Xeomin®, a Commercial Formulation of Botulinum Neurotoxin Type A, Promotes Regeneration in a Preclinical Model of Spinal Cord Injury

 ,
,  and
and 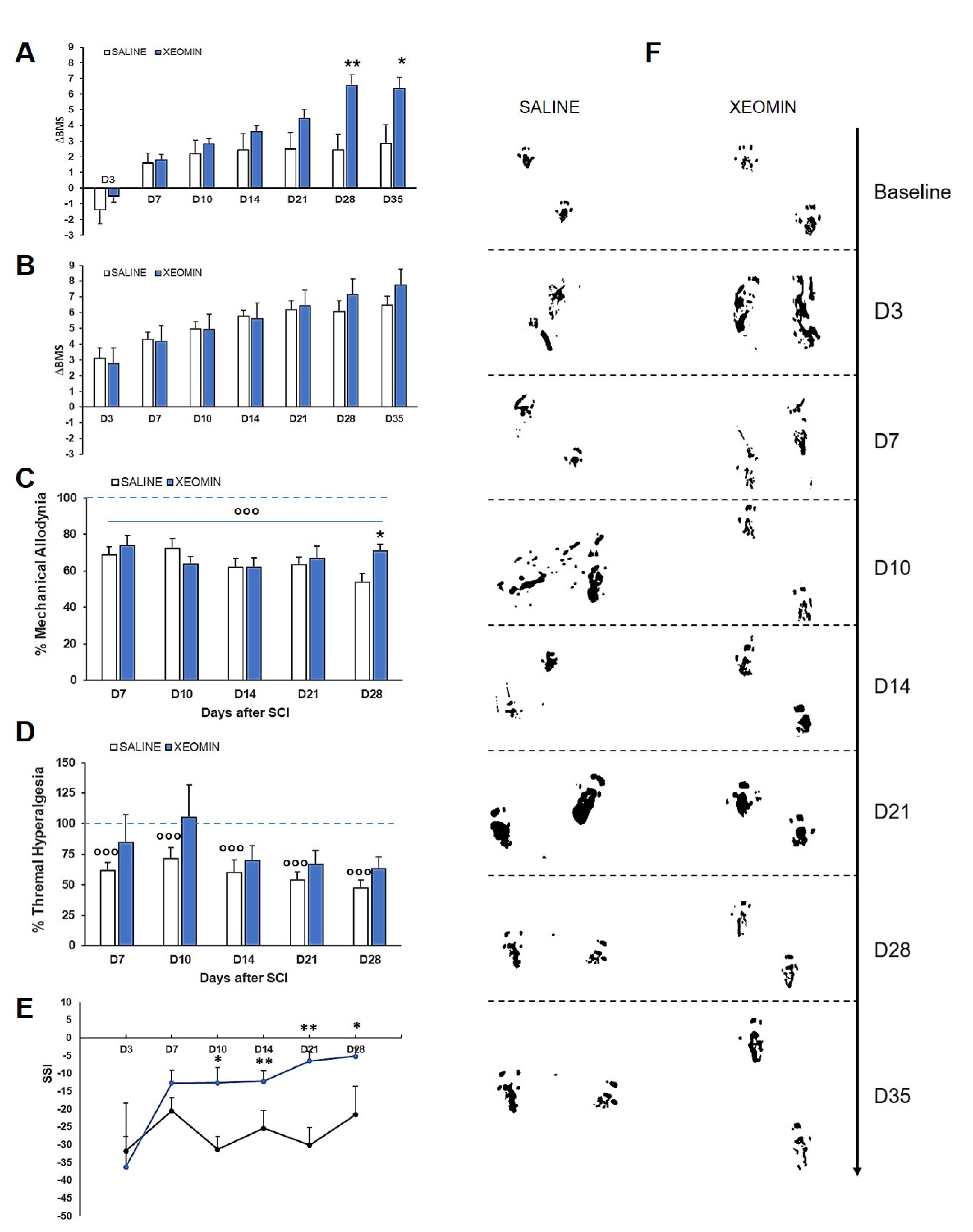{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
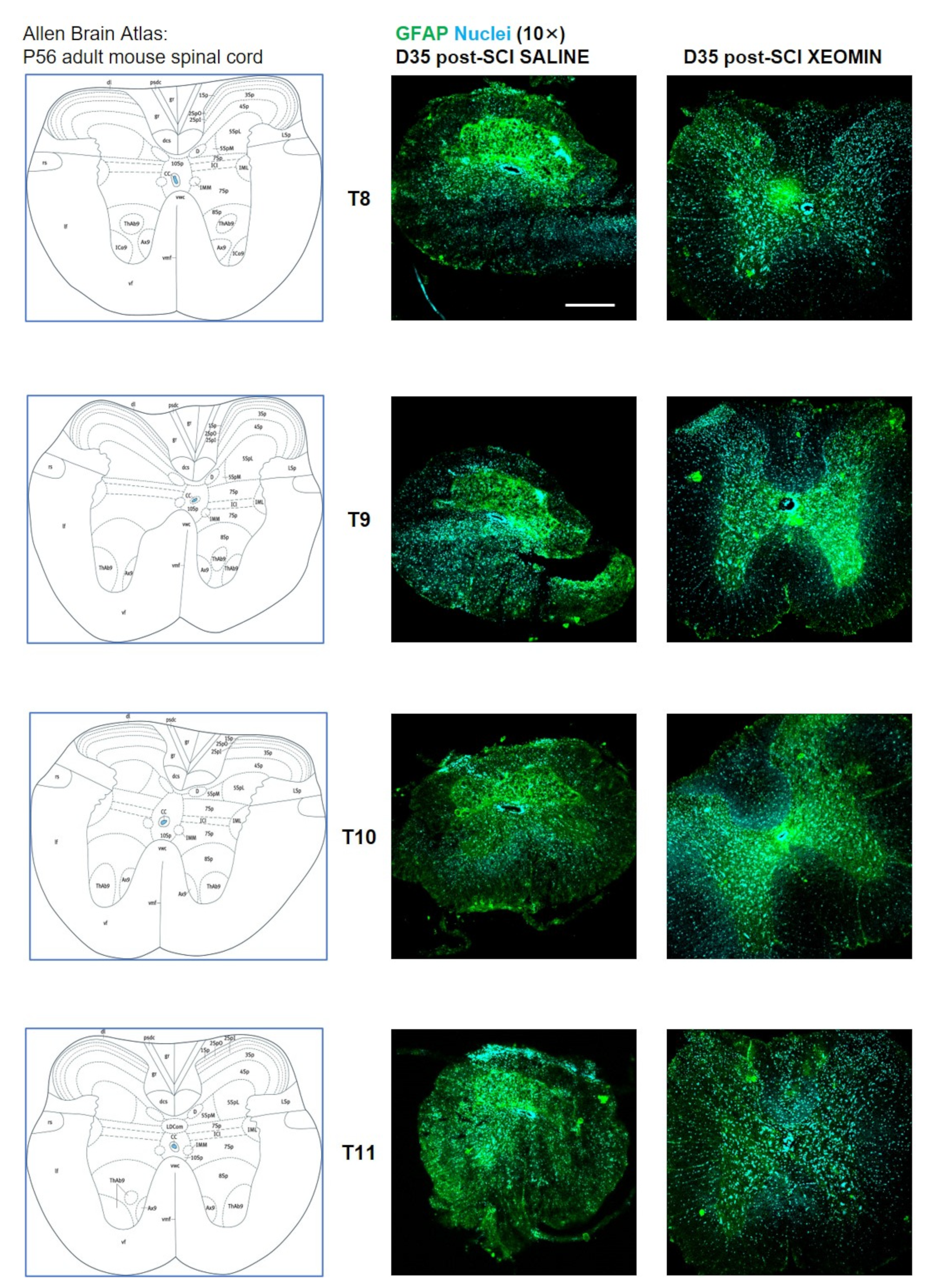
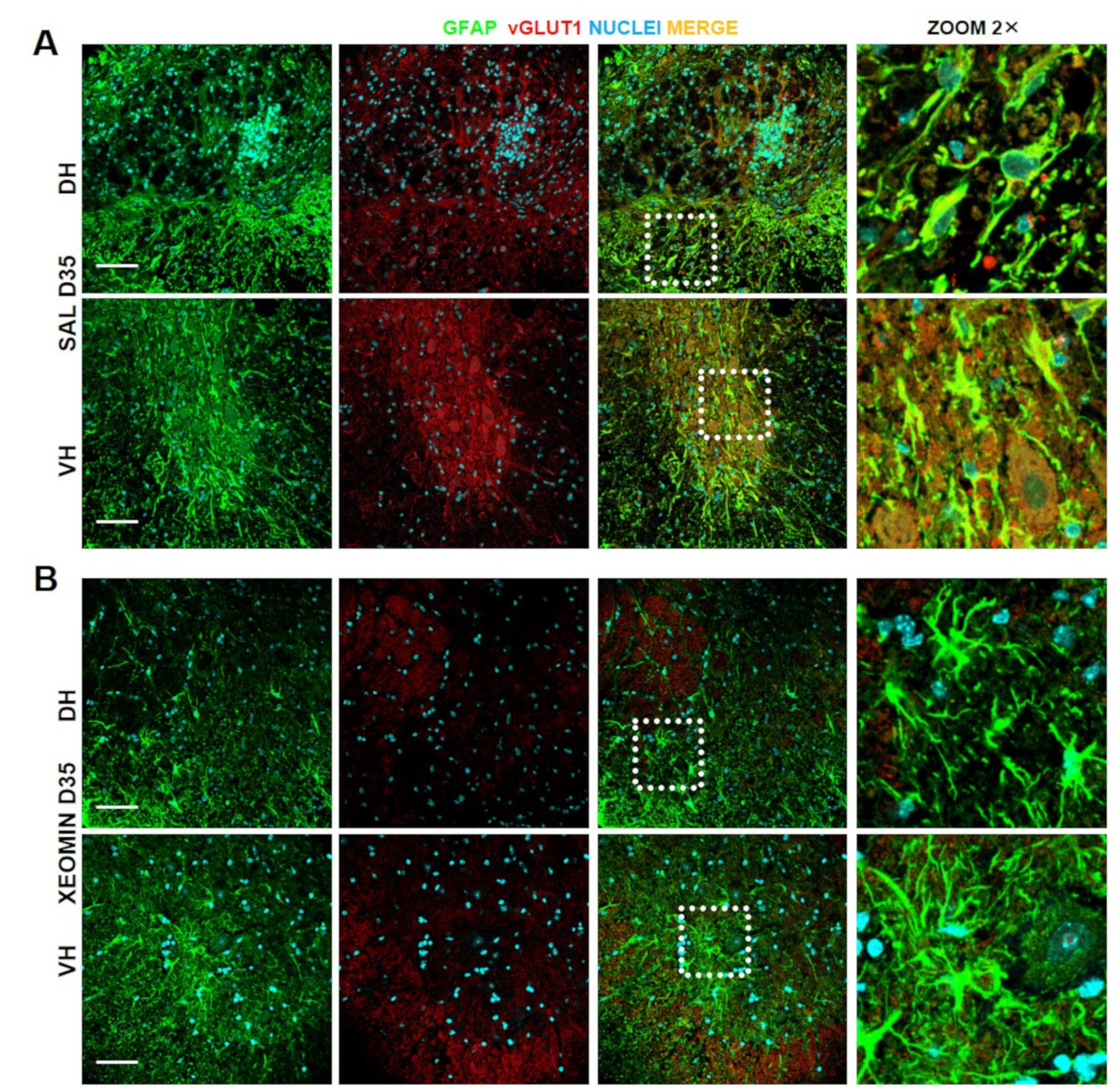
2. Results
3. Discussion and Conclusions
4. Materials and Methods
4.1. Animals
4.2. Surgery
4.3. Drugs
4.4. Behavioral Test: Basso Mouse Scale (BMS)
4.5. Nociceptive Tests
4.6. Sciatic Static Index
4.7. Immunohistochemistry and Confocal Images
4.8. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Montecucco, C.; Schiavo, G. Structure and function of tetanus and botulinum neurotoxins. Q. Rev. Biophys. 1995, 28, 423–472. [Google Scholar] [CrossRef]
- Masuyer, G.; Zhang, S.; Barkho, S.; Shen, Y.; Henriksson, L.; Kosenina, S.; Dong, M.; Stenmar, P. Structural characterisation of the catalytic domain of botulinum neurotoxin X—High activity and unique substrate specificity. Sci. Rep. 2018, 8, 4518. [Google Scholar] [CrossRef] [Green Version]
- Rossetto, O.; Pirazzini, M.; Montecucco, C. Botulinum neurotoxins: Genetic, structural and mechanistic insights. Nat. Rev. Microbiol. 2014, 12, 535–549. [Google Scholar] [CrossRef] [PubMed]
- Steward, L.; Brin, M.F.; Brideau-Andersen, A. Novel Native and Engineered Botulinum Neurotoxins. Handb. Exp. Pharmacol. 2021, 263, 63–89. [Google Scholar] [PubMed]
- Lacy, D.B.; Tepp, W.; Cohen, A.C.; DasGupta, B.R.; Stevens, R.C. Crystal structure of botulinum neurotoxin type A and implications for toxicity. Nat. Struct. Biol. 1998, 5, 898–902. [Google Scholar] [CrossRef]
- Schiavo, G.; Poulain, B.; Rossetto, O.; Benfenati, F.; Tauc, L.; Montecucco, C. Tetanus toxin is a zinc protein and its inhibition of neurotransmitter release and protease activity depend on zinc. EMBO J. 1992, 11, 3577–3583. [Google Scholar] [CrossRef] [PubMed]
- Söllner, T.; Whiteheart, S.W.; Brunner, M.; Erdjument-Bromage, H.; Geromanos, S.; Tempst, P.; Rothman, J.E. SNAP receptors implicated in vesicle targeting and fusion. Nature 1993, 362, 318–324. [Google Scholar] [CrossRef]
- Pirazzini, M.; Rossetto, O.; Eleopra, R.; Montecucco, C. Botulinum neurotoxins: Biology, pharmacology, and toxicology. Pharmacol. Rev. 2017, 69, 200–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jankovic, J. Botulinum toxin: State of the art. Mov. Disord. 2017, 32, 1131–1138. [Google Scholar] [CrossRef]
- Choudhury, S.; Baker, M.R.; Chatterjee, S.; Kumar, H. Botulinum Toxin: An Update on Pharmacology and Newer Products in Development. Toxins 2021, 13, 58. [Google Scholar] [CrossRef] [PubMed]
- Spiegel, L.L.; Ostrem, J.L.; Bledsoe, I.O. FDA Approvals and Consensus Guidelines for Botulinum Toxins in the Treatment of Dystonia. Toxins 2020, 12, 332. [Google Scholar] [CrossRef]
- Lewandowski, M.; Świerczewska, Z.; Barańska-Rybak, W. Off-Label Use of Botulinum Toxin in Dermatology-Current State of the Art. Molecules 2022, 27, 3143. [Google Scholar] [CrossRef]
- Yi, K.H.; Lee, J.H.; Hu, H.W.; Kim, H.J. Anatomical Proposal for Botulinum Neurotoxin Injection for Glabellar Frown Lines. Toxins 2022, 14, 268. [Google Scholar] [CrossRef]
- Jabbari, B. Botulinum neurotoxins in the treatment of refractory pain. Nat. Clin. Pract. Neurol. 2008, 4, 676–685. [Google Scholar] [CrossRef]
- Pavone, F.; Luvisetto, S. Botulinum neurotoxin for pain management: Insights from animal models. Toxins 2010, 2, 2890–2913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matak, I.; Lacković, Z. Botulinum toxin A, brain and pain. Prog. Neurobiol. 2014, 119–120, 39–59. [Google Scholar] [CrossRef] [Green Version]
- Matak, I.; Bölcskei, K.; Bach-Rojecky, L.; Helyes, Z. Mechanisms of Botulinum Toxin Type A Action on Pain. Toxins 2019, 11, 459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luvisetto, S.; Gazerani, P.; Cianchetti, C.; Pavone, F. Botulinum Toxin Type A as a Therapeutic Agent against Headache and Related Disorders. Toxins 2015, 7, 3818–3844. [Google Scholar] [CrossRef] [Green Version]
- Egeo, G.; Fofi, L.; Barbanti, P. Botulinum Neurotoxin for the Treatment of Neuropathic Pain. Front. Neurol. 2020, 11, 716. [Google Scholar] [CrossRef]
- Reyes-Long, S.; Alfaro-Rodríguez, A.; Cortes-Altamirano, J.L.; Lara-Padilla, E.; Herrera-Maria, E.; Romero-Morelos, P.; Salcedo, M.; Bandala, C. The Mechanisms of Action of Botulinum Toxin Type A in Nociceptive and Neuropathic Pathways in Cancer Pain. Curr. Med. Chem. 2021, 28, 2996–3009. [Google Scholar] [CrossRef] [PubMed]
- Frampton, J.E.; Silberstein, S. OnabotulinumtoxinA: A Review in the Prevention of Chronic Migraine. Drugs 2018, 78, 589–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burstein, R.; Blumenfeld, A.M.; Silberstein, S.D.; Manack Adams, A.; Brin, M.F. Mechanism of Action of OnabotulinumtoxinA in Chronic Migraine: A Narrative Review. Headache 2020, 60, 1259–1272. [Google Scholar] [CrossRef]
- Raciti, L.; Raciti, G.; Militi, D.; Casella, C.; Calabrò, R.S. Chronic Migraine: A Narrative Review on the Use of Botulinum Toxin with Clinical Indications and Future Directions. J. Integr. Neurosci. 2022, 21, 141. [Google Scholar] [CrossRef]
- Vacca, V.; Madaro, L.; De Angelis, F.; Proietti, D.; Cobianchi, S.; Orsini, T.; Puri, P.L.; Luvisetto, S.; Pavone, F.; Marinelli, S. Revealing the Therapeutic Potential of Botulinum Neurotoxin Type A in Counteracting Paralysis and Neuropathic Pain in Spinally Injured Mice. Toxins 2020, 12, 491. [Google Scholar] [CrossRef]
- Marinelli, S.; Vacca, V.; De Angelis, F.; Pieroni, L.; Orsini, T.; Parisi, C.; Soligo, M.; Protto, V.; Manni, L.; Guerrieri, R.; et al. Innovative mouse model mimicking human-like features of spinal cord injury: Efficacy of Docosahexaenoic acid on acute and chronic phases. Sci. Rep. 2019, 9, 8883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kundi, S.; Bicknell, R.; Ahmed, Z. Spinal cord injury: Current mammalian models. Am. J. Neurosci. 2013, 4, 1–12. [Google Scholar] [CrossRef]
- Mills, C.D.; Fullwood, S.D.; Hulsebosch, C.E. Changes in Metabotropic Glutamate receptor expression following spinal cord injury. Exp. Neurol. 2001, 170, 244–257. [Google Scholar] [CrossRef] [PubMed]
- Yiu, G.; He, Z. Glial inhibition of CNS axon regeneration. Nat. Rev. Neurosci. 2006, 7, 617–627. [Google Scholar] [CrossRef] [Green Version]
- Karimi-Abdolrezaee, S.; Billakanti, R. Reactive Astrogliosis after spinal cord injury—Beneficial and detrimental effects. Mol. Neurobiol. 2012, 46, 251–264. [Google Scholar] [CrossRef]
- Hausmann, O.N. Post-traumatic inflammation following spinal cord injury. Spinal Cord 2003, 41, 369–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ju, G.; Wang, J.; Wang, Y.; Zhao, X. Spinal cord contusion. Neural Regen. Res. 2014, 9, 789–794. [Google Scholar] [CrossRef] [PubMed]
- Walters, E.T. Neuroinflammatory contributions to pain after SCI: Roles for central glial mechanisms and nociceptor-mediated host defense. Exp. Neurol. 2014, 258, 48–61. [Google Scholar] [CrossRef]
- Rupp, D.; Nicholson, G.; Canty, D.; Wang, J.; Rhéaume, C.; Le, L.; Steward, L.E.; Washburn, M.; Jacky, B.P.; Broide, R.S.; et al. OnabotulinumtoxinA Displays Greater Biological Activity Compared to IncobotulinumtoxinA, Demonstrating Non-Interchangeability in Both In Vitro and In Vivo Assays. Toxins 2020, 12, 393. [Google Scholar] [CrossRef] [PubMed]
- Car, H.; Bogucki, A.; Bonikowski, M.; Dec-Ćwiek, M.; Drużdż, A.; Koziorowski, D.; Rudzińska-Bar, M.; Sarzyńska-Długosz, I.; Sławek, J. Botulinum toxin type-A preparations are not the same medications—Basic science (Part 1). Neurol. Neurochir. Pol. 2021, 55, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Byun, J.; Kwak, S.; Kwon, J.H.; Shin, M.; Lee, D.K.; Rhee, C.H.; Kang, W.H.; Oh, J.W.; Cruz, D.J.M. Comparative Pharmacodynamics of Three Different Botulinum Toxin Type A Preparations following Repeated Intramuscular Administration in Mice. Toxins 2022, 14, 365. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, M. Ethical guidelines for investigations of experimental pain in conscious animals. Pain 1983, 16, 109–110. [Google Scholar] [CrossRef] [PubMed]
- Luvisetto, S.; Marinelli, S.; Cobianchi, S.; Pavone, F. Anti-allodynic efficacy of botulinum neurotoxin A in a model of neuropathic pain. Neuroscience 2007, 145, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Vacca, V.; Luvisetto, S.; Marinelli, S.; Pavone, F. Botulinum toxin A increases analgesic effects of morphine, counters development of morphine tolerance and modulates glia activation and μ opioid receptor expression in neuropathic mice. Brain Behav. Immun. 2013, 32, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Cobianchi, S.; Marinelli, S.; Florenzano, F.; Pavone, F.; Luvisetto, S. Short but not long-lasting treadmill running reduces allodynia and improves functional recovery after peripheral nerve injury. Neuroscience 2010, 168, 273–287. [Google Scholar] [CrossRef]
- Baptista, A.F.; de Souza Gomes, J.R.; Oliveira, J.T.; Santos, S.M.; Vannier-Santos, M.A.; Martinez, A.M. A new approach to assess function after sciatic nerve lesion in the mouse adaptation of the sciatic static index. J. Neurosci. Methods 2007, 161, 259–264. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mastrorilli, V.; De Angelis, F.; Vacca, V.; Pavone, F.; Luvisetto, S.; Marinelli, S. Xeomin®, a Commercial Formulation of Botulinum Neurotoxin Type A, Promotes Regeneration in a Preclinical Model of Spinal Cord Injury. Toxins 2023, 15, 248. https://doi.org/10.3390/toxins15040248
Mastrorilli V, De Angelis F, Vacca V, Pavone F, Luvisetto S, Marinelli S. Xeomin®, a Commercial Formulation of Botulinum Neurotoxin Type A, Promotes Regeneration in a Preclinical Model of Spinal Cord Injury. Toxins. 2023; 15(4):248. https://doi.org/10.3390/toxins15040248
Chicago/Turabian StyleMastrorilli, Valentina, Federica De Angelis, Valentina Vacca, Flaminia Pavone, Siro Luvisetto, and Sara Marinelli. 2023. "Xeomin®, a Commercial Formulation of Botulinum Neurotoxin Type A, Promotes Regeneration in a Preclinical Model of Spinal Cord Injury" Toxins 15, no. 4: 248. https://doi.org/10.3390/toxins15040248
APA StyleMastrorilli, V., De Angelis, F., Vacca, V., Pavone, F., Luvisetto, S., & Marinelli, S. (2023). Xeomin®, a Commercial Formulation of Botulinum Neurotoxin Type A, Promotes Regeneration in a Preclinical Model of Spinal Cord Injury. Toxins, 15(4), 248. https://doi.org/10.3390/toxins15040248