Functional Characterization of a New Degradation Peptide BmTX4-P1 from Traditional Chinese Scorpion Medicinal Material


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
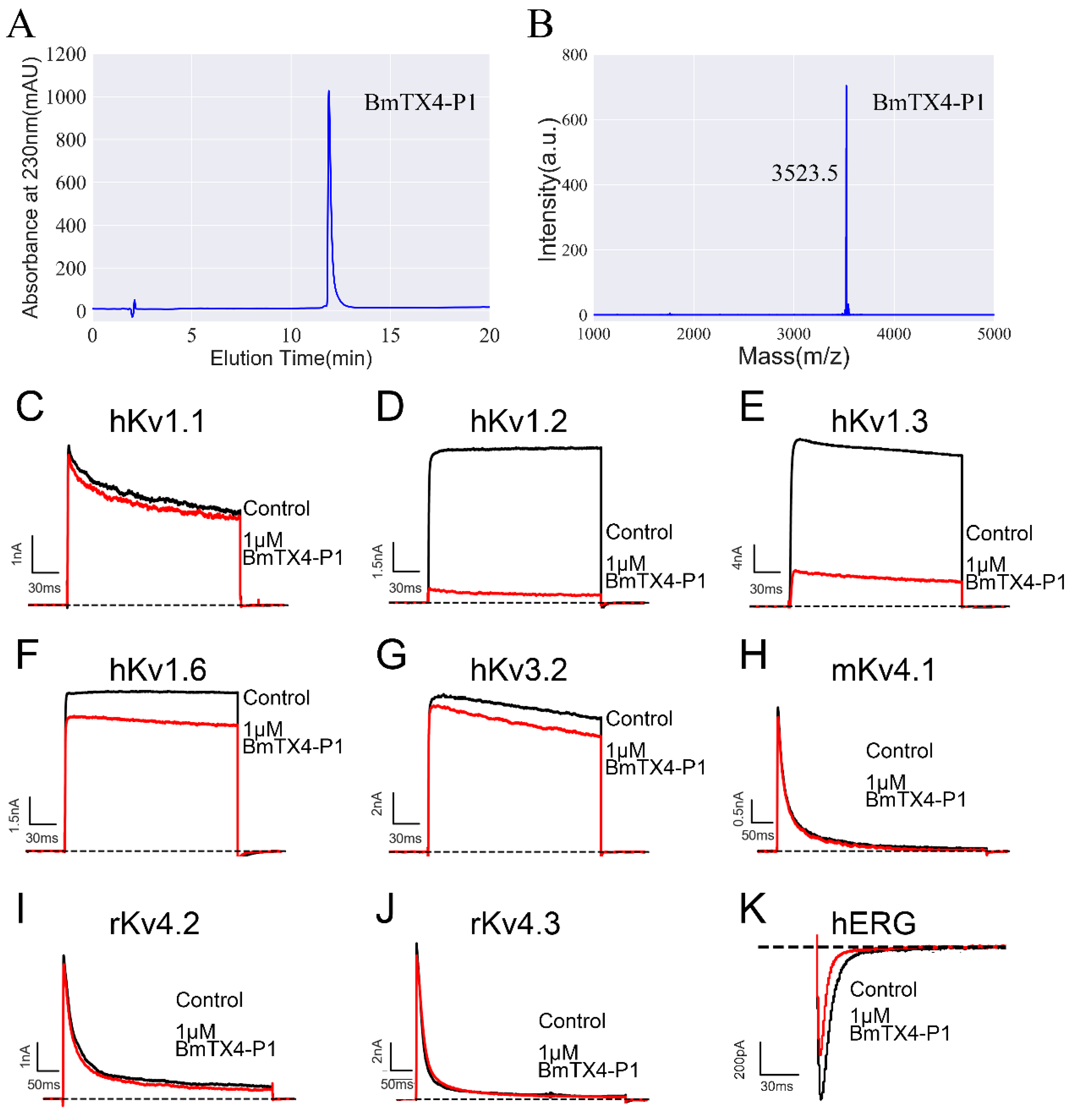
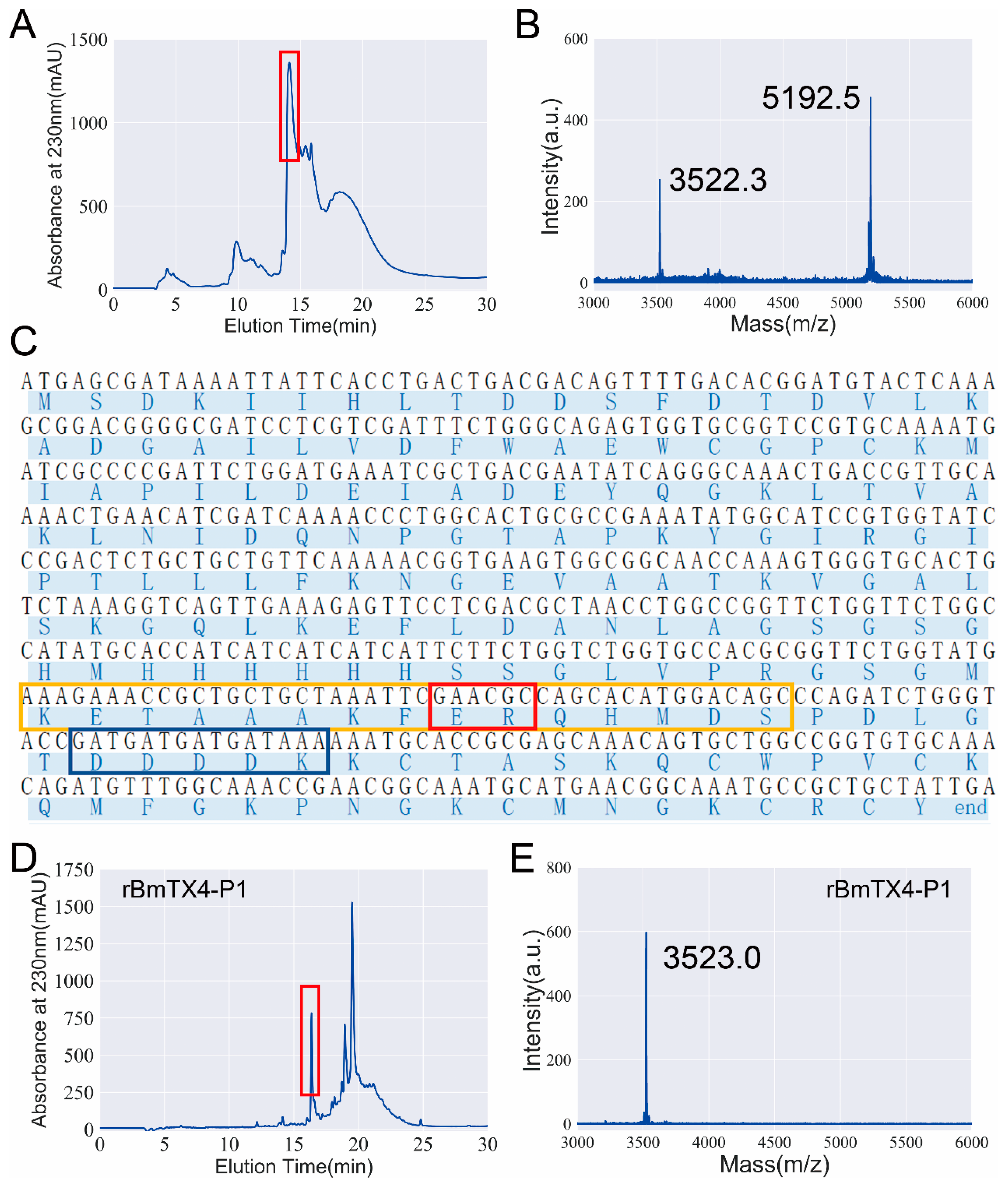
2.1. Identification of a New Degraded Peptide, BmTX4-P1, from Traditional Chinese Scorpion Medicinal Material
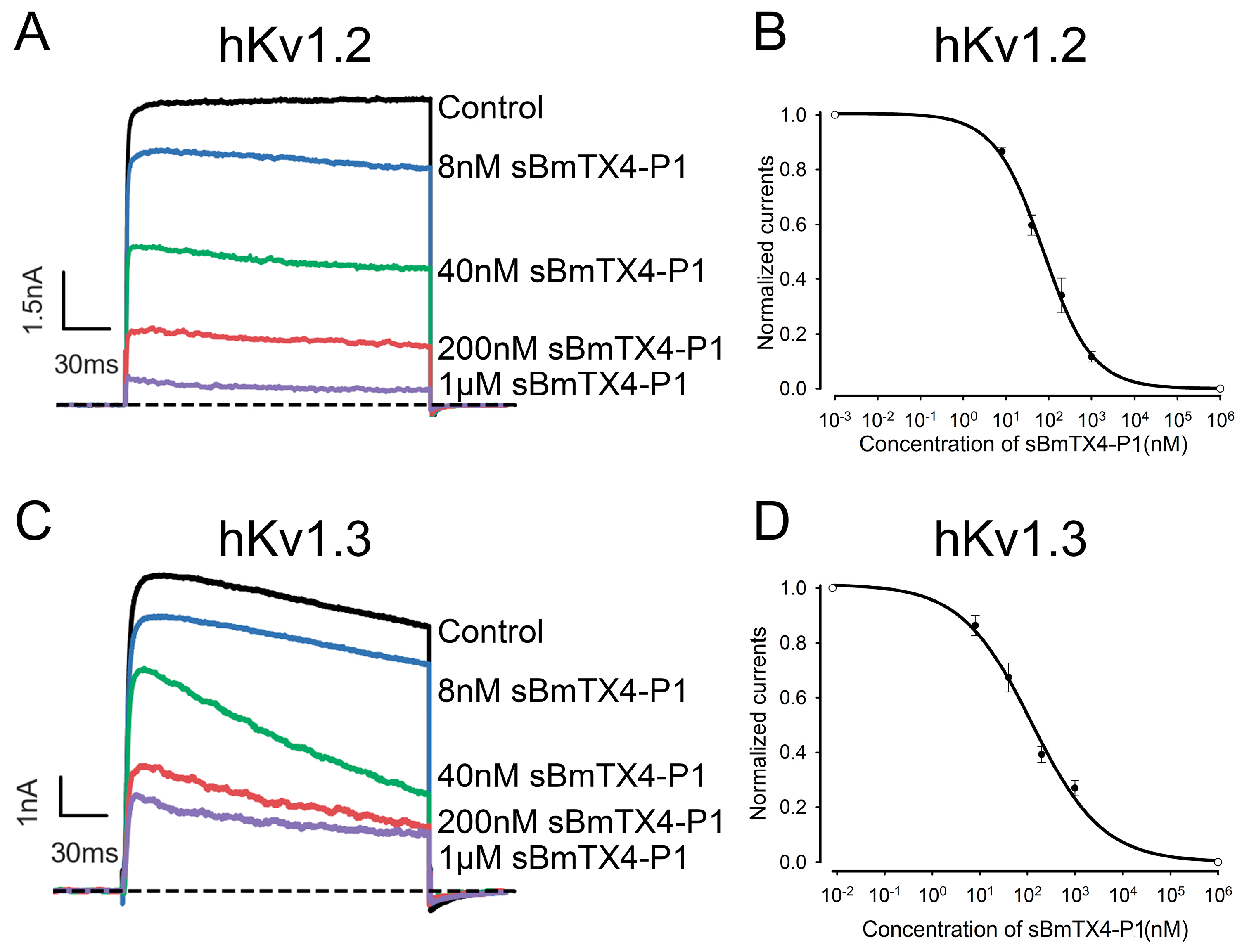
2.2. Functional Characterization of the Chemically Synthesized Peptide BmTX4-P1 (sBmTX4-P1)
2.3. Functional Expression and Pharmacological Property Confirmation of Recombinant BmTX4-P1 (rBmTX4-P1)
2.4. Identification of the Functional Sites of BmTX4-P1 Interacting with the Potassium Channel
3. Discussion
4. Materials and Methods
4.1. Scorpion Toxin Extraction and LC–MS/MS Analysis
4.2. Chemical Synthesis of sBmTX-P1
4.3. Source of Potassium Channel Plasmids
4.4. Cell Culture and Expression of Potassium Channels
4.5. Electrophysiological Recordings
4.6. Construction of the rBmTX4-P1 Peptide Expression Vector
4.7. Expression and Purification of Peptides
4.8. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chinese Pharmacopoeia Comission. Chinese Pharmacopoeia (Part I); China Medical Science and Technology Press: Beijing, China, 2020; p. 143. [Google Scholar]
- Yuan, Y.; Luo, L.; Peigneur, S.; Tytgat, J.; Zhu, S. Two recombinant depressant scorpion neurotoxins differentially affecting mammalian sodium channels. Toxicon 2010, 55, 1425–1433. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.C.; Zhang, L.; Nie, Y.; Luo, X. Identification and molecular characterization of three new K+-channel specific toxins from the Chinese scorpion Mesobuthus martensii Karsch revealing intronic number polymorphism and alternative splicing in duplicated genes. Peptides 2012, 34, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Umetsu, Y.; Gao, B.; Ohki, S.; Zhu, S. Mesomartoxin, a new K(v)1.2-selective scorpion toxin interacting with the channel selectivity filter. Biochem. Pharmacol. 2015, 93, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Kampo, S.; Cui, Y.; Yu, J.; Anabah, T.W.; Falagan, A.A.; Bayor, M.T.; Wen, Q.P. Scorpion Venom peptide, AGAP inhibits TRPV1 and potentiates the analgesic effect of lidocaine. Heliyon 2021, 7, e08560. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, Y.; Zhu, Y.; Zhang, L.; Ji, J.; Gui, M.; Li, C.; Song, Y. Study of Anti-Inflammatory and Analgesic Activity of Scorpion Toxins DKK-SP1/2 from Scorpion Buthus martensii Karsch (BmK). Toxins 2021, 13, 498. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xu, E.; Sang, M.; Wang, Z.; Zhang, Y.; Ye, J.; Zhou, Q.; Zhao, C.; Hu, C.; Lu, W.; et al. Makatoxin-3, a thermostable Nav1.7 agonist from Buthus martensii Karsch (BmK) scorpion elicits non-narcotic analgesia in inflammatory pain models. J. Ethnopharmacol. 2022, 288, 114998. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Hu, P.; Wu, W.; Wang, Y. Peptides with therapeutic potential in the venom of the scorpion Buthus martensii Karsch. Peptides 2019, 115, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Cheng, X.; Chen, J.; Wang, M.; Chen, Y.; Liu, J.; Sang, M.; Zhao, N.; Yan, H.; Cheng, X.; et al. A Buthus martensii Karsch scorpion sting targets Nav1.7 in mice and mimics a phenotype of human chronic pain. Pain 2022, 163, e202–e214. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Zou, X.; Li, S.; He, J.; Xi, C.; Tang, Q.; Wang, Y.; Cao, Z. BmK NSPK, a Potent Potassium Channel Inhibitor from Scorpion Buthus martensii Karsch, Promotes Neurite Outgrowth via NGF/TrkA Signaling Pathway. Toxins 2021, 13, 33. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Tao, J.; Zhang, S.; Lan, W.; Wang, C.; Ji, Y.; Cao, C. Selective Blockade of Neuronal BK (alpha + beta4) Channels Preventing Epileptic Seizure. J. Med. Chem. 2020, 63, 216–230. [Google Scholar] [CrossRef] [PubMed]
- Chandy, K.G.; Norton, R.S. Peptide blockers of Kv1.3 channels in T cells as therapeutics for autoimmune disease. Curr. Opin. Chem. Biol. 2017, 38, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Wang, D.; Tong, Y.; Qin, C.; Yang, L.; Yu, F.; Huang, X.; Liu, S.; Cao, Z.; Guo, L.; et al. Thermostable potassium channel-inhibiting neurotoxins in processed scorpion medicinal material revealed by proteomic analysis: Implications of its pharmaceutical basis in traditional Chinese medicine. J. Proteom. 2019, 206, 103435. [Google Scholar] [CrossRef]
- Corbett, M.A.; Bellows, S.T.; Li, M.; Carroll, R.; Micallef, S.; Carvill, G.L.; Myers, C.T.; Howell, K.B.; Maljevic, S.; Lerche, H.; et al. Dominant KCNA2 mutation causes episodic ataxia and pharmacoresponsive epilepsy. Neurology 2016, 87, 1975–1984. [Google Scholar] [CrossRef] [PubMed]
- Helbig, K.L.; Hedrich, U.B.; Shinde, D.N.; Krey, I.; Teichmann, A.C.; Hentschel, J.; Schubert, J.; Chamberlin, A.C.; Huether, R.; Lu, H.M.; et al. A recurrent mutation in KCNA2 as a novel cause of hereditary spastic paraplegia and ataxia. Ann. Neurol. 2016, 80. [Google Scholar] [CrossRef]
- Masnada, S.; Hedrich, U.B.S.; Gardella, E.; Schubert, J.; Kaiwar, C.; Klee, E.W.; Lanpher, B.C.; Gavrilova, R.H.; Synofzik, M.; Bast, T.; et al. Clinical spectrum and genotype-phenotype associations of KCNA2-related encephalopathies. Brain 2017, 140, 2337–2354. [Google Scholar] [CrossRef]
- Hedrich, U.B.S.; Lauxmann, S.; Wolff, M.; Synofzik, M.; Bast, T.; Binelli, A.; Serratosa, J.M.; Martinez-Ulloa, P.; Allen, N.M.; King, M.D.; et al. 4-Aminopyridine is a promising treatment option for patients with gain-of-function KCNA2-encephalopathy. Sci. Transl. Med. 2021, 13, eaaz4957. [Google Scholar] [CrossRef] [PubMed]
- Tarcha, E.J.; Chi, V.; Munoz-Elias, E.J.; Bailey, D.; Londono, L.M.; Upadhyay, S.K.; Norton, K.; Banks, A.; Tjong, I.; Nguyen, H.; et al. Durable pharmacological responses from the peptide ShK-186, a specific Kv1.3 channel inhibitor that suppresses T cell mediators of autoimmune disease. J. Pharmacol. Exp. Ther. 2012, 342, 642–653. [Google Scholar] [CrossRef] [PubMed]
- Chhabra, S.; Chang, S.C.; Nguyen, H.M.; Huq, R.; Tanner, M.R.; Londono, L.M.; Estrada, R.; Dhawan, V.; Chauhan, S.; Upadhyay, S.K.; et al. Kv1.3 channel-blocking immunomodulatory peptides from parasitic worms: Implications for autoimmune diseases. FASEB J. 2014, 28, 3952–3964. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liang, P.; Zhang, Y.; Wu, Y.; Song, Y.; Wang, X.; Chen, T.; Peng, B.; Liu, W.; Yin, J.; et al. Blockade of Kv1.3 Potassium Channel Inhibits Microglia-Mediated Neuroinflammation in Epilepsy. Int. J. Mol. Sci. 2022, 23, 14693. [Google Scholar] [CrossRef]
- Revuelta, M.; Urrutia, J.; Villarroel, A.; Casis, O. Microglia-Mediated Inflammation and Neural Stem Cell Differentiation in Alzheimer’s Disease: Possible Therapeutic Role of KV1.3 Channel Blockade. Front. Cell. Neurosci. 2022, 16, 868842. [Google Scholar] [CrossRef]
- Qin, C.; Wan, X.; Li, S.; Yang, F.; Yang, L.; Zuo, Z.; Cao, Z.; Chen, Z.; Wu, Y. Different pharmacological properties between scorpion toxin BmKcug2 and its degraded analogs highlight the diversity of K(+) channel blockers from thermally processed scorpions. Int. J. Biol. Macromol. 2021, 178, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, K.N.; Sivaraja, V.; Huys, I.; Sasaki, T.; Cheng, B.; Kumar, T.K.; Sato, K.; Tytgat, J.; Yu, C.; San, B.C.; et al. kappa-Hefutoxin1, a novel toxin from the scorpion Heterometrus fulvipes with unique structure and function. Importance of the functional diad in potassium channel selectivity. J. Biol. Chem. 2002, 277, 30040–30047. [Google Scholar] [CrossRef] [PubMed]
- Mouhat, S.; Mosbah, A.; Visan, V.; Wulff, H.; Delepierre, M.; Darbon, H.; Grissmer, S.; De Waard, M.; Sabatier, J.M. The ‘functional’ dyad of scorpion toxin Pi1 is not itself a prerequisite for toxin binding to the voltage-gated Kv1.2 potassium channels. Biochem. J. 2004, 377 Pt 1, 25–36. [Google Scholar] [CrossRef]
- Qin, C.; Yang, X.; Zuo, Z.; Yang, L.; Yang, F.; Cao, Z.; Chen, Z.; Wu, Y. BmK86-P1, a New Degradation Peptide with Desirable Thermostability and Kv1.2 Channel-Specific Activity from Traditional Chinese Scorpion Medicinal Material. Toxins 2021, 13, 610. [Google Scholar] [CrossRef] [PubMed]
- Miceli, F.; Guerrini, R.; Nappi, M.; Soldovieri, M.V.; Cellini, E.; Gurnett, C.A.; Parmeggiani, L.; Mei, D.; Taglialatela, M. Distinct epilepsy phenotypes and response to drugs in KCNA1 gain- and loss-of function variants. Epilepsia 2022, 63, e7–e14. [Google Scholar] [CrossRef] [PubMed]
- Smallwood, T.B.; Navarro, S.; Cristofori-Armstrong, B.; Watkins, T.S.; Tungatt, K.; Ryan, R.Y.M.; Haigh, O.L.; Lutzky, V.P.; Mulvenna, J.P.; Rosengren, K.J.; et al. Synthetic hookworm-derived peptides are potent modulators of primary human immune cell function that protect against experimental colitis in vivo. J. Biol. Chem. 2021, 297, 100834. [Google Scholar] [CrossRef]
- Mei, Y.; Fang, C.; Ding, S.; Liu, X.; Hu, J.; Xu, J.; Mei, Q. PAP-1 ameliorates DSS-induced colitis with involvement of NLRP3 inflammasome pathway. Int. Immunopharmacol. 2019, 75, 105776. [Google Scholar] [CrossRef]
- Kazama, I. Roles of lymphocyte Kv1.3-channels in gut mucosal immune system: Novel therapeutic implications for inflammatory bowel disease. Med. Hypotheses 2015, 85, 61–63. [Google Scholar] [CrossRef]
- Ousingsawat, J.; Spitzner, M.; Puntheeranurak, S.; Terracciano, L.; Tornillo, L.; Bubendorf, L.; Kunzelmann, K.; Schreiber, R. Expression of voltage-gated potassium channels in human and mouse colonic carcinoma. Clin. Cancer Res. 2007, 13, 824–831. [Google Scholar] [CrossRef]
- Vianna-Jorge, R.; Oliveira, C.F.; Garcia, M.L.; Kaczorowski, G.J.; Suarez-Kurtz, G. Shaker-type Kv1 channel blockers increase the peristaltic activity of guinea-pig ileum by stimulating acetylcholine and tachykinins release by the enteric nervous system. Br. J. Pharmacol. 2003, 138, 57–62. [Google Scholar] [CrossRef]
- Bassotti, G.; Usai-Satta, P.; Bellini, M. Linaclotide for the treatment of chronic constipation. Expert Opin. Pharmacother. 2018, 19, 1261–1266. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, K.; Sugiura, M.; Kimura, T. High Proteolytic Resistance of Spider-Derived Inhibitor Cystine Knots. Int. J. Pept. 2015, 2015, 537508. [Google Scholar] [CrossRef] [PubMed]
- Herzig, V.; King, G.F. The Cystine Knot Is Responsible for the Exceptional Stability of the Insecticidal Spider Toxin omega-Hexatoxin-Hv1a. Toxins 2015, 7, 4366–4380. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Han, S.; Cao, Z.; Wu, Y.; Zhuo, R.; Li, W. Fusion expression and purification of four disulfide-rich peptides reveals enterokinase secondary cleavage sites in animal toxins. Peptides 2013, 39, 145–151. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, C.; Yang, X.; Zhang, Y.; Deng, G.; Huang, X.; Zuo, Z.; Sun, F.; Cao, Z.; Chen, Z.; Wu, Y. Functional Characterization of a New Degradation Peptide BmTX4-P1 from Traditional Chinese Scorpion Medicinal Material. Toxins 2023, 15, 340. https://doi.org/10.3390/toxins15050340
Qin C, Yang X, Zhang Y, Deng G, Huang X, Zuo Z, Sun F, Cao Z, Chen Z, Wu Y. Functional Characterization of a New Degradation Peptide BmTX4-P1 from Traditional Chinese Scorpion Medicinal Material. Toxins. 2023; 15(5):340. https://doi.org/10.3390/toxins15050340
Chicago/Turabian StyleQin, Chenhu, Xuhua Yang, Yuanyuan Zhang, Gang Deng, Xin Huang, Zheng Zuo, Fang Sun, Zhijian Cao, Zongyun Chen, and Yingliang Wu. 2023. "Functional Characterization of a New Degradation Peptide BmTX4-P1 from Traditional Chinese Scorpion Medicinal Material" Toxins 15, no. 5: 340. https://doi.org/10.3390/toxins15050340
APA StyleQin, C., Yang, X., Zhang, Y., Deng, G., Huang, X., Zuo, Z., Sun, F., Cao, Z., Chen, Z., & Wu, Y. (2023). Functional Characterization of a New Degradation Peptide BmTX4-P1 from Traditional Chinese Scorpion Medicinal Material. Toxins, 15(5), 340. https://doi.org/10.3390/toxins15050340