In Vitro High-Throughput Genotoxicity Testing Using γH2AX Biomarker, Microscopy and Reproducible Automatic Image Analysis in ImageJ—A Pilot Study with Valinomycin



Abstract
:1. Introduction
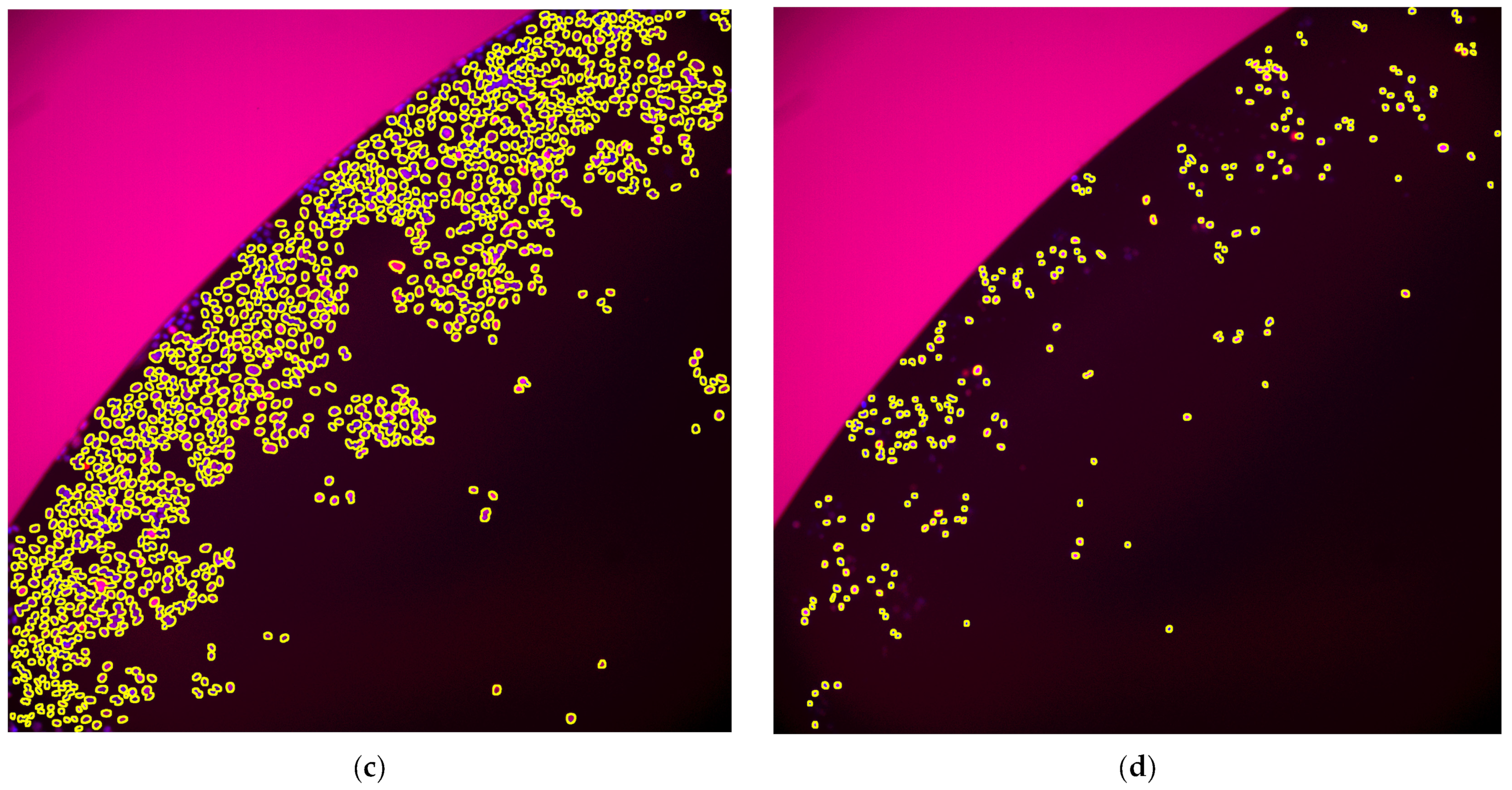
2. Results
3. Discussion
- We are aware of the fact that for the measurement of γH2AX response, cell lines derived from normal tissues are considered to be more reliable than those from cancer tissues [24]. Nevertheless, HeLa cell line was used as a model since it is recommended in the manual of the kit’s manufacturer. The CHO-K1 cell line was used as the second cell type because it is a cell line used for the HPRT mutation assay accepted by OECD and EPA.
- As a model compound, we only used valinomycin because it was used by the manufacturer of the kit. There was, therefore, no need to use metabolic activation, because valinomycin is genotoxic per se.
- To accelerate image acquisition and to minimize the potential risk of channel-to-channel leakage, we presumed that dead cells were washed away during the preparation of the samples and, therefore, we did not use any additional dye to detect dead cells.
- We used nuclei area, not number of nuclei, to evaluate cell mass.
4. Conclusions
5. Materials and Methods
5.1. Chemicals
5.2. Cell Lines and Culture Conditions
5.3. Direct Measurement of DNA DSBs
5.4. Measurement Settings
- Manufacturer and model of microscope: Olympus IX83 P2ZF;
- Objective lens magnification: 10×; NA = 0.3;
- Excitation filters (mounted in the light source);
- Violet: 395/25 nm; LED module 1, DAPI;
- Green: 555/28 nm; LED module 5, Cy3;
- Quad band filter set for DAPI/FITC/Cy3/Cy5;
- Quad band polychroic mirror (mounted in the filter turret);
- BP 411–454 nm;
- BP 495–536 nm;
- BP 577–617 nm;
- BP 655–810 nm;
- Emission filters (mounted in the fast emission filter wheel, in front of the camera);
- DAPI: BP 421–445 nm;
- Cy3: BP 581–619 nm;
- Illumination light source: Lumencor Spectra X Lamp;
- Camera manufacturer and model: Hamamatsu ORCA-Flash4.0;
- Pixel size: 650 nm × 650 nm;
- Software program(s) and version: OLYMPUS cellSens Dimension 3.2 (Build 23706);
- Image acquisition settings including exposure times, gain, and binning: exp 500 ms, gain: 0, binning: 4 × 4;
- Experiment manager: ZDC + autofocus, two channels: DAPI and Cy3 (Figure 3);
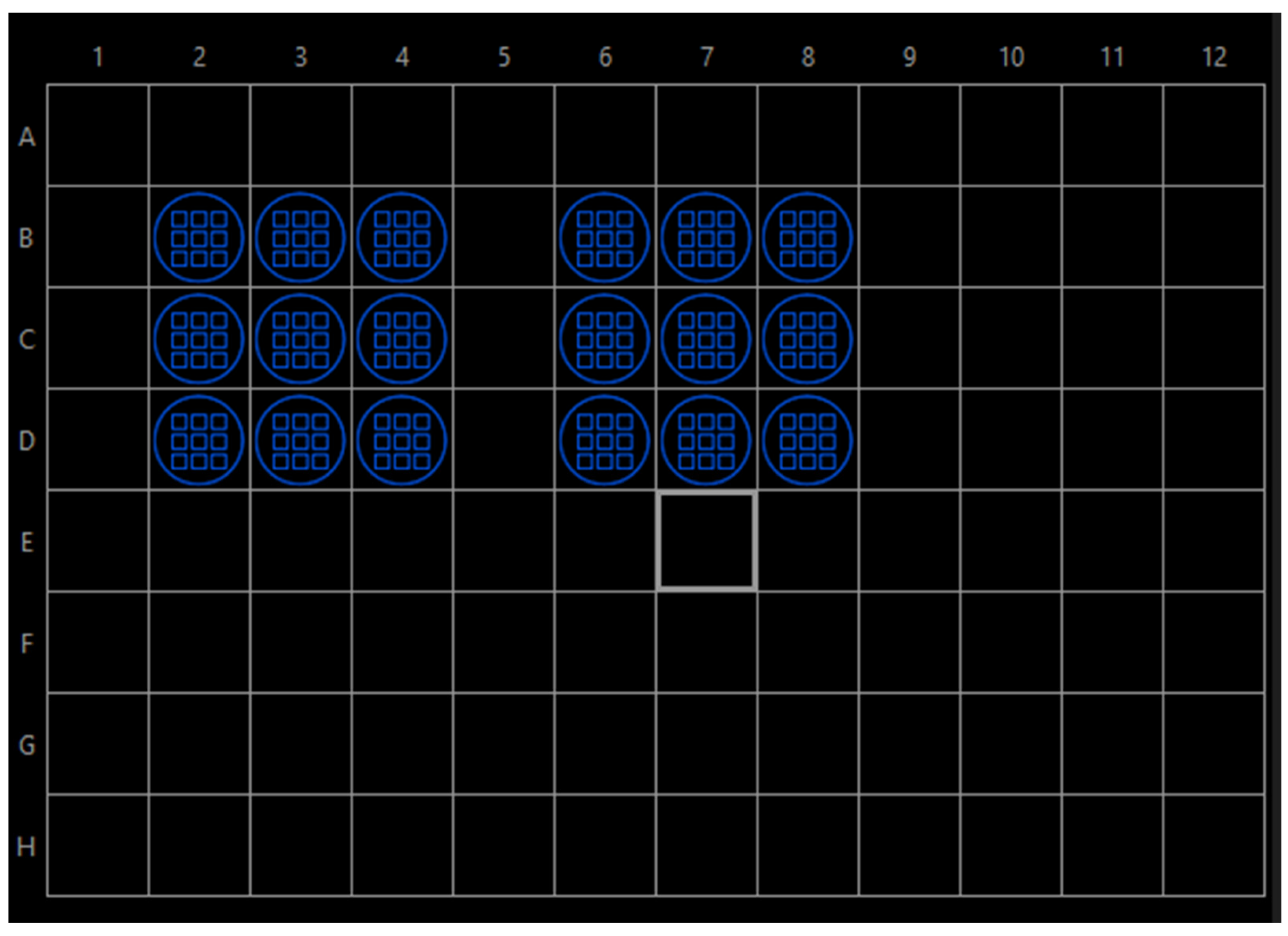
- Well navigator: single frames, 4 × 4 per well (Figure 4).
5.5. BioImage Analysis
- A description of the workflow;
- The code of the workflow;
- The original image data.
5.5.1. Description of the Workflow
5.5.2. The Code of the Workflow and the Original Image Data
- We start our image data flow with image_1, a DAPI channel with nuclei.
- Following this, we apply “Copy” on image_1 and obtain a new image out, image_2.
- As the next step, we apply “Otsu” auto threshold on image_2 and obtain a new image mask out, image_3. The threshold values are saved and used on all DAPI images from the same well.
- Afterward, we apply “Analyze Particles” on image_3, and single out a region of nuclei as a Region of Interest (ROI) set. All ROIs touching edges are skipped.
- In the next step, we open image_4, which is the Cy3 channel. We apply “Copy” on image_4, and obtain a new image, image_5.
- We apply background subtraction with s rolling ball of size 50 on image_5 to subtract the local background value from intensity measurements, and obtain image_6.
- Afterward, image_6 is selected for measuring features under ROIs from the previous step.
- The process Log and measured features from Cy3 channel for the whole well and summary are saved in the “Results” subfolder in the CSV table. A flattened image_4 with ROIs outlined is saved as a JPEG for later inspection.
5.6. Calculations
5.7. Data Analysis and Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix B
- Data
- Underlying data available from Zenodo: https://doi.org/10.5281/zenodo.7673198.
- Naming Convention for images: ChannelName_YYYYMMD_Well_PossitionInWell_AcqRun.FileFormat
- Structure:
- Images;
- 4H—all images for all wells;
- 24H—all images for all wells.
- Results:
- Results_4H;
- Results_24H.
- Available under Creative Commons Attribution 4.0 International.
- Software
- Code available from Zenodo: https://doi.org/10.5281/zenodo.7673497 and GitHub: https://github.com/martinschatz-cz/genotoxicity-bia
- Structure:
- ImageJ_scripts:
- enhance_ROI_outlines_flatten.ijm;
- Process_WFolder_macro_v1.ijm;
- sort_wells.ijm;
- vis_CA_allOpened.ijm.
- Python_scripts:
- SF_dataVis_and_statistics_mean_4h.ipynb;
- SF_dataVis_and_statistics_mean_24h.ipynb;
- README.md;
- Requirements.txt.
- Available under Creative Commons Attribution 4.0 International.
References
- OECD. Overview on Genetic Toxicology TGs; OECD Series on Testing and Assessment; OECD Publishing: Paris, France, 2017. [Google Scholar] [CrossRef]
- Luan, Y.; Honma, M. Genotoxicity testing and recent advances. Genome Instab. Dis. 2021, 3, 1–21. [Google Scholar] [CrossRef]
- Mišík, M.; Nersesyan, A.; Ferk, F.; Holzmann, K.; Krupitza, G.; Herrera Morales, D.; Staudinger, M.; Wultsch, G.; Knasmueller, S. Search for the optimal genotoxicity assay for routine testing of chemicals: Sensitivity and specificity of conventional and new test systems. Mutat. Res./Genet. Toxicol. Environ. Mutagen. 2022, 881, 503524. [Google Scholar] [CrossRef] [PubMed]
- Richmond, J. The 3Rs—Past, Present and Future. Scand. J. Lab. Anim. Sci. 2000, 27, 84–92. [Google Scholar] [CrossRef]
- Mah, L.J.; El-Osta, A.; Karagiannis, T.C. γH2AX: A sensitive molecular marker of DNA damage and repair. Leukemia 2010, 24, 679–686. [Google Scholar] [CrossRef]
- Srivastava, N.; Gochhait, S.; de Boer, P.; Bamezai, R.N.K. Role of H2AX in DNA damage response and human cancers. Mutat. Res./Rev. Mutat. Res. 2009, 681, 180–188. [Google Scholar] [CrossRef]
- Kuo, L.J.; Yang, L.X. γ-H2AX- A novel biomaker for DNA double-strand breaks. In Vivo 2008, 22, 305–310. [Google Scholar]
- Rahmanian, N.; Shokrzadeh, M.; Eskandani, M. Recent advances in γH2AX biomarker-based genotoxicity assays: A marker of DNA damage and repair. DNA Repair 2021, 108, 103243. [Google Scholar] [CrossRef]
- Garcia-Canton, C.; Anadon, A.; Meredith, C. Assessment of the in vitro gammaH2AX assay by High Content Screening as a novel genotoxicity test. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2013, 757, 158–166. [Google Scholar] [CrossRef]
- Reddig, A.; Roggenbuck, D.; Reinhold, D. Comparison of different immunoassays for γH2AX quantification. J. Lab. Precis. Med. 2018, 3, 80. [Google Scholar] [CrossRef]
- Zhang, B.; Li, F.; Shen, L.; Chen, L.; Xia, Z.; Ding, J.; Li, M.; Guo, L.-H. A cathodic photoelectrochemical immunoassay with dual signal amplification for the ultrasensitive detection of DNA damage biomarkers. Biosens. Bioelectron. 2023, 224, 115052. [Google Scholar] [CrossRef]
- Tolosa, L.; Gómez-Lechón, M.J.; Donato, M.T. High-content screening technology for studying drug-induced hepatotoxicity in cell models. Arch. Toxicol. 2015, 89, 1007–1022. [Google Scholar] [CrossRef] [PubMed]
- Popp, H.D.; Brendel, S.; Hofmann, W.-K.; Fabarius, A. Immunofluorescence Microscopy of γH2AX and 53BP1 for Analyzing the Formation and Repair of DNA Double-strand Breaks. JoVE 2017, 129, e56617. [Google Scholar] [CrossRef]
- Willitzki, A.; Lorenz, S.; Hiemann, R.; Guttek, K.; Goihl, A.; Hartig, R.; Conrad, K.; Feist, E.; Sack, U.; Schierack, P.; et al. Fully automated analysis of chemically induced γH2AX foci in human peripheral blood mononuclear cells by indirect immunofluorescence. Cytom. A 2013, 83, 1017–1026. [Google Scholar] [CrossRef]
- Ando, M.; Yoshikawa, K.; Iwase, Y.; Ishiura, S. Usefulness of monitoring γ-H2AX and cell cycle arrest in HepG2 cells for estimating genotoxicity using a high-content analysis system. J. Biomol. Screen. 2014, 19, 1246–1254. [Google Scholar] [CrossRef] [PubMed]
- Quesnot, N.; Rondel, K.; Audebert, M.; Martinais, S.; Glaise, D.; Morel, F.; Loyer, P.; Robin, M.A. Evaluation of genotoxicity using automated detection of γH2AX in metabolically competent HepaRG cells. Mutagenesis 2016, 31, 43–50. [Google Scholar] [CrossRef]
- Marková, E.; Schultz, N.; Belyaev, I.Y. Kinetics and dose-response of residual 53BP1/gamma-H2AX foci: Co-localization, relationship with DSB repair and clonogenic survival. Int. J. Radiat. Biol. 2007, 83, 319–329. [Google Scholar] [CrossRef]
- Kim, Y.J.; Koedrith, P.; Kim, H.S.; Yu, W.J.; Kim, J.C.; Seo, Y.R. Comparative genotoxicity investigation using comet and gammaH2AX assays for screening of genotoxicants in HepG2 human hepatoma cells. Toxicol. Environ. Health Sci. 2016, 8, 68–78. [Google Scholar] [CrossRef]
- Liu, X.L.; Wu, R.Y.; Sun, X.F.; Cheng, S.F.; Zhang, R.Q.; Zhang, T.Y.; Zhang, X.F.; Zhao, Y.; Shen, W.; Li, L. Mycotoxin zearalenone exposure impairs genomic stability of swine follicular granulosa cells in vitro. Int. J. Biol. Sci. 2018, 14, 294–305. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Schneider, J.; Weiss, R.; Ruhe, M.; Jung, T.; Roggenbuck, D.; Stohwasser, R.; Schierack, P.; Rödiger, S. Open source bioimage informatics tools for the analysis of DNA damage and associated biomarkers. J. Lab. Precis. Med. 2019, 4, 21. [Google Scholar] [CrossRef]
- Invitrogen. HCS DNA Damage Kit. Available online: https://www.thermofisher.com/document-connect/document-connect.html?url=https://assets.thermofisher.com/TFS-Assets%2FLSG%2Fmanuals%2Fmp10292.pdf (accessed on 21 February 2023).
- Smart, D.J.; Ahmedi, K.P.; Harvey, J.S.; Lynch, A.M. Genotoxicity screening via the gammaH2AX by flow assay. Mutat. Res. 2011, 715, 25–31. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. The Hallmarks of Cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef]
- Shihan, M.H.; Novo, S.G.; Le Marchand, S.J.; Wang, Y.; Duncan, M.K. A simple method for quantitating confocal fluorescent images. Biochem. Biophys. Rep. 2021, 25, 100916. [Google Scholar] [CrossRef]
- Fowler, P.; Smith, K.; Young, J.; Jeffrey, L.; Kirkland, D.; Pfuhler, S.; Carmichael, P. Reduction of misleading (“false”) positive results in mammalian cell genotoxicity assays. I. Choice of cell type. Mutat. Res./Genet. Toxicol. Environ. Mutagen. 2012, 742, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Donato, M.T.; Tolosa, L.; Gómez-Lechón, M.J. Culture and Functional Characterization of Human Hepatoma HepG2 Cells. In Protocols in In Vitro Hepatocyte Research; Vinken, M., Rogiers, V., Eds.; Springer: New York, NY, USA, 2015; pp. 77–93. [Google Scholar]
- Qu, D.; Gu, Y.; Feng, L.; Han, J. High Content Analysis technology for evaluating the joint toxicity of sunset yellow and sodium sulfite in vitro. Food Chem. 2017, 233, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Qu, D.; Jiang, M.; Huang, D.; Zhang, H.; Feng, L.; Chen, Y.; Zhu, X.; Wang, S.; Han, J. Synergistic Effects of The Enhancements to Mitochondrial ROS, p53 Activation and Apoptosis Generated by Aspartame and Potassium Sorbate in HepG2 Cells. Molecules 2019, 24, 457. [Google Scholar] [CrossRef] [PubMed]
- Rogakou, E.P.; Nieves-Neira, W.; Boon, C.; Pommier, Y.; Bonner, W.M. Initiation of DNA fragmentation during apoptosis induces phosphorylation of H2AX histone at serine 139. J. Biol. Chem. 2000, 275, 9390–9395. [Google Scholar] [CrossRef]
- Duke, R.C.; Witter, R.Z.; Nash, P.B.; Young, J.D.-E.; Ojcius, D.M. Cytolysis mediated by ionophores and pore-forming agents: Role of intracellular calcium in apoptosis. FASEB J. 1994, 8, 237–246. [Google Scholar] [CrossRef]
- Su, Z.; Ran, X.; Leitch, J.J.; Schwan, A.L.; Faragher, R.; Lipkowski, J. How Valinomycin Ionophores Enter and Transport K+ across Model Lipid Bilayer Membranes. Langmuir 2019, 35, 16935–16943. [Google Scholar] [CrossRef]
- Korga, A.; Ostrowska, M.; Jozefczyk, A.; Iwan, M.; Wojcik, R.; Zgorka, G.; Herbet, M.; Vilarrubla, G.G.; Dudka, J. Apigenin and hesperidin augment the toxic effect of doxorubicin against HepG2 cells. BMC Pharmacol. Toxicol. 2019, 20, 22. [Google Scholar] [CrossRef]
- Miura, K.; Nørrelykke, S.F. Reproducible image handling and analysis. EMBO J. 2021, 40, e105889. [Google Scholar] [CrossRef] [PubMed]
- Levet, F.; Carpenter, A.; Eliceiri, K.; Kreshuk, A.; Bankhead, P.; Haase, R. Developing open-source software for bioimage analysis: Opportunities and challenges [version 1; peer review: 2 approved]. F1000Research 2021, 10, 302. [Google Scholar] [CrossRef] [PubMed]
- Haase, R.; Jain, A.; Rigaud, S.; Vorkel, D.; Rajasekhar, P.; Suckert, T.; Lambert, T.J.; Nunez-Iglesias, J.; Poole, D.P.; Tomancak, P.; et al. Interactive design of GPU-accelerated Image Data Flow Graphs and cross-platform deployment using multi-lingual code generation. bioRxiv 2020. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Overall Intensity (OI) or Foci Detection (FD) | Device for Detection | Software for Bioimage Analysis | Ref. |
|---|---|---|---|
| FD | AKLIDES Cell Damage system, Medipan | [14] | |
| FD | AKLIDES Cell Damage system, Medipan | [10] | |
| OI | Cellomics Arrayscan VTI platform 1, Thermo Scientific, TargetActivation Bioapplication software V.6.6.1.4. | [9] | |
| OI | ArrayScan VTI HSC reader Reader 1, Thermo Scientific | [15] | |
| OI | Cellomic Arrayscan VTI HCS Reader 1, Thermo Scientific | [16] | |
| FD | confocal laser scanning microscope (Zeiss LSM 510) | Foci 8.0 software (Schultz and Belyaev, unpublished) | [17] |
| FD | confocal microscope (Nikon) | NE Element software (Nikon) | [18] |
| OI | Olympus fluorescence microscope (BX51) | ImageJ software (v.1.47) | [19] |
| γH2AX Signal Increase | RCC 1 | Classification |
|---|---|---|
| >1.5× | Above 25% | Genotoxic |
| <1.5× | 0–100% | Non-genotoxic |
| >1.5× | Below 25% | Cytotoxicity-driven genotoxicity = false positive |
| 1.5× | ≥25% | Equivocal |
| Cell Line | Sample | MFV (γH2AX Response) | Area (Nuclei) [µm2] | MoA | Fold | RA | ||
|---|---|---|---|---|---|---|---|---|
| Mean | IQR | Mean | IQR | |||||
| CHO-K1 | Ctrl | 1.641 | 1.520 | 2.130 | 1.058 | 0.770 | 1 | 100% |
| Val30 | 2.179 | 1.872 | 1.423 | 0.862 | 1.531 | 1.987 | 66.81% | |
| Val15 | 2.151 | 1.439 | 1.614 | 0.814 | 1.333 | 1.730 | 75.78% | |
| HeLa | Ctrl | 0.132 | 0.166 | 3.249 | 1.334 | 0.041 | 1 | 100% |
| Val30 | 0.127 | 0.149 | 2.955 | 1.109 | 0.043 | 1.063 | 90.94% | |
| Val15 | 0.119 | 0.147 | 3.040 | 1.167 | 0.039 | 0.966 | 93.56% | |
| Cell Line | Sample | MFV (γH2AX Response) | Area (Nuclei) [µm2] | MoA | Fold | RA | ||
|---|---|---|---|---|---|---|---|---|
| Mean | IQR | Mean | IQR | |||||
| CHO-K1 | Ctrl | 1.518 | 1.509 | 2.186 | 0.973 | 0.694 | 1 | 100% |
| Val30 | 2.785 | 3.229 | 0.893 | 0.405 | 3.119 | 4.495 | 40.83% | |
| Val15 | 2.605 | 2.670 | 1.173 | 0.572 | 2.221 | 3.199 | 53.66% | |
| HeLa | Ctrl | 0.064 | 0.082 | 2.919 | 1.276 | 0.022 | 1 | 100% |
| Val30 | 0.595 | 0.324 | 2.070 | 0.806 | 0.287 | 13.200 | 70.91% | |
| Val15 | 0.358 | 0.275 | 2.227 | 0.893 | 0.161 | 7.376 | 76.30% | |
| Sample | γH2AX Response | Cell Number | Mean over Cell Number | Fold | Relative Cell Number |
|---|---|---|---|---|---|
| Ctrl | 90.4 | 184 | 0.491 | 1.0 | 100% |
| Val30 | 201.5 | 72.9 | 2.764 | 5.6 | 40% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Křížkovská, B.; Schätz, M.; Lipov, J.; Viktorová, J.; Jablonská, E. In Vitro High-Throughput Genotoxicity Testing Using γH2AX Biomarker, Microscopy and Reproducible Automatic Image Analysis in ImageJ—A Pilot Study with Valinomycin. Toxins 2023, 15, 263. https://doi.org/10.3390/toxins15040263
Křížkovská B, Schätz M, Lipov J, Viktorová J, Jablonská E. In Vitro High-Throughput Genotoxicity Testing Using γH2AX Biomarker, Microscopy and Reproducible Automatic Image Analysis in ImageJ—A Pilot Study with Valinomycin. Toxins. 2023; 15(4):263. https://doi.org/10.3390/toxins15040263
Chicago/Turabian StyleKřížkovská, Bára, Martin Schätz, Jan Lipov, Jitka Viktorová, and Eva Jablonská. 2023. "In Vitro High-Throughput Genotoxicity Testing Using γH2AX Biomarker, Microscopy and Reproducible Automatic Image Analysis in ImageJ—A Pilot Study with Valinomycin" Toxins 15, no. 4: 263. https://doi.org/10.3390/toxins15040263
APA StyleKřížkovská, B., Schätz, M., Lipov, J., Viktorová, J., & Jablonská, E. (2023). In Vitro High-Throughput Genotoxicity Testing Using γH2AX Biomarker, Microscopy and Reproducible Automatic Image Analysis in ImageJ—A Pilot Study with Valinomycin. Toxins, 15(4), 263. https://doi.org/10.3390/toxins15040263