Proteomics of Vespa velutina nigrithorax Venom Sac Queens and Workers: A Quantitative SWATH-MS Analysis

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Morphology of the Venom Sac of Vespa velutina Queens and Workers
2.2. SDS-PAGE Analysis of the Venom Sac Potential Allergenic Proteins of Vespa velutina Queens and Workers
2.3. Proteomic Quantitative Analysis, Biological Pathways and Molecular Functions of Dysregulated Proteins in the Venom Sac of Vespa velutina Queens and Workers
2.4. Proteomic Quantitative Analysis from Potential Vespa velutina Allergens
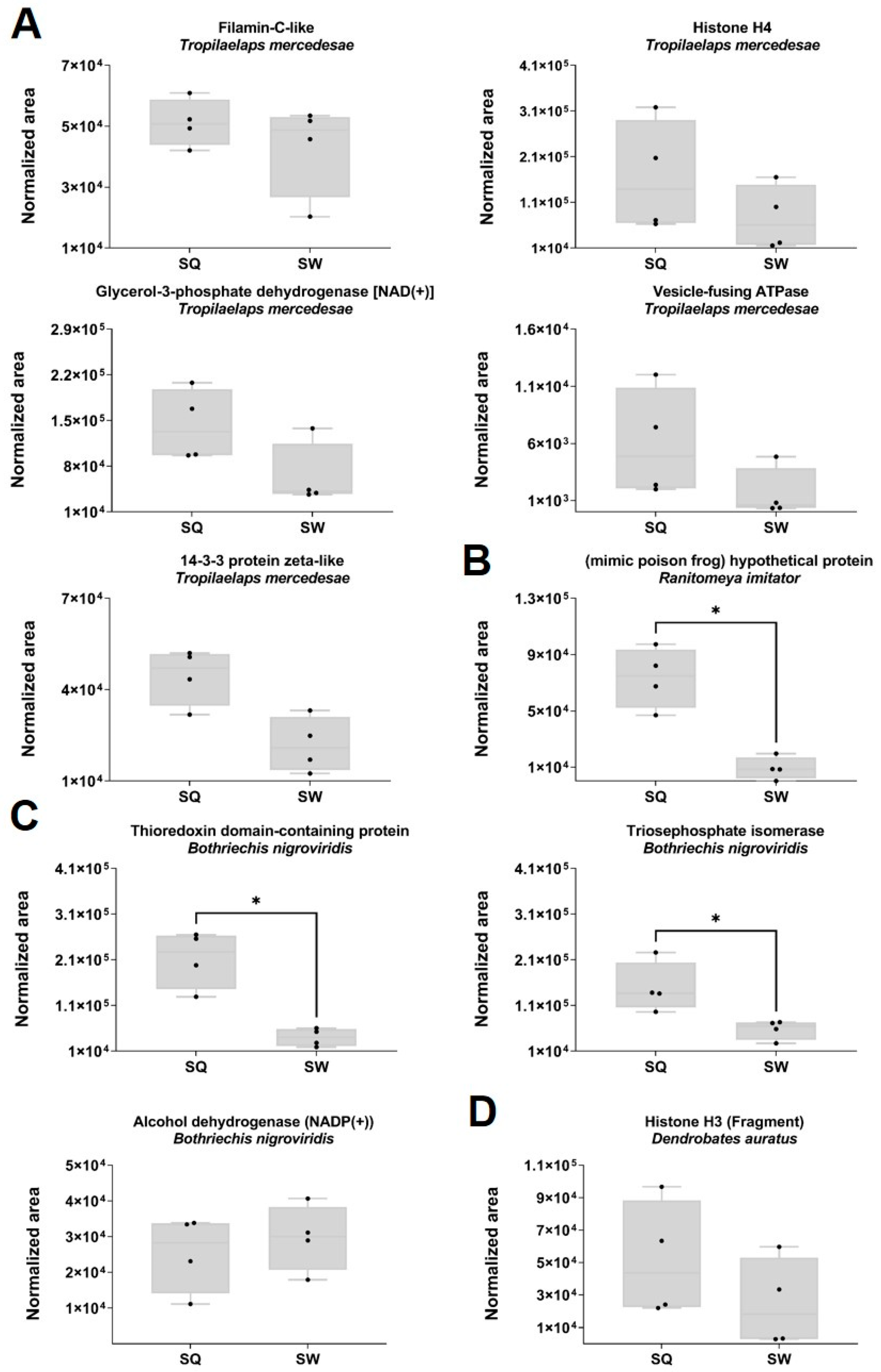
2.5. Proteomic Quantitative Analysis from Dysregulated Proteins of the Class Insecta
2.6. Proteomic Quantitative Analysis from Dysregulated Proteins of the Classes Amphibia, Arachnida and Reptilia
2.7. Proteomic Quantitative Analysis of Dysregulated Proteins from the Classes Bacilla, γ-Proteobacteria and Pisoniviricetes
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Source of Insects
5.2. Venom Collection
5.3. Sample Preparation for Mass Spectrometric Analysis
5.4. Sequential Window Acquisition of All Theoretical Mass Spectra (SWATH-MS) Quantification
5.5. Protein Functional Analysis
5.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brock, R.E.; Cini, A.; Sumner, S. Ecosystem services provided by aculeate wasps. Biol. Rev. Camb. Philos. Soc. 2021, 96, 1645–1675. [Google Scholar] [CrossRef] [PubMed]
- Perrard, A.; Pickett, K.; Villemant, C.; Kojima, J.; Carpenter, J. Phylogeny of hornets: A total evidence approach (Hymenoptera, Vespidae, Vespinae, Vespa). J. Hymenopt. Res. 2013, 2, 1–15. [Google Scholar] [CrossRef]
- Detoni, M.; Féas, X.; Jeanne, R.L.; Loope, K.J.; O’Donnell, S.; Santoro, D.; Sumner, S.; Jandt, J.M. Evolutionary and ecological pressures shaping social wasps collective defenses. Ann. Entomol. Soc. Am. 2020, 113, 407–424. [Google Scholar] [CrossRef]
- Herrera, C.; Leza, M.; Martínez-López, E. Diversity of compounds in Vespa spp. venom and the epidemiology of its sting: A global appraisal. Arch. Toxicol. 2020, 94, 3609–3627. [Google Scholar] [CrossRef] [PubMed]
- Dongol, T.; Dhananjaya, B.L.; Shrestha, R.K.; Aryal, G. Pharmacological and immunological properties of wasp venom. In Pharmacology and Therapeutics; Joghi, S., Gowder, T., Eds.; Intech: Rijeka, Croatia, 2014; pp. 47–81. [Google Scholar] [CrossRef] [Green Version]
- Abd El-Wahed, A.; Yosri, N.; Sakr, H.H.; Du, M.; Algethami, A.F.M.; Zhao, C.; Abdelazeem, A.H.; Tahir, H.E.; Masry, S.H.D.; Abdel-Daim, M.M.; et al. Wasp Venom Biochemical Components and Their Potential in Biological Applications and Nanotechnological Interventions. Toxins 2021, 13, 206. [Google Scholar] [CrossRef]
- Silva, J.; Monge-Fuentes, V.; Gomes, F.; Lopes, K.; dos Anjos, L.; Campos, G.; Arenas, C.; Biolchi, A.; Gonçalves, J.; Galante, P.; et al. Pharmacological Alternatives for the Treatment of Neurodegenerative Disorders: Wasp and Bee Venoms and Their Components as New Neuroactive Tools. Toxins 2015, 7, 3179–3209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chugo, S.; Lizaso, M.T.; Alvarez, M.J.; Arroabaren, E.; Lizarza, S.; Tabar, A.I. Vespa velutina nigritorax: A New Causative Agent in Anaphylaxis. J. Investig. Allergol. Clin. Immunol. 2015, 25, 231–232. [Google Scholar] [PubMed]
- Vega, J.M.; Ortiz-Sánchez, F.J.; Martínez-Arcediano, A.; Castro, L.; Alfaya, T.; Carballada, F.; Cruz, S.; Marqués, L.; Vega, A.; Ruiz-León, B. Social wasps in Spain: The who and where. Allergol. Immunopathol. 2022, 50, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Carriazo, S.; Ortiz, A. Wasp stings and plasma exchange. Clin. Kidney J. 2022, 15, 1455–1458. [Google Scholar] [CrossRef]
- Laborde-Castérot, H.; Darrouzet, E.; Le Roux, G.; Labadie, M.; Delcourt, N.; de Haro, L.; Vodovar, D.; Langrand, J.; French Poison Control Centers Research Group. Ocular Lesions Other Than Stings Following Yellow-Legged Hornet (Vespa velutina nigrithorax) Projections, as Reported to French Poison Control Centers. JAMA Ophthalmol. 2021, 139, 105–108. [Google Scholar] [CrossRef]
- de Haro, L.; Labadie, M.; Chanseau, P.; Cabot, C.; Blanc-Brisset, I.; Penouil, F.; National Coordination Committee for Toxicovigilance. Medical consequences of the Asian black hornet (Vespa velutina) invasion in Southwestern France. Toxicon 2010, 55, 650–652. [Google Scholar] [CrossRef] [PubMed]
- Van der Brempt, X. Les anaphylaxies aux piqûres de frelons asiatiques (Vespa velutina): Cas déclarés au Réseau d’Allergo-Vigilance® (RAV) de 2011 à 2021 et conduite à tenir. Rev. Fr. Allergol. 2022, 62, 166–170. [Google Scholar] [CrossRef]
- Miranda, J.; Mesquita, A.M.; Plácido, J.; Coimbra, A. Vespa velutina nigrithorax venom allergy: Real-life clinical practice. Ann. Allergy Asthma Immunol. 2022, 129, 524–525. [Google Scholar] [CrossRef] [PubMed]
- Esteves Caldeira, L.; Silva, M.I.T.; Pedro, E.; Cosme, J. Hypersensitivity to Vespa velutina nigrithorax: An emerging problem in Portugal? Eur. Ann. Allergy Clin. Immunol. 2023. [Google Scholar] [CrossRef]
- Grossi, V.; Severino, M.; Massolo, A.; Infantino, M.; Laureti, F.; Macchia, D.; Meucci, E.; Francescato, E.; Pantera, B.; Ebbli, A.; et al. Vespa velutina nigrithorax venom allergy: Inhibition studies approach for the choice of specific immunotherapy. Eur. Ann. Allergy Clin. Immunol. 2022. [Google Scholar] [CrossRef]
- Xie, C.; Xu, S.; Ding, F.; Xie, M.; Lv, J.; Yao, J.; Pan, D.; Sun, Q.; Liu, C.; Chen, T.; et al. Clinical features of severe wasp sting patients with dominantly toxic reaction: Analysis of 1091 cases. PLoS ONE 2013, 8, e83164. [Google Scholar] [CrossRef] [Green Version]
- Choi, M.B.; Hong, E.J.; Kwon, O. Defensive behavior of the invasive alien hornet, Vespa velutina, against color, hair and auditory stimuli of potential aggressors. PeerJ 2021, 9, e11249. [Google Scholar] [CrossRef]
- Choi, M.B.; Kim, T.G.; Kwon, O. Recent Trends in Wasp Nest Removal and Hymenoptera Stings in South Korea. J. Med. Entomol. 2019, 56, 254–260. [Google Scholar] [CrossRef]
- Vidal, C. The Asian wasp Vespa velutina nigrithorax: Entomological and allergological characteristics. Clin. Exp. Allergy 2022, 52, 489–498. [Google Scholar] [CrossRef]
- Feás Sánchez, X.; Charles, R.J. Notes on the nest architecture and colony composition in winter of the yellow-legged Asian Hornet, Vespa velutina Lepeletier 1836 (Hym.: Vespidae), in its introduced habitat in Galicia (NW Spain). Insects 2019, 10, 237. [Google Scholar] [CrossRef] [Green Version]
- Feás, X.; Vázquez-Tato, M.P.; Seijas, J.A.; Pratima GNikalje, A.; Fraga-López, F. Extraction and Physicochemical Characterization of Chitin Derived from the Asian Hornet, Vespa velutina Lepeletier 1836 (Hym.: Vespidae). Molecules 2020, 25, 384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feás, X.; Vidal, C.; Remesar, S. What We Know about Sting-Related Deaths? Human Fatalities Caused by Hornet, Wasp and Bee Stings in Europe (1994–2016). Biology 2022, 11, 282. [Google Scholar] [CrossRef] [PubMed]
- Feás, X.; Vidal, C.; Vázquez-Tato, M.P.; Seijas, J.A. Asian Hornet, Vespa velutina Lepeletier 1836 (Hym.: Vespidae), Venom Obtention Based on an Electric Stimulation Protocol. Molecules 2021, 27, 138. [Google Scholar] [CrossRef]
- Feás, X. Human Fatalities Caused by Hornet, Wasp and Bee Stings in Spain: Epidemiology at State and Sub-State Level from 1999 to 2018. Biology 2021, 10, 73. [Google Scholar] [CrossRef] [PubMed]
- Yun, H.S.; Oh, J.; Lim, J.S.; Kim, H.J.; Kim, J.S. Anti-Inflammatory Effect of Wasp Venom in BV-2 Microglial Cells in Comparison with Bee Venom. Insects 2021, 12, 297. [Google Scholar] [CrossRef]
- Meng, Y.C.; Mo, X.G.; He, T.T.; Wen, X.X.; Nieh, J.C.; Yang, X.W.; Tan, K. New bioactive peptides from the venom gland of a social hornet Vespa velutina. Toxicon 2021, 199, 94–100. [Google Scholar] [CrossRef]
- Chae, J.H.; Oh, J.; Lim, J.S.; Jeong, Y.A.; Yun, H.S.; Jang, C.H.; Kim, H.J.; Kim, J.S. Wasp Venom Ameliorates Scopolamine-Induced Learning and Memory Impairment in Mice. Toxins 2022, 14, 256. [Google Scholar] [CrossRef]
- Le, T.N.; Da Silva, D.; Colas, C.; Darrouzet, E.; Baril, P.; Leseurre, L.; Maunit, B. Asian hornet Vespa velutina nigrithorax venom: Evaluation and identification of the bioactive compound responsible for human keratinocyte protection against oxidative stress. Toxicon 2020, 176, 1–9. [Google Scholar] [CrossRef]
- Cappa, F.; Cini, A.; Pepiciello, I.; Petrocelli, I.; Inghilesi, A.F.; Anfora, G.; Dani, F.R.; Bortolotti, L.; Wen, P.; Cervo, R. Female volatiles as sex attractants in the invasive population of Vespa velutina nigrithorax. J. Insect. Physiol. 2019, 119, 103952. [Google Scholar] [CrossRef]
- Cheng, Y.N.; Wen, P.; Dong, S.H.; Tan, K.; Nieh, J.C. Poison and alarm: The Asian hornet. J. Exp. Biol. 2017, 220 Pt 4, 645–651. [Google Scholar] [CrossRef] [Green Version]
- Thiéry, D.; Bonnard, O.; Riquier, L.; De Revel, G.; Monceau, K. An alarm pheromone in the venom gland of Vespa velutina: Evidence revisited from the European invasive population. Entomol. Gen. 2018, 38, 145–156. [Google Scholar] [CrossRef]
- Vidal, C.; Armisén, M.; Monsalve, R.; Gómez-Rial, J.; González-Fernández, T.; Carballada, F.; Lombardero, M.; González-Quintela, A. Vesp v 5 and glycosylated Vesp v 1 are relevant allergens in Vespa velutina nigrithorax anaphylaxis. Clin. Exp. Allergy 2020, 50, 1424–1427. [Google Scholar] [CrossRef] [PubMed]
- Vidal, C.; Armisén, M.; Monsalve, R.; González-Vidal, T.; Lojo, S.; López-Freire, S.; Méndez, P.; Rodríguez, V.; Romero, L.; Galán, A.; et al. Anaphylaxis to Vespa velutina nigrithorax: Pattern of Sensitization for an Emerging Problem in Western Countries. J. Investig. Allergol. Clin. Immunol. 2021, 31, 228–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Vázquez, V.; Gómez-Rial, J.; Monsalve, R.I.; Vidal, C. Consistency of Determination of sIgE and the Basophil Activation Test in Vespa velutina nigrithorax Allergy. J. Investig. Allergol. Clin. Immunol. 2022, 32, 143–145. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Vázquez, V.; Armisén, M.; Gómez-Rial, J.; Lamas-Vázquez, B.; Vidal, C. Immunotherapy with Vespula venom for Vespa velutina nigrithorax anaphylaxis: Preliminary clinical and immunological results. Clin. Exp. Allergy 2022, 52, 345–347. [Google Scholar] [CrossRef]
- Ruiz-Leon, B.; Serrano, P.; Vidal, C.; Moreno-Aguilar, C. Management of Double Sensitization to Vespids in Europe. Toxins 2022, 14, 126. [Google Scholar] [CrossRef] [PubMed]
- Guido-Patiño, J.C.; Plisson, F. Profiling hymenopteran venom toxins: Protein families, structural landscape, biological activities, and pharmacological benefits. Toxicon X 2022, 14, 100119. [Google Scholar] [CrossRef]
- Dos Santos-Pinto, J.R.A.; Perez-Riverol, A.; Lasa, A.M.; Palma, M.S. Diversity of peptidic and proteinaceous toxins from social Hymenoptera venoms. Toxicon 2018, 148, 172–196. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Chen, S.; Zhou, Y.; Xie, C.; Zhu, B.; Zhu, H.; Liu, S.; Wang, W.; Chen, H.; Ji, Y. Deciphering the venomic transcriptome of killer-wasp Vespa velutina. Sci. Rep. 2015, 5, 9454. [Google Scholar] [CrossRef]
- Tan, J.; Wang, W.; Wu, F.; Li, Y.; Fan, Q. Transcriptome profiling of venom gland from wasp species: De novo assembly, functional annotation, and discovery of molecular markers. BMC Genom. 2020, 21, 427. [Google Scholar] [CrossRef]
- Le, T.N.; da Silva, D.; Colas, C.; Darrouzet, E.; Baril, P.; Leseurre, L.; Maunit, B. Development of an LC-MS multivariate nontargeted methodology for differential analysis of the peptide profile of Asian hornet venom (Vespa velutina nigrithorax): Application to the investigation of the impact of collection period variation. Anal. Bioanal. Chem. 2020, 412, 1419–1430. [Google Scholar] [CrossRef]
- Nelsen, D.R.; Nisani, Z.; Cooper, A.M.; Fox, G.A.; Gren, E.C.; Corbit, A.G.; Hayes, W.K. Poisons, toxungens, and venoms: Redefining and classifying toxic biological secretions and the organisms that employ them. Biol. Rev. Camb. Philos. Soc. 2014, 89, 450–465. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, F.; Wang, X.L.; Wang, P.H.; Ma, S.L.; Yang, Y.; Ye, W.-G.; Diao, K.-Y.; Dai, P.L. Midgut Bacterial Communities of Vespa velutina Lepeletier (Hymenoptera: Vespidae). Front. Ecol. Evol. 2022, 10, 934054. [Google Scholar] [CrossRef]
- Suenami, S.; Konishi, N.M.; Miyazaki, R. Community analysis of gut microbiota in hornets, the largest eusocial wasps, Vespa mandarinia and V. simillima. Sci. Rep. 2019, 9, 9830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cini, A.; Meriggi, N.; Bacci, G.; Cappa, F.; Vitali, F.; Cavalieri, D.; Cervo, R. Gut microbial composition in different castes and developmental stages of the invasive hornet Vespa velutina nigrithorax. Sci. Total Environ. 2020, 745, 140873. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Seo, J.; Yang, S.H.; Kim, I.-S.; Koo, Y. Intestine bacterial microbiota of Asian hornet (Vespa velutina nigrithorax) and honey bee. Korean J. Environ. Agric. 2018, 37, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Esmaeilishirazifard, E.; Usher, L.; Trim, C.; Denise, H.; Sangal, V.; Tyson, G.H.; Barlow, A.; Redway, K.F.; Taylor, J.D.; Kremyda-Vlachou, M.; et al. Bacterial Adaptation to Venom in Snakes and Arachnida. Microbiol. Spectr. 2022, 10, e0240821. [Google Scholar] [CrossRef]
- Gkitsaki, I.; Papachristoforou, A.; Michailidou, S.; Karamvalis, N.; Iliadis, I.; Graikini, D.; Sakarikou, C.; Tsoukis, E.; Argyriou, A.; Giaouris, E. The transmittable through stinging microbiota differs between honeybees and wasps: A potentially greater microbial risk of the wasp sting for humans. Int. Microbiol. 2023. [Google Scholar] [CrossRef]
- Ul-Hasan, S.; Rodríguez-Román, E.; Reitzel, A.M.; Adams, R.M.M.; Herzig, V.; Nobile, C.J.; Saviola, A.J.; Trim, S.A.; Stiers, E.E.; Moschos, S.A.; et al. The emerging field of venom-microbiomics for exploring venom as a microenvironment, and the corresponding Initiative for Venom Associated Microbes and Parasites (iVAMP). Toxicon X 2019, 4, 100016. [Google Scholar] [CrossRef]
- Djukic, M.; Brzuszkiewicz, E.; Fünfhaus, A.; Voss, J.; Gollnow, K.; Poppinga, L.; Liesegang, H.; Garcia-Gonzalez, E.; Genersch, E.; Daniel, R. How to kill the honey bee larva: Genomic potential and virulence mechanisms of Paenibacillus larvae. PLoS ONE 2014, 9, e90914. [Google Scholar] [CrossRef]
- Beims, H.; Bunk, B.; Erler, S.; Mohr, K.I.; Spröer, C.; Pradella, S.; Günther, G.; Rohde, M.; von der Ohe, W.V.; Steinert, M. Discovery of Paenibacillus larvae ERIC V: Phenotypic and genomic comparison to genotypes ERIC I-IV reveal different inventories of virulence factors which correlate with epidemiological prevalences of American Foulbrood. Int. J. Med. Mictrobiol. 2020, 310, 151394. [Google Scholar] [CrossRef]
- Nowar, E.E. Oriental Hornet (Vespa orientalis) as AFB Disease Vector to Honeybee (Apis mellifera L.). Colonies. Middle East J. Appl. Sci. 2021, 06, 934–940. [Google Scholar]
- Martin, S.J.; Brettell, L.E. Deformed Wing Virus in Honeybees and Other Insects. Annu. Rev. Virol. 2019, 6, 49–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chauzat, M.-P.; Schurr, F.; Faucon, J.-P.; Ribière-Chabert, M.; Blanchard, P.; Drajnudel, P. First detections of honey bee pathogens in nest of the Asian hornet (Vespa velutina) collected in France. CIHEAM Watch. Lett. 2015, 33, 1–5. [Google Scholar]
- Dalmon, A.; Gayral, P.; Decante, D.; Klopp, C.; Bigot, D.; Thomasson, M.; Herniou, E.A.; Alaux, C.; Le Conte, Y. Viruses in the Invasive Hornet. Viruses 2019, 11, 1041. [Google Scholar] [CrossRef] [Green Version]
- Mazzei, M.; Forzan, M.; Cilia, G.; Sagona, S.; Bortolotti, L.; Felicioli, A. First detection of replicative deformed wing virus (DWV) in Vespa velutina nigrithorax. Bull. Insectol. 2018, 71, 211–216. [Google Scholar]
- Brown, J.L.; Twomey, E.; Amézquita, A.; Souza, M.B.; Caldwell, J.P.; Lötters, S.; May, R.V.; Melo-Sampaio, P.R.; Mejia-Vargas, D.; Pérez-Peña, P.; et al. A taxonomic revision of the Neotropical poison frog genus Ranitomeya (Amphibia: Dendrobatidae). Zootaxa 2011, 3083, 1–120. [Google Scholar] [CrossRef] [Green Version]
- Koludarov, I.; Velasque, M.; Timm, T.; Greve, C.; Ben Hamadou, A.; Kumar Gupta, D.; Lochnit, G.; el Heinzinger, M.; Vilcinskas, A.; Gloag, R.; et al. A common venomous ancestor? Prevalent bee venom genes evolved before the aculeate stinger while few major toxins are bee-specific. bioRxiv 2022. [Google Scholar] [CrossRef]
- Ruiz-León, B.; Martínez San Ireneo, M.; de la Roca, F.; Arenas, L.; Alfaya Arias, T.; Cordobés, C.; Marqués, L.; Vega, A.; Moreno-Aguilar, C. The Lights and the Shadows of Controlled Sting Challenge with Hymenoptera. J. Investig. Allergol. Clin. Immunol. 2022, 32, 357–366. [Google Scholar] [CrossRef]
- Feás, X. The Yellow-Legged Asian Hornet (Vespa velutina): Reproductive Caste—Drone. Available online: https://youtu.be/JgMyW1v5H1Y (accessed on 20 March 2023).
- Feás, X. The Yellow-Legged Asian Hornet (Vespa velutina) collecting water. Available online: https://youtu.be/d9lJFNXtES0 (accessed on 20 March 2023).
- Bonzon-Kulichenko, E.; Pérez-Hernández, D.; Núñez, E.; Martínez-Acedo, P.; Navarro, P.; Trevisan-Herraz, M.; Ramos Mdel, C.; Sierra, S.; Martínez-Martínez, S.; Ruiz-Meana, M.; et al. A robust method for quantitative high-throughput analysis of proteomes by 18O labeling. Mol. Cell. Proteom. 2011, 10, M110.003335. [Google Scholar] [CrossRef] [Green Version]
- Perez-Hernandez, D.; Gutiérrez-Vázquez, C.; Jorge, I.; López-Martín, S.; Ursa, A.; Sánchez-Madrid, F.; Vázquez, J.; Yáñez-Mó, M. The intracellular interactome of tetraspanin-enriched microdomains reveals their function as sorting machineries toward exosomes. J. Biol. Chem. 2013, 288, 11649–11661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shevchenko, A.; Wilm, M.; Vorm, O.; Jensen, O.N.; Podtelejnikov, A.V.; Neubauer, G.; Shevchenko, A.; Mortensen, P.; Mann, M. A strategy for identifying gel-separated proteins in sequence databases by MS alone. Biochem. Soc. Trans. 1996, 24, 893–896. [Google Scholar] [CrossRef] [Green Version]
- Peñas-Martínez, J.; Barrachina, M.N.; Cuenca-Zamora, E.J.; Luengo-Gil, G.; Bravo, S.B.; Caparrós-Pérez, E.; Teruel-Montoya, R.; Eliseo-Blanco, J.; Vicente, V.; García, Á.; et al. Qualitative and quantitative comparison of plasma exosomes from neonates and adults. Int. J. Mol. Sci. 2021, 22, 1926. [Google Scholar] [CrossRef]
- Camino, T.; Lago-Baameiro, N.; Sueiro, A.; Bravo, S.B.; Couto, I.; Santos, F.F.; Teruel-Montoya, R.; Eliseo-Blanco, J.; Vicente, V.; García, Á.; et al. Brown Adipose Tissue Sheds Extracellular Vesicles That Carry Potential Biomarkers of Metabolic and Thermogenesis Activity Which Are Affected by High Fat Diet Intervention. Int. J. Mol. Sci. 2022, 23, 10826. [Google Scholar] [CrossRef] [PubMed]
- Chantada-Vázquez, M.D.P.; Conde-Amboage, M.; Graña-López, L.; Vázquez-Estévez, S.; Bravo, S.B.; Núñez, C. Circulating Proteins Associated with Response and Resistance to Neoadjuvant Chemotherapy in HER2-Positive Breast Cancer. Cancers 2022, 14, 1087. [Google Scholar] [CrossRef]
- Shilov, I.V.; Seymour, S.L.; Patel, A.A.; Loboda, A.; Tang, W.H.; Keating, S.P.; Hunter, C.L.; Nuwaysir, L.M.; Schaeffer, D.A. The Paragon Algorithm, a Next Generation Search Engine That Uses Sequence Temperature Values and Feature Probabilities to Identify Peptides from Tandem Mass Spectra. Mol. Cell. Proteom. 2007, 6, 1638–1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Order/Family | Scientific Name/Common Name | pp | Group |
|---|---|---|---|---|
| Amphibia | Anura/Dendrobatidae | Ranitomeya imitator (Schulte, 1986)/Mimic poison frog. | 16 | 9NEOB |
| Dendrobates auratus (Girard, 1955)/Green and black poison dart frog | 1 | DENAT | ||
| Arachnida | Mesostigmata/Laelapidae | Tropilaelaps mercedesae (Delfinado & Baker, 1961)/Tropilaelaps mite | 5 | 9ACAR |
| Bacilli | Bacillales/Paenibacillaceae | Paenibacillus larvae subsp. larvae (White 1906)/American Foulbrood | 1 | 9BACL |
| Lactobacillales/Lactobacillaceae | Lactobacillus sp. | 1 | 9LACO | |
| Lactobacillus bombicola (Praet et al., 2015) | 1 | 9LACO | ||
| γ-proteobacteria | Enterobacterales/Enterobacteriaceae | Klebsiella sp. | 3 | 9ENTR |
| Enterobacterales/Erwiniaceae | Pantoea agglomerans (Ewing & Fife, 1972) Gavini et al., 1989 | 2 | ENTAG | |
| Orbales/Orbaceae | Candidatus Schmidhempelia bombi str. Bimp | 1 | 9GAMM | |
| Gilliamella apis (Ludvigsen et al., 2018) | 1 | 9GAMM | ||
| Pseudomonadales/Pseudomonadacea | Pseudomonas sp. | 1 | 9PSED | |
| Insecta | Hymenoptera/Apidae | Apis cerana cerana (Fabricius, 1793)/Asian honeybee | 49 | APICC |
| Apis mellifera ligustica (Spinola, 1806)/Italian honeybee | 2 | APILI | ||
| Apis mellifera (Linnaeus, 1758)/European honeybee | 2 | APIME | ||
| Bombus festivus (Smith, 1861)/Bumblebee of Sichuan | 1 | 9HYME | ||
| Bombus humilis (Illiger, 1806)/Brown-banded carder bumblebee | 1 | BOMHU | ||
| Frieseomelitta varia (Lepeletier, 1836)/Yellow marmalade bee | 82 | 9HYME | ||
| Xylocopa caerulea (Fabricius, 1804)/Blue carpenter bee | 1 | 9HYME | ||
| Hymenoptera/ Halictidae | Dufourea novaeangliae (Robertson, 1897)/Sweat bee | 47 | DUFNO | |
| Hymenoptera/ Megachilidae | Megachile rotundata (Fabricius, 1787)/Alfalfa leafcutting bee | 1 | MEGRT | |
| Hymenoptera/Vespidae | Vespa velutina (Lepeletier, 1836)/Yellow-legged Asian hornet | 5 | VESVE | |
| Pisoniviricetes | Picornavirales/Iflaviridae | Deformed wing virus | 1 | 9VIRU |
| Reptilia | Squamata/Viperidae | Bothriechis nigroviridis (Peters, 1859)/Black-speckled palm pit viper | 3 | BOTNI |
| Species | Uniprot Code | Protein | p Value (t Test) | FC (SQ/SW) |
|---|---|---|---|---|
| Dufourea novaeangliae | A0A154P4T7 | Four and a half LIM domains protein 2 | 0.010 | 28.572 |
| A0A154NYM3 | Transgelin | 0.001 | 16.538 | |
| A0A154PS72 | Profilin | 0.011 | 9.233 | |
| A0A154NWX1 | Laminin subunit β-1 | 0.020 | 6.452 | |
| A0A154PD98 | Nidogen-1 | 0.002 | 4.470 | |
| A0A154PLJ1 | Heat shock 70 kDa protein cognate 5 | 0.007 | 4.375 | |
| A0A154PJJ4 | Annexin | 0.002 | 3.627 | |
| A0A154PP88 | α-1,4 glucan phosphorylase | 0.008 | 3.360 | |
| A0A154PQH0 | Glycogenin-1 | 0.004 | 2.693 | |
| A0A154P296 | Acetyl-CoA hydrolase | 0.027 | 2.644 | |
| A0A154P796 | Spectrin α chain | 0.006 | 2.612 | |
| A0A154PAT6 | Malate dehydrogenase | 0.044 | 2.552 | |
| A0A154PNQ0 | Sortilin-related receptor | 0.017 | 2.535 | |
| A0A154P1L6 | ATP synthase subunit β | 0.019 | 2.389 | |
| A0A154P3S2 | Muscle M-line assembly protein unc-89 | 0.003 | 2.335 | |
| A0A154PJP5 | Titin | 0.019 | 2.170 | |
| A0A154PQ65 | Multifunctional fusion protein | 0.041 | 2.158 | |
| A0A154P3Q8 | Elongation factor 2 | 0.028 | 2.003 | |
| A0A154P289 | Vacuolar proton pump subunit B | 0.016 | 1.994 | |
| A0A154PPS0 | 2-phospho-D-glycerate hydro-lyase | 0.031 | 1.930 | |
| Apis cerana cerana | A0A2A3EBK6 | LIM domain-binding protein | 0.004 | 61.974 |
| A0A2A3EA22 | Four and a half LIM domains protein | 0.008 | 7.737 | |
| A0A2A3EL86 | Elongation factor 1-γ | 0.029 | 5.917 | |
| A0A2A3EC59 | Calpain-A | 0.000 | 3.745 | |
| A0A2A3E2W8 | Pyruvate dehydrogenase E1 component subunit α | 0.019 | 3.234 | |
| A0A2A3EGD2 | Arginine kinase | 0.022 | 2.724 | |
| A0A2A3EMK6 | α -actinin, sarcomeric | 0.001 | 2.719 | |
| A0A2A3EKS3 | Fructose-bisphosphate aldolase | 0.049 | 2.583 | |
| A0A2A3E5U7 | Talin-2 | 0.020 | 2.285 | |
| A0A2A3EPX9 | Glutamate dehydrogenase (NAD(p)(+)) | 0.026 | 2.042 | |
| A0A2A3E464 | ATP synthase subunit α | 0.025 | 2.028 | |
| A0A2A3EAL0 | Zn finger protein-like 1 homolog | 0.044 | 0.281 | |
| A0A2A3EJN3 | Acid β-galactosidase | 0.008 | 0.012 | |
| Frieseomelitta varia | A0A833VQ28 | 40S ribosomal protein S4 | 0.010 | 6.475 |
| A0A833VR32 | EF-1-γ-C-terminal domain-containing protein | 0.022 | 6.293 | |
| A0A833S6V3 | Neurochondrin homolog | 0.001 | 5.899 | |
| A0A833VVD0 | Isocitrate dehydrogenase [NAD] subunit, mitochondrial | 0.008 | 5.509 | |
| A0A833R5C8 | Lipocln_cytosolic_FA-bd_dom domain-containing protein | 0.001 | 4.562 | |
| A0A833VMV6 | Eukaryotic translation initiation factor 5A (eIF-5A) | 0.017 | 4.163 | |
| A0A833RMB6 | Muscle LIM protein Mlp84B | 0.008 | 4.161 | |
| A0A833RMX9 | Filamin-A | 0.011 | 4.147 | |
| A0A833W6L4 | Small nuclear ribonucleoprotein-associated protein | 0.041 | 4.017 | |
| A0A5P1MU32 | Glyceraldehyde-3-phosphate dehydrogenase | 0.000 | 4.000 | |
| A0A833WD27 | Pyruvate dehydrogenase E1 component subunit β | 0.049 | 3.428 | |
| A0A833S5I5 | Phosphoglycerate kinase | 0.001 | 3.175 | |
| A0A833R5H5 | DJ-1_PfpI domain-containing protein | 0.008 | 3.170 | |
| A0A833RCZ1 | Thioredoxin domain-containing protein | 0.009 | 3.081 | |
| A0A833S1K4 | Malic enzyme | 0.021 | 2.587 | |
| A0A833VV22 | WD_REPEATS_REGION domain-containing protein | 0.015 | 2.522 | |
| A0A833W6G6 | Pyruvate kinase | 0.002 | 2.501 | |
| A0A833S8P7 | ADF-H domain-containing protein | 0.009 | 2.389 | |
| A0A833RUN1 | Heat shock 70 kDa protein cognate 4 | 0.013 | 2.205 | |
| A0A833W2V9 | Fructose-bisphosphate aldolase | 0.021 | 2.173 | |
| A0A833WDK7 | Calreticulin | 0.024 | 2.003 | |
| A0A833WFT3 | Thioredoxin domain-containing protein | 0.028 | 1.975 | |
| A0A833RKT2 | α-glucosidase | 0.027 | 0.358 | |
| A0A833W1X0 | α-galactosidase | 0.021 | 0.298 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alonso-Sampedro, M.; Feás, X.; Bravo, S.B.; Chantada-Vázquez, M.P.; Vidal, C. Proteomics of Vespa velutina nigrithorax Venom Sac Queens and Workers: A Quantitative SWATH-MS Analysis. Toxins 2023, 15, 266. https://doi.org/10.3390/toxins15040266
Alonso-Sampedro M, Feás X, Bravo SB, Chantada-Vázquez MP, Vidal C. Proteomics of Vespa velutina nigrithorax Venom Sac Queens and Workers: A Quantitative SWATH-MS Analysis. Toxins. 2023; 15(4):266. https://doi.org/10.3390/toxins15040266
Chicago/Turabian StyleAlonso-Sampedro, Manuela, Xesús Feás, Susana Belén Bravo, María Pilar Chantada-Vázquez, and Carmen Vidal. 2023. "Proteomics of Vespa velutina nigrithorax Venom Sac Queens and Workers: A Quantitative SWATH-MS Analysis" Toxins 15, no. 4: 266. https://doi.org/10.3390/toxins15040266
APA StyleAlonso-Sampedro, M., Feás, X., Bravo, S. B., Chantada-Vázquez, M. P., & Vidal, C. (2023). Proteomics of Vespa velutina nigrithorax Venom Sac Queens and Workers: A Quantitative SWATH-MS Analysis. Toxins, 15(4), 266. https://doi.org/10.3390/toxins15040266