J.S.; Codd, G.A.; Kaya, K. Leucine aminopeptidase M inhibitors, cyanostatin A and B, isolated from cyanobacterial water blooms in Scotland. Phytochemistry 2005, 66, 543–548. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, P.R.; do Amaral, S.C.; Siqueira, A.S.; Xavier, L.P.; Santos, A.V. Anabaenopeptins: What we know so far. Toxins 2021, 13, 522. [Google Scholar] [CrossRef] [PubMed]
- Bownik, A.; Adamczuk, M.; Pawlik-Skowrońska, B. Behavioral disturbances induced by cyanobacterial oligopeptides microginin-FR1, anabaenopeptin-A and microcystin-LR are associated with neuromotoric and cytotoxic changes in Brachionus calyciflorus. J. Hazard. Mater. 2022, 438, 129472. [Google Scholar] [CrossRef]
- Metcalf, J.S.; Tischbein, M.; Cox, P.A.; Stommel, E.W. Cyanotoxins and the nervous system. Toxins 2021, 13, 660. [Google Scholar] [CrossRef]
- Breinlinger, S.; Phillips, T.J.; Haram, B.N.; Mareš, J.; Martínez Yerena, J.A.; Hrouzek, P.; Sobotka, R.; Henderson, W.M.; Schmieder, P.; Williams, S.M. Hunting the eagle killer: A cyanobacterial neurotoxin causes vacuolar myelinopathy. Science 2021, 371, eaax9050. [Google Scholar] [CrossRef]
- Bouaïcha, N.; Miles, C.O.; Beach, D.G.; Labidi, Z.; Djabri, A.; Benayache, N.Y.; Nguyen-Quang, T. Structural diversity, characterization and toxicology of microcystins. Toxins 2019, 11, 714. [Google Scholar] [CrossRef] [Green Version]
- Carmichael, W.W.; Beasley, V.; Bunner, D.L.; Eloff, J.N.; Falconer, I.; Gorham, P.; Harada, K.-I.; Krishnamurthy, T.; Min-Juan, Y.; Moore, R.E.; et al. Naming of cyclic heptapeptide toxins of cyanobacteria (blue-green algae). Toxicon 1988, 26, 971–973. [Google Scholar] [CrossRef] [PubMed]
- Díez-Quijada, L.; Prieto, A.I.; Guzmán-Guillén, R.; Jos, A.; Cameán, A.M. Occurrence and toxicity of microcystin congeners other than MC-LR and MC-RR: A review. Food Chem. Toxicol. 2019, 125, 106–132. [Google Scholar] [CrossRef] [PubMed]
- MacKintosh, C.; Beattie, K.A.; Klumpp, S.; Cohen, P.; Codd, G.A. Cyanobacterial microcystin-LR is a potent and specific inhibitor of protein phosphatases 1 and 2A from both mammals and higher plants. FEBS Lett. 1990, 264, 187–192. [Google Scholar] [CrossRef] [Green Version]
- Massey, I.Y.; Yang, F.; Ding, Z.; Yang, S.; Guo, J.; Al-Osman, M.; Kamegni, R.B.; Zeng, W. Exposure routes and health effects of microcystins on animals and humans: A mini-review. Toxicon 2018, 151, 156–162. [Google Scholar] [CrossRef]
- Hu, Y.; Chen, J.; Fan, H.; Xie, P.; He, J. A review of neurotoxicity of microcystins. Environ. Sci. Pollut. Res. 2016, 23, 7211–7219. [Google Scholar] [CrossRef] [PubMed]
- Tzima, E.; Serifi, I.; Tsikari, I.; Alzualde, A.; Leonardos, I.; Papamarcaki, T. Transcriptional and behavioral responses of zebrafish larvae to microcystin-LR exposure. Int. J. Mol. Sci. 2017, 18, 365. [Google Scholar] [CrossRef] [Green Version]
- Pašková, V.; Adamovský, O.; Pikula, J.; Skočovská, B.; Band’ouchová, H.; Horáková, J.; Babica, P.; Maršálek, B.; Hilscherová, K. Detoxification and oxidative stress responses along with microcystins accumulation in Japanese quail exposed to cyanobacterial biomass. Sci. Total Environ. 2008, 398, 34–47. [Google Scholar] [CrossRef]
- Zhao, S.; Li, G.; Chen, J. A proteomic analysis of prenatal transfer of microcystin-LR induced neurotoxicity in rat offspring. J. Proteom. 2015, 114, 197–213. [Google Scholar] [CrossRef] [PubMed]
- Feurstein, D.; Stemmer, K.; Kleinteich, J.; Speicher, T.; Dietrich, D.R. Microcystin congener–and concentration-dependent induction of murine neuron apoptosis and neurite degeneration. Toxicol. Sci. 2011, 124, 424–431. [Google Scholar] [CrossRef] [Green Version]
- Cox, P.A.; Banack, S.A.; Murch, S.J.; Rasmussen, U.; Tien, G.; Bidigare, R.R.; Metcalf, J.S.; Morrison, L.F.; Codd, G.A.; Bergman, B. Diverse taxa of cyanobacteria produce β-N-methylamino-L-alanine, a neurotoxic amino acid. Proc. Natl. Acad. Sci. USA 2005, 102, 5074–5078. [Google Scholar] [CrossRef] [Green Version]
- Violi, J.P.; Mitrovic, S.M.; Colville, A.; Main, B.J.; Rodgers, K.J. Prevalence of β-methylamino-L-alanine (BMAA) and its isomers in freshwater cyanobacteria isolated from eastern Australia. Ecotoxicol. Environ. Saf. 2019, 172, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Schneider, T.; Simpson, C.; Desai, P.; Tucker, M.; Lobner, D. Neurotoxicity of isomers of the environmental toxin L-BMAA. Toxicon 2020, 184, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.M.; Bereman, M.S.; Marsden, K.C. The cyanotoxin 2, 4-DAB reduces viability and causes behavioral and molecular dysfunctions associated with neurodegeneration in larval zebrafish. Neurotox. Res. 2022, 40, 347–364. [Google Scholar] [CrossRef] [PubMed]
- Chernoff, N.; Hill, D.J.; Diggs, D.L.; Faison, B.D.; Francis, B.M.; Lang, J.R.; Larue, M.M.; Le, T.-T.; Loftin, K.A.; Lugo, J.N.; et al. A critical review of the postulated role of the non-essential amino acid, β-N-Methylamino-L-Alanine, in neurodegenerative disease in humans. J. Toxicol. Environ. Health Part B 2017, 20, 183–229. [Google Scholar] [CrossRef] [PubMed]
- Dunlop, R.A.; Banack, S.A.; Bishop, S.L.; Metcalf, J.S.; Murch, S.J.; Davis, D.A.; Stommel, E.W.; Karlsson, O.; Brittebo, E.B.; Chatziefthimiou, A.D.; et al. Is exposure to BMAA a risk factor for neurodegenerative diseases? A response to a critical review of the BMAA hypothesis. Neurotox. Res. 2021, 39, 81–106. [Google Scholar] [CrossRef]
- Sini, P.; Dang, T.B.C.; Fais, M.; Galioto, M.; Padedda, B.M.; Lugliè, A.; Iaccarino, C.; Crosio, C. Cyanobacteria, cyanotoxins, and neurodegenerative diseases: Dangerous liaisons. Inter. J. Mol. Sci. 2021, 22, 8726. [Google Scholar] [CrossRef] [PubMed]
- Courtier, A.; Potheret, D.; Giannoni, P. Environmental bacteria as triggers to brain disease: Possible mechanisms of toxicity and associated human risk. Life Sci. 2022, 304, 120689. [Google Scholar] [CrossRef]
- Faassen, E.J. Presence of the neurotoxin BMAA in aquatic ecosystems: What do we really know? Toxins 2014, 6, 1109–1138. [Google Scholar] [CrossRef] [Green Version]
- Lopicic, S.; Svirčev, Z.; Malešević, T.P.; Kopitović, A.; Ivanovska, A.; Meriluoto, J. Environmental neurotoxin β-N-Methylamino-L-alanine (BMAA) as a widely occurring putative pathogenic factor in neurodegenerative diseases. Microorganisms 2022, 10, 2418. [Google Scholar] [CrossRef]
- Nunes-Costa, D.; Magalhães, J.D.; G-Fernandes, M.; Cardoso, S.M.; Empadinhas, N. Microbial BMAA and the pathway for Parkinson’s disease neurodegeneration. Front. Aging Neurosci. 2020, 12, 26. [Google Scholar] [CrossRef] [Green Version]
- Dunlop, R.A.; Cox, P.A.; Banack, S.A.; Rodgers, K.J. The non-protein amino acid BMAA is misincorporated into human proteins in place of L-serine causing protein misfolding and aggregation. PLoS ONE 2013, 8, e75376. [Google Scholar] [CrossRef] [Green Version]
- Frøyset, A.K.; Khan, E.A.; Fladmark, K.E. Quantitative proteomics analysis of zebrafish exposed to sub-lethal dosages of β-methyl-amino-L-alanine (BMAA). Sci. Rep. 2016, 6, 29631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, P.A.; Davis, D.A.; Mash, D.C.; Metcalf, J.S.; Banack, S.A. Dietary exposure to an environmental toxin triggers neurofibrillary tangles and amyloid deposits in the brain. Proc. Royal Soc. B Biol. Sci. 2016, 283, 20152397. [Google Scholar] [CrossRef] [Green Version]
- Haase, C.; Stieler, J.; Arendt, T.; Holzer, M. Pseudophosphorylation of tau protein alters its ability for self-aggregation. J. Neurochem. 2004, 88, 1509–1520. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Beebe, K.; Nangle, L.A.; Jang, J.; Longo-Guess, C.M.; Cook, S.A.; Davisson, M.T.; Sundberg, J.P.; Schimmel, P.; Ackerman, S.L. Editing-defective tRNA synthetase causes protein misfolding and neurodegeneration. Nature 2006, 443, 50–55. [Google Scholar] [CrossRef] [PubMed]
- De Munck, E.; Munoz-Saez, E.; Miguel, B.G.; Solas, M.T.; Ojeda, I.; Martinez, A.; Gil, C.; Arahuetes, R.M. β-N-methylamino-l-alanine causes neurological and pathological phenotypes mimicking Amyotrophic Lateral Sclerosis (ALS): The first step towards an experimental model for sporadic ALS. Environ. Toxicol. Pharmacol. 2013, 36, 243–255. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Saez, E.; de Munck, E.; Arahuetes, R.M.; Solas, M.T.; Martínez, A.M.; Miguel, B.G. β-N-methylamino-L-alanine induces changes in both GSK3 and TDP-43 in human neuroblastoma. J. Toxicol. Sci. 2013, 38, 425–430. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, O.; Jiang, L.; Ersson, L.; Malmström, T.; Ilag, L.L.; Brittebo, E.B. Environmental neurotoxin interaction with proteins: Dose-dependent increase of free and protein-associated BMAA (β-N-methylamino-L-alanine) in neonatal rat brain. Sci. Rep. 2015, 5, 15570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, X.; Basile, M.; Mash, D.C. Cerebral uptake and protein incorporation of cyanobacterial toxin β-N-methylamino-L-alanine. Neuroreport 2013, 24, 779–784. [Google Scholar] [CrossRef]
- Han, N.-C.; Bullwinkle, T.J.; Loeb, K.F.; Faull, K.F.; Mohler, K.; Rinehart, J.; Ibba, M. The mechanism of β-N-methylamino-l-alanine inhibition of tRNA aminoacylation and its impact on misincorporation. J. Biol. Chem. 2020, 295, 1402–1410. [Google Scholar] [CrossRef]
- Rochet, J.-C. Errors in translation cause selective neurodegeneration. ACS Chem. Biol. 2006, 1, 562–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmichael, W.W.; Biggs, D.F.; Peterson, M.A. Pharmacology of anatoxin-a, produced by the freshwater cyanophyte Anabaena flos-aquae NRC-44-1. Toxicon 1979, 17, 229–236. [Google Scholar] [CrossRef]
- Christensen, V.G.; Khan, E. Freshwater neurotoxins and concerns for human, animal, and ecosystem health: A review of anatoxin-a and saxitoxin. Sci. Total Environ. 2020, 736, 139515. [Google Scholar] [CrossRef] [PubMed]
- Fiore, M.F.; de Lima, S.T.; Carmichael, W.W.; McKinnie, S.M.; Chekan, J.R.; Moore, B.S. Guanitoxin, re-naming a cyanobacterial organophosphate toxin. Harmful Algae 2020, 92, 101737. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, J.; Richer, R.; Cox, P.; Codd, G. Cyanotoxins in desert environments may present a risk to human health. Sci. Total Environ. 2012, 421, 118–123. [Google Scholar] [CrossRef]
- Metcalf, J.; Banack, S.; Richer, R.; Cox, P. Neurotoxic amino acids and their isomers in desert environments. J. Arid. Environ. 2015, 112, 140–144. [Google Scholar] [CrossRef]
- Backer, L.C.; Manassaram-Baptiste, D.; LePrell, R.; Bolton, B. Cyanobacteria and algae blooms: Review of health and environmental data from the harmful algal bloom-related illness surveillance system (HABISS) 2007–2011. Toxins 2015, 7, 1048–1064. [Google Scholar] [CrossRef] [Green Version]
- Carmichael, W.W.; Boyer, G.L. Health impacts from cyanobacteria harmful algae blooms: Implications for the North American Great Lakes. Harmful Algae 2016, 54, 194–212. [Google Scholar] [CrossRef] [PubMed]
- Ohtani, I.; Moore, R.E.; Runnegar, M.T. Cylindrospermopsin: A potent hepatotoxin from the blue-green alga Cylindrospermopsis raciborskii. J. Amer. Chem. Soc. 1992, 114, 7941–7942. [Google Scholar] [CrossRef]
- Rzymski, P.; Poniedziałek, B. In search of environmental role of cylindrospermopsin: A review on global distribution and ecology of its producers. Wat. Res. 2014, 66, 320–337. [Google Scholar] [CrossRef]
- Chen, G.; Wang, L.; Wang, M.; Hu, T. Comprehensive insights into the occurrence and toxicological issues of nodularins. Mar. Pollut. Bull. 2021, 162, 111884. [Google Scholar] [CrossRef]
- Lehtonen, K.K.; Kankaanpää, H.; Leiniö, S.; Sipiä, V.O.; Pflugmacher, S.; Sandberg-Kilpi, E. Accumulation of nodularin-like compounds from the cyanobacterium Nodularia spumigena and changes in acetylcholinesterase activity in the clam Macoma balthica during short-term laboratory exposure. Aquat. Toxicol. 2003, 64, 461–476. [Google Scholar] [CrossRef]
- Kankaanpää, H.; Leiniö, S.; Olin, M.; Sjövall, O.; Meriluoto, J.; Lehtonen, K.K. Accumulation and depuration of cyanobacterial toxin nodularin and biomarker responses in the mussel Mytilus edulis. Chemosphere 2007, 68, 1210–1217. [Google Scholar] [CrossRef]
- Christensen, V.G.; Olds, H.T.; Norland, J.; Khan, E. Phytoplankton community interactions and cyanotoxin mixtures in three recurring surface blooms within one lake. J. Hazard. Mater. 2022, 427, 128142. [Google Scholar] [CrossRef]
- Martin, R.M.; Stallrich, J.; Bereman, M.S. Mixture designs to investigate adverse effects upon co-exposure to environmental cyanotoxins. Toxicology 2019, 421, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.M.; Bereman, M.S.; Marsden, K.C. BMAA and MCLR interact to modulate behavior and exacerbate molecular changes related to neurodegeneration in larval zebrafish. Toxicol. Sci. 2021, 179, 251–261. [Google Scholar] [CrossRef]
- Roy-Lachapelle, A.; Duy, S.V.; Munoz, G.; Dinh, Q.T.; Bahl, E.; Simon, D.F.; Sauvé, S. Analysis of multiclass cyanotoxins (microcystins, anabaenopeptins, cylindrospermopsin and anatoxins) in lake waters using on-line SPE liquid chromatography high-resolution orbitrap mass spectrometry. Anal. Methods 2019, 11, 5289–5300. [Google Scholar] [CrossRef] [Green Version]
- Janssen, E.M.-L. Cyanobacterial peptides beyond microcystins—A review on co-occurrence, toxicity, and challenges for risk assessment. Water Res. 2019, 151, 488–499. [Google Scholar] [CrossRef] [PubMed]
- Natumi, R.; Janssen, E.M.-L. Cyanopeptide co-production dynamics beyond mirocystins and effects of growth stages and nutrient availability. Environ. Sci. Technol. 2020, 54, 6063–6072. [Google Scholar] [CrossRef] [PubMed]
- Nagarajan, M.; Maruthanayagam, V.; Sundararaman, M. SAR analysis and bioactive potentials of freshwater and terrestrial cyanobacterial compounds: A review. J. Appl. Toxicol. 2013, 33, 313–349. [Google Scholar] [CrossRef] [PubMed]
- Caban-Holt, A.; Mattingly, M.; Cooper, G.; Schmitt, F.A. Neurodegenerative memory disorders: A potential role of environmental toxins. Neurol. Clin. 2005, 23, 485–521. [Google Scholar] [CrossRef]
- Shaw, C.A.; Höglinger, G.U. Neurodegenerative diseases: Neurotoxins as sufficient etiologic agents? Neuromolecular. Med. 2008, 10, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caller, T.A.; Doolin, J.W.; Haney, J.F.; Murby, A.J.; West, K.G.; Farrar, H.E.; Ball, A.; Harris, B.T.; Stommel, E.W. A cluster of amyotrophic lateral sclerosis in New Hampshire: A possible role for toxic cyanobacteria blooms. Amyotrop. Lateral Scler. 2009, 10, 101–108. [Google Scholar] [CrossRef]
- Banack, S.A.; Caller, T.; Henegan, P.; Haney, J.; Murby, A.; Metcalf, J.S.; Powell, J.; Alan, P.; Stommel, E. Detection of cyanotoxins, β-N-methylamino-l-alanine and microcystins, from a lake surrounded by cases of amyotrophic lateral sclerosis. Toxins 2015, 7, 322–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torbick, N.; Ziniti, B.; Stommel, E.; Linder, E.; Andrew, A.; Caller, T.; Haney, J.; Bradley, W.; Henegan, P.L.; Shi, X. Assessing cyanobacterial harmful algal blooms as risk factors for amyotrophic lateral sclerosis. Neurotox. Res. 2018, 33, 199–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanhope, J.M.; Brody, J.A.; Morris, C.E. Epidemiologic features of amyotrophic lateral sclerosis and parkinsonism-dementia in Guam, Mariana Islands. Int. J. Epidemiol. 1972, 1, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Cox, P.A.; Sacks, O.W. Cycad neurotoxins, consumption of flying foxes, and ALS-PDC disease in Guam. Neurology 2002, 58, 956–959. [Google Scholar] [CrossRef] [PubMed]
- Yanagihara, R.T.; Garruto, R.M.; Gajdusek, D.C. Epidemiological surveillance of amyotrophic lateral sclerosis and parkinsonism-dementia in the commonwealth of the Northern Mariana Islands. Ann. Neurol. 1983, 13, 79–86. [Google Scholar] [CrossRef]
- Cox, P.A.; Banack, S.; Murch, S.J. Biomagnification of cyanobacterial neurotoxins and neurodegenerative disease among the Chamorro people of Guam. Proc. Natl. Acad. Sci. USA 2003, 100, 13380–13383. [Google Scholar] [CrossRef] [Green Version]
- Murch, S.J.; Cox, P.A.; Banack, S.A.; Steele, J.C.; Sacks, O.W. Occurrence of β-methylamino-l-alanine (BMAA) in. ALS/PDC patients from Guam. Acta Neurol. Scand. 2004, 110, 267–269. [Google Scholar] [CrossRef] [PubMed]
- Kimura, K.; Yase, Y.; Higashi, Y.; Yamamoto, K.; Sugiura, M.; Tsumoto, T.; Uno, S.; Yoshimura, S.; Namikawa, K.; Kumura, J.; et al. Endemiological and geomedical studies on amyotrophic lateral sclerosis and allied diseases in Kii Peninsula, Japan (preliminary report). Psychiatry Clin. Neurosci. 1961, 15, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Yase, Y. The pathogenesis of amyotrophic lateral sclerosis. Lancet 1972, 300, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Gunnarsson, L.-G.; Lygner, P.-E.; Veiga-Cabo, J.; de Pedro-Cuesta, J. An epidemic-like cluster of motor neuron disease in a Swedish county during the period 1973–1984. Neuroepidemiology 1996, 15, 142–152. [Google Scholar] [CrossRef]
- Sienko, D.G.; Davis, J.P.; Taylor, J.A.; Brooks, B.R. Amyotrophic lateral sclerosis: A case-control study following detection of a cluster in a small Wisconsin community. Arch. Neurol. 1990, 47, 38–41. [Google Scholar] [CrossRef] [PubMed]
- Corcia, P.; Jafari-Schluep, H.-F.; Lardillier, D.; Mazyad, H.; Giraud, P.; Clavelou, P.; Pouget, J.; Camu, W. A clustering of conjugal amyotrophic lateral sclerosis in southeastern France. Arch. Neurol. 2003, 60, 553–557. [Google Scholar] [CrossRef] [Green Version]
- Uccelli, R.; Binazzi, A.; Altavista, P.; Belli, S.; Comba, P.; Mastrantonio, M.; Vanacore, N. Geographic distribution of amyotrophic lateral sclerosis through motor neuron disease mortality data. Eur. J. Epidemiol. 2007, 22, 781–790. [Google Scholar] [CrossRef] [PubMed]
- Sabel, C.E.; Boyle, P.J.; Löytönen, M.; Gatrell, A.C.; Jokelainen, M.; Flowerdew, R.; Maasilta, P. Spatial clustering of amyotrophic lateral sclerosis in Finland at place of birth and place of death. Am. J. Epidemiol. 2003, 157, 898–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horner, R.D.; Grambow, S.C.; Coffman, C.J.; Lindquist, J.H.; Oddone, E.Z.; Allen, K.D.; Kasarskis, E.J. Amyotrophic lateral sclerosis among 1991 Gulf War veterans: Evidence for a time-limited outbreak. Neuroepidemiology 2008, 31, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Miranda, M.L.; Alicia Overstreet Galeano, M.; Tassone, E.; Allen, K.D.; Horner, R.D. Spatial analysis of the etiology of amyotrophic lateral sclerosis among 1991 Gulf War veterans. Neurotoxicology 2008, 29, 964–970. [Google Scholar] [CrossRef]
- Stommel, E.W.; Field, N.C.; Caller, T.A. Aerosolization of cyanobacteria as a risk factor for amyotrophic lateral sclerosis. Med. Hypotheses 2013, 80, 142–145. [Google Scholar] [CrossRef]
- Masseret, E.; Banack, S.; Boumédiène, F.; Abadie, E.; Brient, L.; Pernet, F.; Juntas-Morales, R.; Pageot, N.; Metcalf, J.; Cox, P. Dietary BMAA exposure in an amyotrophic lateral sclerosis cluster from southern France. PLoS ONE 2013, 8, e83406. [Google Scholar] [CrossRef] [Green Version]
- Caller, T.A.; Chipman, J.W.; Field, N.C.; Stommel, E.W. Spatial analysis of amyotrophic lateral sclerosis in Northern New England, USA, 1997–2009. Muscle Nerve 2013, 48, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Torbick, N.; Hession, S.; Stommel, E.; Caller, T. Mapping amyotrophic lateral sclerosis lake risk factors across northern New England. Int. J. Health Geogr. 2014, 13, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delzor, A.; Couratier, P.; Boumédiène, F.; Nicol, M.; Druet-Cabanac, M.; Paraf, F.; Méjean, A.; Ploux, O.; Leleu, J.-P.; Brient, L.; et al. Searching for a link between the L-BMAA neurotoxin and amyotrophic lateral sclerosis: A study protocol of the French BMAA-LS programme. BMJ Open 2014, 4, e005528. [Google Scholar] [CrossRef]
- Lee, S.; Choi, B.; Kim, S.J.; Kim, J.; Kang, D.; Lee, J. Relationship between freshwater harmful algal blooms and neurodegenerative disease incidence rates in South Korea. Environ. Health 2022, 21, 116. [Google Scholar] [CrossRef]
- Lannuzel, A.; Mecharles, S.; Tressières, B.; Demoly, A.; Alhendi, R.; Hédreville-Tablon, M.-A.; Alecu, C. Clinical varieties and epidemiological aspects of amyotrophic lateral sclerosis in the Caribbean island of Guadeloupe: A new focus of ALS associated with Parkinsonism. Amyotroph. Lateral Scler. Front. Degener. 2015, 16, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Filippini, T.; Fiore, M.; Tesauro, M.; Malagoli, C.; Consonni, M.; Violi, F.; Arcolin, E.; Iacuzio, L.; Conti, G.O.; Cristaldi, A.; et al. Clinical and lifestyle factors and risk of amyotrophic lateral sclerosis: A population-based case-control study. Int. J. Environ. Res. Public Health 2020, 17, 857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Field, N.C.; Metcalf, J.S.; Caller, T.A.; Banack, S.A.; Cox, P.A.; Stommel, E.W. Linking β-methylamino-L-alanine exposure to sporadic amyotrophic lateral sclerosis in Annapolis, MD. Toxicon 2013, 70, 179–183. [Google Scholar] [CrossRef]
- Berntzon, L.; Ronnevi, L.; Bergman, B.; Eriksson, J. Detection of BMAA in the human central nervous system. Neuroscience 2015, 292, 137–147. [Google Scholar] [CrossRef] [Green Version]
- Pablo, J.; Banack, S.; Cox, P.; Johnson, T.; Papapetropoulos, S.; Bradley, W.; Buck, A.; Mash, D. Cyanobacterial neurotoxin BMAA in ALS and Alzheimer’s disease. Acta Psychiatr. Scand. 2009, 120, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Montine, T.J.; Li, K.; Perl, D.P.; Galasko, D. Lack of β-methylamino-L-alanine in brain from controls, AD, or Chamorros with PDC. Neurology 2005, 65, 768–769. [Google Scholar] [CrossRef] [PubMed]
- Meneely, J.P.; Chevallier, O.P.; Graham, S.; Greer, B.; Green, B.D.; Elliott, C.T. β-methylamino-L-alanine (BMAA) is not found in the brains of patients with confirmed Alzheimer’s disease. Sci. Rep. 2016, 6, 36363. [Google Scholar] [CrossRef] [PubMed]
- Spencer, P.S.; Palmer, V.S.; Kisby, G.E. Western Pacific ALS-PDC: Evidence implicating cycad genotoxins. J. Neurol. Sci. 2020, 419, 117185. [Google Scholar] [CrossRef] [PubMed]
- Jonasson, S.; Eriksson, J.; Berntzon, L.; Spáčil, Z.; Ilag, L.L.; Ronnevi, L.-O.; Rasmussen, U.; Bergman, B. Transfer of a cyanobacterial neurotoxin within a temperate aquatic ecosystem suggests pathways for human exposure. Proc. Natl. Acad. Sci. USA 2010, 107, 9252–9257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Eriksson, J.; Lage, S.; Jonasson, S.; Shams, S.; Mehine, M.; Ilag, L.L.; Rasmussen, U. Diatoms: A novel source for the neurotoxin BMAA in aquatic environments. PLoS ONE 2014, 9, e84578. [Google Scholar] [CrossRef]
- Johnson, H.E.; King, S.R.; Banack, S.A.; Webster, C.; Callanaupa, W.J.; Cox, P.A. Cyanobacteria (Nostoc commune) used as a dietary item in the Peruvian highlands produce the neurotoxic amino acid BMAA. J. Ethnopharmacol. 2008, 118, 159–165. [Google Scholar] [CrossRef]
- Cox, P.A.; Richer, R.; Metcalf, J.S.; Banack, S.A.; Codd, G.A.; Bradley, W.G. Cyanobacteria and BMAA exposure from desert dust: A possible link to sporadic ALS among Gulf War veterans. Amyotroph. Lateral Scler. 2009, 10, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Lage, S.; Costa, P.R.; Moita, T.; Eriksson, J.; Rasmussen, U.; Rydberg, S.J. BMAA in shellfish from two Portuguese transitional water bodies suggests the marine dinoflagellate Gymnodinium catenatum as a potential BMAA source. Aquat. Toxicol. 2014, 152, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Bradley, W.G.; Miller, R.; Levine, T.; Stommel, E.; Cox, P. Studies of environmental risk factors in amyotrophic lateral sclerosis (ALS) and a phase I clinical trial of L-serine. Neurotox. Res. 2018, 33, 192–198. [Google Scholar] [CrossRef]
- Saitou, T.; Sugiura, N.; Itayama, T.; Inamori, Y.; Matsumura, M. Degradation characteristics of microcystins by isolated bacteria from Lake Kasumigaura. J. Water Supply Res. Technol. AQUA 2003, 52, 13–18. [Google Scholar] [CrossRef]
- Edwards, C.; Graham, D.; Fowler, N.; Lawton, L.A. Biodegradation of microcystins and nodularin in freshwaters. Chemosphere 2008, 73, 1315–1321. [Google Scholar] [CrossRef] [PubMed]
- Gibble, C.M.; Peacock, M.B.; Kudela, R.M. Evidence of freshwater algal toxins in marine shellfish: Implications for human and aquatic health. Harmful Algae 2016, 59, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Chiswell, R.K.; Shaw, G.R.; Eaglesham, G.; Smith, M.J.; Norris, R.L.; Seawright, A.A.; Moore, M.R. Stability of cylindrospermopsin, the toxin from the cyanobacterium, Cylindrospermopsis raciborskii: Effect of pH, temperature, and sunlight on decomposition. Environ. Toxicol. 1999, 14, 155–161. [Google Scholar] [CrossRef]
- Adamski, M.; Żmudzki, P.; Chrapusta, E.; Bober, B.; Kaminski, A.; Zabaglo, K.; Latkowska, E.; Bialczyk, J. Effect of pH and temperature on the stability of cylindrospermopsin. Characterization of decomposition products. Algal Res. 2016, 15, 129–134. [Google Scholar] [CrossRef]
- Sedmak, B.; Carmeli, S.; Eleršek, T. "Non-toxic" cyclic peptides induce lysis of cyanobacteria—An effective cell population density control mechanism in cyanobacterial blooms. Microb. Ecol. 2008, 56, 201–209. [Google Scholar] [CrossRef]
- Sedmak, B.; Sukenik, A.; Eleršek, T.; Kosi, G. The biological role of cyclic hepatotoxic and non-hepatotoxic cyanopeptides and its ecological consequences. In Ecotoxicology Research Developments; Santos, E.B., Ed.; Nova Science Pub Inc.: London, UK, 2009; pp. 169–300. [Google Scholar]
- Bionda, N. Cyclic Lipodepsipeptides as Lead Structures for the Discovery of New Antibiotics. Ph.D. Thesis, Florida Atlantic University, Boca Raton, FL, USA, 2013. [Google Scholar]
- Kurtz, T.; Zeng, T.; Rosario-Ortiz, F.L. Photodegradation of cyanotoxins in surface waters. Water Res. 2021, 192, 116804. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Xing, P.; Wu, Q.L. The high resilience of the bacterioplankton community in the face of a catastrophic disturbance by a heavy Microcystis bloom. FEMS Microbiol. Ecol. 2012, 82, 192–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kormas, K.A.; Lymperopoulou, D.S. Cyanobacterial toxin degrading bacteria: Who are they? BioMed Res. Int. 2013, 2013, 463894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Wichelen, J.; Vanormelingen, P.; Codd, G.A.; Vyverman, W. The common bloom-forming cyanobacterium Microcystis is prone to a wide array of microbial antagonists. Harmful Algae 2016, 55, 97–111. [Google Scholar] [CrossRef]
- Mazur-Marzec, H.; Plinski, M. Do toxic cyanobacteria blooms pose a threat to the Baltic ecosystem? Oceanologia 2009, 51, 293–319. [Google Scholar] [CrossRef] [Green Version]
- Stolz, A. Molecular characteristics of xenobiotic-degrading sphingomonads. Appl. Microbiol. Biotechnol. 2009, 81, 793–811. [Google Scholar] [CrossRef] [PubMed]
- Kato, H.; Imanishi, S.Y.; Tsuji, K.; Harada, K.-I. Microbial degradation of cyanobacterial cyclic peptides. Water Res. 2007, 41, 1754–1762. [Google Scholar] [CrossRef]
- Yang, F.; Huang, F.; Feng, H.; Wei, J.; Massey, I.Y.; Liang, G.; Zhang, F.; Yin, L.; Kacew, S.; Zhang, X.; et al. A complete route for biodegradation of potentially carcinogenic cyanotoxin microcystin-LR in a novel indigenous bacterium. Water Res. 2020, 174, 115638. [Google Scholar] [CrossRef] [PubMed]
- Massey, I.Y.; Yang, F. A mini review on microcystins and bacterial degradation. Toxins 2020, 12, 268. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.; Zuo, Y.; Peng, L.; Gan, N.; Song, L. Widespread distribution and adaptive degradation of microcystin degrader (mlr-genotype) in Lake Taihu, China. Toxins 2021, 13, 864. [Google Scholar] [CrossRef] [PubMed]
- Okano, K.; Shimizu, K.; Maseda, H.; Kawauchi, Y.; Utsumi, M.; Itayama, T.; Zhang, Z.; Sugiura, N. Whole-genome sequence of the microcystin-degrading bacterium Sphingopyxis sp. strain C-1. Genome Announc. 2015, 3, e0083815. [Google Scholar] [CrossRef] [Green Version]
- Qin, L.; Zhang, X.; Chen, X.; Wang, K.; Shen, Y.; Li, D. Isolation of a novel microcystin-degrading bacterium and the evolutionary origin of mlr gene cluster. Toxins 2019, 11, 269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maghsoudi, E.; Fortin, N.; Greer, C.; Maynard, C.; Page, A.; Duy, S.V.; Sauve, S.; Prevost, M.; Dorner, S. Cyanotoxin degradation activity and mlr gene expression profiles of a Sphygopyxis sp. isolated from Lake Champlain, Canada. Environ. Sci. Process. Impacts 2016, 18, 1417–1426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imanishi, S.; Kato, H.; Mizuno, M.; Tsuji, K.; Harada, K.-I. Bacterial degradation of microcystins and nodularin. Chem. Res. Toxicol. 2005, 18, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, Z.A.; Alamri, S.A. Biodegradation of cylindrospermopsin toxin by microcystin-degrading bacteria isolated from cyanobacterial blooms. Toxicon 2012, 60, 1390–1395. [Google Scholar] [CrossRef] [PubMed]
- Dziga, D.; Kokocinski, M.; Maksylewicz, A.; Czaja-Prokop, U.; Barylski, J. Cylindrospermopsin biodegradation abilities of Aeromonas sp. isolated from Rusałka Lake. Toxins 2016, 8, 55. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, Z.A.; Hashem, M.; Alamri, S.A. Growth inhibition of the cyanobacterium Microcystis aeruginosa and degradation of its microcystin toxins by the fungus Trichoderma citrinoviride. Toxicon 2014, 86, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, Z.A.; Hashem, M.; Alamri, S.; Campos, A.; Vasconcelos, V. Fungal biodegradation and removal of cyanobacteria and microcystins: Potential applications and research needs. Environ. Sci. Pollut. Res. 2021, 28, 37041–37050. [Google Scholar] [CrossRef] [PubMed]
- Kurmayer, R.; Jüttner, F. Strategies for the co-existence of zooplankton with the toxic cyanobacterium Planktothrix rubescens in Lake Zurich. J. Plankton Res. 1999, 21, 659–683. [Google Scholar] [CrossRef]
- Akbar, S.; Huang, J.; Zhou, Q.; Gu, L.; Sun, Y.; Zhang, L.; Lyu, K.; Yang, Z. Elevated temperature and toxic Microcystis reduce Daphnia fitness and modulate gut microbiota. Environ. Pollut. 2021, 271, 116409. [Google Scholar] [CrossRef] [PubMed]
- Combes, A.; Dellinger, M.; Cadel-six, S.; Amand, S.; Comte, K. Ciliate Nassula sp. grazing on a microcystin-producing cyanobacterium (Planktothrix agardhii): Impact on cell growth and in the microcystin fractions. Aquatic Toxicol. 2013, 126, 435–441. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, Z.; Wang, N.; Gu, L.; Sun, Y.; Huang, Y.; Chen, Y.; Yang, Z. Mixotrophic Ochromonas addition improves the harmful Microcystis-dominated phytoplankton community in in situ microcosms. Environ. Sci. Technol. 2020, 54, 4609–4620. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, R.; Balasubramanian, R. Methods and approaches used for detection of cyanotoxins in environmental samples: A review. Crit. Rev. Environ. Sci. Technol. 2013, 43, 1349–1383. [Google Scholar] [CrossRef]
- Duncan, M.W. Good mass spectrometry and its place in good science. J. Mass Spectrom. 2012, 47, 795–809. [Google Scholar] [CrossRef] [PubMed]
- Faassen, E.J.; Antoniou, M.G.; Beekman-Lukassen, W.; Blahova, L.; Chernova, E.; Christophoridis, C.; Combes, A.; Edwards, C.; Fastner, J.; Harmsen, J. A collaborative evaluation of LC-MS/MS based methods for BMAA analysis: Soluble bound BMAA found to be an important fraction. Mar. Drugs 2016, 14, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanseverino, I.; António, D.C.; Loos, R.; Lettieri, T. Cyanotoxins: Methods and Approaches for Their Analysis and Detection; Centre, J.R., Ed.; Publications Office of the European Union: Luxembourg, 2017. [Google Scholar] [CrossRef]
- Sundaravadivelu, D.; Sanan, T.T.; Venkatapathy, R.; Mash, H.; Tettenhorst, D.; DAnglada, L.; Frey, S.; Tatters, A.O.; Lazorchak, J. Determination of cyanotoxins and prymnesins in water, fish tissue, and other matrices: A review. Toxins 2022, 14, 213. [Google Scholar] [CrossRef]
- Van Apeldoorn, M.E.; Van Egmond, H.P.; Speijers, G.J.; Bakker, G.J. Toxins of cyanobacteria. Mol. Nutr. Food Res. 2007, 51, 7–60. [Google Scholar] [CrossRef] [PubMed]
- Testai, E.; Buratti, F.M.; Funari, E.; Manganelli, M.; Vichi, S.; Arnich, N.; Biré, R.; Fessard, V.; Sialehaamoa, A. Review and analysis of occurrence, exposure and toxicity of cyanobacteria toxins in food. EFSA Support. Publ. 2016, 13, 998E. [Google Scholar] [CrossRef]
- Merel, S.; Walker, D.; Chicana, R.; Snyder, S.; Baurès, E.; Thomas, O. State of knowledge and concerns on cyanobacterial blooms and cyanotoxins. Environ. Int. 2013, 59, 303–327. [Google Scholar] [CrossRef] [PubMed]
- Abbes, S.; Vo Duy, S.; Munoz, G.; Dinh, Q.T.; Simon, D.F.; Husk, B.; Baulch, H.M.; Vinçon-Leite, B.; Fortin, N.; Greer, C.W.; et al. Occurrence of BMAA isomers in bloom-impacted lakes and reservoirs of Brazil, Canada, France, Mexico, and the United Kingdom. Toxins 2022, 14, 251. [Google Scholar] [CrossRef]
- Altenburger, R.; Ait-Aissa, S.; Antczak, P.; Backhaus, T.; Barceló, D.; Seiler, T.-B.; Brion, F.; Busch, W.; Chipman, K.; de Alda, M.L.; et al. Future water quality monitoring-adapting tools to deal with mixtures of pollutants in water resource management. Sci. Total Environ. 2015, 512, 540–551. [Google Scholar] [CrossRef] [Green Version]
- Hollender, J.; Schymanski, E.L.; Singer, H.P.; Ferguson, P.L. Nontarget screening with high resolution mass spectrometry in the environment: Ready to go? Environ. Sci. Technol. 2017, 51, 11505–11512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escher, B.I.; Stapleton, H.M.; Schymanski, E.L. Tracking complex mixtures of chemicals in our changing environment. Science 2020, 367, 388–392. [Google Scholar] [CrossRef]
- Dom, I.; Biré, R.; Hort, V.; Lavison-Bompard, G.; Nicolas, M.; Guérin, T. Extended targeted and non-targeted strategies for the analysis of marine toxins in mussels and oysters by (LC-HRMS). Toxins 2018, 10, 375. [Google Scholar] [CrossRef] [Green Version]
- Krauss, M.; Singer, H.; Hollender, J. LC–high resolution MS in environmental analysis: From target screening to the identification of unknowns. Anal. Bioanal. Chem. 2010, 397, 943–951. [Google Scholar] [CrossRef] [Green Version]
- Bogialli, S.; Bortolini, C.; Di Gangi, I.M.; Di Gregorio, F.N.; Lucentini, L.; Favaro, G.; Pastore, P. Liquid chromatography-high resolution mass spectrometric methods for the surveillance monitoring of cyanotoxins in freshwaters. Talanta 2017, 170, 322–330. [Google Scholar] [CrossRef]
- Gonzalez-Gaya, B.; Lopez-Herguedas, N.; Bilbao, D.; Mijangos, L.; Iker, A.; Etxebarria, N.; Irazola, M.; Prieto, A.; Olivares, M.; Zuloaga, O. Suspect and non-target screening: The last frontier in environmental analysis. Anal. Methods 2021, 13, 1876–1904. [Google Scholar] [CrossRef] [PubMed]
- Picardo, M.; Sanchís, J.; Núñez, O.; Farré, M. Suspect screening of natural toxins in surface and drinking water by high performance liquid chromatography and high-resolution mass spectrometry. Chemosphere 2020, 261, 127888. [Google Scholar] [CrossRef] [PubMed]
- Bishop, C.T.; Anet, E.F.L.J.; Gorham, P.R. Isolation and identification of the fast-death factor in Microcystis aeruginosa NRC-1. Can. J. Biochem. Physiol. 1959, 37, 453–471. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Wang, W.; Xu, Q.; Liu, Z.; Teng, J.; Yan, H.; Liu, X. Microcystins in water: Detection, microbial degradation strategies, and mechanisms. Int. J. Environ. Res. Public Health 2022, 19, 13175. [Google Scholar] [CrossRef] [PubMed]
- Romera-García, E.; Helmus, R.; Ballesteros-Gómez, A.; Visser, P.M. Multi-class determination of intracellular and extracellular cyanotoxins in freshwater samples by ultra-high performance liquid chromatography coupled to high resolution mass spectrometry. Chemosphere 2021, 274, 129770. [Google Scholar] [CrossRef] [PubMed]
- Van Hassel, W.H.R.; Huybrechts, B.; Masquelier, J.; Wilmotte, A.; Andjelkovic, M. Development, validation and application of a targeted LC-MS method for quantification of microcystins and nodularin: Towards a better characterization of drinking water. Water 2022, 14, 1195. [Google Scholar] [CrossRef]
- Panda, D.; Dash, B.P.; Manickam, S.; Boczkaj, G. Recent advancements in LC-MS based analysis of biotoxins: Present and future challenges. Mass Spectrom. Rev. 2022, 41, 766–803. [Google Scholar] [CrossRef]
- Triantis, T.; Tsimeli, K.; Kaloudis, T.; Thanassoulias, N.; Lytras, E.; Hiskia, A. Development of an integrated laboratory system for the monitoring of cyanotoxins in surface and drinking waters. Toxicon 2010, 55, 979–989. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, J.; Zhang, D.; Luo, L.; Liao, Q.; Yuan, L.; Wu, N. Seasonal and spatial variations of microcystins in Poyang Lake, the largest freshwater lake in China. Environ. Sci. Pollut. Res. Int. 2018, 25, 6300–6307. [Google Scholar] [CrossRef]
- Turner, A.D.; Dhanji-Rapkova, M.; O’Neill, A.; Coates, L.; Lewis, A.; Lewis, K. Analysis of microcystins in cyanobacterial blooms from freshwater bodies in England. Toxins 2018, 10, 39. [Google Scholar] [CrossRef] [Green Version]
- Mantzouki, E.; Lurling, M.; Fastner, J.; de Senerpont Domis, L.; Wilk-Wozniak, E.; Koreiviene, J.; Seelen, L.; Teurlincx, S.; Verstijnen, Y.; Krzton, W.; et al. Temperature effects explain continental scale distribution of cyanobacterial toxins. Toxins 2018, 10, 156. [Google Scholar] [CrossRef] [Green Version]
- Main, B.J.; Bowling, L.C.; Padula, M.P.; Bishop, D.P.; Mitrovic, S.M.; Guillemin, G.J.; Rodgers, K.J. Detection of the suspected neurotoxin beta-methylamino-l-alanine (BMAA) in cyanobacterial blooms from multiple water bodies in Eastern Australia. Harmful Algae 2018, 74, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Pitois, F.; Fastner, J.; Pagotto, C.; Dechesne, M. Multi-toxin occurrences in ten French water resource reservoirs. Toxins 2018, 10, 283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, A.D.; Waack, J.; Lewis, A.; Edwards, C.; Lawton, L. Development and single-laboratory validation of a UHPLC-MS/MS method for quantitation of microcystins and nodularin in natural water, cyanobacteria, shellfish and algal supplement tablet powders. J. Chromatogr. B 2018, 1074–1075, 111–123. [Google Scholar] [CrossRef]
- Thuret-Benoist, H.; Pallier, V.; Feuillade-Cathalifaud, G. Quantification of microcystins in natural waters by HPLC-UV after a pre-concentration step: Validation of the analytical performances and study of the interferences. Environ. Toxicol. Pharmacol. 2019, 72, 103223. [Google Scholar] [CrossRef]
- Botha, C.J.; Laver, P.; Singo, A.; Venter, E.; Ferreira, G.C.H.; Rösemann, M.; Myburgh, J.G. Evaluation of a Norwegian-developed ELISA to determine microcystin concentrations in fresh water. Water Supply 2019, 19, 743–752. [Google Scholar] [CrossRef]
- Haddad, S.P.; Bobbitt, J.M.; Taylor, R.B.; Lovin, L.M.; Conkle, J.L.; Chambliss, C.K.; Brooks, B.W. Determination of microcystins, nodularin, anatoxin-a, cylindrospermopsin, and saxitoxin in water and fish tissue using isotope dilution liquid chromatography tandem mass spectrometry. J. Chromatogr. A 2019, 1599, 66–74. [Google Scholar] [CrossRef]
- Leon, C.; Penuela, G.A. Detected cyanotoxins by UHPLC MS/MS technique in tropical reservoirs of northeastern Colombia. Toxicon 2019, 167, 38–48. [Google Scholar] [CrossRef]
- Vo Duy, S.; Munoz, G.; Dinh, Q.T.; Tien Do, D.; Simon, D.F.; Sauve, S. Analysis of the neurotoxin beta-N-methylamino-L-alanine (BMAA) and isomers in surface water by FMOC derivatization liquid chromatography high resolution mass spectrometry. PLoS ONE 2019, 14, e0220698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKindles, K.M.; Zimba, P.V.; Chiu, A.S.; Watson, S.B.; Gutierrez, D.B.; Westrick, J.; Kling, H.; Davis, T.W. A multiplex analysis of potentially toxic cyanobacteria in Lake Winnipeg during the 2013 bloom season. Toxins 2019, 11, 587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, B.; Liu, Z.; Huang, R.; Xu, Y.; Liu, D.; Wang, W.; Zhao, Z.; Cui, F.; Shi, W. Impact factors on the production of beta-methylamino-L-alanine (BMAA) by cyanobacteria. Chemosphere 2020, 243, 125355. [Google Scholar] [CrossRef] [PubMed]
- Abbas, F.; Porojan, C.; Mowe, M.A.D.; Lehane, M.; Mitrovic, S.M.; Lim, R.P.; Yeo, D.C.J.; Furey, A. Sample extraction and liquid chromatography–tandem mass spectrometry (LC-MS/MS) method development and validation for the quantitative detection of cyanobacterial hepatotoxins and neurotoxins in Singapore’s reservoirs. Mar. Freshw. Res. 2020, 71, 673–688. [Google Scholar] [CrossRef]
- Stoyneva-Gärtner, M.; Stefanova, K.; Descy, J.-P.; Uzunov, B.; Radkova, M.; Pavlova, V.; Mitreva, M.; Gärtner, G. Microcystis aeruginosa and M. wesenbergii were the primary planktonic microcystin producers in several Bulgarian waterbodies (August 2019). Appl. Sci. 2020, 11, 357. [Google Scholar] [CrossRef]
- Zervou, S.K.; Moschandreou, K.; Paraskevopoulou, A.; Christophoridis, C.; Grigoriadou, E.; Kaloudis, T.; Triantis, T.M.; Tsiaoussi, V.; Hiskia, A. Cyanobacterial toxins and peptides in Lake Vegoritis, Greece. Toxins 2021, 13, 394. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Husk, B.R.; Duy, S.V.; Dinh, Q.T.; Sanchez, J.S.; Sauve, S.; Whalen, J.K. Quantitative screening for cyanotoxins in soil and groundwater of agricultural watersheds in Quebec, Canada. Chemosphere 2021, 274, 129781. [Google Scholar] [CrossRef]
- Choi, J.-W.; Jang, J.-H.; Kim, A.-K.; Lee, S.-H. Determination of β-N-Methylamino-L-Alanine (BMAA) levels in surface water and treated water by liquid chromatography-tandem mass spectrometry. J. Environ. Anal. Health Toxicol. 2021, 24, 171–175. [Google Scholar] [CrossRef]
- Dinh, Q.T.; Munoz, G.; Simon, D.F.; Vo Duy, S.; Husk, B.; Sauve, S. Stability issues of microcystins, anabaenopeptins, anatoxins, and cylindrospermopsin during short-term and long-term storage of surface water and drinking water samples. Harmful Algae 2021, 101, 101955. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Xie, H.; Cao, Y.; Xu, Q.; Gan, N. Magnetic solid phase extraction coupled with high-performance liquid chromatography-diode array detection based on assembled magnetic covalent organic frameworks for selective extraction and detection of microcystins in aquatic foods. J. Chromatogr. A 2022, 1685, 463614. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Huang, J.J.; Chen, J.; He, X.; Wang, Y.; Wang, J.; Wang, B. Trace determination of multiple hydrophilic cyanotoxins in freshwater by off- and on-line solid phase extraction coupled to liquid chromatography-tandem mass spectrometry. Sci. Total Environ. 2022, 853, 158545. [Google Scholar] [CrossRef]
- Van Hassel, W.H.R.; Andjelkovic, M.; Durieu, B.; Marroquin, V.A.; Masquelier, J.; Huybrechts, B.; Wilmotte, A. A summer of cyanobacterial blooms in Belgian waterbodies: Microcystin quantification and molecular characterizations. Toxins 2022, 14, 61. [Google Scholar] [CrossRef]
- Choi, J.-W.; Jang, J.-H.; Lee, S.-H.; Yoon, M.-A. Determination of cylindrospermopsin in surface and treated water using liquid chromatography-tandem mass spectrometry. Environ. Anal. Health Toxicol. 2022, 25, 71–76. [Google Scholar] [CrossRef]
- Duy, S.V.; Munoz, G.; Dinh, Q.T.; Zhang, Y.; Simon, D.F.; Sauve, S. Fast screening of saxitoxin, neosaxitoxin, and decarbamoyl analogues in fresh and brackish surface waters by on-line enrichment coupled to HILIC-HRMS. Talanta 2022, 241, 123267. [Google Scholar] [CrossRef] [PubMed]
- Aparicio-Muriana, M.M.; Carmona-Molero, R.; Lara, F.J.; Garcia-Campana, A.M.; Del Olmo-Iruela, M. Multiclass cyanotoxin analysis in reservoir waters: Tandem solid-phase extraction followed by zwitterionic hydrophilic interaction liquid chromatography-mass spectrometry. Talanta 2022, 237, 122929. [Google Scholar] [CrossRef]
- Zhao, P.; Qiu, J.; Li, A.; Yan, G.; Li, M.; Ji, Y. Matrix effect of diverse biological samples extracted with different extraction ratios on the detection of beta-N-methylamino-L-alanine by two common LC-MS/MS analysis methods. Toxins 2022, 14, 387. [Google Scholar] [CrossRef] [PubMed]
- Emmons, R.V.; Karaj, E.; Cudjoe, E.; Bell, D.S.; Tillekeratne, L.M.V.; Gionfriddo, E. Leveraging multi-mode microextraction and liquid chromatography stationary phases for quantitative analysis of neurotoxin beta-N-methylamino-L-alanine and other non-proteinogenic amino acids. J. Chromatogr. A 2022, 1685, 463636. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, Y.; Abdallah, M.F.; Tan, H.; Li, J.; Liu, S.; Zhang, R.; Sun, F.; Li, Y.; Yang, S. Novel one-point calibration strategy for high-throughput quantitation of microcystins in freshwater using LC-MS/MS. Sci. Total Environ. 2023, 858, 159345. [Google Scholar] [CrossRef]
- Kulabhusan, P.K.; Campbell, K. Recent trends in the detection of freshwater cyanotoxins with a critical note on their occurrence in Asia. Trends Environ. Anal. Chem. 2021, 32, e00150. [Google Scholar] [CrossRef]
- Lee, J.; Lee, S.; Jiang, X. Cyanobacterial toxins in freshwater and food: Important sources of exposure to humans. Annu. Rev. Food Sci. Technol. 2017, 8, 281–304. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, M.C.; Jiang, S.C. Can cyanotoxins penetrate human skin during water recreation to cause negative health effects? Harmful Algae 2020, 98, 101872. [Google Scholar] [CrossRef]
- Tatters, A.O.; Howard, M.D.; Nagoda, C.; Busse, L.; Gellene, A.G.; Caron, D.A. Multiple stressors at the land-sea interface: Cyanotoxins at the land-sea interface in the Southern California Bight. Toxins 2017, 9, 95. [Google Scholar] [CrossRef] [Green Version]
- Tatters, A.O.; Howard, M.D.; Nagoda, C.; Fetscher, A.E.; Kudela, R.M.; Caron, D.A. Heterogeneity of toxin-producing cyanobacteria and cyanotoxins in coastal watersheds of southern California. Estuaries Coasts 2019, 42, 958–975. [Google Scholar] [CrossRef]
- Tatters, A.O.; Smith, J.; Kudela, R.M.; Hayashi, K.; Howard, M.D.; Donovan, A.R.; Loftin, K.A.; Caron, D.A. The tide turns: Episodic and localized cross-contamination of a California coastline with cyanotoxins. Harmful Algae 2021, 103, 102003. [Google Scholar] [CrossRef]
- Howard, M.D.; Smith, J.; Caron, D.A.; Kudela, R.M.; Loftin, K.; Hayashi, K.; Fadness, R.; Fricke, S.; Kann, J.; Roethler, M.; et al. Integrative monitoring strategy for marine and freshwater harmful algal blooms and toxins across the freshwater-to-marine continuum. Integr. Environ. Assess. Manag. 2022, 1, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Andraka, D.; Ospanov, K.; Myrzakhmetov, M. Current state of communal sewage treatment in the republic of Kazakhstan. J. Ecol. Eng. 2015, 16, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez-Praena, D.; Jos, Á.; Pichardo, S.; Moreno, I.M.; Cameán, A.M. Presence and bioaccumulation of microcystins and cylindrospermopsin in food and the effectiveness of some cooking techniques at decreasing their concentrations: A review. Food Chem. Toxicol. 2013, 53, 139–152. [Google Scholar] [CrossRef]
- Abdallah, M.F.; Van Hassel, W.H.; Andjelkovic, M.; Wilmotte, A.; Rajkovic, A. Cyanotoxins and food contamination in developing countries: Review of their types, toxicity, analysis, occurrence and mitigation strategies. Toxins 2021, 13, 786. [Google Scholar] [CrossRef] [PubMed]
- Downing, S.; Contardo-Jara, V.; Pflugmacher, S.; Downing, T.G. The fate of the cyanobacterial toxin β-N-methylamino-L-alanine in freshwater mussels. Ecotoxicol. Environ. Safety 2014, 101, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Baptista, M.S.; Vasconcelos, R.G.; Ferreira, P.C.; Almeida, C.M.R.; Vasconcelos, V.M. Assessment of the non-protein amino acid BMAA in Mediterranean mussel Mytilus galloprovincialis after feeding with estuarine cyanobacteria. Environ. Sci. Pollution Res. 2015, 22, 12501–12510. [Google Scholar] [CrossRef] [PubMed]
- Lance, E.; Arnich, N.; Maignien, T.; Biré, R. Occurrence of β-N-methylamino-l-alanine (BMAA) and isomers in aquatic environments and aquatic food sources for humans. Toxins 2018, 10, 83. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.Y.; Rydberg, S. Transfer of the neurotoxin β-N-methylamino-l-alanine (BMAA) in the agro–aqua cycle. Mar. Drugs 2020, 18, 244. [Google Scholar] [CrossRef] [PubMed]
- Crush, J.; Briggs, L.; Sprosen, J.; Nichols, S. Effect of irrigation with lake water containing microcystins on microcystin content and growth of ryegrass, clover, rape, and lettuce. Environ. Toxicol. 2008, 23, 246–252. [Google Scholar] [CrossRef]
- Mohamed, Z.A.; Al Shehri, A.M. Microcystins in groundwater wells and their accumulation in vegetable plants irrigated with contaminated waters in Saudi Arabia. J. Hazard. Mater. 2009, 172, 310–315. [Google Scholar] [CrossRef]
- Kittler, K.; Schreiner, M.; Krumbein, A.; Manzei, S.; Koch, M.; Rohn, S.; Maul, R. Uptake of the cyanobacterial toxin cylindrospermopsin in Brassica vegetables. Food Chem. 2012, 133, 875–879. [Google Scholar] [CrossRef]
- Roney, B.R.; Renhui, L.; Banack, S.A.; Murch, S.; Honegger, R.; Cox, P.A. Consumption of fa cai Nostoc soup: A potential for BMAA exposure from Nostoc cyanobacteria in China? Amyotroph. Lateral Scler. 2009, 10, 44–49. [Google Scholar] [CrossRef]
- Han, D.; Deng, Z.; Lu, F.; Hu, Z. Biology and biotechnology of edible Nostoc. In Handbook of Microalgal Culture: Applied Phycology and Biotechnology, 2nd ed.; Richmond, A., Hu, Q., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2013; pp. 433–444. [Google Scholar] [CrossRef]
- Fidor, A.; Konkel, R.; Mazur-Marzec, H. Bioactive peptides produced by cyanobacteria of the genus Nostoc: A review. Mar. Drugs 2019, 17, 561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Lloréns, J.L. Microalgae: From staple foodstuff to avant-garde cuisine. Inter. J. Gastr. Food Sci. 2020, 21, 100221. [Google Scholar] [CrossRef]
- McCarron, P.; Logan, A.C.; Giddings, S.D.; Quilliam, M.A. Analysis of β-N-methylamino-L-alanine (BMAA) in spirulina-containing supplements by liquid chromatography-tandem mass spectrometry. Aquat. Biosyst. 2014, 10, 5. [Google Scholar] [CrossRef] [Green Version]
- Roy-Lachapelle, A.; Solliec, M.; Bouchard, M.F.; Sauvé, S. Detection of cyanotoxins in algae dietary supplements. Toxins 2017, 9, 76. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Parra, E.; Boutarfa, S.; Aboal, M. Are cyanotoxins the only toxic compound potentially present in microalgae supplements? Results from a study of ecological and non-ecological products. Toxins 2020, 12, 552. [Google Scholar] [CrossRef]
- Murch, S.J.; Cox, P.A.; Banack, S.A. A mechanism for slow release of biomagnified cyanobacterial neurotoxins and neuro-degenerative disease in Guam. Proc. Natl. Acad. Sci. USA 2004, 101, 12228–12231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, R.M., Jr.; Larson, D.A.; Bold, H.C. Airborne algae: Their abundance and heterogeneity. Science 1964, 143, 583–585. [Google Scholar] [CrossRef] [PubMed]
- Olson, N.E.; Cooke, M.E.; Shi, J.H.; Birbeck, J.A.; Westrick, J.A.; Ault, A.P. Harmful algal bloom toxins in aerosol generated from inland lake water. Environ. Sci. Technol. 2020, 54, 4769–4780. [Google Scholar] [CrossRef] [PubMed]
- Harb, C.; Pan, J.; DeVilbiss, S.; Badgley, B.; Marr, L.C.; Schmale, D.G., III; Foroutan, H. Increasing freshwater salinity impacts aerosolized bacteria. Environ. Sci. Technol. 2021, 55, 5731–5741. [Google Scholar] [CrossRef] [PubMed]
- Backer, L.C.; McNeel, S.V.; Barber, T.; Kirkpatrick, B.; Williams, C.; Irvin, M.; Zhou, Y.; Johnson, T.B.; Nierenberg, K.; Aubel, M.; et al. Recreational exposure to microcystins during algal blooms in two California lakes. Toxicon 2010, 55, 909–921. [Google Scholar] [CrossRef]
- Brodie, E.L.; DeSantis, T.Z.; Parker, J.P.M.; Zubietta, I.X.; Piceno, Y.M.; Andersen, G.L. Urban aerosols harbor diverse and dynamic bacterial populations. Proc. Natl. Acad. Sci. USA 2007, 104, 299–304. [Google Scholar] [CrossRef] [Green Version]
- Wood, S.A.; Dietrich, D.R. Quantitative assessment of aerosolized cyanobacterial toxins at two New Zealand lakes. J. Environ. Monit. 2011, 13, 1617–1624. [Google Scholar] [CrossRef] [Green Version]
- Genitsaris, S.; Kormas, K.A.; Moustaka-Gouni, M. Airborne algae and cyanobacteria: Occurrence and related health effects. Front. Biosci.-Elite 2011, 3, 772–787. [Google Scholar]
- Wisniewska, K.A.; Śliwińska-Wilczewska, S.; Lewandowska, A.U. Airborne microalgal and cyanobacterial diversity and composition during rain events in the southern Baltic Sea region. Sci. Rep. 2022, 12, 2029. [Google Scholar] [CrossRef]
- Plaas, H.E.; Paerl, R.W.; Baumann, K.; Karl, C.; Popendorf, K.J.; Barnard, M.A.; Chang, N.Y.; Curtis, N.P.; Huang, H.; Mathieson, O.L.; et al. Harmful cyanobacterial aerosolization dynamics in the airshed of a eutrophic estuary. Sci. Total Environ. 2022, 852, 158383. [Google Scholar] [CrossRef]
- Berstein, I.I.; Safferman, R.S. Viable algae in house dust. Nature 1970, 227, 851–852. [Google Scholar] [CrossRef]
- Després, V.; Huffman, J.A.; Burrows, S.M.; Hoose, C.; Safatov, A.; Buryak, G.; Fröhlich-Nowoisky, J.; Elbert, W.; Andreae, M.; Pöschl, U.; et al. Primary biological aerosol particles in the atmosphere: A review. Tellus B Chem. Phys. Meteorol. 2012, 64, 15598. [Google Scholar] [CrossRef] [Green Version]
- Wisniewska, K.; Lewandowska, A.U.; Śliwińska-Wilczewska, S. The importance of Cyanobacteria and microalgae present in aerosols to human health and the environment—Review study. Environ. Int. 2019, 131, 104964. [Google Scholar] [CrossRef]
- Wiśniewska, K.A.; Śliwińska-Wilczewska, S.; Lewandowska, A.U. The first characterization of airborne cyanobacteria and microalgae in the Adriatic Sea region. PLoS ONE 2020, 15, e0238808. [Google Scholar] [CrossRef] [PubMed]
- Gärtner, G.; Stoyneva-Gärtner, M.; Uzunov, B. Algal toxic compounds and their aeroterrestrial, airborne and other extremophilic producers with attention to soil and plant contamination: A review. Toxins 2021, 13, 322. [Google Scholar] [CrossRef]
- Scott, L.L.; Downing, S.; Downing, T.G. The evaluation of BMAA inhalation as a potential exposure route using a rat model. Neurotox. Res. 2018, 33, 6–14. [Google Scholar] [CrossRef]
- Plaas, H.E.; Paerl, H.W. Toxic cyanobacteria: A growing threat to water and air quality. Environ. Sci. Technol. 2020, 55, 44–64. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, V.R.; Mancin, V.G.; Pinto, E.F.; Soares, R.M.; Azevedo, S.M.; Macchione, M.; Carvalho, A.R.; Zin, W.A. Repeated intranasal exposure to microcystin-LR affects lungs but not nasal epithelium in mice. Toxicon 2015, 104, 14–18. [Google Scholar] [CrossRef]
- Li, X.; Xu, L.; Zhou, W.; Zhao, Q.; Wang, Y. Chronic exposure to microcystin-LR affected mitochondrial DNA maintenance and caused pathological changes of lung tissue in mice. Environ. Pollut. 2016, 210, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, G.M.; Oliveira, V.R.; Soares, R.M.; Azevedo, S.M.; Lima, L.M.; Barreiro, E.J.; Valença, S.S.; Saldiva, P.H.; Faffe, D.S.; Zin, W.A. Can LASSBio 596 and dexamethasone treat acute lung and liver inflammation induced by microcystin-LR? Toxicon 2010, 56, 604–612. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Sun, H.; Yan, W.; Xu, D.; Shen, T. A proteomic study of the pulmonary injury induced by microcystin-LR in mice. Toxicon 2018, 150, 304–314. [Google Scholar] [CrossRef]
- Breidenbach, J.D.; French, B.W.; Gordon, T.T.; Kleinhenz, A.L.; Khalaf, F.K.; Willey, J.C.; Hammersley, J.R.; Wooten, R.M.; Crawford, E.L.; Modyanov, N.N. Microcystin-LR aerosol induces inflammatory responses in healthy human primary airway epithelium. Environ. Int. 2022, 169, 107531. [Google Scholar] [CrossRef]
- Facciponte, D.N.; Bough, M.W.; Seidler, D.; Carroll, J.L.; Ashare, A.; Andrew, A.S.; Tsongalis, G.J.; Vaickus, L.J.; Henegan, P.L.; Butt, T.H. Identifying aerosolized cyanobacteria in the human respiratory tract: A proposed mechanism for cyanotoxin-associated diseases. Sci. Total Environ. 2018, 645, 1003–1013. [Google Scholar] [CrossRef]
- Pierozan, P.; Piras, E.; Brittebo, E.; Karlsson, O. The cyanobacterial neurotoxin β-N-methylamino-l-alanine (BMAA) targets the olfactory bulb region. Arch. Toxicol. 2020, 94, 2799–2808. [Google Scholar] [CrossRef]
- Sarasa, M.; Pesini, P. Natural non-transgenic animal models for research in Alzheimer’s disease. Curr. Alzheimer Res. 2009, 6, 171–178. [Google Scholar] [CrossRef] [Green Version]
- Davis, D.A.; Cox, P.A.; Banack, S.A.; Lecusay, P.D.; Garamszegi, S.P.; Hagan, M.J.; Powell, J.T.; Metcalf, J.S.; Palmour, R.M.; Beierschmitt, A.; et al. L-serine reduces spinal cord pathology in a vervet model of preclinical ALS/MND. J. Neuropathol. Exp. Neurol. 2020, 79, 393–406. [Google Scholar] [CrossRef] [PubMed]
- Mondo, K.; Hammerschlag, N.; Basile, M.; Pablo, J.; Banack, S.A.; Mash, D.C. Cyanobacterial neurotoxin β-N-methylamino-L-alanine (BMAA) in shark fins. Mar. Drugs 2012, 10, 509–520. [Google Scholar] [CrossRef] [Green Version]
- Davis, D.A.; Garamszegi, S.P.; Banack, S.A.; Dooley, P.D.; Coyne, T.M.; McLean, D.W.; Rotstein, D.S.; Mash, D.C.; Cox, P.A. BMAA, Methylmercury, and mechanisms of neurodegeneration in dolphins: A natural model of toxin exposure. Toxins 2021, 13, 697. [Google Scholar] [CrossRef] [PubMed]
- Davis, D.A.; Mondo, K.; Stern, E.; Annor, A.K.; Murch, S.J.; Coyne, T.M.; Brand, L.E.; Niemeyer, M.E.; Sharp, S.; Bradley, W.G.; et al. Cyanobacterial neurotoxin BMAA and brain pathology in stranded dolphins. PLoS ONE 2019, 14, e0213346. [Google Scholar] [CrossRef]
- Lobner, D.; Piana, P.M.T.; Salous, A.K.; Peoples, R.W. β-N-methylamino-L-alanine enhances neurotoxicity through multiple mechanisms. Neurobiol. Dis. 2007, 25, 360–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedrosa, C.D.S.G.; Souza, L.R.Q.; Gomes, T.A.; de Lima, C.V.F.; Ledur, P.F.; Karmirian, K.; Barbeito-Andres, J.; Costa, M.D.N.; Higa, L.M.; Rossi, Á.D.; et al. The cyanobacterial saxitoxin exacerbates neural cell death and brain malformations induced by Zika virus. PLoS Negl. Trop. Dis. 2020, 14, e0008060. [Google Scholar] [CrossRef] [Green Version]
- Wiese, M.; D’agostino, P.M.; Mihali, T.K.; Moffitt, M.C.; Neilan, B.A. Neurotoxic alkaloids: Saxitoxin and its analogs. Mar. Drugs 2010, 8, 2185–2211. [Google Scholar] [CrossRef] [Green Version]
- Smith, F.M.; Wood, S.A.; van Ginkel, R.; Broady, P.A.; Gaw, S. First report of saxitoxin production by a species of the freshwater benthic cyanobacterium, Scytonema Agardh. Toxicon 2011, 57, 566–573. [Google Scholar] [CrossRef]
- Cusick, K.D.; Sayler, G.S. An overview on the marine neurotoxin, saxitoxin: Genetics, molecular targets, methods of detection and ecological functions. Mar. Drugs 2013, 11, 991–1018. [Google Scholar] [CrossRef] [Green Version]
- Roué, M.; Gugger, M.; Golubic, S.; Amzil, Z.; Aráoz, R.; Turquet, J.; Chinain, M.; Laurent, D. Marine cyanotoxins potentially harmful to human health. In Outstanding Marine Molecules; Chapter 1; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2014; pp. 1–22. [Google Scholar] [CrossRef]
- Bonnard, I.; Bornancin, L.; Dalle, K.; Chinain, M.; Zubia, M.; Banaigs, B.; Roué, M. Assessment of the chemical diversity and potential toxicity of benthic cyanobacterial blooms in the lagoon of Moorea Island (French Polynesia). J. Mar. Sci. Eng. 2020, 8, 406. [Google Scholar] [CrossRef]
- Weiss, C.; Figueras, E.; Borbely, A.N.; Sewald, N. Cryptophycins: Cytotoxic cyclodepsipeptides with potential for tumor targeting. J. Pept. Sci. 2017, 23, 514–531. [Google Scholar] [CrossRef] [Green Version]
- Kust, A.; Řeháková, K.; Vrba, J.; Maicher, V.; Mareš, J.; Hrouzek, P.; Chiriac, M.C.; Benedová, Z.; Tesařová, B.; Saurav, K. Insight into unprecedented diversity of cyanopeptides in eutrophic ponds using an ms/ms networking approach. Toxins 2020, 12, 561. [Google Scholar] [CrossRef]
- Panda, D.; Himes, R.H.; Moore, R.E.; Wilson, L.; Jordan, M.A. Mechanism of action of the unusually potent microtubule inhibitor cryptophycin 1. Biochemistry 1997, 36, 12948–12953. [Google Scholar] [CrossRef]
- Costa, M.; Costa-Rodrigues, J.; Fernandes, M.H.; Barros, P.; Vasconcelos, V.; Martins, R. Marine cyanobacteria compounds with anticancer properties: A review on the implication of apoptosis. Mar. Drugs 2012, 10, 2181–2207. [Google Scholar] [CrossRef] [Green Version]
- Luesch, H.; Chanda, S.K.; Raya, R.M.; DeJesus, P.D.; Orth, A.P.; Walker, J.R.; Izpisúa Belmonte, J.C.; Schultz, P.G. A functional genomics approach to the mode of action of apratoxin A. Nat. Chem. Biol. 2006, 2, 158–167. [Google Scholar] [CrossRef]
- Hastie, C.J.; Borthwick, E.B.; Morrison, L.F.; Codd, G.A.; Cohen, P.T.W. Inhibition of several protein phosphatases by a non-covalently interacting microcystin and a novel cyanobacterial peptide, nostocyclin. Biochim. Biophys. Acta 2005, 1726, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Maynes, J.T.; Luu, H.A.; Cherney, M.M.; Andersen, R.J.; Williams, D.; Holmes, C.F.; James, M.N. Crystal structures of protein phosphatase-1 bound to motuporin and dihydromicrocystin-LA: Elucidation of the mechanism of enzyme inhibition by cyanobacterial toxins. J. Mol. Biol. 2006, 356, 111–120. [Google Scholar] [CrossRef]
- Tapia-Larios, C.; Mendoza, E.G.; Rubio-Armendáriz, M.C.; Olivero-Verbel, J. Identification of novel molecular targets of four microcystin variants by high-throughput virtual screening. Appl. Sci. 2022, 12, 2612. [Google Scholar] [CrossRef]
- Liu, L.; Xu, T.C.; Zhao, Z.A.; Zhang, N.N.; Li, J.; Chen, H.S. Toll-like receptor 4 signaling in neurons mediates cerebral ischemia/reperfusion injury. Mol. Neurobiol. 2022, 60, 864–874. [Google Scholar] [CrossRef]
- Mello, F.D.; Braidy, N.; Marçal, H.; Guillemin, G.; Nabavi, S.M.; Neilan, B.A. Mechanisms and effects posed by neurotoxic products of cyanobacteria/microbial eukaryotes/dinoflagellates in algae blooms: A review. Neurotox. Res. 2018, 33, 153–167. [Google Scholar] [CrossRef] [PubMed]
- Hinojosa, M.G.; Gutiérrez-Praena, D.; Prieto, A.I.; Guzmán-Guillén, R.; Jos, A.; Cameán, A.M. Neurotoxicity induced by microcystins and cylindrospermopsin: A review. Sci. Total Environ. 2019, 668, 547–565. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Shi, T.; Wang, Y.T.; He, J.; Zhao, X.; Wang, Y.K.; Giesy, J.P.; Chen, F.; Chen, Y.; Tuo, X.; et al. Effects of acute exposure to microcystins on hypothalamic-pituitary-adrenal (HPA), -gonad (HPG) and -thyroid (HPT) axes of female rats. Sci. Total Environ. 2021, 778, 145196. [Google Scholar] [CrossRef]
- Jin, H.; How, J.; Meng, X.; Ma, T.; Wang, B.; Liu, Z.; Sha, X.; Ding, J.; Han, X. Microcystin-leucine arginine induced the apoptosis of GnRH neurons by activating the endoplasmic reticulum stress resulting in a decrease of serum testosterone level in mice. Ecotoxicol. Environ. Saf. 2021, 208, 111748. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, H.; Du, X.; Chen, X.; Petlulu, P.; Tian, Z.; Shi, L.; Zhang, B.; Yuan, S.; Guo, X.; et al. A new identity of microcystins: Environmental endocrine disruptors? An evidence-based review. Sci. Total Environ. 2022, 851, 158262. [Google Scholar] [CrossRef] [PubMed]
- Casas-Rodriguez, A.; Cameán, A.M.; Jos, A. Potential endocrine disruption of cyanobacterial toxins, microcystins and cylindrospermopsin: A review. Toxins 2022, 14, 882. [Google Scholar] [CrossRef]
- Shi, T.; Xu, L.L.; Chen, L.; He, J.; Wang, Y.K.; Chen, F.; Chen, Y.; Giesy, J.P.; Wang, Y.T.; Wu, Q.H.; et al. Acute exposure to microcystins affects hypothalamic-pituitary axes of male rats. Environ. Pollut. 2023, 318, 120843. [Google Scholar] [CrossRef]
- Wei, L.; Sun, B.; Chang, M.; Liu, Y.; Nie, P. Effects of cyanobacterial toxin microcystin-LR on the transcription levels of immune-related genes in grass carp Ctenopharyngodon idella. Environ. Biol. Fish. 2009, 85, 231–238. [Google Scholar] [CrossRef]
- Rymuszka, A.; Adaszek, Ł. Cytotoxic effects and changes in cytokine gene expression induced by microcystin-containing extract in fish immune cells–An in vitro and in vivo study. Fish Shellfish Immun. 2013, 34, 1524–1532. [Google Scholar] [CrossRef]
- Saha, P.; Bose, D.; Stebliankin, V.; Cickovski, T.; Seth, R.K.; Porter, D.E.; Brooks, B.W.; Mathee, K.; Narasimhan, G.; Colwell, R.; et al. Prior exposure to microcystin alters host gut resistome and is associated with dysregulated immune homeostasis in translatable mouse models. Sci. Rep. 2022, 12, 11516. [Google Scholar] [CrossRef]
- Diez-Quijada, L.; Benítez-González, M.D.M.; Puerto, M.; Jos, A.; Cameán, A.M. Immunotoxic effects induced by microcystins and cylindrospermopsin: A review. Toxins 2021, 13, 711. [Google Scholar] [CrossRef]
- Yan, M.; Jin, H.; Pan, C.; Hang, H.; Li, D.; Han, X. Movement disorder and neurotoxicity induced by chronic exposure to microcystin-LR in mice. Mol. Neurobiol. 2022, 59, 5516–5531. [Google Scholar] [CrossRef] [PubMed]
- Goto, J.J.; Koenig, J.H.; Ikeda, K. The physiological effect of ingested β-N-methylamino-L-alanine on a glutamatergic synapse in an in vivo preparation. Comp. Biochem. Physiol. C Toxicol. Pharm. 2012, 156, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Okle, O.; Rath, L.; Galizia, C.G.; Dietrich, D.R. The cyanobacterial neurotoxin beta-N-methylamino-l-alanine (BMAA) induces neuronal and behavioral changes in honeybees. Toxicol. Appl. Pharm. 2013, 270, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Qiu, J.; Zhao, M.; Li, F.; Yu, R.; Li, A. Accumulation and distribution of neurotoxin BMAA in aquatic animals and effect on the behavior of zebrafish in a T-maze test. Toxicon 2020, 173, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Scott, L.; Downing, T. Dose-dependent adult neurodegeneration in a rat model after neonatal exposure to β-n-methylamino-l-alanine. Neurotox. Res. 2019, 35, 711–723. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, O.; Lindquist, N.G.; Brittebo, E.B.; Roman, E. Selective brain uptake and behavioral effects of the cyanobacterial toxin BMAA (β-N-methylamino-L-alanine) following neonatal administration to rodents. Toxicol. Sci. 2009, 109, 286–295. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, O.; Berg, A.-L.; Hanrieder, J.; Arnerup, G.; Lindström, A.-K.; Brittebo, E.B. Intracellular fibril formation, calcification, and enrichment of chaperones, cytoskeletal, and intermediate filament proteins in the adult hippocampus CA1 following neonatal exposure to the nonprotein amino acid BMAA. Arch. Toxicol. 2015, 89, 423–436. [Google Scholar] [CrossRef] [Green Version]
- Pierozan, P.; Karlsson, O. Mitotically heritable effects of BMAA on striatal neural stem cell proliferation and differentiation. Cell Death Dis. 2019, 10, 478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierozan, P.; Cattani, D.; Karlsson, O. Hippocampal neural stem cells are more susceptible to the neurotoxin BMAA than primary neurons: Effects on apoptosis, cellular differentiation, neurite outgrowth, and DNA methylation. Cell Death Dis. 2020, 11, 910. [Google Scholar] [CrossRef] [PubMed]
- Laugeray, A.; Oummadi, A.; Jourdain, C.; Feat, J.; Meyer-Dilhet, G.; Menuet, A.; Plé, K.; Gay, M.; Routier, S.; Mortaud, S.; et al. Perinatal exposure to the cyanotoxin β-N-méthylamino-l-alanine (BMAA) results in long-lasting behavioral changes in offspring—Potential involvement of DNA damage and oxidative stress. Neurotox. Res. 2018, 33, 87–112. [Google Scholar] [CrossRef]
- Méresse, S.; Larrigaldie, V.; Oummadi, A.; de Concini, V.; Morisset-Lopez, S.; Reverchon, F.; Menuet, A.; Montécot-Dubourg, C.; Mortaud, S. β-N-Methyl-Amino-L-Alanine cyanotoxin promotes modification of undifferentiated cells population and disrupts the inflammatory status in primary cultures of neural stem cells. Toxicology 2022, 482, 153358. [Google Scholar] [CrossRef]
- Li, X.; Zhang, X.; Ju, J.; Li, Y.; Yin, L.; Pu, Y. Maternal repeated oral exposure to microcystin-LR affects neurobehaviors in developing rats. Environ. Toxicol. Chem. 2015, 34, 64–69. [Google Scholar] [CrossRef]
- Montagne, A.; Zhao, Z.; Zlokovic, B.V. Alzheimer’s disease: A matter of blood-brain barrier dysfunction? J. Exp. Med. 2017, 214, 3151–3169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Bachari, S.; Naish, J.H.; Parker, G.J.; Emsley, H.C.; Parkes, L.M. Blood–brain barrier leakage is increased in Parkinson’s disease. Front. Physiol. 2020, 11, 593026. [Google Scholar] [CrossRef]
- Pan, Y.; Nicolazzo, J.A. Altered blood–brain barrier and blood–spinal cord barrier dynamics in amyotrophic lateral sclerosis: Impact on medication efficacy and safety. Br. J. Pharmacol. 2022, 179, 2577–2588. [Google Scholar] [CrossRef] [PubMed]
- Barisano, G.; Montagne, A.; Kisler, K.; Schneider, J.A.; Wardlaw, J.M.; Zlokovic, B.V. Blood–brain barrier link to human cognitive impairment and Alzheimer’s disease. Nat. Cardiovasc. Res. 2022, 1, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Smith, Q.R.; Nagura, H.; Takada, Y.; Duncan, M.W. Facilitated transport of the neurotoxin, β-N-methylamino-L-alanine, across the blood-brain barrier. J. Neurochem. 1992, 58, 1330–1337. [Google Scholar] [CrossRef]
- Myhre, O.; Eide, D.M.; Kleiven, S.; Utkilen, H.C.; Hofer, T. Repeated five-day administration of L-BMAA, microcystin-LR, or as mixture, in adult C57BL/6 mice-lack of adverse cognitive effects. Sci. Rep. 2018, 8, 2308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlsson, O.; Berg, A.L.; Lindström, A.K.; Hanrieder, J.; Arnerup, G.; Roman, E.; Bergquist, J.; Lindquist, N.G.; Brittebo, E.B.; Andersson, M. Neonatal exposure to the cyanobacterial toxin BMAA induces changes in protein expression and neurodegeneration in adult hippocampus. Toxicol. Sci. 2012, 130, 391–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, T.L.; Bergeron, C.; Biro, A.J.; Hansen, S. Chronic oral administration of β-N-methylamino-l-alanine is not neurotoxic to mice. J. Neurol. Sci. 1989, 94, 173–180. [Google Scholar] [CrossRef]
- Duncan, M.W.; Villacreses, N.E.; Pearson, P.G.; Wyatt, L.; Rapoport, S.I.; Kopin, I.J.; Markey, S.P.; Smith, Q.R. 2-Amino-3-(methylamino)-propanoic acid (BMAA) pharmacokinetics and blood-brain barrier permeability in the rat. J. Pharmacol. Exp. Ther. 1991, 258, 27–35. [Google Scholar]
- Cruz-Aguado, R.; Winkler, D.; Shaw, C.A. Lack of behavioural and neuropathological effects of dietary β-methylamino-l-alanine (BMAA) in mice. Pharmacol. Biochem. Behav. 2006, 84, 294–299. [Google Scholar] [CrossRef]
- Scott, L.L.; Downing, T.G. A single neonatal exposure to BMAA in a rat model produces neuropathology consistent with neurodegenerative diseases. Toxins 2017, 10, 22. [Google Scholar] [CrossRef] [Green Version]
- Garamszegi, S.P.; Banack, S.A.; Duque, L.L.; Metcalf, J.S.; Stommel, E.W.; Cox, P.A.; Davis, D.A. Detection of β-N-methylamino-L-alanine in postmortem olfactory bulbs of Alzheimer’s disease patients using UHPLC-MS/MS: An autopsy case-series study. Toxicol. Rep. 2023, 10, 87–96. [Google Scholar] [CrossRef]
- Fischer, W.J.; Altheimer, S.; Cattori, V.; Meier, P.J.; Dietrich, D.R.; Hagenbuch, B. Organic anion transporting polypeptides expressed in liver and brain mediate uptake of microcystin. Toxicol. Appl. Pharmacol. 2005, 203, 257–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondal, A.; Saha, P.; Bose, D.; Chatterjee, S.; Seth, R.K.; Xiao, S.; Porter, D.E.; Brooks, B.W.; Scott, G.I.; Nagarkatti, M.; et al. Environmental microcystin exposure in underlying NAFLD-induced exacerbation of neuroinflammation, blood-brain barrier dysfunction, and neurodegeneration are NLRP3 and S100B dependent. Toxicology 2021, 461, 152901. [Google Scholar] [CrossRef]
- Wang, J.; Chen, Y.; Zhang, C.; Xiang, Z.; Ding, J.; Han, X. Learning and memory deficits and Alzheimer’s disease-like changes in mice after chronic exposure to microcystin-LR. J. Hazardous Mater. 2019, 373, 504–518. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, C.; Zhu, J.; Ding, J.; Chen, Y.; Han, X. Blood-brain barrier disruption and inflammation reaction in mice after chronic exposure to microcystin-LR. Sci. Total Environ. 2019, 689, 662–678. [Google Scholar] [CrossRef] [PubMed]
- Feurstein, D.; Holst, K.; Fischer, A.; Dietrich, D.R. Oatp-associated uptake and toxicity of microcystins in primary murine whole brain cells. Toxicol. Appl. Pharmacol. 2009, 234, 247–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feurstein, D.; Kleinteich, J.; Heussner, A.H.; Stemmer, K.; Dietrich, D.R. Investigation of microcystin congener–dependent uptake into primary murine neurons. Environ. Health Perspect. 2010, 118, 1370–1375. [Google Scholar] [CrossRef]
- Rudrabhatla, P.; Albers, W.; Pant, H.C. Peptidyl-prolyl isomerase 1 regulates protein phosphatase 2A-mediated topographic phosphorylation of neurofilament proteins. J. Neurosci. 2009, 29, 14869–14880. [Google Scholar] [CrossRef] [Green Version]
- Yan, M.; Jin, H.; Pan, C.; Han, X. Chronic microcystin-LR-induced α-synuclein promotes neuroinflammation through activation of the NLRP3 inflammasome in microglia. Mol. Neurobiol. 2022, 60, 884–900. [Google Scholar] [CrossRef]
- Goedert, M. Alpha-synuclein and neurodegenerative diseases. Nat. Rev. Neurosci. 2001, 2, 492–501. [Google Scholar] [CrossRef]
- Bougea, A. Synuclein in neurodegeneration. Adv. Clin. Chem. 2001, 103, 97–134. [Google Scholar] [CrossRef]
- Verheijen, J.; Sleegers, K. Understanding Alzheimer disease at the interface between genetics and transcriptomics. Trends Genet. 2018, 34, 434–447. [Google Scholar] [CrossRef] [Green Version]
- Chiu, A.S.; Gehringer, M.M.; Braidy, N.; Guillemin, G.J.; Welch, J.H.; Neilan, B.A. Gliotoxicity of the cyanotoxin, β-methyl-amino-L-alanine (BMAA). Sci. Rep. 2013, 3, 1482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, A.S.; Braidy, N.; Marçal, H.; Welch, J.H.; Gehringer, M.M.; Guillemin, G.J.; Neilan, B.A. Global cellular responses to β-methyl-amino-l-alanine (BMAA) by olfactory ensheathing glial cells (OEC). Toxicon 2015, 99, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Bubik, A.; Sedmak, B.; Novinec, M.; Lenarčič, B.; Lah, T.T. Cytotoxic and peptidase inhibitory activities of selected non-hepatotoxic cyclic peptides from cyanobacteria. Biol. Chem. 2008, 389, 1339–1346. [Google Scholar] [CrossRef] [PubMed]
- D‘Mello, F.; Braidy, N.; Marçal, H.; Guillemin, G.; Rossi, F.; Chinian, M.; Laurent, D.; Teo, C.; Neilan, B.A. Cytotoxic effects of environmental toxins on human glial cells. Neurotox. Res. 2017, 31, 245–258. [Google Scholar] [CrossRef] [PubMed]
- Soto, T.; Buzzi, E.D.; Rotstein, N.P.; German, O.L.; Politi, L.E. Damaging effects of BMAA on retina neurons and Müller glial cells. Exp. Eye Res. 2021, 202, 108342. [Google Scholar] [CrossRef] [PubMed]
- Tan, V.X.; Mazzocco, C.; Varney, B.; Bodet, D.; Guillemin, T.A.; Bessede, A.; Guillemin, G.J. Detection of the cyanotoxins L-BMAA uptake and accumulation in primary neurons and astrocytes. Neurotox. Res. 2018, 33, 55–61. [Google Scholar] [CrossRef]
- Dias, F.R.P.; de Souza Almeida, R.R.; Sovrani, V.; Thomaz, N.K.; Gonçalves, C.A.; Quincozes-Santos, A.; Bobermin, L.D. Glioprotective effects of resveratrol against BMAA-induced astroglial dysfunctions. Neurotox. Res. 2022, 40, 530–541. [Google Scholar] [CrossRef]
- Takser, L.; Benachour, N.; Husk, B.; Cabana, H.; Gris, D. Cyanotoxins at low doses induce apoptosis and inflammatory effects in murine brain cells: Potential implications for neurodegenerative diseases. Toxicol. Rep. 2016, 3, 180–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forum on Neuroscience and Nervous System Disorders; Board on Health Sciences Policy; Institute of Medicine. Neurodegeneration: Exploring Commonalities across Diseases: Workshop Summary; National Academies Press: Washington, DC, USA, 2013. [Google Scholar] [CrossRef]
- Drummond, D.A.; Wilke, C.O. Mistranslation-induced protein misfolding as a dominant constraint on coding-sequence evolution. Cell 2008, 134, 341–352. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Miguel, V.E.; Lujan, C.; Espie, T.; Martinez-Martinez, D.; Moore, S.; Backes, C.; Gonzalez, S.; Galimov, E.R.; Brown, A.E.; Halic, M. Increased fidelity of protein synthesis extends lifespan. Cell Metab. 2021, 33, 2288–2300. [Google Scholar] [CrossRef]
- Brilkova, M.; Nigri, M.; Kumar, H.S.; Moore, J.; Mantovani, M.; Keller, C.; Grimm, A.; Eckert, A.; Shcherbakov, D.; Akbergenov, R.; et al. Error-prone protein synthesis recapitulates early symptoms of Alzheimer disease in aging mice. Cell Rep. 2022, 40, 111433. [Google Scholar] [CrossRef]
- Lant, J.T.; Kiri, R.; Duennwald, M.L.; O’Donoghue, P. Formation and persistence of polyglutamine aggregates in mistranslating cells. Nucleic Acids Res. 2021, 49, 11883–11899. [Google Scholar] [CrossRef] [PubMed]
- Tawfik, D.S.; Gruic-Sovulj, I. How evolution shapes enzyme selectivity–lessons from aminoacyl-tRNA synthetases and other amino acid utilizing enzymes. FEBS J. 2020, 287, 1284–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapur, M.; Ackerman, S.L. mRNA translation gone awry: Translation fidelity and neurological disease. Trends Genet. 2018, 34, 218–231. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yang, X.L.; Sun, L. The uniqueness of AlaRS and its human disease connections. RNA Biol. 2021, 18, 1501–1511. [Google Scholar] [CrossRef]
- Terrey, M.; Adamson, S.I.; Chuang, J.H.; Ackerman, S.L. Defects in translation-dependent quality control pathways lead to convergent molecular and neurodevelopmental pathology. Elife 2021, 10, e66904. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Zhou, H.; Vo, M.N.; Shi, Y.; Nawaz, M.H.; Vargas-Rodriguez, O.; Diedrich, J.K.; Yates, J.R.; Kishi, S.; Musier-Forsyth, K.; et al. Double mimicry evades tRNA synthetase editing by toxic vegetable-sourced non-proteinogenic amino acid. Nat. Commun. 2017, 8, 2281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodgers, K.J. Non-protein amino acids and neurodegeneration: The enemy within. Exp. Neurol. 2014, 253, 192–196. [Google Scholar] [CrossRef] [PubMed]
- Glover, W.; Mash, D.C.; Murch, S.J. The natural non-protein amino acid N-β-methylamino-L-alanine (BMAA) is incorporated into protein during synthesis. Amino Acids 2014, 46, 2553–2559. [Google Scholar] [CrossRef]
- Wang, Z.-Q.; Zhang, C.-C. A tRNA t6A modification system contributes to the sensitivity towards the toxin β-N-methylamino-L-alanine (BMAA) in the cyanobacterium Anabaena sp. PCC 7120. Aquat. Toxicol. 2022, 245, 106121. [Google Scholar] [CrossRef]
- Okamoto, S.; Esumi, S.; Hamaguchi-Hamada, K.; Hamada, S. β-N-methylamino-L-alanine (BMAA) suppresses cell cycle progression of non-neuronal cells. Sci. Rep. 2018, 8, 17995. [Google Scholar] [CrossRef] [Green Version]
- Meyer, J.L.; Gunasekera, S.P.; Brown, A.L.; Ding, Y.; Miller, S.; Teplitski, M.; Paul, V.J. Cryptic diversity of Black Band Disease cyanobacteria in Siderastrea siderea corals revealed by chemical ecology and comparative genome-resolved metagenomics. Mar. Drugs 2023, 21, 76. [Google Scholar] [CrossRef]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.; Sandhu, K.V.; Bastiaanssen, T.F.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The microbiota-gut-brain axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef] [PubMed]
- Morais, L.H.; Schreiber, H.L., IV; Mazmanian, S.K. The gut microbiota-brain axis in behavior and brain disorders. Nat. Rev. Microbiol. 2021, 19, 241–255. [Google Scholar] [CrossRef]
- Silva, D.F.; Candeias, E.; Esteves, A.R.; Magalhaes, J.D.; Fereira, I.L.; Nunes-Costa, D.; Rego, A.C.; Empadinhas, N.; Cardoso, S.M. Microbial BMAA elicits mitochondrial dysfunction, innate immunity activation, and Alzheimer’s disease features in cortical neurons. J. Neuroinflammation 2020, 17, 332. [Google Scholar] [CrossRef] [PubMed]
- Esteves, A.R.; Munoz-Pinto, M.F.; Nunes-Costa, D.; Candeias, E.; Silva, D.F.; Magalhães, J.D.; Pereira-Santos, A.R.; Ferreira, I.L.; Alarico, S.; Tiago, I.; et al. Footprints of a microbial toxin from the gut microbiome to mesencephalic mitochondria. Gut 2023, 72, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Li, A.; Qiu, J.; Yan, W.; Yan, C.; Zhang, L.; Li, M. Effects of the neurotoxin β-N-methylamino-L-alanine (BMAA) on the early embryonic development of marine shellfish and fish. Frontiers Mar. Sci. 2022, 9, 1033851. [Google Scholar] [CrossRef]



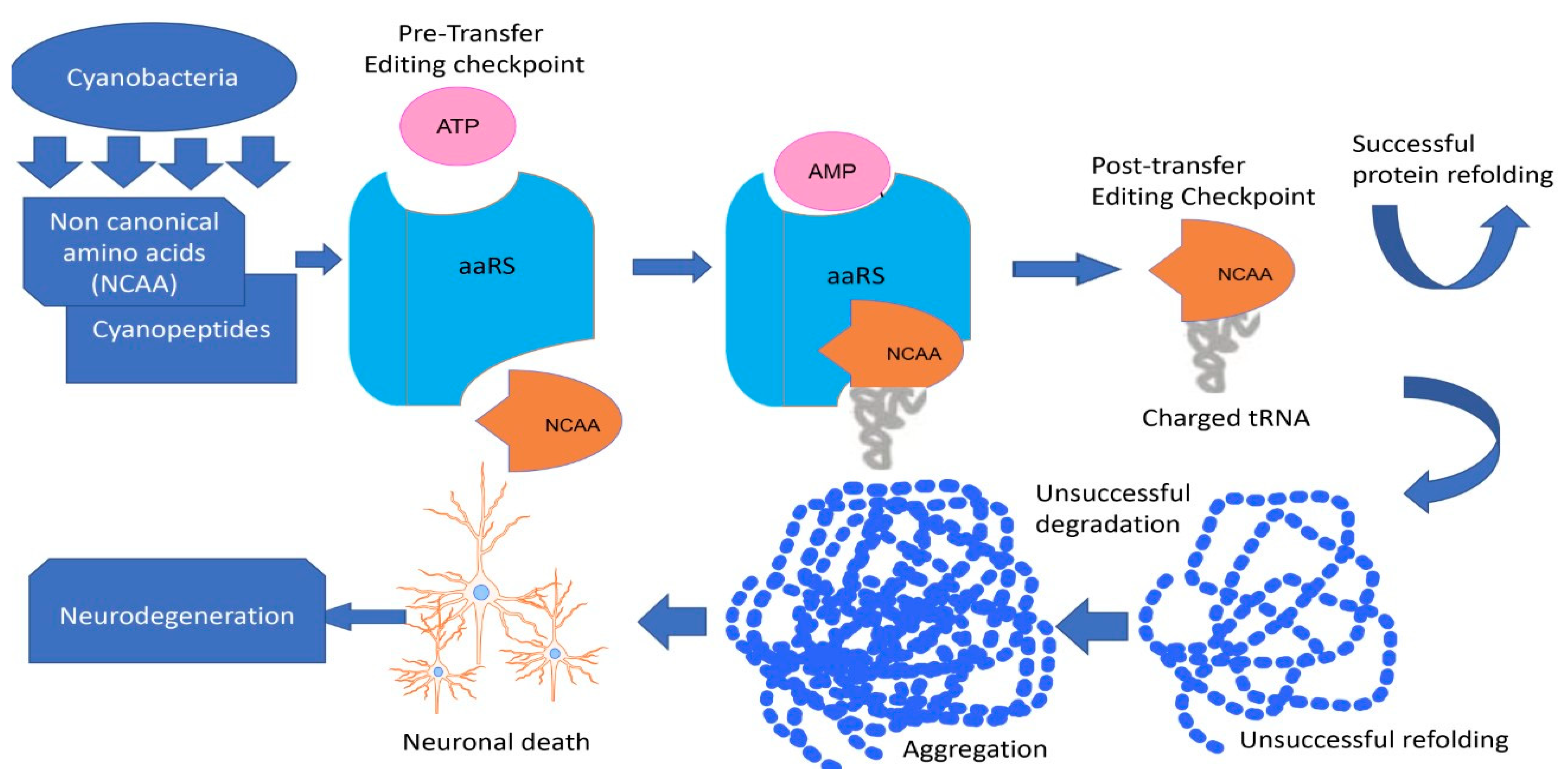{kind=link}
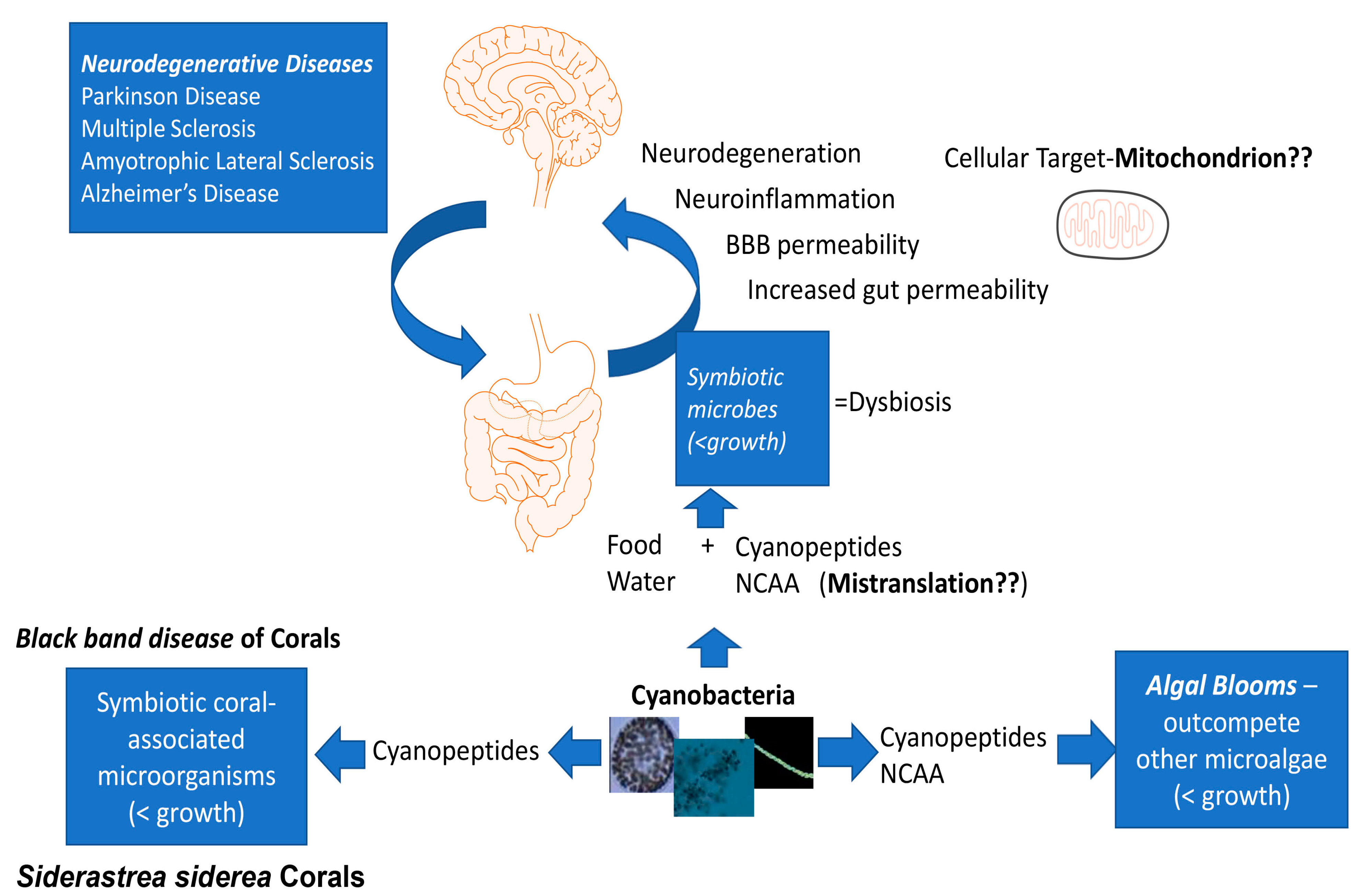{kind=link}
| Location | Period | ALS/PD Cases | Water Quality | Toxic Food or Dietary Components | Key Findings | Reference |
|---|---|---|---|---|---|---|
| Guam | 1945–1969 | 492 | Mn↑ | Cycad flour | Cycad toxic effect; Biochemical and neuropathologic abnormalities in ALS/PD diagnosed locals | [98] |
| Guam | 1940s–1960s | - | - | Cycad flour, flying foxes; food containing phytotoxins | Accumulation of cycad neurotoxins (BMAA, cycasin) in flying foxes; Flying foxes consumption → ALS-PDC | [99] |
| Guam and other Mariana Islands | 1956–1980 | 39 | - | - | Similar genotypic composition of Chamorros on all the Mariana Islands but different mortality rates of ALS/PD on Saipan than on Guam; Environmental factors of ALS > genetic | [100] |
| Guam, Canada | - | 23 | HABs: BMAA | Cycad flour, flying foxes (for Guam) | BMAA in tissues from frontal cortex; BMAA-containing food relates to ALS/PDC; HABs → cyanobacterial contamination water supplies →BMAA biomagnification | [101,102] |
| Kii Peninsula, Japan | 1961 | >4 | Ca, Mg, Na, KHCO3, Cl ↓ | - | Low mineral content in water supplies possibly leads to ND; > ALS—Mitogawa area | [103] |
| KII Peninsula, Japan | 1972 | 40 | Mn↑ | - | Possible association of Mn to ALS. | [104] |
| Skaraborg, Sweden | 1973–1984 | 70 males | - | - | Cluster of MND in Skaraborg; Agricultural occupation → MND risk. | [105] |
| Two Rivers, Small Wisconsin, USA | 1975–1983 | 6 | - | fish | Polychlorinated biphenyl Contaminated fish consumption → ALS risk. | [106] |
| France | 1975–1999 | 18 | - | - | ALS cluster in south-eastern France; Infections or environmental factors of ALS > genetic. | [107] |
| Italy | 1980–2001 | 634 | - | - | 16 ALS clusters; Low efficiency in detoxification systems; Environmental factors of ALS (toxins) | [108] |
| Finland | 1985–1995 | 576 | Pb, Cd, Zn↑ | - | Two ALS clusters; Environmental factors of ALS. | [109] |
| Enfield, NH, northeastern USA | 1990–2007 | 278 | HABs: BMAA, MC | Fish, shellfish | High ALS incidence near Lake Mascoma; Chronic exposure to cyanotoxins → ALS; Combined impact of multiple cyanotoxins. | [95] |
| Iraq, Saudi Arabia | 1991–2001 | 48 | BMAA | - | 48 ALS cases in Persian Gulf war veterans linked to desert’s crust contains BMAA; Aerosolization of cyanobacteria → inhalation of dust → BMAA exposure | [110,111,112] |
| Southern France, Hérault district | 1994–2009 | 381 | HABs: BMAA | shellfish | ALS cluster in Thau lagoon; Association with high concentrations of BMAA in mussels and oysters | [113] |
| Northern New England, USA | 1997–2009 | 688 | HABs, [CH3Hg]+ | - | 11 clusters of ALS grouped in 4 regions; Location of ALS cases are close to water bodies where HABs occurs; Environmental factors → ALS risk | [114] |
| Northern New England, USA | 1997–2009 | >800 | HABs | - | HABs → water-quality → ALS risk | [115] |
| Northern New England, USA | 1999–2009 | - | HABs: BMAA | - | Mapping cyanobacterial HABs for northern New England lakes; Cyanotoxins increase ALS risk. | [97] |
| Western NH, USA | - | - | HABs: BMAA | fish | High concentrations of BMAA and DAB were found in the Lake Mascoma fish; BMAA, DAB, AEG in the air filters; ALS linked to BMAA. | [96] |
| France | 2003–2011 | 72 | HABs: BMAA | Nine ALS clusters; ALS linked to BMAA. | [116] | |
| South Korea | 2005–2017 | - | HABs: BMAA mycrocystin, and other cyanotoxins | - | HABs severity → ND occurrence; HABs→ long-term impacts on human health | [117] |
| Guadeloupe | 1996–2011 | 63 | - | - | The highest incidence of ALS - Marie-Galante island; Environmental factor(s) → ALS risk | [118] |
| Northern and Southern Italy | 2002–2012 | 95 | - | dietary supplements | Private wells using → ALS risk↑; Amino acid supplements → ALS risk | [119] |
| Annapolis, Maryland, USA | 2013 | 3 | HABs: BMAA | blue crab | High concentrations of BMAA in the crabs originated Chesapeake Bay HABs exposure → ALS occurrence | [120] |
| № | Cyanotoxins | Detection Techniques | Sensivity | Reference | |
|---|---|---|---|---|---|
| LOD | LOQ | ||||
| 1. | MC-LR and 2 congeners | UHPLC-MS/MS | 0.02–0.04 ng/mL | - | [185] |
| 2. | MC-LR and 11 congeners | UHPLC-MS/MS | - | 0.2 µg/L | [186] |
| 3. | MC-LR and 4 congeners | LC-MS/MS | 0.005–0.0817 µg/L | 0.005–0.0817 µg/L | [187] |
| Nodularin | 0.0048 µg/L | 0.0048 µg/L | |||
| Anatoxin-a | 0.0001 µg/L | 0.0004 µg/L | |||
| Cylindrospermopsin | 0.0001 µg/L | 0.0004 µg/L | |||
| 4. | BMAA | UHPLC-MS/MS | 0.02 pg/µL | 0.05 pg/µL | [188] |
| 2,4-DAB | 0.04 pg/µL | 0.13 pg/µL | |||
| 5. | MC-LR and 7 congeners | LC-MS/MS | - | 0.04–0.5 µg/L | [189] |
| Anatoxin-a | - | 0.02 µg/L | |||
| Cylindrospermopsin (and deoxyCYN) | - | 0.01–0.02 µg/L | |||
| Saxitoxins (4 congeners), GTX (5 congeners), decarbamoylgonyautoxin, N-sulfogonyautoxins-1 and -2 | - | 0.1–2 µg/L | |||
| 6. | MC-LR and 11 congeners | HPLC-MS/MS | 0.01 ± 0.01–0.19 ± 0.2 μg/L | 0.04 ± 0.04–0.64 ± 0.65 μg/L | [190] |
| Nodularin | 0.04 ± 0.02 μg/L | 0.13 ± 0.06 μg/L | |||
| 7. | MC-LR and 2 congeners | HPLC-UV/PDA | 3–4 μg/L | 9–13 μg/L | [191] |
| 8. | MC-LR and 2 congeners | HPLC-HRMS | 0.002 μg/L | - | [192] |
| 9. | Anatoxin-a, | HILIC-MS/MS | 0.004 ng/mL | 0.01 ng/mL | [193] |
| Cylindrospermopsin | 0.07 ng/mL | 0.23 ng/mL | |||
| Saxitoxin | 0.01 ng/mL | 0.04 ng/mL | |||
| MC-LR and 4 congeners | RPLC- MS/MS | 0.02–0.08 ng/mL | 0.07–0.28 ng/mL | ||
| Nodularin | 0.05 ng/mL | 0.18 ng/mL | |||
| 10. | MC-LR and 5 congeners | UHPLC-MS/MS | - | 0.046–0.052 µg/L | [194] |
| Nodularin | - | 0.049 µg/L | |||
| Cylindrospermopsin | - | 0.052 µg/L | |||
| 11. | BMAA | UHPLC-HRMS | 5 ng/L | 10 ng/L | [195] |
| DAB | 3 ng/L | 5 ng/L | |||
| AEG | 2 ng/L | 5 ng/L | |||
| BAMA | 5 ng/L | 10 ng/L | |||
| 12. | MC-LR and 5 congeners | HPLC-MS/MS | 0.0003–0.0009 µg/L | - | [196] |
| Cylindrospermopsin | 0.0005 µg/L | - | |||
| Saxitoxin, dcSTX | 0.0009–0.0013 µg/L | - | |||
| 13. | BMAA | LC-MS/MS | 10 ng/L | - | [197] |
| 14. | MC-LR and 5 congeners | LC-MS/MS | 0.04–0.8 μg/L | 0.1–2.3 μg/L | [198] |
| Nodularin | 0.3 μg/L | 0.9 μg/L | |||
| Anatoxin-a | 0.27 μg/L | 0.81 μg/L | |||
| Cylindrospermopsin | 0.14 μg/L | 0.4 μg/L | |||
| 15. | MC-LR and 2 congeners | HPLC-DAD | 0.08–0.15 µg/L | - | [199] |
| 16. | MC-LR and 11 congeners | LC-MS/MS | 0.001–0.007 μg/L | 0.003–0.020 μg/L | [200] |
| Nodularin | 0.002 μg/L | 0.006 μg/L | |||
| Anatoxin-a | 0.001 μg/L | 0.003 μg/L | |||
| Cylindrospermopsin | 0.001 μg/L | 0.003 μg/L | |||
| 17. | MCs | UPLC-MS/MS | 0.005 µg/L | - | [201] |
| Anatoxin-a | 0.02 µg/L | - | |||
| Cylindrospermopsin | 0.02 µg/L | - | |||
| Saxitoxin | 0.8 µg/L | - | |||
| BMAA | 0.03 µg/L | - | |||
| 18. | BMAA | LC-MS/MS | 0.030 μg/L | 0.096 μg/L | [202] |
| 19. | MC-LR and 11 congeners | on-line SPE – UHPLC-HRMS | 5–37 ng/L | 15–130 ng/L | [203] |
| Anatoxin-a | 15–18 ng/L | 50–60 ng/L | |||
| Homoanatoxin-a | 11–12 ng/L | 30–40 ng/L | |||
| Cylindrospermopsin | 41–53 ng/L | 130–170 ng/L | |||
| 20. | MC-LR and 1 congener | HPLC-DAD | 0.2-0.3 μg/L | - | [204] |
| 21. | Anatoxin-a | UHPLC-MS/MS | 1.1 ng/L | 2.5 ng/L | [205] |
| Cylindrospermopsin | 10.9 ng/L | 21.7 ng/L | |||
| Saxitoxins (4 congeners) | 3.5-9 ng/L | 7.1–26.9 ng/L | |||
| GTX (7 congeners) | 18.5–54.5 ng/L | 42.2–227.6 ng/L | |||
| 22. | MC-LR and 7 congeners | UHPLC-MS/MS | 0.1 µg/L | 0.5 µg/L | [181] |
| Nodularin | 0.1 µg/L | 0.5 µg/L | |||
| 23. | MC-LR and 2 congeners | UHPLC-MS/MS | 0.1 µg/L | 24 µg/L | [206] |
| 24. | Cylindrospermopsin | UHPLC-MS/MS | 0.029 μg/L | 0.091 μg/L | [207] |
| 25. | Saxitoxins (4 congeners) | on-line SPE–HILIC-HRMS | 0.72–3.9 ng/L | 2.4–13 ng/L | [208] |
| 26. | MC-LR and 1 congener | tandem-SPE-HILIC-MS/MS | 0.0012–0.0021 μg/L | 0.004–0.007 μg/L | [209] |
| Nodularin | 0.0021 μg/L | 0.007 μg/L | |||
| Anatoxin-a | 0.03 μg/L | 0.1 μg/L | |||
| Cylindrospermopsin | 0.0012 μg/L | 0.004 μg/L | |||
| BMAA | 0.015 μg/L | 0.05 μg/L | |||
| DAB | 0.009 μg/L | 0.03 μg/L | |||
| AEG | 0.006 μg/L | 0.02 μg/L | |||
| 27. | BMAA | LC-MS/MS | 2.8 ng/mL | - | [210] |
| DAB | 1.7 ng/mL | - | |||
| 28. | BMAA | on-line SPE-UHPLC-HRMS | 10 ng/L | - | [211] |
| BAMA | 10 ng/L | - | |||
| DAB | 10 ng/L | - | |||
| AEG | 5 ng/L | - | |||
| 29. | BMAA | UHPLC-MS/MS | - | 2.5 µg/L | [212] |
| AEG | - | 2.5 µg/L | |||
| DABA | - | 5 µg/L | |||
| 30. | MC-LR and 7 congeners | UHPLC-MS/MS (ESI) | 0.02–0.2 µg/L | 0.05–0.5 µg/L | [213] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nugumanova, G.; Ponomarev, E.D.; Askarova, S.; Fasler-Kan, E.; Barteneva, N.S. Freshwater Cyanobacterial Toxins, Cyanopeptides and Neurodegenerative Diseases. Toxins 2023, 15, 233. https://doi.org/10.3390/toxins15030233
Nugumanova G, Ponomarev ED, Askarova S, Fasler-Kan E, Barteneva NS. Freshwater Cyanobacterial Toxins, Cyanopeptides and Neurodegenerative Diseases. Toxins. 2023; 15(3):233. https://doi.org/10.3390/toxins15030233
Chicago/Turabian StyleNugumanova, Galina, Eugene D. Ponomarev, Sholpan Askarova, Elizaveta Fasler-Kan, and Natasha S. Barteneva. 2023. "Freshwater Cyanobacterial Toxins, Cyanopeptides and Neurodegenerative Diseases" Toxins 15, no. 3: 233. https://doi.org/10.3390/toxins15030233
APA StyleNugumanova, G., Ponomarev, E. D., Askarova, S., Fasler-Kan, E., & Barteneva, N. S. (2023). Freshwater Cyanobacterial Toxins, Cyanopeptides and Neurodegenerative Diseases. Toxins, 15(3), 233. https://doi.org/10.3390/toxins15030233